Pinniped- and Cetacean-Derived ETosis Contributes to Combating Emerging Apicomplexan Parasites (Toxoplasma gondii, Neospora caninum) Circulating in Marine Environments

 , ,
, , 
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Marine Environment Affected by Emerging Neozoan Parasites
3. Cetacean/Pinniped Leukocytes of the Innate Immune System
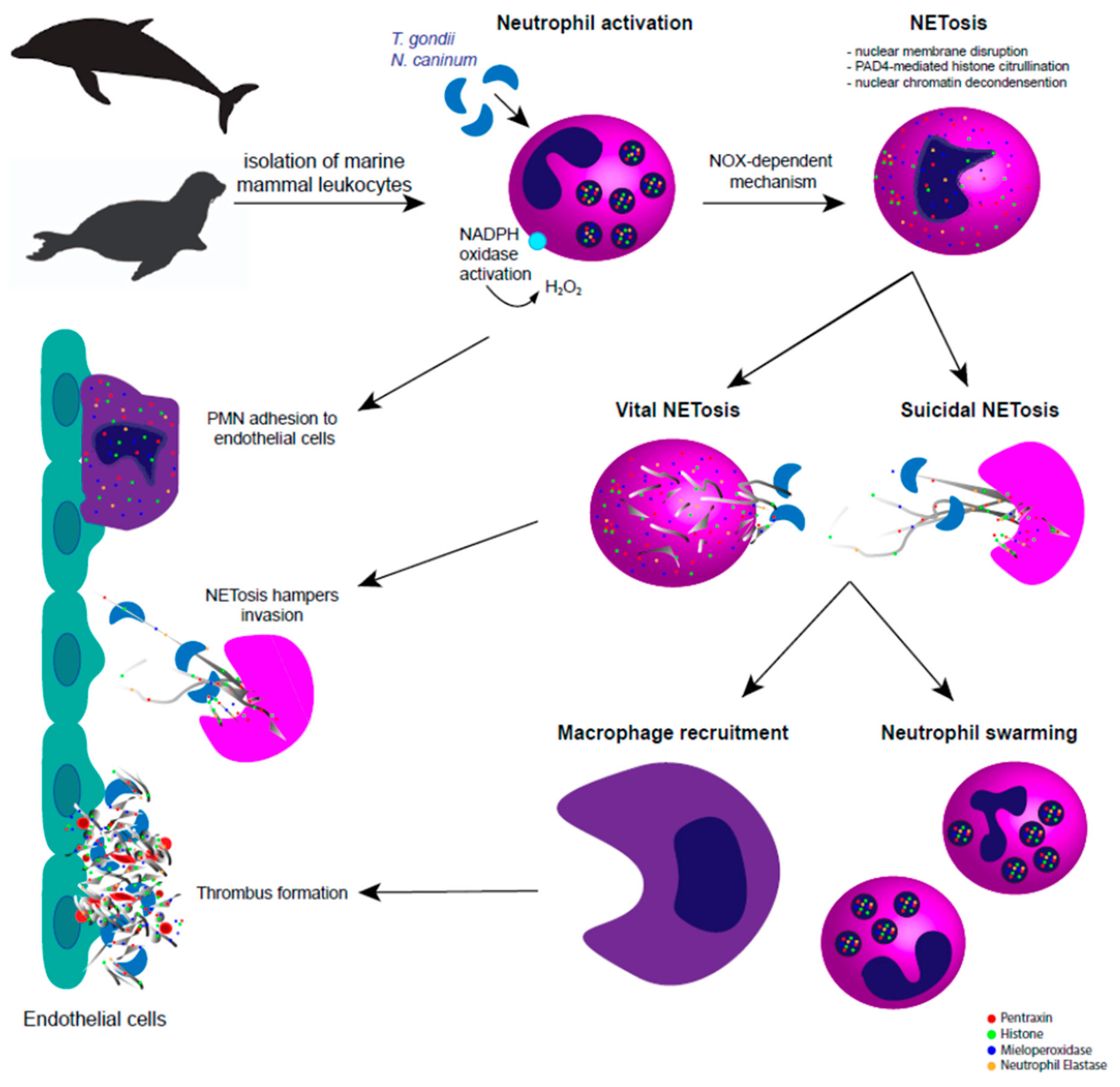
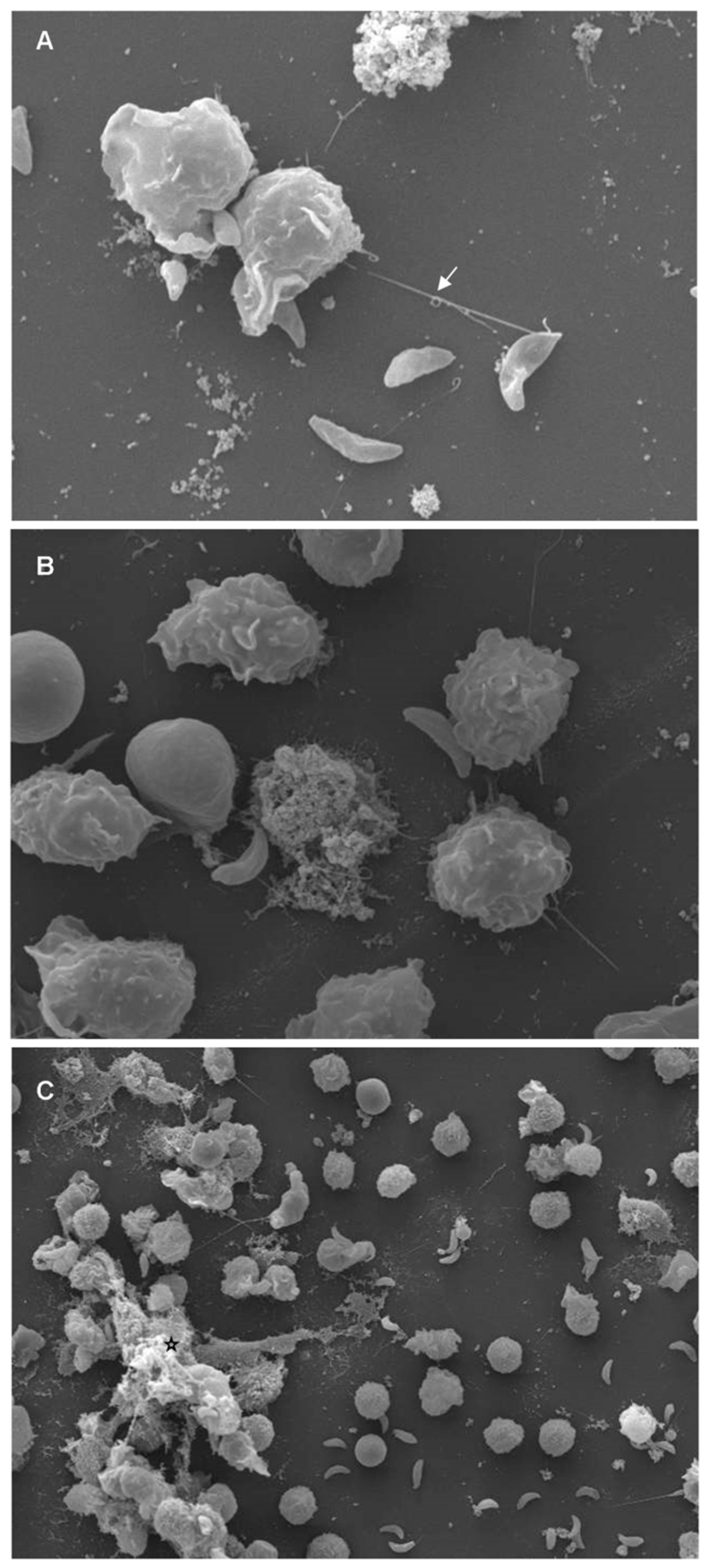
4. ETosis in Terrestrial and Marine Mammals
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abbas, A.K.; Litchman, A.H.; Pillai, S. Cellular and Molecular Immunology; Elsevier/Saunders: Philadelphia, PA, USA, 2012. [Google Scholar]
- Riera Romo, M.; Pérez-Martínez, D.; Castillo Ferrer, C. Innate immunity in vertebrates: An overview. Immunology 2016, 148, 125–139. [Google Scholar] [CrossRef] [PubMed]
- King, D.P.; Schrenzel, M.D.; McKnight, M.L.; Reidarson, T.H.; Hanni, K.D.; Stott, J.L.; Ferrick, D.A. Molecular cloning and sequencing of interleukin 6 cDNA fragments from the harbour seal (Phoca vitulina), killer whale (Orcinus orca), and southern sea otter (Enhydra lutris nereis). Immunogenetics 1996, 43, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, J.H.; Hermosilla, C.; Hardt, M.; Failing, K.; Zahner, H.; Taubert, A. PMN-mediated immune reactions against Eimeria bovis. Vet. Parasitol. 2008, 151, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Taubert, A.; Wimmers, K.; Ponksuksili, S.; Jimenez, C.A.; Zahner, H.; Hermosilla, C. Microarray-based transcriptional profiling of Eimeria bovis-infected bovine endothelial host cells. Vet. Res. 2010, 41, 70. [Google Scholar] [CrossRef] [PubMed]
- Abi Abdallah, D.S.; Denkers, E.Y. Neutrophils cast extracellular traps in response to protozoan parasites. Front. Immunol. 2012, 3, 382. [Google Scholar] [CrossRef] [PubMed]
- Pérez, D.; Muñoz, M.C.; Molina, J.M.; Muñoz-Caro, T.; Silva, L.M.R.; Taubert, A.; Hermosilla, C.; Ruiz, A. Eimeria ninakohlyakimovae induces NADPH oxidase-dependent monocyte extracellular trap formation and upregulates IL-12 and TGF-α, IL-6 and CCL2 gene transcription. Vet. Parasitol. 2016, 227, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef]
- Fonfara, S.; Siebert, U.; Prange, A.; Colijn, F. The impact of stress on cytokine and haptoglobin mRNA expression in blood samples from harbour porpoises (Phocoena phocoena). J. Mar. Biol. Assoc. UK 2007, 87, 305–311. [Google Scholar] [CrossRef]
- Kakuschke, A.; Valentine-Thon, E.; Griesel, S.; Fonfara, S.; Siebert, U.; Prange, A. Immunological impact of metals in harbor seals (Phoca vitulina) of the North Sea. Environ. Sci. Technol. 2005, 39, 7568–7575. [Google Scholar] [CrossRef]
- Beineke, A.; Siebert, U.; Wohlsein, P.; Baumgärtner, W. Immunology of whales and dolphins. Vet. Immunol. Immunopathol. 2010, 133, 81–94. [Google Scholar] [CrossRef]
- Charlton-Robb, K.; Gershwin, L.; Thompson, R.; Austin, J.; Owen, K.; McKechnie, S. A new dolphin species, the Burrunan dolphin Tursiops australis sp. nov., endemic to southern Australian coastal waters. PLoS ONE 2011, 6, e24047. [Google Scholar] [CrossRef] [PubMed]
- Hermosilla, C.; Silva, L.M.R.; Navarro, M.; Taubert, A. Anthropozoonotic endoparasites in free-ranging “urban” South American sea lions (Otaria flavescens). J. Vet. Med. 2016, 7507145. [Google Scholar] [CrossRef] [PubMed]
- Pompa, S.; Ehrlich, P.R.; Ceballos, G. Global distribution and conservation of marine mammals. Proc. Natl. Acad. Sci. USA 2011, 108, 13600–13605. [Google Scholar] [CrossRef] [PubMed]
- Cavagnolo, R.Z. The immunology of marine mammals. Dev. Comp. Immunol. 1979, 3, 245–257. [Google Scholar] [CrossRef]
- Noda, K.; Aoki, M.; Akiyoshi, H.; Asaki, H.; Shimada, T.; Ohashi, F. Evaluation of the polymorphonuclear cell functions of bottlenose dolphins. J. Vet. Med. Sci. 2003, 65, 727–729. [Google Scholar] [CrossRef] [PubMed]
- Noda, K.; Akiyoshi, H.; Aoki, M.; Shimada, T.; Ohashi, F. Relationship between transportation stress and polymorphonuclear cell functions of bottlenose dolphins, Tursiops truncatus. J. Vet. Med. Sci. 2007, 69, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.; Morsey, B.; Mori, C.; Nambiar, P.R.; De Guise, S. PCBs and TCDD, alone and in mixtures, modulate marine mammal but not B6C3F1 mouse leukocyte phagocytosis. Toxicol. Environ. Health A 2005, 68, 635–656. [Google Scholar] [CrossRef]
- Levin, M.; Morsey, B.; De Guise, S. Modulation of the respiratory burst by organochlorine mixtures in marine mammals, humans, and mice. J. Toxicol. Environ. Health A 2007, 70, 73–83. [Google Scholar] [CrossRef]
- Sokolova, O.V. Peculiarities of phagocytosis in bottlenose dolphin (Tursiops truncatus) during the period of adaptation to the captivity conditions. Dokl. Biol. Sci. 2005, 403, 263–266. [Google Scholar] [CrossRef]
- Pellisso, S.C.; Muñoz, M.J.; Carballo, M.; Sanchez-Vizcaino, J.M. Determination of the immunotoxic potential of heavy metals on the functional activity of bottlenose dolphin leukocytes in vitro. Vet. Immunol. Immunopathol. 2008, 121, 189–198. [Google Scholar] [CrossRef]
- Kato, H.; Perrin, W.F. Bryde´s whale: Balaenoptera edeni/brydei. InEncyclopaedia of Marine Mammals, 2nd ed.; Perrin, W.F., Würsig, B., Thewissen, J.G.M., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 158–163. ISBN 9780123735539. [Google Scholar]
- Frouin, H.; Lebeuf, M.; Hammill, M.; Fournier, M. Phagocytosis in pup and adult harbour, grey and harp seals. Vet. Immunol. Immunopathol. 2010, 134, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Schwacke, L.H.; Twiner, M.J.; De Guise, S.; Balmer, B.C.; Wells, R.S.; Townsend, F.I.; Rotstein, D.C.; Varela, R.A.; Hansen, L.J.; Zolman, E.S.; et al. Eosinophilia and biotoxin exposure in bottlenose dolphins (Tursiops truncatus) from a coastal area impacted by repeated mortality events. Environ. Res. 2010, 110, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Keogh, M.J.; Spoon, T.; Ridgway, S.H.; Jensen, E.; Van Bonn, W.; Romano, T.A. Simultaneous measurement of phagocytosis and respiratory burst of leukocytes in whole blood from bottlenose dolphins (Tursiops truncatus) utilizing flow cytometry. Vet. Immunol. Immunopathol. 2011, 144, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Zychlinsky, A. Neutrophil extracellular traps: Is immunity the second function of chromatin? J. Cell Biol. 2012, 198, 773–783. [Google Scholar] [CrossRef]
- Silva, L.M.R.; Muñoz-Caro, T.; Burgos, R.A.; Hidalgo, M.A.; Taubert, A.; Hermosilla, C. Far beyond phagocytosis: Phagocyte-derived extracellular traps act efficiently against protozoan parasites in vitro and in vivo. Mediat. Inflamm. 2016, 2016, 5898074. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Espinosa, O.; Rojas-Espinosa, O.; Moreno-Altamirano, M.M.; López-Villegas, E.O.; Sánchez-García, F.J. Metabolic requirements for neutrophil traps formation. Immunology 2015, 145, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S. Morbillivirus infections in aquatic mammals. J. Comp. Pathol. 1998, 119, 201–225. [Google Scholar] [CrossRef]
- Müller, G.; Wohlsein, P.; Beineke, A.; Haas, L.; Greiser-Wilke, I.; Siebert, U.; Fonfara, S.; Harder, T.; Stede, M.; Gruber, A.D.; et al. Phocine distemper in German seals, 2002. Emerg. Infect. Dis. 2004, 10, 723–725. [Google Scholar] [CrossRef]
- Conrad, P.A.; Miller, M.A.; Kreuder, C.; James, E.R.; Mazet, J.; Dabritz, H.; Jessup, D.A.; Gulland, F.; Grigg, M.E. Transmission of Toxoplasma: Clues from the study of sea otters as sentinels of Toxoplasma gondii flow into the marine environment. Int. J. Parasitol. 2005, 35, 1155–1168. [Google Scholar] [CrossRef]
- Miller, M.A.; Miller, W.A.; Conrad, P.A.; James, E.R.; Melli, A.C.; Leutenegger, C.M.; Dabritz, H.A.; Packham, A.E.; Paradies, D.; Harris, M.; et al. Type X Toxoplasma gondii in wild mussel and terrestrial carnivores from coastal California: New linkages between terrestrial mammals, runoff and toxoplasmosis of sea otters. Int. J. Parasitol. 2008, 38, 1319–1328. [Google Scholar] [CrossRef]
- Wohlsein, P.; Puff, C.; Kreutzer, M.; Siebert, U.; Baumgärtner, W. Distemper in a dolphin. Emerg. Infect. Dis. 2007, 13, 1959–1961. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Fair, P.A.; Sundar, N.; Velmurugan, G.; Kwok, O.C.; McFee, W.E.; Majumdar, D.; Su, C. Isolation of Toxoplasma gondii from bottlenose dolphins (Tursiops truncatus). J. Parasitol. 2008, 94, 821–823. [Google Scholar] [CrossRef]
- Dubey, J.P. History of the discovery of the life cycle of Toxoplasma gondii. Int. J. Parasitol. 2009, 39, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Van Bressem, M.F.; Raga, J.A.; Di Guardo, G.; Jepson, P.D.; Duignan, P.J.; Siebert, U.; Barrett, T.; Santos, M.C.; Moreno, I.B.; Siciliano, S.; et al. Emerging infectious diseases in cetaceans worldwide and the possible role of environmental stressors. Dis. Aquat. Organ. 2009, 86, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Kleinertz, S.; Hermosilla, C.; Ziltener, A.; Kreicker, S.; Hirzmann, J.; Abdel-Ghaffar, F.; Taubert, A. Gastrointestinal parasites of free-living Indo-Pacific bottlenose dolphins (Tursiops aduncus) in the Northern Red Sea, Egypt. Parasitol. Res. 2014, 113, 1405–1415. [Google Scholar] [CrossRef] [PubMed]
- Reichel, M.; Muñoz-Caro, T.; Sánchez-Contreras, G.; Rubio-García, A.; Magdowski, G.; Gärtner, U.; Hermosilla, C.; Taubert, A. Harbour seal (Phoca vitulina) PMN and monocytes release extracellular traps to capture the apicomplexan parasite Toxoplasma gondii. Dev. Comp. Immunol. 2015, 50, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Villagra-Blanco, R.; Silva, L.M.R.; Aguilella-Segura, A.; Arcenillas-Hernández, I.; Martínez-Carrasco, C.; Seipp, A.; Gärtner, U.; Ruiz de Ybañez, R.; Taubert, A.; Hermosilla, C. Bottlenose dolphins (Tursiopstruncatus) do also cast neutrophilextracellular traps against the apicomplexan parasite Neosporacaninum. Int. J. Parasitol Parasites Wildl. 2017, 6, 287–294. [Google Scholar] [CrossRef]
- Imlau, M.; Conejeros, I.; Muñoz-Caro, T.; Zhou, E.; Gärtner, U.; Ternes, K.; Taubert, A.; Hermosilla, C. Institute of Parasitology; Justus Liebig University Giessen: Giessen, Germany, 2019; unpublished data. [Google Scholar]
- Moore, M.J. How we all kill whales. ICES J. Mar. Sci. 2014, 71, 760–763. [Google Scholar] [CrossRef]
- Resendes, A.R.; Juan-Salles, C.; Almeria, S.; Majo, N.; Domingo, M.; Dubey, J.P. Hepatic sarcocystosis in a striped dolphin (Stenella coeruleoalba) from the Spanish Mediterranean coast. J. Parasitol. 2002, 88, 206–209. [Google Scholar] [CrossRef]
- Hughes-Hanks, J.M.; Rickard, L.G.; Panuska, C.; Saucier, J.R.; O’Hara, T.M.; Dehn, L.; Rolland, R.M. Prevalence of Cryptosporidium spp. and Giardia spp. in five marine mammal species. J. Parasitol. 2005, 91, 1225–1228. [Google Scholar] [CrossRef]
- Fujii, K.; Kakumoto, C.; Kobayashi, M.; Saito, S.; Kariya, T.; Watanabe, Y.; Xuan, X.; Igarashi, I.; Suzuki, M. Seroepidemiology of Toxoplasma gondii and Neospora caninum in seals around Hokkaido, Japan. J. Vet. Med. Sci. 2007, 69, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Raga, J.A.; Fernandez, M.; Balbuena, J.A.; Aznar, F.J. Parasites. In Encyclopaedia of Marine Mammals, 2nd ed.; Perrin, W.F., Thewissen, H.G.M., Würsing, B., Eds.; Academic Press Elsevier Inc.: San Diego, CA, USA, 2008; pp. 821–830. ISBN 9780123735539. [Google Scholar]
- Barbosa, L.; Johnson, C.K.; Lambourn, D.M.; Gibson, A.K.; Haman, K.H.; Huggins, J.L.; Sweeny, A.R.; Sundar, N.; Raverty, S.A.; Grigg, M.E. A novel Sarcocystis neurona genotype XIII is associated with severe encephalitis in an unexpectedly broad range of marine mammals from the northeastern Pacific Ocean. Int. J. Parasitol. 2015, 45, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Hermosilla, C.; Muñoz-Caro, T.; Silva, L.M.R.; Ruiz, A.; Taubert, A. The intriguing host innate immune response: Novel anti-parasitic defence by neutrophil extracellular traps. Parasitology 2014, 141, 1489–1498. [Google Scholar] [CrossRef] [PubMed]
- Hermosilla, C.; Silva, L.M.R.; Kleinertz, S.; Prieto, R.; Silva, M.A.; Taubert, A. Endoparasite survey of free-swimming baleen whales (Balaenoptera musculus, B. physalus, B. borealis) and sperm whales (Physeter macrocephalus) using non/minimally invasive methods. Parasitol. Res. 2016, 115, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Hermosilla, C.; Hirzmann, J.; Silva, L.M.R.; Brotons, J.M.; Cerdá, M.; Prenger-Berninghoff, E.; Ewers, C.; Taubert, A. Occurrence of anthropozoonotic parasitic infections and faecal microbes in free-ranging sperm whales (Physeter macrocephalus) from the Mediterranean Sea. Parasitol. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Inskeep, W., II; Gardiner, C.H.; Harris, R.K.; Dubey, J.P.; Goldston, R.T. Toxoplasmosis in Atlantic bottle-nosed dolphins (Tursiops truncatus). J. Wildl. Dis. 1990, 26, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Omata, Y.; Umeshita, Y.; Watarai, M.; Tachibana, M.; Sasaki, M.; Murata, K.; Yamada, T.K. Investigation for presence of Neospora caninum, Toxoplasma gondii and Brucella-species infection in killer whales (Orcinus orca) mass-stranded on the coast of Shiretoko, Hokkaido. Jpn. J. Vet. Med. Sci. 2006, 68, 523–526. [Google Scholar] [CrossRef]
- Mazzariol, S.; Marker, F.; Mignone, W.; Serracca, L.; Goria, M.; Marsili, L.; DiGuardo, G.; Casalone, C. Dolphin Morbillivirus and Toxoplasma gondii co-infection in a Mediterranean fin whale (Balaenoptera physalus). BMC Vet. Res. 2012, 8, 20. [Google Scholar] [CrossRef]
- Cabezón, O.; Hall, A.J.; Vincent, C.; Pabon, M.; García-Bocanegra, I.; Dubey, J.P.; Almería, S. Seroprevalence of Toxoplasma gondii in North-eastern Atlantic harbor seal (Phoca vitulina) and grey seal (Halichoerus grypus). Vet. Parasitol. 2011, 179, 253–256. [Google Scholar] [CrossRef]
- Massie, N.G.; Ware, M.W.; Villegas, E.; Black, M. Uptake and transmission of Toxoplasma gondii oocysts by migratory, filter-feeding fish. Vet. Parasitol. 2010, 169, 296–303. [Google Scholar] [CrossRef]
- Jardine, J.E.; Dubey, J.P. Congenital toxoplasmosis in a Indo-Pacific bottlenose dolphin (Tursiops aduncus). J. Parasitol. 2002, 88, 197–199. [Google Scholar] [CrossRef]
- Dubey, J.P.; Morales, J.A.; Sundar, N.; Velmurugan, G.V.; González-Barrientos, C.R.; Hernández-Mora, G.; Su, C. Isolation and genetic characterization of Toxoplasma gondii from striped dolphin (Stenella coeruleoalba) from Costa Rica. J. Parasitol. 2007, 93, 710–711. [Google Scholar] [CrossRef] [PubMed]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife–threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Gulland, F.M.; Hall, A.J. Is marine mammal health deterioring? Trends of global reporting of marine mammal disease. EcoHealth 2007, 4, 135–150. [Google Scholar] [CrossRef]
- Dubey, J.P. Review of Neospora caninum and neosporosis in animals. Korean J. Parasitol. 2003, 41, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Buergelt, C.D.; Bonde, R.K. Toxoplasmic meningoencephalitis in a West Indian manatee. J. Am. Vet. Med. Assoc. 1983, 183, 1294–1296. [Google Scholar]
- Olsen, M.E.; Roch, P.D.; Stabler, M.; Chan, W. Giardiasis in ringed seals from the western Arctic. J. Wildl. Dis. 1997, 33, 646–648. [Google Scholar] [CrossRef]
- Johnson, S.P.; Nolan, S.; Gulland, F.M.D. Antimicrobial susceptibility of bacteria isolated from pinnipeds stranded in central and northern California. J. Zoo Wildl. Med. 1998, 29, 288–294. [Google Scholar]
- LaPointe, J.M.; Gulland FM, D.; Haines, D.M.; Barr, B.C.; Duignan, P.J. Placentitis due to Coxiella burnetii in a Pacific harbor seal (Phoca vitulina richardsi). J. Vet. Diagn. Investig. 1999, 11, 541–543. [Google Scholar] [CrossRef]
- Van Pelt, R.W.; Dietrich, R.A. Staphylococcal infection and toxoplasmosis in a young harbor seal. J. Wildl. Dis. 1973, 9, 258–261. [Google Scholar] [CrossRef]
- Holshuh, H.J.; Sherrod, A.E.; Taylor, C.R.; Andrews, B.E.; Howard, E.B. Toxoplasmosis in a feral northern fur seal. J. Am. Vet. Med. Assoc. 1985, 187, 1229–1230. [Google Scholar] [PubMed]
- LaPointe, J.M.; Duignan, P.J.; Marsh, A.E.; Gulland, F.M.; Barr, B.C.; Naydan, D.K.; King, D.P.; Farman, C.A.; Huntingdon, K.A.B.; Lowenstine, L.J. Meningoencephalitis due to a Sarcocystis neurona-like protozoan in Pacific harbour seals (Phoca vitulina richardsi). J. Parasitol. 1999, 84, 1184–1189. [Google Scholar] [CrossRef]
- Bowater, R.O.; Norton, J.; Johnson, S.; Hill, B.; O’Donoghue, P.; Prior, H. Toxoplasmosis in Indo-Pacific humpbacked dolphins (Sousa chinensis), from Queensland. Aust. Vet.J. 2003, 81, 627–632. [Google Scholar] [CrossRef]
- Cabezón, O.; Resendes, A.R.; Domingo, M.; Raga, J.A.; Agustí, C.; Alegre, F.; Mons, J.L.; Dubey, J.P.; Almería, S. Seroprevalence of Toxoplasma gondii antibodies in wild dolphins from the Spanish Mediterranean coast. J. Parasitol. 2004, 90, 643–644. [Google Scholar] [CrossRef] [PubMed]
- Santos, P.S.; Albuquerque, G.R.; da Silva, V.M.; Martin, A.R.; Marvulo, M.F.; Souza, S.L.; Ragozo, A.M.; Nascimento, C.C.; Gennari, S.M.; Dubey, J.P.; et al. Seroprevalence of Toxoplasma gondii in free-living Amazon River dolphins (Inia geoffrensis) from central Amazon, Brazil. Vet. Parasitol. 2011, 183, 171–173. [Google Scholar] [CrossRef]
- Mikaelian, I.; Boisclair, J.; Dubey, J.P.; Kennedy, S.; Martineau, D. Toxoplasmosis in beluga whales from the St. Lawrence estuary: Two case reports and a serological survey. J. Comp. Pathol. 2000, 122, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Cole, R.A.; Lindsay, D.S.; Howe, D.K.; Roderick, C.L.; Dubey, J.P.; Thomas, N.J.; Baeten, L.A. Biological and molecular characterizations of Toxoplasma gondii strains obtained from southern sea otters (Enhydra lutris nereis). J. Parasitol. 2000, 86, 526–530. [Google Scholar] [CrossRef]
- Goldstein, J.D.; Reese, E.; Reif, J.S.; Varela, R.A.; McCulloch, S.D.; Dfran, R.H.; Fair, P.A.; Bossart, G.D. Hematologic, biochemical, and cytologic findings from apparently healthy Atlantic bottlenose dolphins (Tursiops truncatus) inhabiting the India River Lagoon, Florida, USA. J. Wildl. Dis. 2006, 42, 447–454. [Google Scholar] [CrossRef]
- Hall, A.J.; Wells, R.S.; Sweeney, J.C.; Townsend, F.I.; Balmer, B.C.; Hohn, A.A.; Rhinehart, H.L. Annual, seasonal and individual variation in hematology and clinical blood chemistry profiles in bottlenose dolphins (Tursiops truncatus) from Sarasota Bay, Florida. Comp. Biochem. Physiol. A 2007, 148, 266–277. [Google Scholar] [CrossRef]
- Venn-Watson, S.; Jensen, E.D.; Ridgway, S.H. Effects of age and sex on clinic-pathologic reference ranges in a healthy managed Atlantic bottlenose dolphin population. J. Am. Vet. Med. Assoc. 2007, 231, 596–601. [Google Scholar] [CrossRef]
- Schwacke, L.H.; Hall, A.J.; Townsend, F.I.; Wells, R.S.; Hansen, L.J.; Hohn, A.A.; Bossart, G.D.; Fair, P.A.; Rowles, T.K. Hematologic and serum biochemical reference intervals for free-ranging common bottlenose dolphins (Tursiops truncatus) and variation in the distributions of clinicopathologic values related to geographic sampling site. Am. J. Vet. Res. 2009, 70, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Asper, E.D.; Cornell, L.; Duffield, D.A.; Odell, D.K.; Joseph, B.E.; Stark, B.I.; Perry, C.A. Hematology and serum chemistry values in bottlenose dolphins. In The Bottlenose Dolphin; Leatherwood, S., Reeves, R.R., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 479–485. ISBN 9780323139618. [Google Scholar]
- Woolford, L.; Wong, A.; Sneath, H.L.; Long, T.; Boyd, S.P.; Lanyon, J.M. Hematology of dugongs (Dugong dugon) in southern Queensland. Vet. Clin. Pathol. 2015, 44, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Bossart, G.D. Immunocytes of the Atlantic Bottlenose Dolphin (Tursiops truncatus) and West Indian Manatee (Trichechus manatus latirostris): Morphologic Characterizations and Correlations Between Healthy and Disease States and under Free-Ranging and Captive Conditions. Ph.D. Thesis, Florida International University, Miami, FL, USA, 1995. [Google Scholar]
- Reidarson, T.H.; Duffield, D.; McBain, J. Normal hematology of marine mammals. In Schalm’s Veterinary Hematology, 5th ed.; Feldman, B.F., Zinkl, J.G., Jain, N.C., Eds.; Lippincott Williams& Wilkins: Philadelphia, PA, USA, 2000; pp. 1164–1173. ISBN 978-0683306927. [Google Scholar]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, A.S.; Foster, D.N.; Troedsson, M.H. Equine seminal plasma reduces sperm binding to polymorphonuclear neutrophils (PMNs) and improves the fertility of fresh semen inseminated into inflamed uteri. Reproduction 2004, 127, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.M.R.; Muñoz-Caro, T.; Rüdiger, G.; Vila-Viçosa, M.J.M.; Cortes, H.C.E.; Hermosilla, C.; Taubert, A. The apicomplexan parasite Eimeria arloingi induces caprine neutrophil extracellular traps. Parasitol. Res. 2014, 113, 2797–2807. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Caro, T.; Conejeros, I.; Zhou, E.; Pikhovych, A.; Gärtner, U.; Hermosilla, C.; Kulke, D.; Taubert, A. Dirofiliaria immitis microfilariae and third-stage larvae induce canine NETosis resulting in different types of neutrophil extracellular traps. Front. Immunol. 2018, 9, 968. [Google Scholar] [CrossRef] [PubMed]
- Kiehl, A.R.; Schiller, C.A. A study of manatee leukocytes using peroxidase stain. Vet. Clin. Pathol. 1994, 23, 50–53. [Google Scholar] [CrossRef]
- Muñoz-Caro, T.; Silva, L.M.R.; Rentería-Solís, Z.; Taubert, A.; Hermosilla, C. Neutrophil extracellular traps in the intestinal mucosa of Eimeria-infected animals. Asian Pac. J. Trop. Biomed. 2016, 6, 301–307. [Google Scholar] [CrossRef]
- Meyer, D.J.; Harvey, J.W. Veterinary Laboratory Medicine: Interpretation and Diagnosis; W.B. Saunders: Philadelphia, PA, USA, 1998; p. 350. [Google Scholar]
- Roth, J.A.; Kaeberle, M.L. Isolation of neutrophils and eosinophils from the peripheral blood of cattle and comparison of functional activities. J. Immunol. Methods 1981, 45, 153–156. [Google Scholar] [CrossRef]
- Doyle, A.D.; Jacobsen, E.A.; Ochkur, S.I.; McGarry, M.P.; Shim, K.G.; Nguyen, D.T.; Protheroe, C.; Colbert, D.; Kloeber, J.; Neely, J.; et al. Expression of the secondary granule proteins major basic protein 1 (MBP-1) and eosinophil peroxidase (EPX) is required for eosinophilopoiesis in mice. Blood 2013, 122, 781–790. [Google Scholar] [CrossRef]
- Desforges, J.P.W.; Sonne, C.; Levin, M.; Siebert, U.; de Guise, S.; Dietz, R. Immunotoxic effects of environmental pollutants in marine mammals. Environ. Int. 2016, 86, 126–139. [Google Scholar] [CrossRef] [PubMed]
- De Swart, R.L.; Kluten, R.M.G.; Huizing, C.J.; Vedder, L.J.; Reijnders, P.J.H.; Visser, I.K.G.; UytdeHaag, F.G.C.M.; Osterhaus, A.D.M.E. Mitogen and antigen induced B and T cell responses of peripheral blood mononuclear cells from the harbour seal (Phoca vitulina). Vet. Immunol. Immunopathol. 1993, 37, 217–230. [Google Scholar] [CrossRef]
- De Swart, R.; Ross, P.S.; Vedder, L.J.; Boink, F.B.T.J.; Reijnders, P.J.H.; Mulduer, P.G.H.; Osterhaus, A.D.M.E. Haematology and clinical chemistry values for harbour seals (Phoca vitulina) fed environmentally contaminated herring remain within normal ranges. Can. J. Zool. 1995, 73, 2035–2043. [Google Scholar] [CrossRef]
- Lehnert, K.; Schwanke, E.; Hahnb, K.; Wohlsein, P.; Siebert, U. Heartworm (Acanthocheilonema spirocauda) and seal louse (Echinophthirius horridus) infections in harbour seals (Phoca vitulina) from the North and Baltic Seas. J. Sea Res. 2015, 113, 65–72. [Google Scholar] [CrossRef]
- Hasselmeier, I.; Fonfara, S.; Driver, J.; Siebert, U. Differential hematology profiles of free-ranging, rehabilitated, and captive harbor seals (Phoca vitulina) of the German North Sea. Aquat. Mammals 2008, 34, 149–156. [Google Scholar] [CrossRef]
- Gupta, A.K.; Hasler, P.; Holzgreve, W.; Gebhardt, S.; Hahn, S. Induction of neutrophil extracellular DNA lattices by placental microparticles and IL-8 and their presence in preeclampsia. Hum. Immunol. 2005, 66, 1146–1154. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.; Giaglis, S.; Hoesli, I.; Hasler, P. Neutrophil NETs in reproduction: From infertility to preeclampsia and the possibility of fetal loss. Front. Immunol. 2012, 3, 362. [Google Scholar] [CrossRef]
- Hahn, S.; Giaglis, S.; Chowdhury, C.S.; Hösli, I.; Hasler, P. Modulation of neutrophil NETosis: Interplay between infectious agents and underlying host physiology. Semin. Immunopathol. 2013, 35, 439–453. [Google Scholar] [CrossRef]
- Cheng, O.Z.; Palaniyar, N. NET Balancing: A problem in inflammatory lung diseases. Front. Immunol. 2013, 4, 1. [Google Scholar] [CrossRef]
- Zambrano, F.; Carrau, T.; Gärtner, U.; Seipp, A.; Taubert, A.; Felmer, R.; Sánchez, R.; Hermosilla, C. Leukocytes co-incubated with human sperm trigger classic neutrophil extracellular traps formation, reducing sperm motility. Fertil. Steril. 2016, 106, 1053–1060. [Google Scholar] [CrossRef]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2017, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Von Köckritz-Blickwede, M.; Goldmann, O.; Thulin, P.; Heinemann, K.; Norrby-Teglund, A.; Rohde, M.; Medina, E. Phagocytosis-independent antimicrobial activity of mast cells by means of extracellular trap formation. Blood 2008, 111, 3070–3080. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.M.; Rubin, C.J.; Khandpur, R.; Wang, J.Y.; Riblett, M.; Yalavarthi, S.; Villanueva, E.C.; Shah, P.; Kaplan, M.J.; Bruce, A.T. Mast cells and neutrophils release IL-17 through extracellular trap formation in psoriasis. J. Immunol. 2011, 187, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Möllerherm, H.; von Köckritz-Blickwede, M.; Branitzki-Heinemann, K. Antimicrobial activityof mast cells: Role and relevance of extracellular DNA traps. Front. Immunol. 2016, 7, 265. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, S.; Gold, J.A.; Andina, N.; Lee, J.J.; Kelly, A.M.; Kozlowski, E.; Schmid, I.; Straumann, A.; Reichenbach, J.; Gleich, G.J.; et al. Catapult-like release of mitochondrial DNA by eosinophils contributes to antibacterial defense. Nat. Med. 2008, 14, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Aulik, N.A.; Hellenbrand, K.M.; Czuprynski, C.J. Mannheimia haemolytica and its leukotoxin cause macrophage extracellular trap formation by bovine macrophages. Infect. Immun. 2010, 80, 1923–1933. [Google Scholar] [CrossRef]
- Hellenbrand, K.M.; Forsythe, K.M.; Rivera-Rivas, J.J.; Czuprynski, C.J.; Aulik, N.A. Histophilus somni causes extracellular trap formation by bovine neutrophils and macrophages. Microb. Pathog. 2013, 54, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Boe, D.M.; Curtis, B.J.; Chen, M.M.; Ippolito, J.A.; Kovacs, E.J. Extracellular traps and macrophages: New roles for the versatile phagocyte. J. Leukoc. Biol. 2015, 97, 1023–1035. [Google Scholar] [CrossRef]
- Doster, R.S.; Rogers, L.M.; Gaddy, J.A.; Aronoff, D.M. Macrophage extracellular traps: A scoping review. J. Innate Immun. 2018, 10, 3–13. [Google Scholar] [CrossRef]
- Schorn, C.; Janko, C.; Latzko, M.; Chaurio, R.; Schett, G.; Herrmann, M. Monosodium urate crystals induce extracellular DNA traps in neutrophils, eosinophils, and basophils but not in mononuclear cells. Front. Immunol. 2012, 3, 277. [Google Scholar] [CrossRef]
- Morshed, M.; Hlushchuk, R.; Simon, D.; Walls, A.F.; Obata-Ninomiya, K.; Karasuyama, H.; Djonov, V.; Eggel, A.; Kaufmann, T.; Simon, H.U.; et al. NADPH oxidase-independent formation of extracellular DNA traps by basophils. J. Immunol. 2014, 192, 5314–5323. [Google Scholar] [CrossRef] [PubMed]
- Taubert, A.; Behrendt, J.H.; Sühwold, A.; Zahner, H.; Hermosilla, C. Monocyte- and macrophage-mediated immune reactions against Eimeria bovis. Vet. Parasitol. 2009, 164, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Caro, T.; Hermosilla, C.; Silva, L.M.R.; Cortes, H.; Taubert, A. Neutrophil extracellular traps as innate immune reaction against the emerging apicomplexan parasite Besnoitia besnoiti. PLoS ONE 2014, 9, e91415. [Google Scholar]
- Halder, L.D.; Abdelfatah, M.A.; Jo, E.A.; Jacobsen, I.D.; Westermann, M.; Beyersdorf, N.; Lorkowski, S.; Zipfel, P.F.; Skerka, C. Factor H binds to extracellular DNA traps released from human blood monocytes in response to Candida albicans. Front. Immunol. 2016, 7, 671. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, S.; Urosevic, M.; Daryadel, A.; Oberholzer, P.A.; Baumann, C.; Fey, M.F.; Dummer, R.; Simon, H.U.; Yousefi, S. Induction of genes mediating interferon dependent extracellular trap formation during neutrophil differentiation. J. Biol. Chem. 2004, 279, 44123–44132. [Google Scholar] [CrossRef] [PubMed]
- Guimarães-Costa, A.B.; Rochael, N.C.; Oliveira, F.; Echevarria-Lima, J.; Saraiva, E.M. Neutrophil extracellular traps reprogram IL-4/GM-CSF-induced monocyte differentiation to anti-inflammatory macrophages. Front. Immunol. 2017, 8, 523. [Google Scholar] [CrossRef] [PubMed]
- Caudrillier, A.; Kessenbrock, K.; Gilliss, B.M.; Nguyen, J.X.; Marques, M.B.; Monestier, M.; Toy, P.; Werb, Z.; Looney, M.R. Platelets induce neutrophil extracellular traps in transfusion-related acute lung injury. J. Clin. Investig. 2012, 122, 2661–2671. [Google Scholar] [CrossRef]
- Clark, S.R.; Ma, A.C.; Tavener, S.A.; McDonald, B.; Goodarzi, Z.; Kelly, M.M.; Patel, K.D.; Chakrabarti, S.; McAvoy, E.; Sinclair, G.D.; et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat. Med. 2007, 13, 463–469. [Google Scholar] [CrossRef]
- Pijanowski, L.; Golbach, L.; Kolaczkowska, E.; Scheer, M.; Verburg-van Kemenade, B.M.L.; Chadzinska, M. Carp neutrophilic granulocytes form extracellular traps via ROS-dependent and independent pathways. Fish Shellfish Immunol. 2013, 34, 1244–1252. [Google Scholar] [CrossRef]
- Desai, J.; Kumar, S.V.; Mulay, S.R.; Konrad, L.; Romoli, S.; Schauer, C.; Herrmann, M.; Bilyy, R.; Müller, S.; Popper, B.; et al. PMA and crystal-induced neutrophil extracellular trap formation involves RIPK1-RIPK3-MLKL signaling. Eur. J. Immunol. 2016, 46, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Kenny, E.F.; Herzig, A.; Krüger, R.; Muth, A.; Mondal, S.; Thompson, P.R.; Brinkmann, V.; Bernuth, H.V.; Zychlinsky, A. Diverse stimuli engage different neutrophil extracellular trap pathways. eLife 2017, 6, e24437. [Google Scholar] [CrossRef] [PubMed]
- Nishinaka, Y.; Arai, T.; Adachi, S.; Takaori-Kondo, A.; Yamashita, K. Singlet oxygen is essential for neutrophil extracellular trap formation. Biochem. Biophys. Res.Commun. 2011, 413, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Urban, C.F.; Reichard, U.; Brinkmann, V.; Zychlinsky, A. Neutrophil extracellular traps capture and kill Candida albicans yeast and hyphal forms. Cell Microbiol. 2006, 8, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Alemán, O.R.; Mora, N.; Cortes-Vieyra, R.; Uribe-Querol, E.; Rosales, C. Differential use of human neutrophil Fcγ receptors for inducing neutrophil extracellular trap formation. J. Immunol. Res. 2016, 2016, 2908034. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Shi, X.; Tang, X.; Wang, Y.; Shen, F.; Zhang, Q.; Wang, C.; Jiang, M.; Liu, M.; Yu, L. Aflatoxin B1 induces reactive oxygen species-mediated autophagy and extracellular trap formation in macrophages. Front. Cell. Infect. Microbiol. 2017, 7, 53. [Google Scholar] [CrossRef]
- Pilsczek, F.H.; Salina, D.; Poon, K.K.H.; Fahey, C.; Yipp, B.G.; Sibley, C.D.; Robbins, S.M.; Green, F.H.Y.; Surette, M.G.; Sugai, M. A novel mechanism of rapid nuclear neutrophil extracellular trap formation in response to Staphylococcus aureus. J. Immunol. 2010, 185, 7413–7426. [Google Scholar] [CrossRef]
- Johnson, C.J.; Davis, J.M.; Huttenlocher, A.; Kernien, J.F.; Nett, J.E. Emerging fungal pathogen Candida auris evades neutrophil attack. mBio 2018, 9, e01403-18. [Google Scholar] [CrossRef]
- Jenne, C.N.; Kubes, P. NETs tangle with HIV. Cell Host Microbe 2012, 12, 5–7. [Google Scholar] [CrossRef]
- Jenne, C.N.; Kubes, P. Virus-induced NETs-critical component of host defense or pathogenic mediator? PLoS Pathog. 2015, 11, e1004546. [Google Scholar] [CrossRef] [PubMed]
- Guimarães-Costa, A.B.; Nascimento, M.T.; Froment, G.S.; Soares, R.P.; Morgado, F.N.; Conceição-Silva, F.; Saraiva, E.M. Leishmania amazonensis promastigotes induce and are killed by neutrophil extracellular traps. Proc. Natl. Acad. Sci. USA 2009, 106, 6748–6753. [Google Scholar] [CrossRef] [PubMed]
- Abi Abdallah, D.S.; Lin, C.; Ball, C.J.; King, M.R.; Duhamel, G.E.; Denkers, E.Y. Toxoplasma gondii triggers release of human and mouse neutrophil extracellular traps. Infect. Immun. 2012, 80, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Hermosilla, C.; Ruiz, A.; Taubert, A. Eimeria bovis: An update on parasite-host interactions. Int. J. Med. Microbiol. 2012, 302, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Y.; Xin, L.; Beverley, S.M.; Carlsen, E.D.; Popov, V.; Chang, K.P.; Wang, M.; Soong, L. Differential microbicidal effects of human histone proteins H2A and H2B on Leishmania promastigotes and amastigotes. Infect. Immun. 2011, 79, 1124–1133. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Leshner, M.; Wang, S.; Lewis, C.; Zheng, H.; Chen, X.A.; Santy, L.; Wang, Y. PAD4 mediated histone hypercitrullination induces heterochromatin decondensation and chromatin unfolding to form neutrophil extracellular trap like structures. Front. Immunol. 2012, 3, 307. [Google Scholar] [CrossRef]
- Muñoz-Caro, T.; Mena Huertas, J.S.; Conejeros, I.; Alarcón, P.; Hidalgo, M.A.; Burgos, R.A.; Taubert, A.; Hermosilla, C. CD11b-, ERK/MAP kinase- and SOCE-dependent Eimeria bovis-triggered neutrophil extracellular trap formation. Vet. Res. 2015, 46, 23. [Google Scholar] [CrossRef]
- Muñoz-Caro, T.; Lendner, M.; Daugschies, A.; Hermosilla, C.; Taubert, A. NADPH oxidase, MPO, NE, ERK1/2, p38 MAPK and Ca2+ influx are essential for Cryptosporidium parvum-induced NET formation. Dev. Comp. Immunol. 2015, 52, 245–254. [Google Scholar] [CrossRef]
- Li, P.; Li, M.; Lindberg, M.R.; Kennett, M.J.; Xiong, N.; Wang, Y. PAD4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. J. Exp. Med. 2010, 207, 1853–1862. [Google Scholar] [CrossRef]
- Yipp, B.G.; Kubes, P. NETosis: How vital is it? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef] [PubMed]
- De Guise, S.; Flipo, D.; Boehm, J.; Martineau, D.; Béland, P.; Fournier, M. Immune functions in beluga whales (Delphinapterus leucas): Evaluation of phagocytosis and respiratory burst with peripheral blood using flow cytometry. Vet. Immunol. Immunopathol. 1995, 47, 351–362. [Google Scholar] [CrossRef]
- Hakkim, A.; Fuchs, T.A.; Martinez, N.E.; Hess, S.; Prinz, H.; Zychlinsky, A.; Waldmann, H. Activation of the Raf-MEK-ERK pathway is required for neutrophil extracellular trap formation. Nat. Chem. Biol. 2011, 7, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Burgos, R.A.; Conejeros, I.; Hidalgo, M.A.; Werling, D.; Hermosilla, C. Calcium influx, a new potential therapeutic target in the control of neutrophil- dependent inflammatory diseases in bovines. Vet. Immunol. Immunopathol. 2011, 143, 1–10. [Google Scholar] [CrossRef]
- Conejeros, I.; Jara, E.; Carretta, M.D.; Alarcón, P.; Hidalgo, M.A.; Burgos, R.A. 2-Aminoethoxydiphenyl borate (2-APB) reduces respiratory burst, MMP-9 release and CD11b expression, and increases l-selectin shedding in bovine neutrophils. Res. Vet. Sci. 2012, 92, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Villagra-Blanco, R.; Silva, L.M.R.; Gärtner, U.; Wagner, H.; Failing, K.; Wehrend, A.; Taubert, A.; Hermosilla, C. Molecular analyses on Neospora caninum-triggered NETosis in the caprine system. Dev. Comp. Immunol. 2017, 72, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Douda, D.N.; Khan, M.A.; Grasemann, H.; Palaniyar, N. SK3 channel and mitochondrial ROS mediate NADPH oxidase-independent NETosis induced by calcium influx. Proc. Natl. Acad. Sci. USA 2015, 112, 2817–2822. [Google Scholar] [CrossRef] [PubMed]
- Rochael, N.C.; Guimarães-Costa, A.B.; Nascimento, M.T.C.; De Souza-Vieira, T.S.; Oliveira, M.P.; Garcia e Souza, L.F.; Oliveira, M.F.; Saraiva, E.M. Classical ROS-dependent and early/rapid ROS-independent release of neutrophil extracellular traps triggered by Leishmania parasites. Sci. Rep. 2015, 5, 18302. [Google Scholar] [CrossRef]
- Stoiber, W.; Obermayer, A.; Steinbacher, P.; Krautgartner, W.D. The role of reactive oxygen species (ROS) in the formation of extracellular traps (ETs) in humans. Biomolecules 2015, 5, 702–723. [Google Scholar] [CrossRef]
- Malawista, S.E.; Van Blaricom, G.; Breitenstein, M.G. Cryopreservable neutrophil surrogates. Stored cytoplasts from human polymorphonuclear leukocytes retain chemotactic, phagocytic, and microbicidal function. J. Clin.Investig. 1989, 83, 728–732. [Google Scholar] [CrossRef]
- Yousefi, S.; Mihalache, C.; Kozlowski, E.; Schmid, I.; Simon, H.U. Viable neutrophils release mitochondrial DNA to form neutrophil extracellular traps. Cell Death Differ. 2009, 16, 1438–1444. [Google Scholar] [CrossRef] [PubMed]
- Yipp, B.G.; Petri, B.; Salina, D.; Jenne, C.N.; Scott, B.N.V.; Zbytnuik, L.D.; Pittman, K.; Asaduzzaman, M.; Wu, K.; Meijndert, H.C.; et al. Infection-induced NETosis is a dynamicprocess involving neutrophil multitasking in vivo. Nat. Med. 2012, 18, 1386–1393. [Google Scholar] [CrossRef] [PubMed]
- Schauer, C.; Janko, C.; Munoz, L.E.; Zhao, Y.; Kienhöfer, D.; Frey, B.; Lell, M.; Manger, B.; Rech, J.; Naschberger, E.; et al. Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines. Nat. Med. 2014, 20, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Coppens, I.; Dunn, J.D.; Romano, J.D.; Pypaert, M.; Zhang, H.; Boothroyd, J.C.; Joiner, K.A. Toxoplasma gondii sequesters lysosomes from mammalian hosts in the vacuolar space. Cell 2006, 125, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.M.; Jones, A.R.; Carmen, J.C.; Sinai, A.P.; Burchmore, R.; Wastling, J.M. Modulation of the host cell proteome by the intracellular apicomplexan parasite Toxoplasma gondii. Infect. Immun. 2007, 76, 828–844. [Google Scholar] [CrossRef]
- Hamid, P.H.; Hirzmann, J.; Hermosilla, C.; Taubert, A. Differential inhibition of host cell cholesterol de novo biosynthesis and processing abrogates Eimeria bovis intracellular development. Parasitol. Res. 2014, 113, 4165–4176. [Google Scholar] [CrossRef]
- Hamid, P.H.; Hirzmann, J.; Kerner, K.; Gimpl, G.; Lochnit, G.; Hermosilla, C.R.; Taubert, A. Eimeria bovis infection modulates endothelial host cell cholesterol metabolism for successful replication. Vet. Res. 2015, 46, 100. [Google Scholar] [CrossRef]
- Taubert, A.; Hermosilla, C.; Silva, L.M.R.; Wieck, A.; Failing, K.; Mazurek, S. Metabolic signatures of Besnoitia besnoiti-infected endothelial host cells and blockage of key metabolic pathways indicate high glycolytic and glutaminolytic needs of the parasite. Parasitol. Res. 2016, 115, 2023–2034. [Google Scholar] [CrossRef]
- Wei, Z.; Hermosilla, C.; Taubert, A.; He, X.; Wang, X.; Gong, P.; Li, J.; Yang, Z.; Zhang, X. Canine neutrophil extracellular traps release induced by the apicomplexan parasite Neospora caninum in vitro. Front. Immunol. 2016, 7, 436. [Google Scholar] [CrossRef]
- Behrendt, J.H.; Ruiz, A.; Zahner, H.; Taubert, A.; Hermosilla, C. Neutrophil extracellular trap formation as innate immune reactions against the apicomplexan parasite Eimeria bovis. Vet. Immunol. Immunopathol. 2010, 133, 1–8. [Google Scholar] [CrossRef]
- Villagra-Blanco, R.; Silva, L.M.R.; Muñoz-Caro, T.; Yang, Z.; Li, J.; Gärtner, U.; Taubert, A.; Zhang, X.; Hermosilla, C. Bovine PMN cast neutrophil extracellular traps against the abortive parasite Neospora caninum. Front. Immunol. 2017, 8, 606. [Google Scholar] [CrossRef]
- Yang, Z.; Wei, Z.; Hermosilla, C.; Taubert, A.; He, X.; Wang, X.; Gong, P.; Li, J.; Zhang, X. Caprine monocytes release extracellular traps against Neospora caninum in vitro. Front. Immunol. 2018, 8. [Google Scholar] [CrossRef]
- Yildiz, K.; Gokpinar, S.; Gazyagci, A.N.; Babur, C.; Sursal, N.; Azkur, A.K. Role of NETs in the difference in host susceptibility to Toxoplasma gondii between sheep and cattle. Vet. Immunol. Immunopathol. 2017, 189, 1–10. [Google Scholar] [CrossRef]
- Van Wormer, E.; Conrad, P.A.; Miller, M.A.; Melli, A.C.; Carpenter, T.E.; Mazet, J. Toxoplasma gondii, source to sea: Higher contribution of domestic felids to terrestrial parasite loading despite lower infection prevalence. EcoHealth 2013. [Google Scholar] [CrossRef] [PubMed]
- Begde, D.; Bundale, S.; Mashitha, P.; Rudra, J.; Nashikkar, N.; Upadhyay, A. Immunomodulatory efficacy of nisin—A bacterial lantibiotic peptide. J. Pept. Sci. 2011, 17, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Pullar, J.M.; Vissers, M.C.; Winterbourn, C.C. Living with a killer: The effects of hypochlorous acid on mammalian cells. IUBMB Life 2000, 50, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Klebanoff, S.J. Myeloperoxidase: Friend and foe. J. Leukoc. Biol. 2005, 77, 598–625. [Google Scholar] [CrossRef] [PubMed]
- De Buhr, N.; von Köckritz-Blickwede, M. How Neutrophil Extracellular Traps Become Visible. J. Immunol. Res. 2016, 4604713. [Google Scholar] [CrossRef] [PubMed]
- Itou, T.; Sugisawa, H.; Inoue, Y.; Jinbo, T.; Sakai, T. Oxygen radical generationand expression of NADPH oxidase genes in bottlenose dolphin (Tursiops truncatus) neutrophils. Dev. Comp. Immunol. 2001, 25, 47–53. [Google Scholar] [CrossRef]
- Von Köckritz-Blickwede, M.; Chow, O.; Ghochani, M.; Nizet, V. Visualization and functional evaluation of phagocyte extracellular traps. Methods Microbiol. 2010, 37, 139–160. [Google Scholar]
- Von Köckritz-Blickwede, M.; Nizet, V. Innate immunity turned inside-out: Antimicrobial defense by phagocyte extracellular traps. J. Mol. Med. 2009, 87, 775–783. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villagra-Blanco, R.; Silva, L.M.R.; Conejeros, I.; Taubert, A.; Hermosilla, C. Pinniped- and Cetacean-Derived ETosis Contributes to Combating Emerging Apicomplexan Parasites (Toxoplasma gondii, Neospora caninum) Circulating in Marine Environments. Biology 2019, 8, 12. https://doi.org/10.3390/biology8010012
Villagra-Blanco R, Silva LMR, Conejeros I, Taubert A, Hermosilla C. Pinniped- and Cetacean-Derived ETosis Contributes to Combating Emerging Apicomplexan Parasites (Toxoplasma gondii, Neospora caninum) Circulating in Marine Environments. Biology. 2019; 8(1):12. https://doi.org/10.3390/biology8010012
Chicago/Turabian StyleVillagra-Blanco, Rodolfo, Liliana M. R. Silva, Iván Conejeros, Anja Taubert, and Carlos Hermosilla. 2019. "Pinniped- and Cetacean-Derived ETosis Contributes to Combating Emerging Apicomplexan Parasites (Toxoplasma gondii, Neospora caninum) Circulating in Marine Environments" Biology 8, no. 1: 12. https://doi.org/10.3390/biology8010012
APA StyleVillagra-Blanco, R., Silva, L. M. R., Conejeros, I., Taubert, A., & Hermosilla, C. (2019). Pinniped- and Cetacean-Derived ETosis Contributes to Combating Emerging Apicomplexan Parasites (Toxoplasma gondii, Neospora caninum) Circulating in Marine Environments. Biology, 8(1), 12. https://doi.org/10.3390/biology8010012