
Ornithine Aminotransferase, an Important Glutamate-Metabolizing Enzyme at the Crossroads of Multiple Metabolic Pathways


Abstract
:1. Introduction
1.1. Physical And Chemical Aspects of the Oat-Mediated Reaction
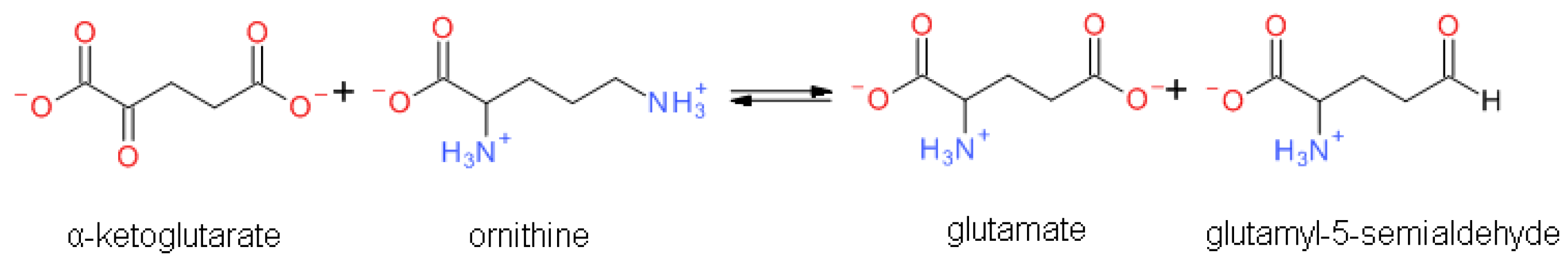
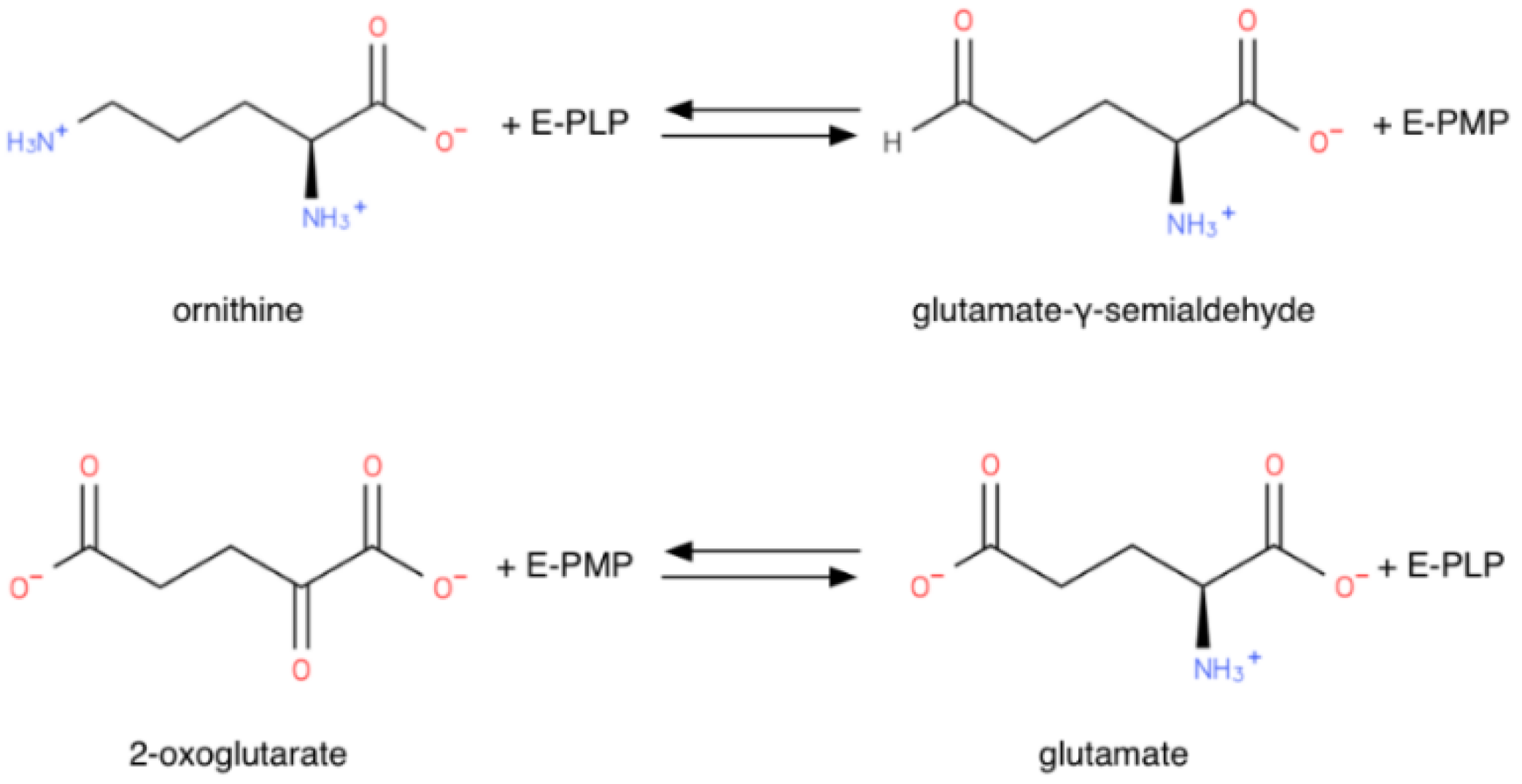
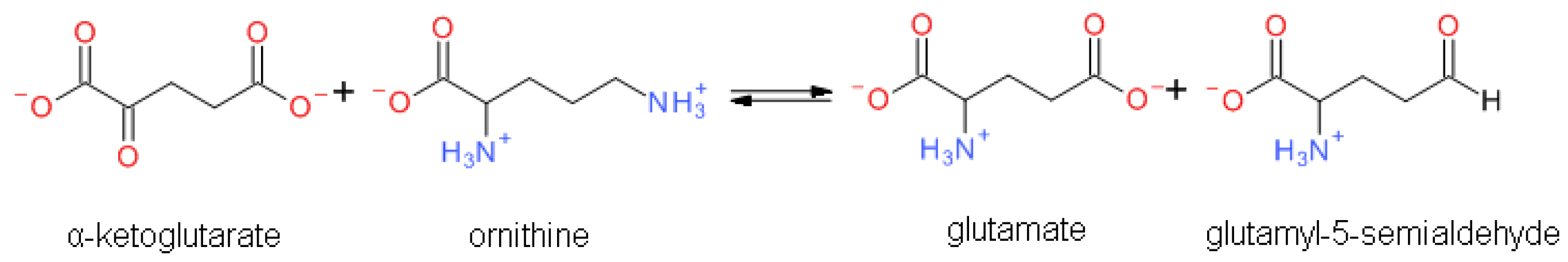
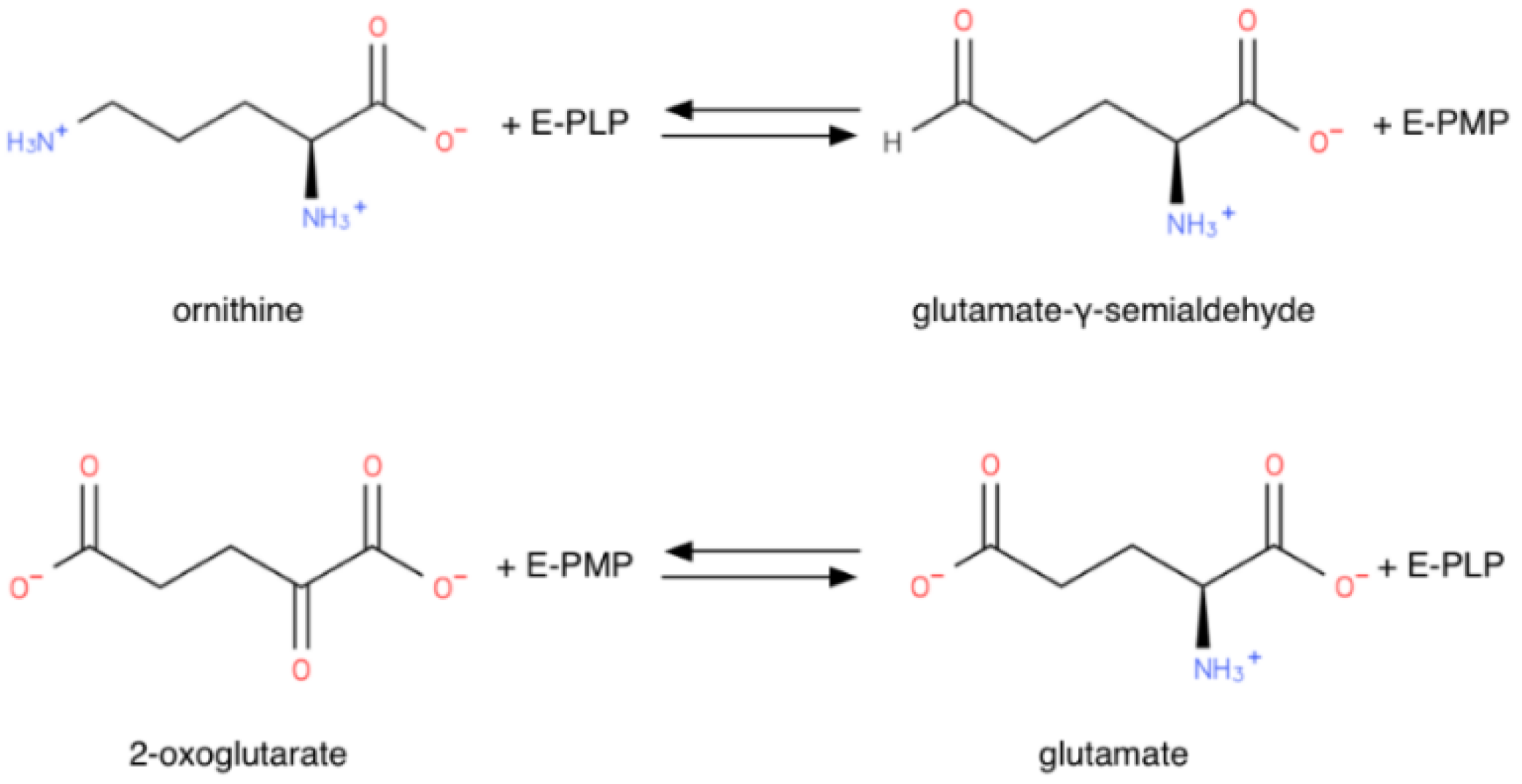
1.1.1. Reaction
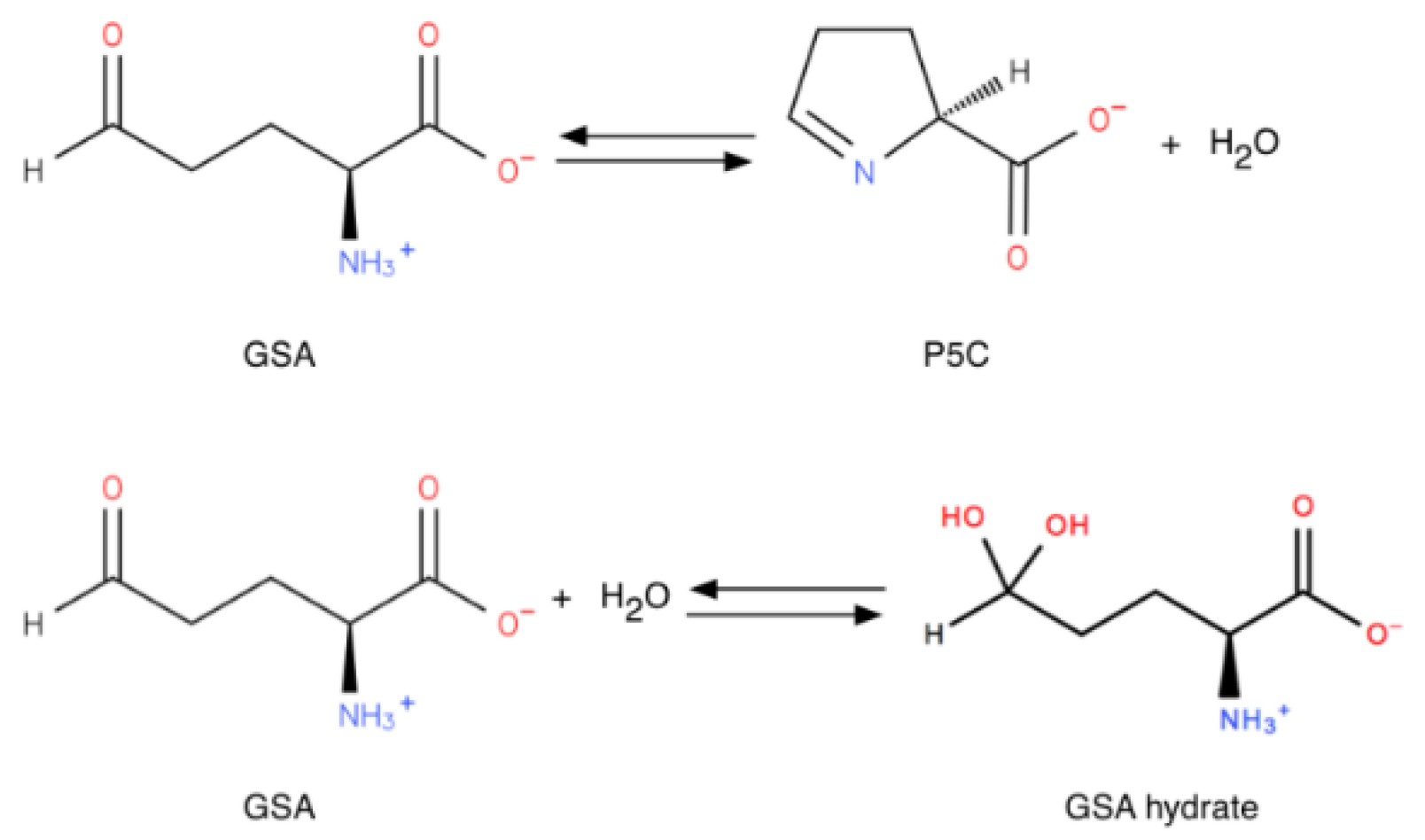
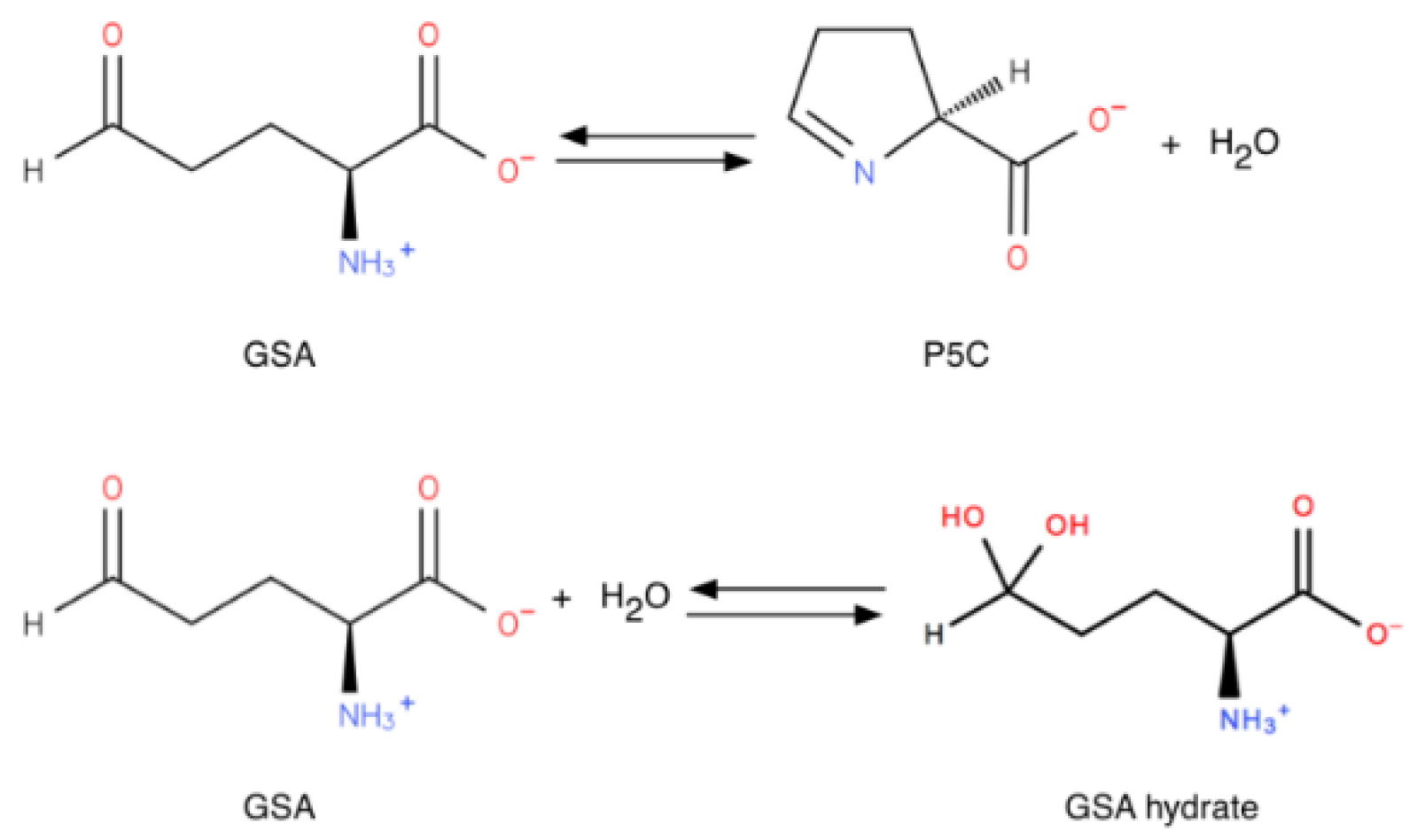
1.1.2. A Key Feature: Spontaneous Cyclization of GSA
1.1.3. Kinetic Characteristics of OAT
1.2. Inhibitors
2. Structure of OAT
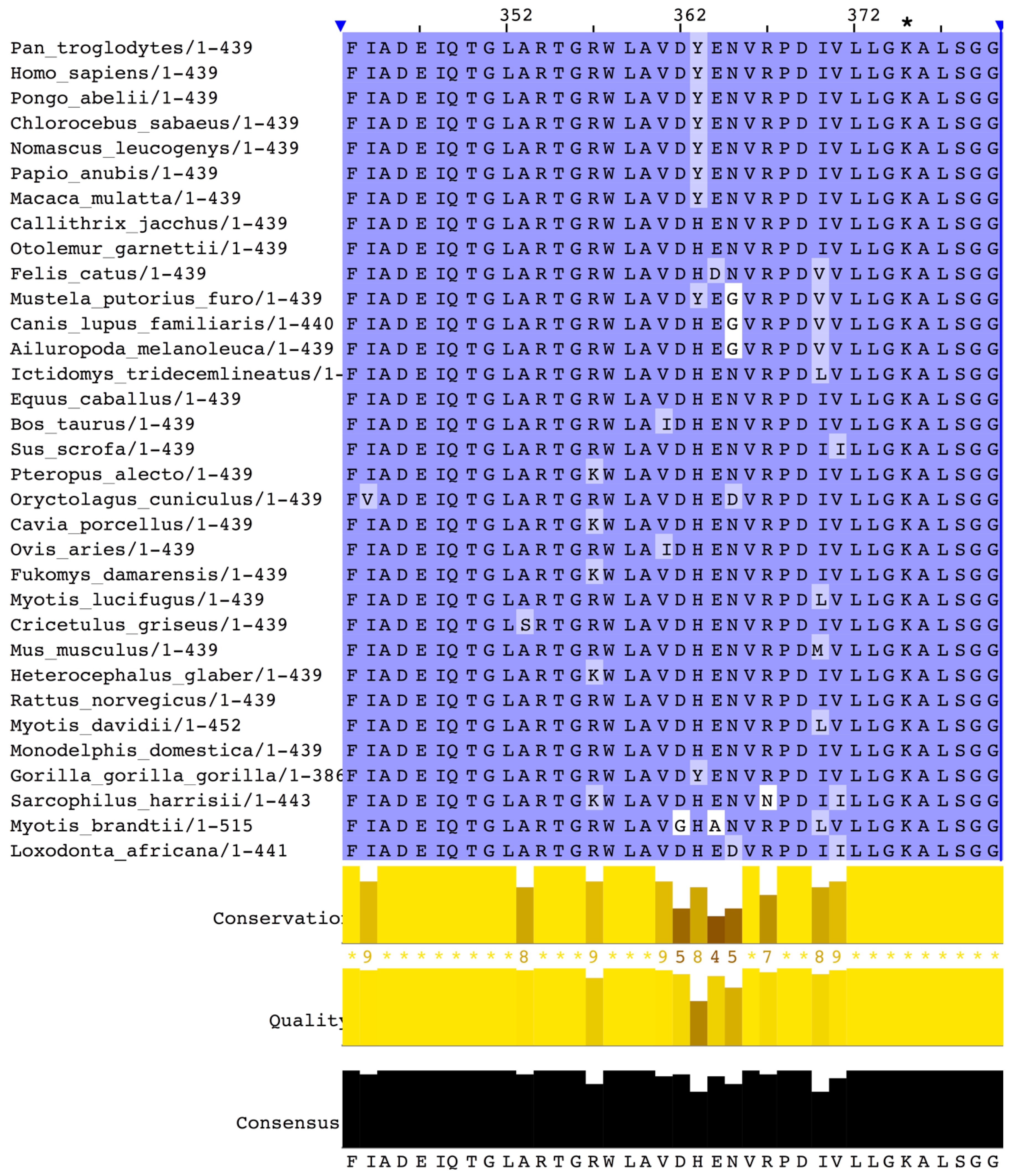
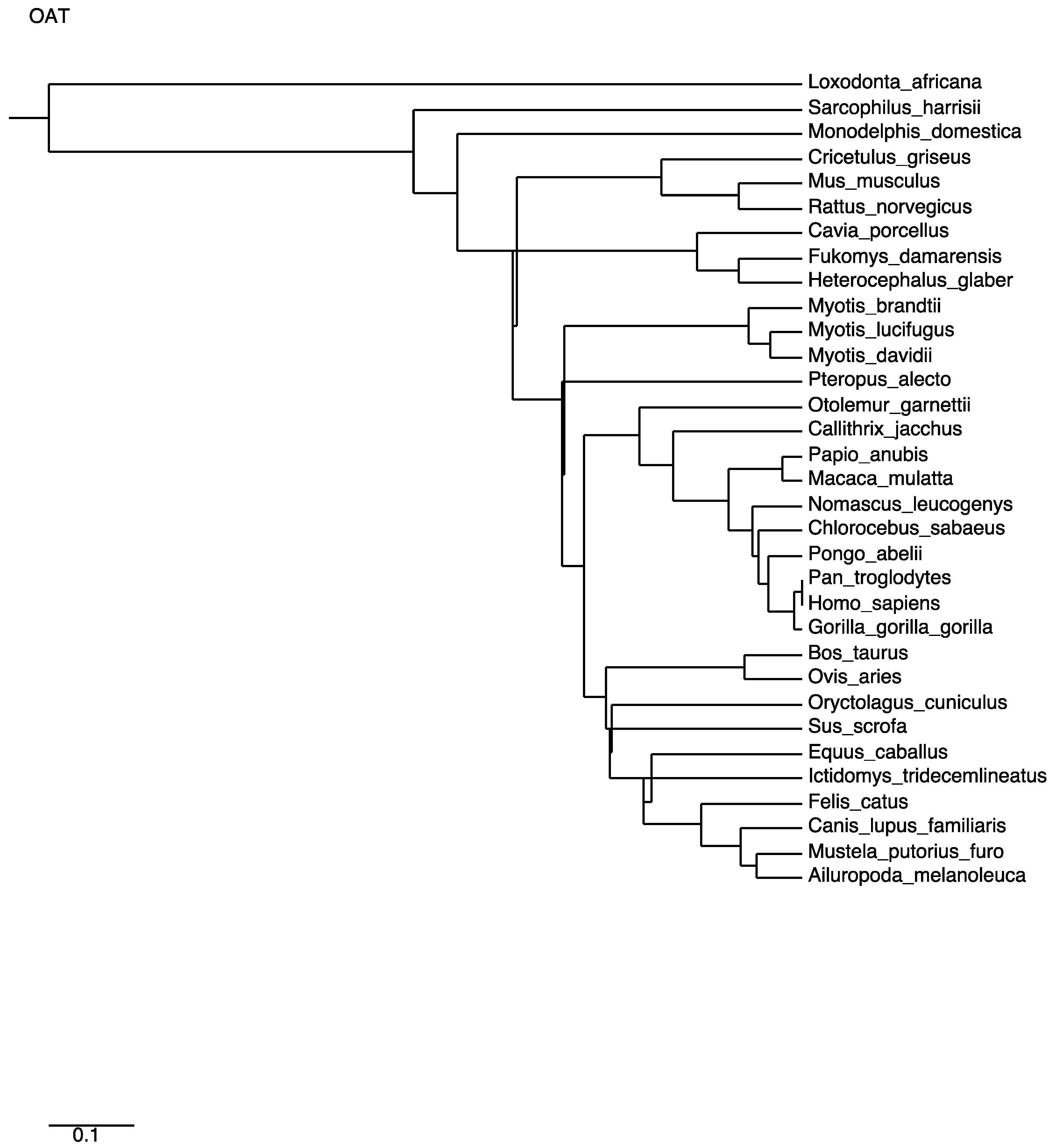
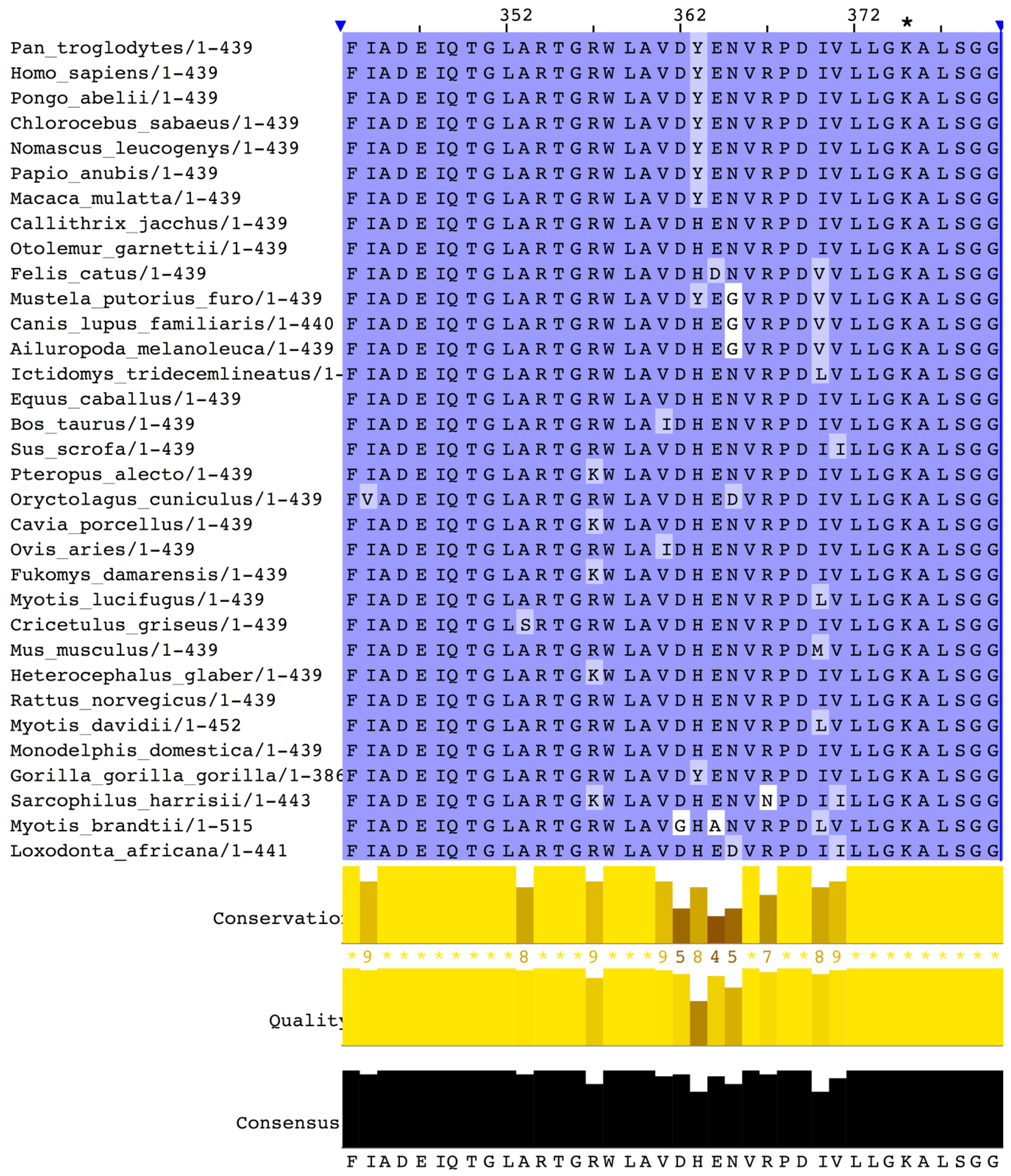
2.1. Primary Sequence and Leading Peptide
2.2. The Three-Dimensional Structure
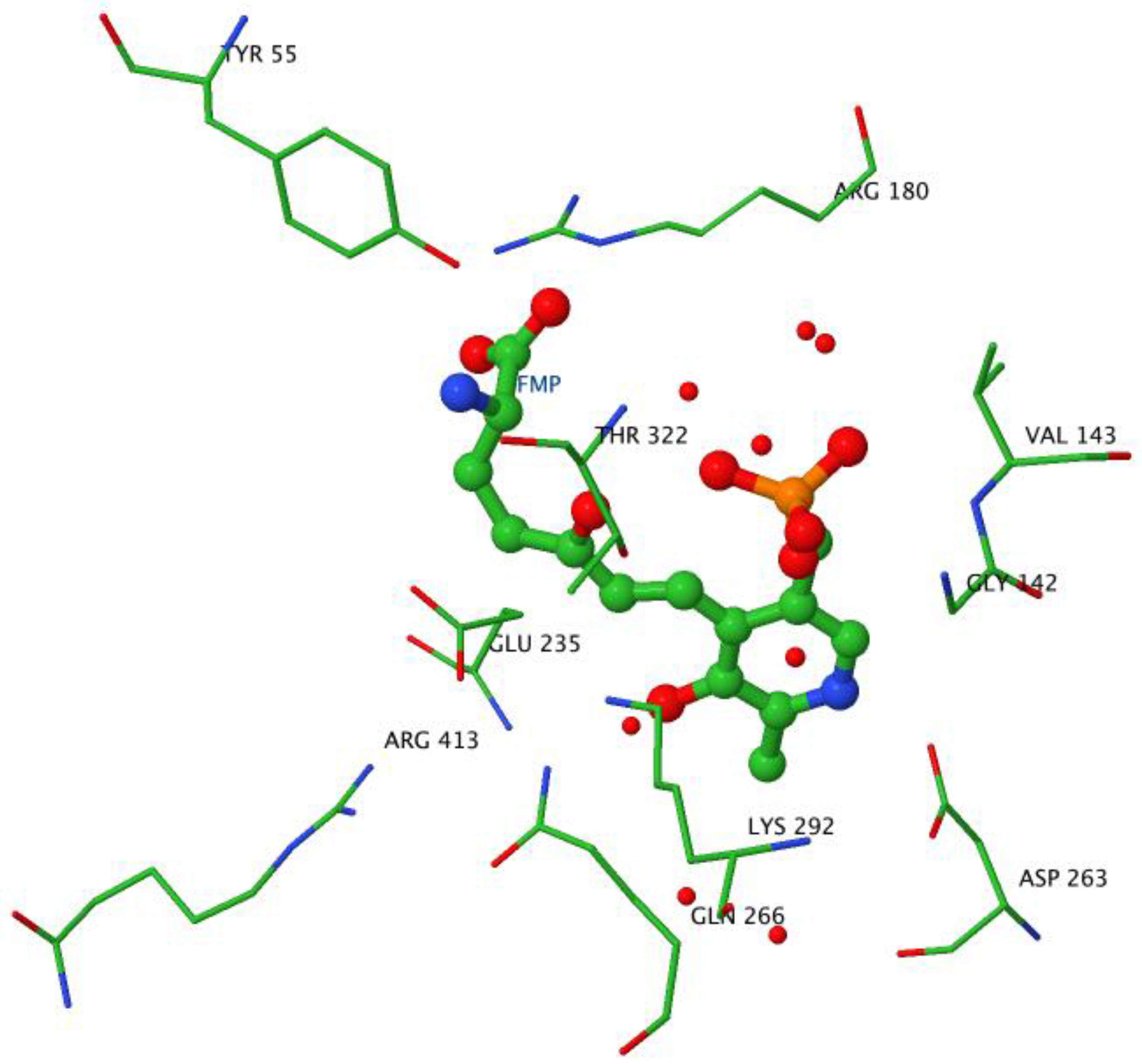
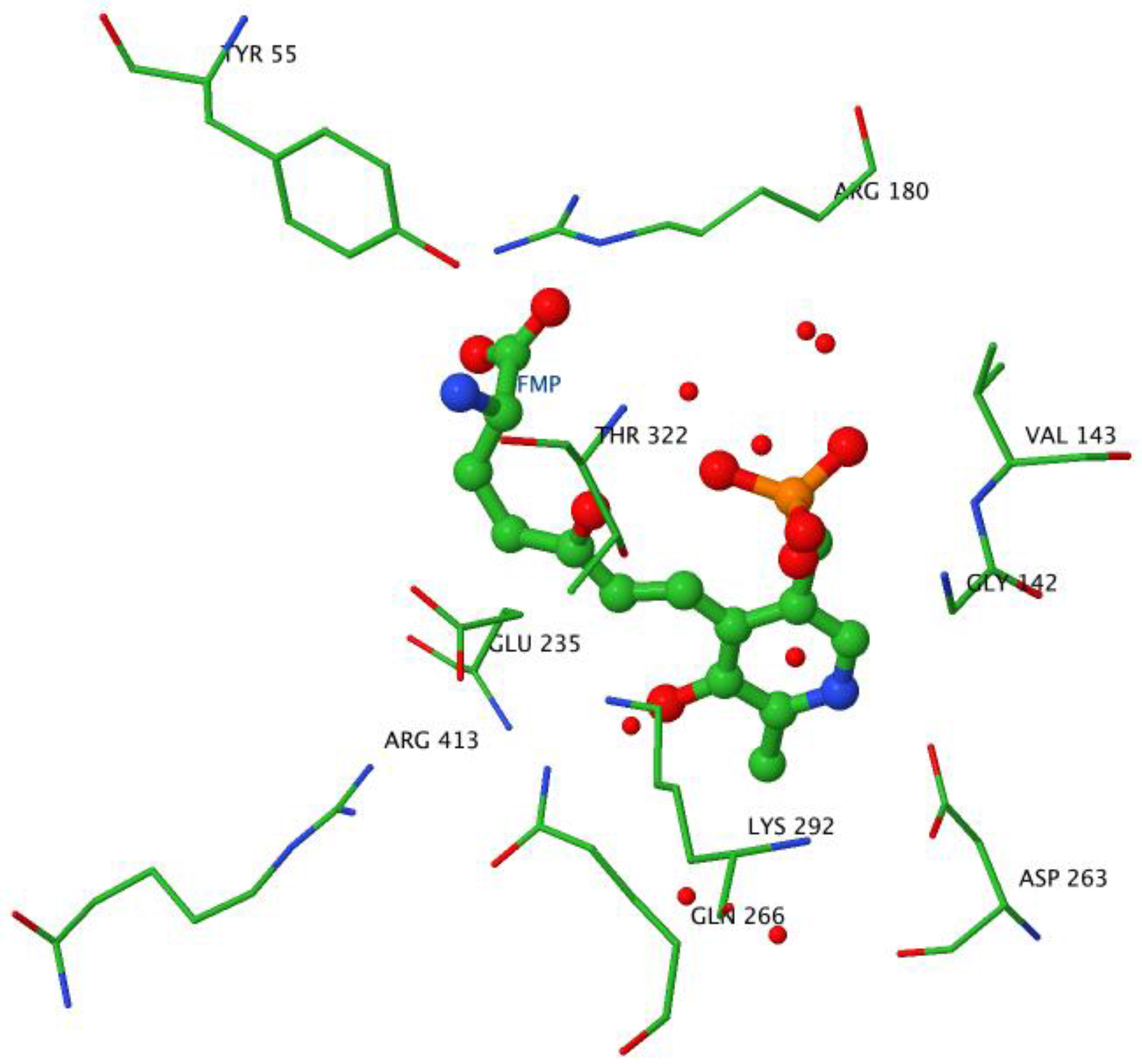
2.3. The Active Site
3. Synthesis and Fate of OAT—From Gene to Protein
3.1. From Gene to Protein
3.1.1. Genes Encoding OAT
3.1.2. Transcription
3.1.3. Translation
3.2. Changes in Protein
3.2.1. Peptide Maturation
3.2.2. Post-Translational Modifications
3.3. Life and Death of the Protein
3.3.1. Interaction with Other Proteins
3.3.2. OAT Degradation
4. Localization of OAT
4.1. Cellular Localization
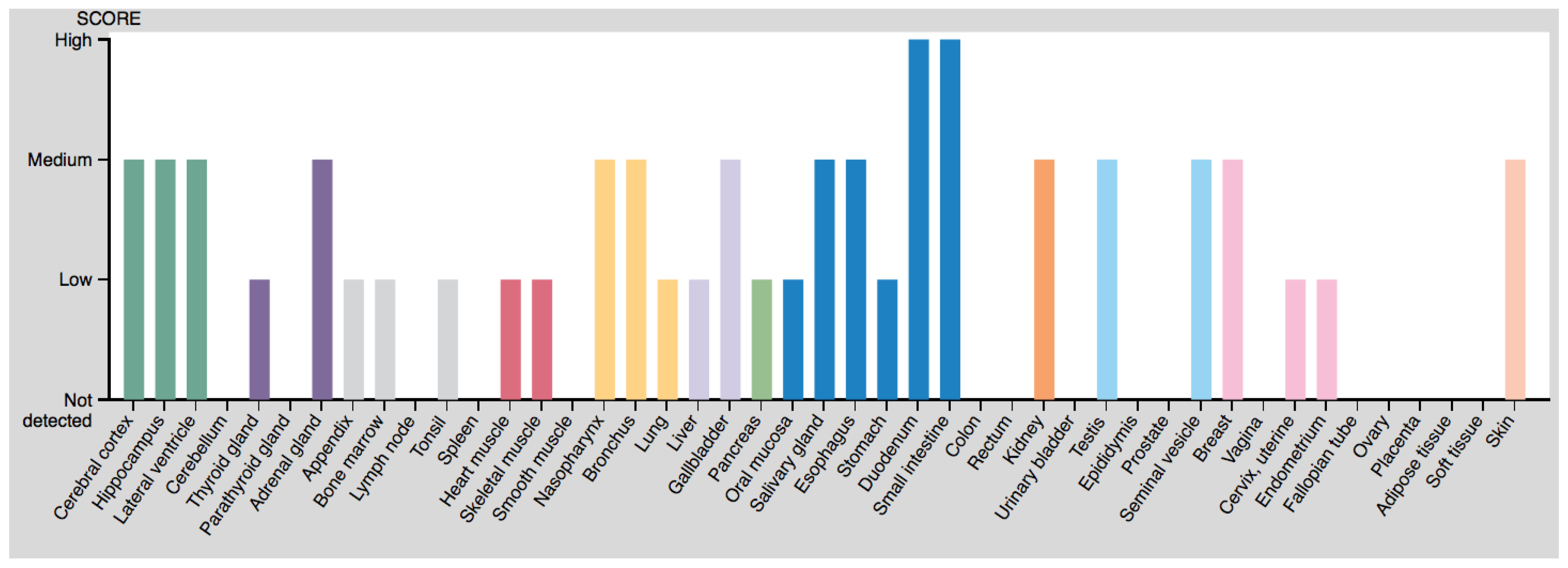
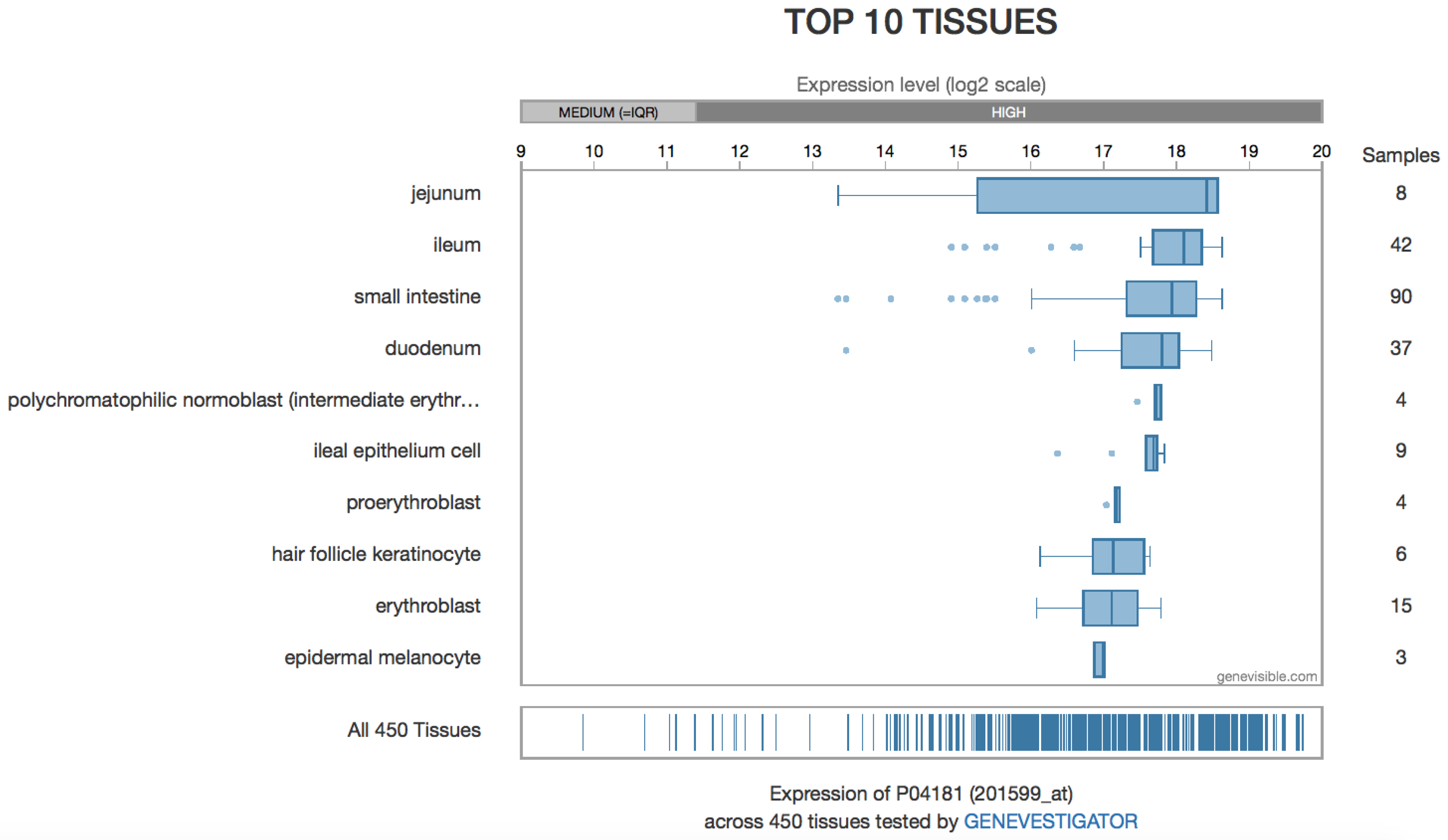
4.2. Tissue Localization
4.3. OAT in the Liver
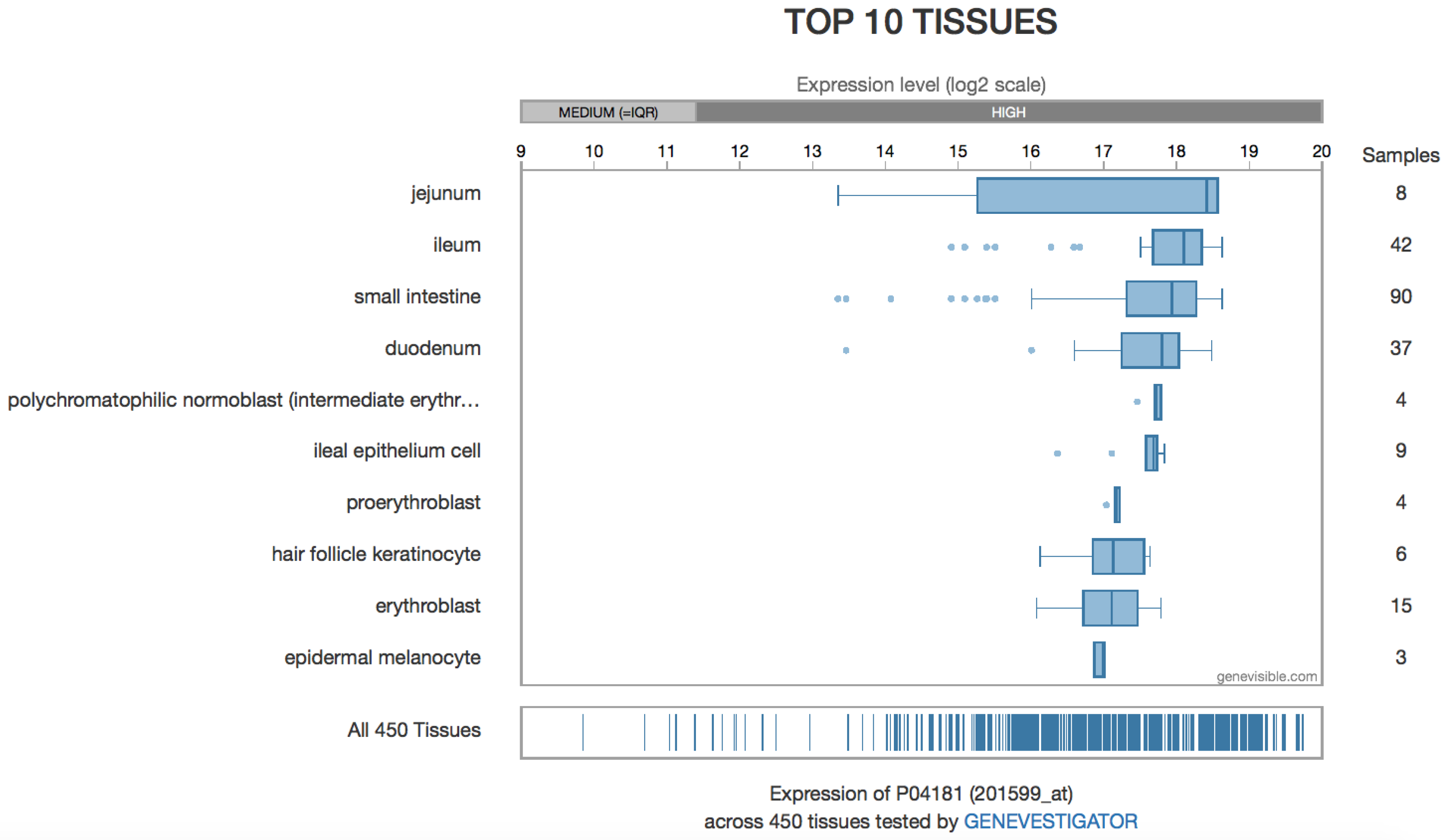
4.4. OAT in the Digestive Tract
4.5. OAT in the Kidney
4.6. OAT in the Brain and the Nervous and Sensory System
4.7. OAT in the Other Tissues
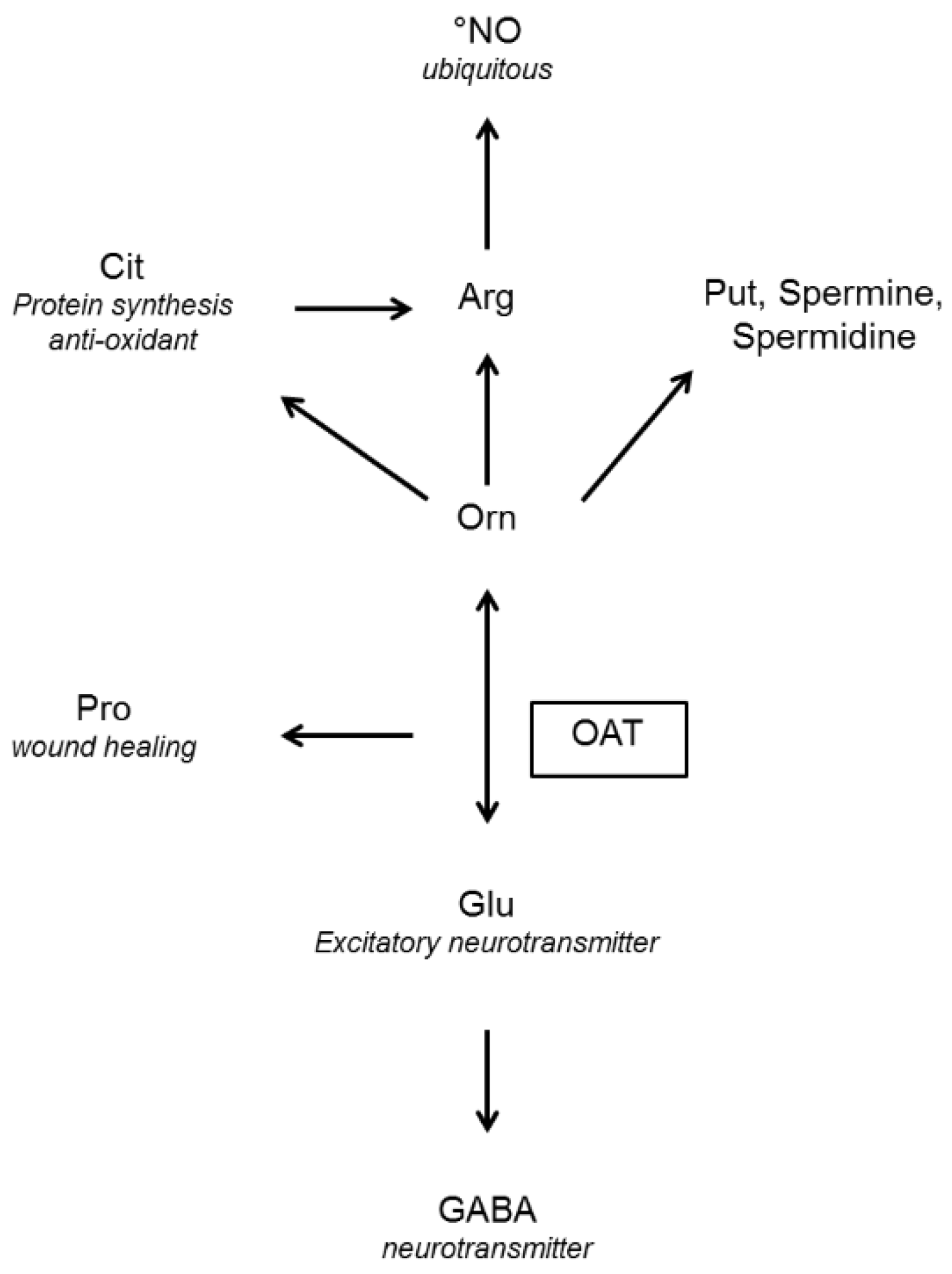
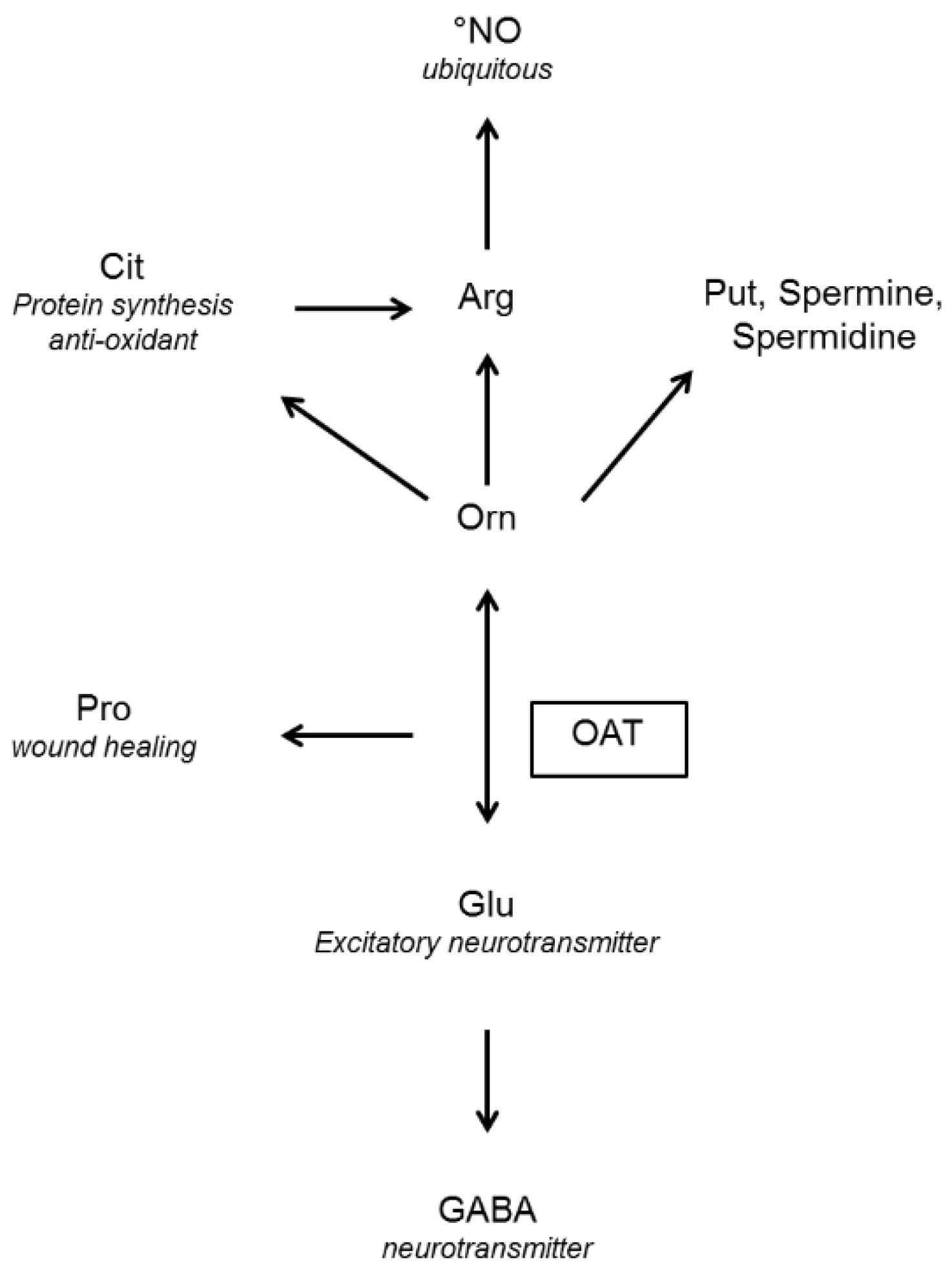
5. The Role of OAT in Ornithine Fluxes
5.1. Fluxes in Non-Mammals: Orn Degradation
5.1.1. Prokaryotes
5.1.2. Plants
5.1.3. Fungi
5.1.4. Unicellular Eukaryotes
5.2. Fluxes in Mammals: A Subtle Compromise between Orn Degradation and Orn Synthesis
5.2.1. OAT and Orn Degradation
5.2.2. OAT and Orn Synthesis
6. OAT in Health and Disease
6.1. OAT and the Development of Mammals
6.1.1. OAT and Arg Supply in the Pre-Weaning Period
6.1.2. Changes in Tissue-Specific OAT Expression
6.1.2.1. Pre-Weaning Period
6.1.2.2. Weaning Period
6.1.2.3. Post-Weaning Period and Puberty
6.1.3. Other Implications
6.2. Consequences of OAT Deficiency: Gyrate Atrophy of the Choroid and Retina (GA)
6.2.1. Overview
6.2.2. Pathophysiology
6.2.3. Diagnosis
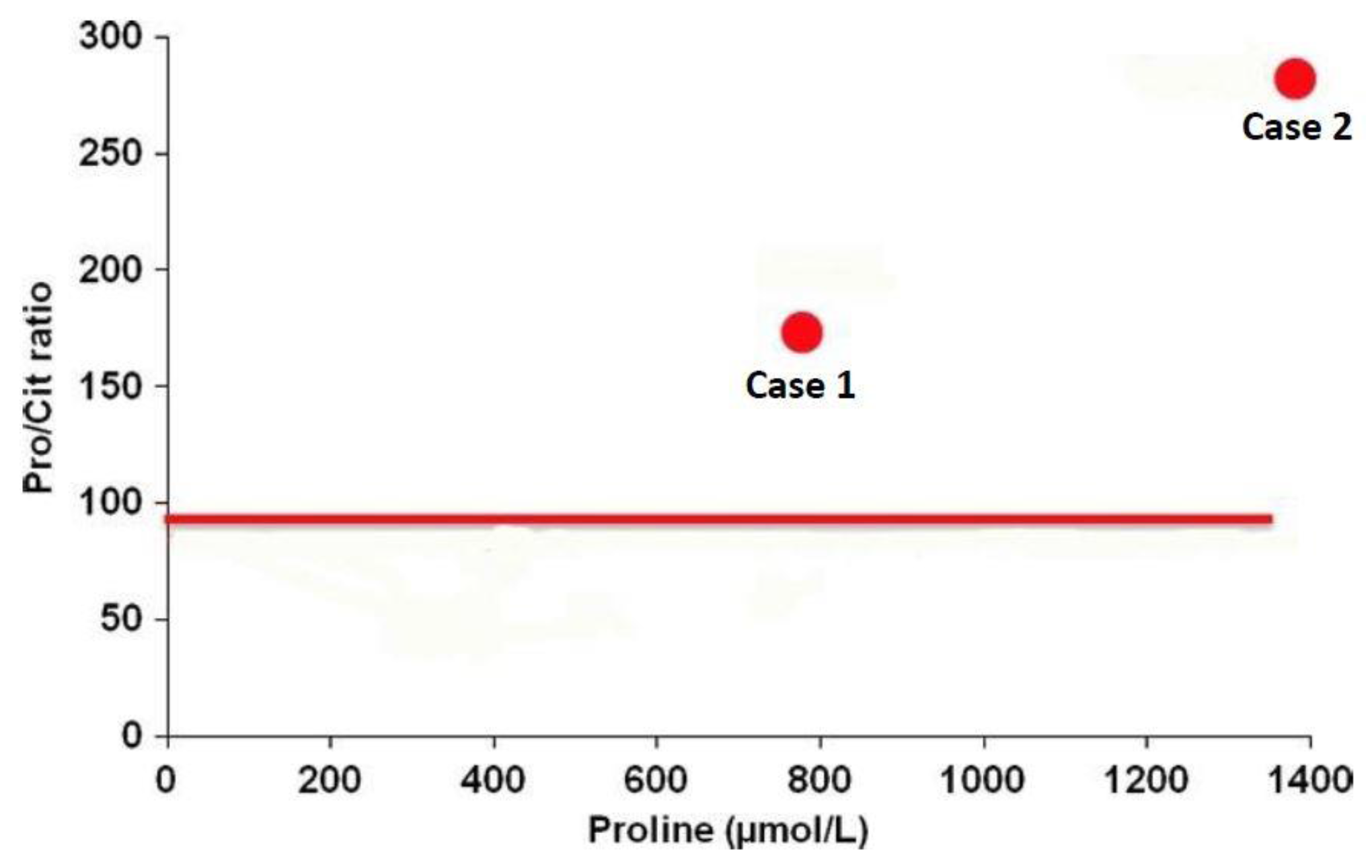
6.2.4. Diagnosis in Newborns
6.2.5. Therapeutic Approaches
6.3. OAT Inhibition As A Possible Therapeutic Strategy
6.3.1. Potential Utility of OAT Inhibitors for the Treatment of Cancer
6.3.2. Treatment of Hyperammonemia
6.3.3. OAT in Sepsis
7. Conclusions
Acknowledgments
Conflicts of interest
References
- Yasuo, W. The Glutamate Crossway. In Metabolic and Therapeutic Aspects of Amino Acids in Clinical Nutrition; Cynober, L.A., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 135–152. [Google Scholar]
- Morris, S.M. Arginine: Beyond protein. Am. J. Clin. Nutr. 2006, 83, 508S–512S. [Google Scholar] [PubMed]
- Melis, G.C.; Ter Wengel, N.; Boelens, P.G.; Van Leeuwen, P.A.M. Glutamine: Recent developments in research on the clinical significance of glutamine. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Breuillard, C.; Cynober, L.; Moinard, C. Citrulline and nitrogen homeostasis: An overview. Amino Acids 2015, 47, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Ramani, D.; De Bandt, J.P.; Cynober, L. Aliphatic polyamines in physiology and diseases. Clin. Nutr. 2014, 33, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Strecker, H.J. Purification and properties of rat liver ornithine δ-transaminase. J. Biol. Chem. 1965, 240, 1225–1230. [Google Scholar] [PubMed]
- Schöpf, C.; Oechler, F. Zur Frage der Biogenese des Vasicins (Peganins). Die Synthese des Desoxyvasicins unter physiologischen Bedingungen. Justus Liebigs Ann. Chem. 1936, 523, 1–29. (In German) [Google Scholar] [CrossRef]
- Schöpf, C.; Steuer, H. Zur Frage der Biogenese des Rutaecarpins und Evodamins. Die Synthese des Rutaecarpins unter zellmöglichen Bedingungen. Justus Liebigs Ann. Chem. 1947, 558, 124–136. (In German) [Google Scholar] [CrossRef]
- Mezl, V.A.; Knox, W.E. Properties and analysis of a stable derivative of pyrroline-5-carboxylic acid for use in metabolic studies. Anal. Biochem. 1976, 74, 430–440. [Google Scholar] [CrossRef]
- Lewis, M.L.; Rowe, C.J.; Sewald, N.; Sutherland, J.D.; Wilson, E.J.; Wright, M.C. The effect of pH on the solution structure of Δ1-pyrroline-2-carboxylic acid as revealed by NMR and electrospray mass spectroscopy. Bioorg. Med. Chem. Lett. 1993, 3, 1193–1196. [Google Scholar] [CrossRef]
- Bearne, S.L.; Wolfenden, R. Glutamate gamma-semialdehyde as a natural transition state analog inhibitor of escherichia coli glucosamine-6-phosphate synthase. Biochemistry 1995, 34, 11515–11520. [Google Scholar] [CrossRef]
- Shen, B.W.; Hennig, M.; Hohenester, E.; Jansonius, J.N.; Schirmer, T. Crystal structure of human recombinant ornithine aminotransferase. J. Mol. Biol. 1998, 277, 81–102. [Google Scholar] [CrossRef] [PubMed]
- Marvin Calculator Plugins. Available online: https://www.chemaxon.com/ (accessed on 6 September 2015).
- Matsuzawa, T. Characteristics of the inhibition of ornithine-δ-aminotransferase by branched-chain amino acids. J. Biochem. 1974, 75, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Rao, G.N.; Cotlier, E. Ornithine delta-aminotransferase activity in retina and other tissues. Neurochem. Res. 1984, 9, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Shiono, T.; Hayasaka, S.; Mizuno, K. Partial purification and certain properties of ornithine ketoacid aminotransferases in the ciliary body and iris and the retinal pigment epithelium of the bovine eye. Exp. Eye Res. 1981, 32, 475–490. [Google Scholar] [CrossRef]
- Burcham, J.M.; Giometti, C.S.; Tollaksen, S.L.; Peraino, C. Comparison of rat and mouse ornithine aminotransferase with respect to molecular properties and regulation of synthesis. Arch. Biochem. Biophys. 1988, 262, 501–507. [Google Scholar] [CrossRef]
- Sanada, Y.; Suemori, I.; Katunuma, N. Properties of ornithine aminotransferase from rat liver, kidney and small intestine. Biochim. Biophys. Acta Enzymol. 1970, 220, 42–50. [Google Scholar] [CrossRef]
- Peraino, C. Functional properties of ornithine-ketoacid aminotransferase from rat liver. Biochim. Biophys. Acta 1972, 289, 117–127. [Google Scholar] [CrossRef]
- Daune, G.; Seiler, N. Interrelationships between ornithine, glutamate, and GABA-III. An ornithine aminotransferase activity that is resistant to inactivation by 5-fluoromethylornithine. Neurochem. Int. 1988, 13, 383–391. [Google Scholar] [CrossRef]
- Wekell, M.M.; Brown, G.W. Ornithine aminotransferase of fishes. Comp. Biochem. Physiol. B 1973, 46, 779–795. [Google Scholar] [CrossRef]
- McGivan, J.D.; Bradford, N.M.; Beavis, A.D. Factors influencing the activity of ornithine aminotransferase in isolated rat liver mitochondria. Biochem. J. 1977, 162, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, D.R.; Srivastava, S.K. Purification and properties of ornithine aminotransferase from rat brain. Experientia 1984, 40, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Drejer, J.; Schousboe, A. Ornithine-delta-aminotransferase exhibits different kinetic properties in astrocytes, cerebral cortex interneurons, and cerebellar granule cells in primary culture. J. Neurochem. 1984, 42, 1194–1197. [Google Scholar] [CrossRef] [PubMed]
- Bishop, S.H.; Ellis, L.L.; Burcham, J.M. Amino Acid Metabolism in Molluscs. In The Mollusca; Academic Press: New York, NY, USA, 1983; pp. 243–327. [Google Scholar]
- Gafan, C.; Wilson, J.; Berger, L.C.; Berger, B.J. Characterization of the ornithine aminotransferase from Plasmodium falciparum. Mol. Biochem. Parasitol. 2001, 118, 1–10. [Google Scholar] [CrossRef]
- Stránská, J.; Tylichová, M.; Kopecný, D.; Snégaroff, J.; Sebela, M. Biochemical characterization of pea ornithine-delta-aminotransferase: Substrate specificity and inhibition by di- and polyamines. Biochimie 2010, 92, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Delauney, A.J.; Hu, C.A.; Kishor, P.B.; Verma, D.P. Cloning of ornithine delta-aminotransferase cDNA from Vigna aconitifolia by trans-complementation in Escherichia coli and regulation of proline biosynthesis. J. Biol. Chem. 1993, 268, 18673–18678. [Google Scholar] [PubMed]
- Fagan, R.J.; Lazaris-Karatzas, A.; Sonenberg, N.; Rozen, R. Translational control of ornithine aminotransferase. Modulation by initiation factor eIF-4E. J. Biol. Chem. 1991, 266, 16518–16523. [Google Scholar] [PubMed]
- Boernke, W.E.; Stevens, F.J.; Edwards, J.J.; Peraino, C. Differential changes in ornithine aminotransferase self-affinity produced by exposure to basic amino acids and increases in the intrinsic electronegativity of the enzyme monomer. Arch. Biochem. Biophys. 1982, 216, 152–157. [Google Scholar] [CrossRef]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L.; Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry: International Version (Hardcover). Available online: http://www.sidalc.net/cgi-bin/wxis.exe/?IsisScript=AGRIUAN.xis&method=post&formato=2&cantidad=1&expresion=mfn=029110 (accessed on 6 September 2002).
- Williams, J.A.; Bridge, G.; Fowler, L.J.; John, R.A. The reaction of ornithine aminotransferase with ornithine. Biochem. J. 1982, 201, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Ilan, Y.; Zigmond, E.; Silverman, R.B.; Lu, H. Inhibition of ornithine aminotransferase for the treatment of proliferative disorders. U.S. patent 8686041 B2; issued April 2014,
- Lee, H.; Juncosa, J.I.; Silverman, R.B. ornithine aminotransferase versus GABA Aminotransferase: Implications for the design of new anticancer drugs. Med. Res. Rev. 2015, 35, 286–305. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.D.; Basford, J.M.; John, R.A. An investigation of the properties of ornithine aminotransferase after inactivation by the “suicide” inhibitor aminohexynoate and use of the compound as a probe of intracellullar protein turnover. Biochem. J. 1983, 209, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Ducep, J.B.; Heintzelmann, B.; Jund, K.; Lesur, B.; Schleimer, M.; Zimmermann, P.R. Synthesis of (2S,5S)-5-fluoromethylornithine; a potent inhibitor of ornithine aminotransferase. Tetrahedron Asymmetry 1997, 8, 327–335. [Google Scholar] [CrossRef]
- Daune, G.; Gerhart, F.; Seiler, N. 5-Fluoromethylornithine, an irreversible and specific inhibitor of L-ornithine: 2-oxo-acid aminotransferase. Biochem. J. 1988, 253, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Daune, G.; Seiler, N. Interrelationships between ornithine, glutamate, and GABA. II. Consequences of inhibition of GABA-T and ornithine aminotransferase in brain. Neurochem. Res. 1988, 13, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Bolkenius, F.N.; Knödgen, B.; Seiler, N. DL-canaline and 5-fluoromethylornithine. Comparison of two inactivators of ornithine aminotransferase. Biochem. J. 1990, 268, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Kito, K.; Sanada, Y.; Katunuma, N. Mode of inhibition of ornithine aminotransferase by L-canaline. J. Biochem. 1978, 83, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Kalita, C.C.; Kerman, J.D.; Strecker, H.J. Preparation and properties of ornithine-oxo-acid aminotransferase of rat kidney comparison with the liver enzyme. Biochim. Biophys. Acta Enzymol. 1976, 429, 780–797. [Google Scholar] [CrossRef]
- Seiler, N.; Spraggs, H.; Daune, G. Interrelationships between ornithine, glutamate and GABA-I. Feed-back inhibition of ornithine aminotransferase by elevated brain GABA levels. Neurochem. Int. 1987, 10, 391–397. [Google Scholar] [CrossRef]
- Jung, M.J.; Seiler, N. Enzyme-activated irreversible inhibitors of L-ornithine: 2-oxoacid aminotransferase. Demonstration of mechanistic features of the inhibition of ornithine aminotransferase by 4-aminohex-5-ynoic acid and gabaculine and correlation with in vivo activity. J. Biol. Chem. 1978, 253, 7431–7439. [Google Scholar] [PubMed]
- Seiler, N. Ornithine aminotransferase, a potential target for the treatment of hyperammonemias. Curr. Drug Targets 2000, 1, 119–153. [Google Scholar] [CrossRef] [PubMed]
- Volpe, P.; Menna, T.; Pagano, G. Ornithine-δ-transaminase heterogeneity and regulation. Eur. J. Biochem. 1974, 44, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Volpe, P.; Sawamura, R.; Strecker, H.J. Control of ornithine δ-transaminase in rat liver and kidney. J. Biol. Chem. 1969, 244, 719–726. [Google Scholar] [PubMed]
- Yip, M.C.; Collins, R.K. Purification and properties of rat kidney and liver ornithine aminotransferase. Enzyme 1971, 12, 187–200. [Google Scholar] [PubMed]
- Kobayashi, T.; Nishii, M.; Takagi, Y.; Titani, K.; Matsuzawa, T. Molecular cloning and nucleotide sequence analysis of mRNA for human kidney ornithine aminotransferase: An examination of ornithine aminotransferase isozymes between liver and kidney. FEBS Lett. 1989, 255, 300–304. [Google Scholar] [CrossRef]
- Simmaco, M.; John, R.A.; Barra, D.; Bossa, F. The primary structure of ornithine aminotransferase: Identification of active-site sequence and site of post-translational proteolysis. FEBS Lett. 1986, 199, 39–42. [Google Scholar] [CrossRef]
- Oyama, R.; Suzuki, M.; Matsuzawa, T.; Titani, K. Complete amino acid sequence of rat kidney ornithine aminotransferase: Identity with liver ornithine aminotransferase. J. Biochem. 1990, 108, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Inana, G.; Totsuka, S.; Redmond, M.; Dougherty, T.; Nagle, J.; Shiono, T.; Ohura, T.; Kominami, E.; Katunuma, N. Molecular cloning of human ornithine aminotransferase mRNA. Proc. Natl. Acad. Sci. USA 1986, 83, 1203–1207. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M.M.; Pitot, H.C. Sequence of the precursor to rat ornithine aminotransferase deduced from a cDNA clone. J. Biol. Chem. 1985, 260, 12993–12997. [Google Scholar] [PubMed]
- Sigrist, C.J.A.; De Castro, E.; Cerutti, L.; Cuche, B.A.; Hulo, N.; Bridge, A.; Bougueleret, L.; Xenarios, I. New and continuing developments at PROSITE. Nucleic Acids Res. 2013, 41, D344–D347. [Google Scholar] [CrossRef] [PubMed]
- Grishin, N.V.; Phillips, M.A.; Goldsmith, E.J. Modeling of the spatial structure of eukaryotic ornithine decarboxylases. Protein Sci. Publ. Protein Soc. 1995, 4, 1291–1304. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.K.; Hale, T.I.; Christen, P. Aminotransferases: demonstration of homology and division into evolutionary subgroups. Eur. J. Biochem. 1993, 214, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Steffen-Munsberg, F.; Vickers, C.; Kohls, H.; Land, H.; Mallin, H.; Nobili, A.; Skalden, L.; Van Den Bergh, T.; Joosten, H.-J.; Berglund, P.; et al. Bioinformatic analysis of a PLP-dependent enzyme superfamily suitable for biocatalytic applications. Biotechnol. Adv. 2015, 33, 566–604. [Google Scholar] [CrossRef] [PubMed]
- Schiroli, D.; Peracchi, A. A subfamily of PLP-dependent enzymes specialized in handling terminal amines. Biochim. Biophys. Acta 2015, 1854, 1200–1211. [Google Scholar] [CrossRef] [PubMed]
- Storici, P.; Capitani, G.; Müller, R.; Schirmer, T.; Jansonius, J.N. Crystal structure of human ornithine aminotransferase complexed with the highly specific and potent inhibitor 5-fluoromethylornithine. J. Mol. Biol. 1999, 285, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.A.; Shen, B.W.; Brünger, A.T. Human ornithine aminotransferase complexed with L-canaline and gabaculine: Structural basis for substrate recognition. Struct. Lond. Engl. 1997, 5, 1067–1075. [Google Scholar] [CrossRef]
- Markova, M.; Peneff, C.; Hewlins, M.J.E.; Schirmer, T.; John, R.A. Determinants of substrate specificity in omega-aminotransferases. J. Biol. Chem. 2005, 280, 36409–36416. [Google Scholar] [CrossRef] [PubMed]
- Marković-Housley, Z.; Kania, M.; Lustig, A.; Vincent, M.G.; Jansonius, J.N.; John, R.A. Quaternary structure of ornithine aminotransferase in solution and preliminary crystallographic data. Eur. J. Biochem. 1987, 162, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Lünsdorf, H.; Hecht, H.J.; Tsai, H. High-resolution electron microscopic studies on the quaternary structure of ornithine aminotransferase from pig kidney. Eur. J. Biochem. 1994, 225, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.N.; Rho, H.W.; Park, J.W.; Jhee, E.C.; Kim, J.S.; Kim, H.R. A variant of ornithine aminotransferase from mouse small intestine. Exp. Mol. Med. 1998, 30, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Brody, L.C.; Mitchell, G.A.; Obie, C.; Michaud, J.; Steel, G.; Fontaine, G.; Robert, M.F.; Sipila, I.; Kaiser-Kupfer, M.; Valle, D. Ornithine delta-aminotransferase mutations in gyrate atrophy. Allelic heterogeneity and functional consequences. J. Biol. Chem. 1992, 267, 3302–3307. [Google Scholar] [PubMed]
- Michaud, J.; Brody, L.C.; Steel, G.; Fontaine, G.; Martin, L.S.; Valle, D.; Mitchell, G. Strand-separating conformational polymorphism analysis: Efficacy of detection of point mutations in the human ornithine delta-aminotransferase gene. Genomics 1992, 13, 389–394. [Google Scholar] [CrossRef]
- Malashkevich, V.N.; Toney, M.D.; Jansonius, J.N. Crystal structures of true enzymatic reaction intermediates: Aspartate and glutamate ketimines in aspartate aminotransferase. Biochem. 1993, 32, 13451–13462. [Google Scholar] [CrossRef]
- Kirsch, J.F.; Eichele, G.; Ford, G.C.; Vincent, M.G.; Jansonius, J.N.; Gehring, H.; Christen, P. Mechanism of action of aspartate aminotransferase proposed on the basis of its spatial structure. J. Mol. Biol. 1984, 174, 497–525. [Google Scholar] [CrossRef]
- Barrett, D.J.; Bateman, J.B.; Sparkes, R.S.; Mohandas, T.; Klisak, I.; Inana, G. Chromosomal localization of human ornithine aminotransferase gene sequences to 10q26 and Xp11. 2. Invest. Ophthalmol. Vis. Sci. 1987, 28, 1037–1042. [Google Scholar] [PubMed]
- Ramesh, V.; Eddy, R.; Bruns, G.A.; Shih, V.E.; Shows, T.B.; Gusella, J.F. Localization of the ornithine aminotransferase gene and related sequences on two human chromosomes. Hum. Genet. 1987, 76, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Hotta, Y.; Inana, G. Gene transfer and expression of human ornithine aminotransferase. Invest. Ophthalmol. Vis. Sci. 1989, 30, 1024–1031. [Google Scholar] [PubMed]
- Ramesh, V.; Cheng, S.V.; Kozak, C.A.; Herron, B.J.; Shih, V.E.; Taylor, B.A.; Gusella, J.F. Mapping of ornithine aminotransferase gene sequences to mouse Chromosomes 7, X, and 3. Mamm. Genome 1992, 3, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhang, S. Computational prediction of amphioxus microRNA genes and their targets. Gene 2009, 428, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Selbach, M.; Schwanhäusser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread changes in protein synthesis induced by microRNAs. Nature 2008, 455, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Ladeuix, B.; Duchamp, C.; Levillain, O. Underestimated contribution of skeletal muscle in ornithine metabolism during mouse postnatal development. Amino Acids 2014, 46, 167–176. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, B.; Suijkerbuijk, R.F.; Balemans, M.; Sinke, R.J.; De Jong, B.; Molenaar, W.M.; Meloni, A.M.; Sandberg, A.A.; Geraghty, M.; Hofker, M. Sublocalization of the synovial sarcoma-associated t (X; 18) chromosomal breakpoint in Xp11.2 using cosmid cloning and fluorescence in situ hybridization. Oncogene 1993, 8, 1457–1463. [Google Scholar] [PubMed]
- Shipley, J.M.; Clark, J.; Crew, A.J.; Birdsall, S.; Rocques, P.J.; Gill, S.; Chelly, J.; Monaco, A.P.; Abe, S.; Gusterson, B.A. The t (X; 18) (p11.2; q11.2) translocation found in human synovial sarcomas involves two distinct loci on the X chromosome. Oncogene 1994, 9, 1447–1453. [Google Scholar] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, G.A.; Looney, J.E.; Brody, L.C.; Steel, G.; Suchanek, M.; Engelhardt, J.F.; Willard, H.F.; Valle, D. Human ornithine-delta-aminotransferase. cDNA cloning and analysis of the structural gene. J. Biol. Chem. 1988, 263, 14288–14295. [Google Scholar] [PubMed]
- Fagan, R.J.; Sheffield, W.P.; Rozen, R. Regulation of ornithine aminotransferase in retinoblastomas. J. Biol. Chem. 1989, 264, 20513–20517. [Google Scholar] [PubMed]
- Kimura, T.; Chowdhury, S.; Tanaka, T.; Shimizu, A.; Iwase, K.; Oyadomari, S.; Gotoh, T.; Matsuzaki, H.; Mori, M.; Akira, S.; et al. CCAAT/enhancer-binding protein β is required for activation of genes for ornithine cycle enzymes by glucocorticoids and glucagon in primary-cultured hepatocytes. FEBS Lett. 2001, 494, 105–111. [Google Scholar] [CrossRef]
- Hayashi, H.; Katunuma, N.; Chiku, K.; Endo, Y.; Natori, Y. Cell-free synthesis of ornithine aminotransferase of rat liver. J. Biochem. 1981, 90, 1229–1232. [Google Scholar] [CrossRef] [PubMed]
- Ewing, R.M.; Chu, P.; Elisma, F.; Li, H.; Taylor, P.; Climie, S.; McBroom-Cerajewski, L.; Robinson, M.D.; O’Connor, L.; Li, M.; et al. Large-scale mapping of human protein-protein interactions by mass spectrometry. Mol. Syst. Biol. 2007, 3, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.; Tuboi, S. Translocation of proteins into rat liver mitochondria. Eur. J. Biochem. 1986, 155, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Chen, Y.; Tishkoff, D.X.; Peng, C.; Tan, M.; Dai, L.; Xie, Z.; Zhang, Y.; Zwaans, B.M.M.; Skinner, M.E.; et al. SIRT5-mediated lysine desuccinylation impacts diverse metabolic pathways. Mol. Cell 2013, 50, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Rardin, M.J.; Newman, J.C.; Held, J.M.; Cusack, M.P.; Sorensen, D.J.; Li, B.; Schilling, B.; Mooney, S.D.; Kahn, C.R.; Verdin, E.; et al. Label-free quantitative proteomics of the lysine acetylome in mitochondria identifies substrates of SIRT3 in metabolic pathways. Proc. Natl. Acad. Sci. USA 2013, 110, 6601–6606. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Xu, Z.; Xia, L.; Liu, X.; Tu, Y.; Lei, H.; Wang, W.; Wang, T.; Song, L.; Ma, C.; et al. Important role of SUMOylation of spliceosome factors in prostate cancer cells. J. Proteome Res. 2014, 13, 3571–3582. [Google Scholar] [CrossRef] [PubMed]
- Stark, C.; Breitkreutz, B.-J.; Reguly, T.; Boucher, L.; Breitkreutz, A.; Tyers, M. BioGRID: A general repository for interaction datasets. Nucleic Acids Res. 2006, 34, D535–D539. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.; Wu, K.; Yang, Y.; Guerrero, C.; Nillegoda, N.; Pan, Z.-Q.; Huang, L. A targeted proteomic analysis of the ubiquitin-like modifier nedd8 and associated proteins. J. Proteome Res. 2008, 7, 1274–1287. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, J.M.R.; Sylvestersen, K.B.; Bekker-Jensen, S.; Szklarczyk, D.; Poulsen, J.W.; Horn, H.; Jensen, L.J.; Mailand, N.; Nielsen, M.L. Mass spectrometric analysis of lysine ubiquitylation reveals promiscuity at site level. Mol. Cell. Proteomics 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Bennett, E.J.; Huttlin, E.L.; Guo, A.; Li, J.; Possemato, A.; Sowa, M.E.; Rad, R.; Rush, J.; Comb, M.J.; et al. Systematic and quantitative assessment of the ubiquitin-modified proteome. Mol. Cell 2011, 44, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.A.; Beli, P.; Weinert, B.T.; Nielsen, M.L.; Cox, J.; Mann, M.; Choudhary, C. A proteome-wide, quantitative survey of in vivo ubiquitylation sites reveals widespread regulatory roles. Mol. Cell. Proteomics 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Stes, E.; Laga, M.; Walton, A.; Samyn, N.; Timmerman, E.; De Smet, I.; Goormachtig, S.; Gevaert, K. A COFRADIC protocol to study protein ubiquitination. J. Proteome Res. 2014, 13, 3107–3113. [Google Scholar] [CrossRef] [PubMed]
- Falsone, S.F.; Gesslbauer, B.; Rek, A.; Kungl, A.J. A proteomic approach towards the Hsp90-dependent ubiquitinylated proteome. Proteomics 2007, 7, 2375–2383. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Borgeson, B.; Phanse, S.; Tu, F.; Drew, K.; Clark, G.; Xiong, X.; Kagan, O.; Kwan, J.; Bezginov, A.; et al. Panorama of ancient metazoan macromolecular complexes. Nature 2015, 525, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Emdal, K.B.; Pedersen, A.-K.; Bekker-Jensen, D.B.; Tsafou, K.P.; Horn, H.; Lindner, S.; Schulte, J.H.; Eggert, A.; Jensen, L.J.; Francavilla, C.; et al. Temporal proteomics of NGF-TrkA signaling identifies an inhibitory role for the E3 ligase Cbl-b in neuroblastoma cell differentiation. Sci. Signal. 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Greco, T.M.; Yu, F.; Guise, A.J.; Cristea, I.M. Nuclear import of histone deacetylase 5 by requisite nuclear localization signal phosphorylation. Mol. Cell. Proteomics 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Christianson, J.C.; Olzmann, J.A.; Shaler, T.A.; Sowa, M.E.; Bennett, E.J.; Richter, C.M.; Tyler, R.E.; Greenblatt, E.J.; Harper, J.W.; Kopito, R.R. Defining human ERAD networks through an integrative mapping strategy. Nat. Cell Biol. 2011, 14, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Vinayagam, A.; Stelzl, U.; Foulle, R.; Plassmann, S.; Zenkner, M.; Timm, J.; Assmus, H.E.; Andrade-Navarro, M.A.; Wanker, E.E. A directed protein interaction network for investigating intracellular signal transduction. Sci. Signal. 2011, 4. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-C.; Greco, T.M.; Boonmee, A.; Miteva, Y.; Cristea, I.M. Functional proteomics establishes the interaction of SIRT7 with chromatin remodeling complexes and expands its role in regulation of RNA polymerase I transcription. Mol. Cell. Proteomics 2012, 11, 60–76. [Google Scholar] [CrossRef] [PubMed]
- Richter, R.; Rorbach, J.; Pajak, A.; Smith, P.M.; Wessels, H.J.; Huynen, M.A.; Smeitink, J.A.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M. A functional peptidyl-tRNA hydrolase, ICT1, has been recruited into the human mitochondrial ribosome. EMBO J. 2010, 29, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Stelzl, U.; Worm, U.; Lalowski, M.; Haenig, C.; Brembeck, F.H.; Goehler, H.; Stroedicke, M.; Zenkner, M.; Schoenherr, A.; Koeppen, S.; et al. A human protein-protein interaction network: a resource for annotating the proteome. Cell 2005, 122, 957–968. [Google Scholar] [CrossRef] [PubMed]
- Sowa, M.E.; Bennett, E.J.; Gygi, S.P.; Harper, J.W. Defining the human deubiquitinating enzyme interaction landscape. Cell 2009, 138, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Neganova, I.; Vilella, F.; Atkinson, S.P.; Lloret, M.; Passos, J.F.; Von Zglinicki, T.; O’Connor, J.-E.; Burks, D.; Jones, R.; Armstrong, L.; et al. An important role for CDK2 in G1 to S checkpoint activation and DNA damage response in human embryonic stem cells. Stem Cells Dayt. Ohio 2011, 29, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Hao, T.; Shaw, C.; Patel, A.J.; Szabó, G.; Rual, J.-F.; Fisk, C.J.; Li, N.; Smolyar, A.; Hill, D.E.; et al. A protein-protein interaction network for human inherited ataxias and disorders of Purkinje cell degeneration. Cell 2006, 125, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.W.; Nagel, J.; Hoving, S.; Gerrits, B.; Bauer, A.; Thomas, J.R.; Kirschner, M.W.; Schirle, M.; Luchansky, S.J. Quantitative Lys-ϵ-Gly-Gly (diGly) proteomics coupled with inducible RNAi reveals ubiquitin-mediated proteolysis of DNA damage-inducible transcript 4 (DDIT4) by the E3 ligase HUWE1. J. Biol. Chem. 2014, 289, 28942–28955. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jiang, X.; Chen, G.; Xu, J. Interaction of amyotrophic lateral sclerosis/frontotemporal lobar degeneration-associated fused-in-sarcoma with proteins involved in metabolic and protein degradation pathways. Neurobiol. Aging 2015, 36, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Taylor, P.; Moran, M.F. Proteomic analysis of the epidermal growth factor receptor (EGFR) interactome and post-translational modifications associated with receptor endocytosis in response to EGF and stress. Mol. Cell. Proteomics 2014, 13, 1644–1658. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.J.; Glazkova, I.; Fréchette, L.; Iorio-Morin, C.; Binda, C.; Pétrin, D.; Trieu, P.; Robitaille, M.; Angers, S.; Hébert, T.E.; et al. Novel, gel-free proteomics approach identifies RNF5 and JAMP as modulators of GPCR stability. Mol. Endocrinol. 2013, 27, 1245–1266. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, Y.; Wang, T.-D.; Xu, H.-Z.; Xia, L.; Zhang, J.; Wu, Y.-L.; Chen, G.-Q.; Wang, L.-S. Proteomic identification of common SCF ubiquitin ligase FBXO6-interacting glycoproteins in three kinds of cells. J. Proteome Res. 2012, 11, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Havugimana, P.C.; Hart, G.T.; Nepusz, T.; Yang, H.; Turinsky, A.L.; Li, Z.; Wang, P.I.; Boutz, D.R.; Fong, V.; Phanse, S.; et al. A census of human soluble protein complexes. Cell 2012, 150, 1068–1081. [Google Scholar] [CrossRef] [PubMed]
- Lau, E.; Kluger, H.; Varsano, T.; Lee, K.; Scheffler, I.; Rimm, D.L.; Ideker, T.; Ronai, Z.A. PKCε promotes oncogenic functions of ATF2 in the nucleus while blocking its apoptotic function at mitochondria. Cell 2012, 148, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.; Zhang, R.; Verdoodt, B.; Bailey, A.; Zhang, C.-D.; Yates, J.R.; Menssen, A.; Hermeking, H. Large-scale identification of c-MYC-associated proteins using a combined TAP/MudPIT approach. Cell Cycle 2007, 6, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Dillon, C.P.; Shi, L.Z.; Milasta, S.; Carter, R.; Finkelstein, D.; McCormick, L.L.; Fitzgerald, P.; Chi, H.; Munger, J.; et al. The transcription factor myc controls metabolic reprogramming upon T lymphocyte activation. Immunity 2011, 35, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Chee, P.Y.; Swick, R.W. Effect of dietary protein and tryptophan and the turnover of rat liver ornithine aminotransferase. J. Biol. Chem. 1976, 251, 1029–1034. [Google Scholar] [PubMed]
- Ip, M.M.; Chee, P.Y.; Swick, R.W. Turnover of hepatic mitochondril ornithine aminotransferase and cytochrome oxidase using [14C] carbonate as tracer. Biochim. Biophys. Acta 1974, 354, 29–38. [Google Scholar] [CrossRef]
- Kobayashi, K.; Morris, H.P.; Katunuma, N. Studies on the turnover rates of ornithine aminotransferase in morris hepatoma 44 and host liver. J. Biochem. 1976, 80, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Kominami, E.; Katunuma, N. Studies on new intracellular proteases in various organs of rats. Eur. J. Biochem. 1976, 62, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Weiss, R.L. Intracellular localization of ornithine and arginine pools in neurospora. J. Biol. Chem. 1973, 248, 5409–5413. [Google Scholar] [PubMed]
- Jauniaux, J.C.; Urrestarazu, L.A.; Wiame, J.M. Arginine metabolism in Saccharomyces cerevisiae: subcellular localization of the enzymes. J. Bacteriol. 1978, 133, 1096–1107. [Google Scholar] [PubMed]
- Stránská, J.; Kopecný, D.; Tylichová, M.; Snégaroff, J.; Sebela, M. Ornithine delta-aminotransferase: An enzyme implicated in salt tolerance in higher plants. Plant Signal. Behav. 2008, 3, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Roosens, N.H.; Thu, T.T.; Iskandar, H.M.; Jacobs, M. Isolation of the ornithine-delta-aminotransferase cDNA and effect of salt stress on its expression in Arabidopsis thaliana. Plant Physiol. 1998, 117, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Shang, L.; Burgett, A.W.G.; Harran, P.G.; Wang, X. Diazonamide toxins reveal an unexpected function for ornithine δ-amino transferase in mitotic cell division. Proc. Natl. Acad. Sci. 2007, 104, 2068–2073. [Google Scholar] [CrossRef] [PubMed]
- Furic, L.; Maher-Laporte, M.; DesGroseillers, L. A genome-wide approach identifies distinct but overlapping subsets of cellular mRNAs associated with Staufen1- and Staufen2-containing ribonucleoprotein complexes. RNA 2008, 14, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, S.L.; Lautenberger, J.A.; Chinn, L.W.; Malasky, M.; Sezgin, E.; Kingsley, L.A.; Goedert, J.J.; Kirk, G.D.; Gomperts, E.D.; Buchbinder, S.P.; et al. Genetic variants in nuclear-encoded mitochondrial genes influence AIDS progression. PLoS ONE 2010, 5, e12862. [Google Scholar] [CrossRef] [PubMed]
- Antibody Staining Consistent with RNA Expression Data. Available online: http://www.proteinatlas.org/ENSG00000065154-OAT/ (accessed on 5 September 2016).
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347. [Google Scholar] [CrossRef] [PubMed]
- Gene: OAT-ENSG00000065154-Homo Sapiens (Human). Available online: http://bgee.org/?page=gene&gene_id=ENSG00000065154 (accessed on 5 September 2016).
- Bastian, F.; Parmentier, G.; Roux, J.; Moretti, S.; Laudet, V.; Robinson-Rechavi, M. Bgee: Integrating and Comparing Heterogeneous Transcriptome Data among Species. In Data Integration in the Life Sciences; Bairoch, A., Cohen-Boulakia, S., Froidevaux, C., Eds.; Springer: Heidelberg, Germany, 2008; pp. 124–131. [Google Scholar]
- Affymetrix Human Genome U133 Plus 2.0 Array. Available online: https://genevisible.com/tissues/HS/UniProt/P04181 (accessed on 3 September 2016).
- Hruz, T.; Laule, O.; Szabo, G.; Wessendorp, F.; Bleuler, S.; Oertle, L.; Widmayer, P.; Gruissem, W.; Zimmermann, P. Genevestigator V3: A reference expression database for the meta-analysis of transcriptomes. Adv. Bioinforma. 2008, 2008, e420747. [Google Scholar] [CrossRef] [PubMed]
- Hayasaka, S.; Shiono, T.; Takaku, Y.; Mizuno, K. Ornithine ketoacid aminotransferase in the bovine eye. Invest. Ophthalmol. Vis. Sci. 1980, 19, 1457–1460. [Google Scholar] [PubMed]
- Ratzlaff, K.; Baich, A. Comparison of ornithine aminotransferase activities in the pigment epithelium and retina of vertebrates. Comp. Biochem. Physiol. B 1987, 88, 35–37. [Google Scholar] [CrossRef]
- Baich, A. Alternative pathways for proline synthesis in mammalian cells. Somatic Cell Genet. 1977, 3, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, T.; Obara, Y. Amino acid synthesis from ornithine: enzymes and quantitative comparison in brain slices and detached retinas from rats and chicks. Brain Res. 1987, 413, 314–319. [Google Scholar] [CrossRef]
- Berglund, L.; Björling, E.; Jonasson, K.; Rockberg, J.; Fagerberg, L.; Al-Khalili Szigyarto, C.; Sivertsson, A.; Uhlén, M. A whole-genome bioinformatics approach to selection of antigens for systematic antibody generation. Proteomics 2008, 8, 2832–2839. [Google Scholar] [CrossRef] [PubMed]
- Herzfeld, A.; Knox, W.E. The Properties, developmental formation, and estrogen induction of ornithine aminotransferase in rat tissues. J. Biol. Chem. 1968, 243, 3327–3332. [Google Scholar] [PubMed]
- Katunuma, N.; Matsuda, Y.; Tomino, I. Studies on ornithine-ketoacid transaminase: I. purification and properties. J. Biochem. 1964, 56, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Katunuma, N.; Okada, M.; Matsuzawa, T.; Otsuka, Y. Studies on ornithine ketoacid transaminase II. role in metabolic pathway. J. Biochem. 1965, 57, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Merrill, M.J.; Pitot, H.C. Regulation of ornithine aminotransferase by cyclic AMP and glucose in primary cultures of adult rat hepatocytes. Arch. Biochem. Biophys. 1985, 237, 373–385. [Google Scholar] [CrossRef]
- Brosnan, M.E.; Brosnan, J.T. Hepatic glutamate metabolism: A tale of 2 hepatocytes. Am. J. Clin. Nutr. 2009, 90, 857S–861S. [Google Scholar] [CrossRef] [PubMed]
- Kuo, F.C.; Hwu, W.L.; Valle, D.; Darnell, J.E. Colocalization in pericentral hepatocytes in adult mice and similarity in developmental expression pattern of ornithine aminotransferase and glutamine synthetase mRNA. Proc. Natl. Acad. Sci. USA 1991, 88, 9468–9472. [Google Scholar] [CrossRef] [PubMed]
- Colombatto, S.; Giribaldi, G.; Vargiu, C.; Grillo, M.A. Modulation of ornithine aminotransferase activity by oxygen in rat hepatocyte cultures. Biochim. Biophys. Acta 1994, 1224, 329–332. [Google Scholar] [CrossRef]
- Swick, R.W.; Tollaksen, S.L.; Nance, S.L.; Thomson, J.F. The unique distribution of ornithine aminotransferase in rat liver mitochondria. Arch. Biochem. Biophys. 1970, 136, 212–218. [Google Scholar] [CrossRef]
- Boon, L.; Geerts, W.J.; Jonker, A.; Lamers, W.H.; Van Noorden, C.J. High protein diet induces pericentral glutamate dehydrogenase and ornithine aminotransferase to provide sufficient glutamate for pericentral detoxification of ammonia in rat liver lobules. Histochem. Cell Biol. 1999, 111, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Pitot, H.C.; Peraino, C. Carbohydrate repression of enzyme induction in rat liver. J. Biol. Chem. 1963, 238, PC1910–PC1912. [Google Scholar]
- Peraino, C. Interactions of diet and cortisone in the regulation of adaptive enzymes in rat liver. J. Biol. Chem. 1967, 242, 3860–3867. [Google Scholar] [PubMed]
- Peraino, C.; Pitot, H.C. Studies on the induction and repression of enzymes in rat liver: II. Carbohydrate repression of dietary and hormonal induction of threonine dehydrase and ornithine δ-transaminase. J. Biol. Chem. 1964, 239, 4308–4313. [Google Scholar] [PubMed]
- Lyons, R.T.; Pitot, H.C. The regulation of ornithine aminotransferase synthesis by glucagon in the rat. Arch. Biochem. Biophys. 1976, 174, 262–272. [Google Scholar] [CrossRef]
- Nakajima, K.; Ishikawa, E. Hormonal and dietary control of enzymes in the rat: III. The regulation of liver serine dehydratase and ornithine δ-aminotransferase in hypophysectomized-pancreatectomized R. J. Biochem. 1971, 69, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Merrill, M.J.; Pitot, H.C. Inhibition of cyclic AMP-dependent induction of ornithine aminotransferase by simple carbohydrates in cultured hepatocytes. Arch. Biochem. Biophys. 1987, 259, 250–261. [Google Scholar] [CrossRef]
- Mueckler, M.M.; Merrill, M.J.; Pitot, H.C. Translational and pretranslational control of ornithine aminotransferase synthesis in rat liver. J. Biol. Chem. 1983, 258, 6109–6114. [Google Scholar] [PubMed]
- Shull, J.D.; Esumi, N.; Colwell, A.S.; Pennington, K.L.; Jendoubi, M. Sequence of the promoter region of the mouse gene encoding ornithine aminotransferase. Gene 1995, 162, 275–277. [Google Scholar] [CrossRef]
- Peraino, C. Regulatory effects of gluococorticoids on ornithine amino-transferase and serine dehydratase in rat liver. Biochim. Biophys. Acta 1968, 165, 108–112. [Google Scholar] [CrossRef]
- Peraino, C.; Emory Morris, J.; Shenoy, S.T. Evidence for different mechanisms in the circadian and glucocorticoid control of rat liver ornithine aminotransferase synthesis. Life Sci. 1976, 19, 1435–1437. [Google Scholar] [CrossRef]
- Ventura, G.; Moinard, C.; Segaud, F.; Le Plenier, S.; Cynober, L.; De Bandt, J.P. Adaptative response of nitrogen metabolism in early endotoxemia: Role of ornithine aminotransferase. Amino Acids 2010, 39, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.E.; Peraino, C. Immunochemical studies of serine dehydratase and ornithine aminotransferase regulation in rat liver in vivo. J. Biol. Chem. 1976, 251, 2571–2578. [Google Scholar] [PubMed]
- Ekelman, K.B.; Peraino, C. Effects of feeding and lighting stimuli on the synthesis of ornithine aminotransferase and serine dehydratase in rat liver. Arch. Biochem. Biophys. 1981, 209, 677–681. [Google Scholar] [CrossRef]
- Yuan, Y.; Hilliard, G.; Ferguson, T.; Millhorn, D.E. Cobalt Inhibits the Interaction between hypoxia-inducible factor- and von hippel-lindau protein by direct binding to hypoxia-inducible factor-α. J. Biol. Chem. 2003, 278, 15911–15916. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, C.; Rafii, M.; Ball, R.O.; Pencharz, P.B. Arginine can be synthesized from enteral proline in healthy adult humans. J. Nutr. 2011, 141, 1432–1436. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, T.; Kobayashi, T.; Tashiro, K.; Kasahara, M. Changes in ornithine metabolic enzymes induced by dietary protein in small intestine and liver: Intestine-liver relationship in ornithine supply to liver. J. Biochem. 1994, 116, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Dekaney, C.M.; Wu, G.; Jaeger, L.A. Ornithine aminotransferase messenger RNA expression and enzymatic activity in fetal porcine intestine. Pediatr. Res. 2001, 50, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Levillain, O.; Diaz, J.-J.; Reymond, I.; Soulet, D. Ornithine metabolism along the female mouse nephron: Localization of ornithine decarboxylase and ornithine aminotransferase. Pflügers Arch. 2000, 440, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M.M.; Moran, S.; Pitot, H.C. Transcriptional control of ornithine aminotransferase synthesis in rat kidney by estrogen and thyroid hormone. J. Biol. Chem. 1984, 259, 2302–2305. [Google Scholar] [PubMed]
- Lyons, R.T.; Pitot, H.C. Hormonal regulation of ornithine aminotransferase biosynthesis in rat liver and kidney. Arch. Biochem. Biophys. 1977, 180, 472–479. [Google Scholar] [CrossRef]
- Wu, C. Estrogen receptor translocation and ornithine aminotransferase induction by estradiol in rat kidney. Biochem. Biophys. Res. Commun. 1979, 89, 769–776. [Google Scholar] [CrossRef]
- Levillain, O.; Diaz, J.-J.; Blanchard, O.; Déchaud, H. Testosterone down-regulates ornithine aminotransferase gene and up-regulates arginase II and ornithine decarboxylase genes for polyamines synthesis in the murine kidney. Endocrinology 2005, 146, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Fagan, R.J.; Rozen, R. Translational control of ornithine-delta-aminotransferase (OAT) by estrogen. Mol. Cell. Endocrinol. 1993, 90, 171–177. [Google Scholar] [CrossRef]
- Kasahara, M.; Matsuzawa, T.; Kokubo, M.; Gushiken, Y.; Tashiro, K.; Koide, T.; Watanabe, H.; Katunuma, N. Immunohistochemical localization of ornithine aminotransferase in normal rat tissues by Fab’-horseradish peroxidase conjugates. J. Histochem. Cytochem. 1986, 34, 1385–1388. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, J.T.; Blaker, W.D.; Meek, J.L. Ornithine as a precursor of neurotransmitter glutamate: Effect of canaline on ornithine aminotransferase activity and glutamate content in the septum of rat brain. Brain Res. 1985, 329, 161–168. [Google Scholar] [CrossRef]
- Lapinjoki, S.P.; Pajunen, A.E.I.; Pulkka, A.E.; Piha, R.S. On the metabolism of ornithine in synaptosomal preparations. Neurochem. Res. 1982, 7, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Seiler, N.; Knödgen, B. Transformation of glutamic acid, putrescine and ornithine into gamma-aminobutyric acid in brain. Hoppe Seylers Z. Physiol. Chem. 1971, 352, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Seiler, N. On the role of GABA in vertebrate polyamine metabolism. Physiol. Chem. Phys. 1980, 12, 411–429. [Google Scholar] [PubMed]
- Wong, P.T.-H.; McGeer, E.G.; McGeer, P.L. A sensitive radiometric assay for ornithine aminotransferase: Regional and subcellular distributions in rat brain. J. Neurochem. 1981, 36, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.T.-H.; McGeer, P.L.; Rossor, M.; McGeer, E.G. Ornithine aminotransferase in Huntington’s disease. Brain Res. 1982, 231, 466–471. [Google Scholar] [CrossRef]
- Wong, P.T.; McGeer, E.G.; McGeer, P.L. Effects of kainic acid injection and cortical lesion on ornithine and aspartate aminotransferases in rat striatum. J. Neurosci. Res. 1982, 8, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Shiono, T.; Hayasaka, S.; Mizuno, K. Development of ornithine ketoacid aminotransferase in rabbit ocular tissues and liver. Invest. Ophthalmol. Vis. Sci. 1982, 23, 419–424. [Google Scholar] [PubMed]
- Shiono, T.; Hayasaka, S.; Mizuno, K. Presence of ornithine ketoacid aminotransferase in human ocular tissues. Graefes Arch. Clin. Exp. Ophthalmol. 1982, 218, 34–36. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Kazarinoff, M.N.; Seiler, N.; Stanley, B.A. Rat colon ornithine and arginine metabolism: Coordinated effects after proliferative stimuli. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G389–G399. [Google Scholar] [PubMed]
- Durante, W.; Liao, L.; Reyna, S.V.; Peyton, K.J.; Schafer, A.I. Transforming growth factor-beta (1) stimulates L-arginine transport and metabolism in vascular smooth muscle cells: Role in polyamine and collagen synthesis. Circulation 2001, 103, 1121–1127. [Google Scholar] [CrossRef] [PubMed]
- Ladeuix, B. Contributions de l’ornithine aminotransférase dans les réponses physiologiques aux contraintes métaboliques chez la souris. Ph.D. Thesis, University Claude Bernard, Villeurbanne, France, 2013. [Google Scholar]
- Hampel, A.; Huber, C.; Geffers, R.; Spona-Friedl, M.; Eisenreich, W.; Bange, F.-C. Mycobacterium tuberculosis is a natural ornithine aminotransferase (rocD) mutant and depends on Rv2323c for growth on arginine. PLoS ONE 2015, 10, e0136914. [Google Scholar] [CrossRef] [PubMed]
- Hervieu, F.; Le Dily, F.; Huault, C.; Billard, J.-P. Contribution of ornithine aminotransferase to proline accumulation in NaCl-treated radish cotyledons. Plant Cell Environ. 1995, 18, 205–210. [Google Scholar] [CrossRef]
- You, J.; Hu, H.; Xiong, L. An ornithine δ-aminotransferase gene OsOAT confers drought and oxidative stress tolerance in rice. Plant Sci. 2012, 197, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Fincham, J.R.S. Ornithine transaminase in neurospora and its relation to the biosynthesis of proline. Biochem. J. 1953, 53, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Dzikowska, A.; Swianiewicz, M.; Talarczyk, A.; Wisniewska, M.; Goras, M.; Scazzocchio, C.; Weglenski, P. Cloning, characterisation and regulation of the ornithine transaminase (otaA) gene of Aspergillus nidulans. Curr. Genet. 1999, 35, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Hellebust, J.A.; Larochelle, J. Compartmentation and activities of enzymes Involved in the metabolism of amino acids implicated in osmoregulatory mechanisms in acanthamoeba castellanii1. J. Protozool. 1988, 35, 498–502. [Google Scholar] [CrossRef]
- Jortzik, E.; Fritz-Wolf, K.; Sturm, N.; Hipp, M.; Rahlfs, S.; Becker, K. Redox regulation of plasmodium falciparum ornithine δ-aminotransferase. J. Mol. Biol. 2010, 402, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Herzfeld, A.; Raper, S.M. Enzymes of ornithine metabolism in adult and developing rat intestine. Biochim. Biophys. Acta 1976, 428, 600–610. [Google Scholar] [CrossRef]
- Jones, M.E. Conversion of glutamate to ornithine and proline: Pyrroline-5-carboxylate, a possible modulator of arginine requirements. J. Nutr. 1985, 115, 509–515. [Google Scholar] [PubMed]
- Smith, R.J.; Phang, J.M. The importance of ornithine as a precursor for proline in mammalian cells. J. Cell. Physiol. 1979, 98, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Sipilä, I.; Simell, O.; O’Donnell, J.J. Gyrate atrophy of the choroid and retina with hyperornithinemia: Characterization of mutant liver L-ornithine: 2-oxoacid aminotransferase kinetics. J. Clin. Invest. 1981, 67, 1805–1807. [Google Scholar] [CrossRef] [PubMed]
- Valle, D.L.; Boison, A.P.; Jezyk, P.; Aguirre, G. Gyrate atrophy of the choroid and retina in a cat. Invest. Ophthalmol. Vis. Sci. 1981, 20, 251–255. [Google Scholar] [PubMed]
- Simell, O.; Takki, K. Raised plasma-ornithine and gyrate atrophy of the choroid and retina. Lancet Lond. Engl. 1973, 1, 1031–1033. [Google Scholar] [CrossRef]
- Ventura, G.; De Bandt, J.-P.; Segaud, F.; Perret, C.; Robic, D.; Levillain, O.; Le Plenier, S.; Godard, C.; Cynober, L.; Moinard, C. Overexpression of ornithine aminotransferase: Consequences on amino acid homeostasis. Br. J. Nutr. 2009, 101, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Marini, J.C.; Stoll, B.; Didelija, I.C.; Burrin, D.G. De novo synthesis is the main source of ornithine for citrulline production in neonatal pigs. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E1348–E1353. [Google Scholar] [CrossRef] [PubMed]
- Ross, G.; Dunn, D.; Jones, M.E. Ornithine synthesis from glutamate in rat intestinal mucosa homogenates: Evidence for the reduction of glutamate to gamma-glutamyl semialdehyde. Biochem. Biophys. Res. Commun. 1978, 85, 140–147. [Google Scholar] [CrossRef]
- Henslee, J.G.; Jones, M.E. Ornithine synthesis from glutamate in rat small intestinal mucosa. Arch. Biochem. Biophys. 1982, 219, 186–197. [Google Scholar] [CrossRef]
- Alonso, E.; Rubio, V. Participation of ornithine aminotransferase in the synthesis and catabolism of ornithine in mice. Studies using gabaculine and arginine deprivation. Biochem. J. 1989, 259, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Curis, E.; Nicolis, I.; Moinard, C.; Osowska, S.; Zerrouk, N.; Benazeth, S.; Cynober, L. Almost all about citrulline in mammals. Amino Acids 2005. [Google Scholar] [CrossRef] [PubMed]
- Cynober, L.; Moinard, C.; De Bandt, J.-P. The 2009 ESPEN Sir David Cuthbertson. Citrulline: A new major signaling molecule or just another player in the pharmaconutrition game? Clin. Nutr. 2010, 29, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Bensaci, J.; Curis, E.; Nicolis, I.; De Bandt, J.-P.; Bénazeth, S. An in silico model of enterocytic glutamine to citrulline conversion pathway. Amino Acids 2012, 43, 1727–1737. [Google Scholar] [CrossRef] [PubMed]
- Flynn, N.E.; Wu, G. An important role for endogenous synthesis of arginine in maintaining arginine homeostasis in neonatal pigs. Am. J. Physiol. 1996, 271, R1149–R1155. [Google Scholar] [PubMed]
- Wang, T.; Lawler, A.M.; Steel, G.; Sipila, I.; Milam, A.H.; Valle, D. Mice lacking ornithine-delta-aminotransferase have paradoxical neonatal hypoornithinaemia and retinal degeneration. Nat. Genet. 1995, 11, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, C.; Rafii, M.; Sgro, M.; Ball, R.O.; Pencharz, P. Arginine is synthesized from proline, not glutamate, in enterally fed human preterm neonates. Pediatr. Res. 2011, 69, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Köhler, E.S.; Sankaranarayanan, S.; Van Ginneken, C.J.; Van Dijk, P.; Vermeulen, J.L.M.; Ruijter, J.M.; Lamers, W.H.; Bruder, E. The human neonatal small intestine has the potential for arginine synthesis; developmental changes in the expression of arginine-synthesizing and -catabolizing enzymes. BMC Dev. Biol. 2008, 8, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Knabe, D.A.; Kim, S.W. Arginine nutrition in neonatal pigs. J. Nutr. 2004, 134, 2783S–2790S. [Google Scholar] [PubMed]
- Wu, G.; Knabe, D.A. Arginine synthesis in enterocytes of neonatal pigs. Am. J. Physiol. 1995, 269, R621–R629. [Google Scholar] [PubMed]
- Wu, G. Synthesis of citrulline and arginine from proline in enterocytes of postnatal pigs. Am. J. Physiol. 1997, 272, G1382–1390. [Google Scholar] [PubMed]
- De Jonge, W.J.; Dingemanse, M.A.; De Boer, P.A.; Lamers, W.H.; Moorman, A.F. Arginine-metabolizing enzymes in the developing rat small intestine. Pediatr. Res. 1998, 43, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Shull, J.D.; Pennington, K.L.; Rader, A.E. Promoter region of the rat gene encoding ornithine aminotransferase: Transcriptional activity, sequence, and DNase-I-hypersensitive sites. Gene 1993, 125, 169–175. [Google Scholar] [CrossRef]
- Ozaki, M.; Gotoh, T.; Nagasaki, A.; Miyanaka, K.; Takeya, M.; Fujiyama, S.; Tomita, K.; Mori, M. Expression of arginase II and related enzymes in the rat small intestine and kidney. J. Biochem. 1999, 125, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Borbolla, A.G.; Knabe, D.A. The uptake of glutamine and release of arginine, citrulline and proline by the small intestine of developing pigs. J. Nutr. 1994, 124, 2437–2444. [Google Scholar] [PubMed]
- Dekaney, C.M.; Wu, G.; Yin, Y.-L.; Jaeger, L.A. Regulation of ornithine aminotransferase gene expression and activity by all-transretinoic acid in Caco-2 intestinal epithelial cells. J. Nutr. Biochem. 2008, 19, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Riby, J.E.; Hurwitz, R.E.; Kretchmer, N. Development of ornithine metabolism in the mouse intestine. Pediatr. Res. 1990, 28, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Cooper, S.K.; Li, Y.; Mei, J.M.; Qiu, S.; Borchert, G.L.; Donald, S.P.; Kung, H.-F.; Phang, J.M. Ornithine-δ-aminotransferase inhibits neurogenesis during xenopus embryonic development. Invest. Ophthalmol. Vis. Sci. 2015, 56, 2486–2497. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, S.; Gekka, T.; Hayashi, T.; Ida, H.; Ohashi, T.; Eto, Y.; Tsuneoka, H. OAT mutations and clinical features in two Japanese brothers with gyrate atrophy of the choroid and retina. Doc. Ophthalmol. 2014, 128, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Heinänen, K.; Näntö-Salonen, K.; Leino, L.; Pulkki, K.; Heinonen, O.; Valle, D.; Simell, O. Gyrate atrophy of the choroid and retina: Lymphocyte ornithine-delta-aminotransferase activity in different mutations and carriers. Pediatr. Res. 1998, 44, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Kaiser-Kupfer, M.I.; Ludwig, I.H.; De Monasterio, F.M.; Valle, D.; Krieger, I. Gyrate atrophy of the choroid and retina: Early findings. Ophthalmology 1985, 92, 394–401. [Google Scholar] [CrossRef]
- Hasanoğlu, A.; Biberoğlu, G.; Tümer, L. Gyrate atrophy of the choroid and retina. Turk. J. Pediatr. 1996, 38, 253–256. [Google Scholar] [PubMed]
- Khan, M.Y.; Ibraheim, A.S.; Firoozmand, S. Gyrate atrophy of the choroid and retina with hyperornithinaemia, cystinuria and lysinuria. Eye 1994, 8, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Tanzer, F.; Firat, M.; Alagoz, M.; Erdogan, H. Gyrate atrophy of the choroid and retina with hyperornithinemia, cystinuria and lysinuria responsive to vitamin B6. BMJ Case Rep. 2011. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, S.; Ueda-Yamada, M.; Ando, A.; Matsumura, S.; Okuda-Ashitaka, E.; Matsumura, M.; Uyama, M.; Ito, S. Cytotoxic effect of spermine on retinal pigment epithelial cells. Invest. Ophthalmol. Vis. Sci. 2007, 48, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Hayasaka, S.; Saito, T.; Nakajima, H.; Takahashi, O.; Mizuno, K.; Tada, K. Clinical trials of vitamin B6 and proline supplementation for gyrate atrophy of the choroid and retina. Br. J. Ophthalmol. 1985, 69, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Sipilä, I. Inhibition of arginine-glycine amidinotransferase by ornithine. A possible mechanism for the muscular and chorioretinal atrophies in gyrate atrophy of the choroid and retina with hyperornithinemia. Biochim. Biophys. Acta 1980, 613, 79–84. [Google Scholar] [CrossRef]
- Stöckler, S.; Hanefeld, F.; Frahm, J. Creatine replacement therapy in guanidinoacetate methyltransferase deficiency, a novel inborn error of metabolism. Lancet Lond. Engl. 1996, 348, 789–790. [Google Scholar] [CrossRef]
- Valayannopoulos, V.; Boddaert, N.; Mention, K.; Touati, G.; Barbier, V.; Chabli, A.; Sedel, F.; Kaplan, J.; Dufier, J.L.; Seidenwurm, D.; et al. Secondary creatine deficiency in ornithine delta-aminotransferase deficiency. Mol. Genet. Metab. 2009, 97, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Cleary, M.A.; Dorland, L.; De Koning, T.J.; Poll-The, B.T.; Duran, M.; Mandell, R.; Shih, V.E.; Berger, R.; Olpin, S.E.; Besley, G.T.N. Ornithine aminotransferase deficiency: Diagnostic difficulties in neonatal presentation. J. Inherit. Metab. Dis. 2005, 28, 673–679. [Google Scholar] [CrossRef] [PubMed]
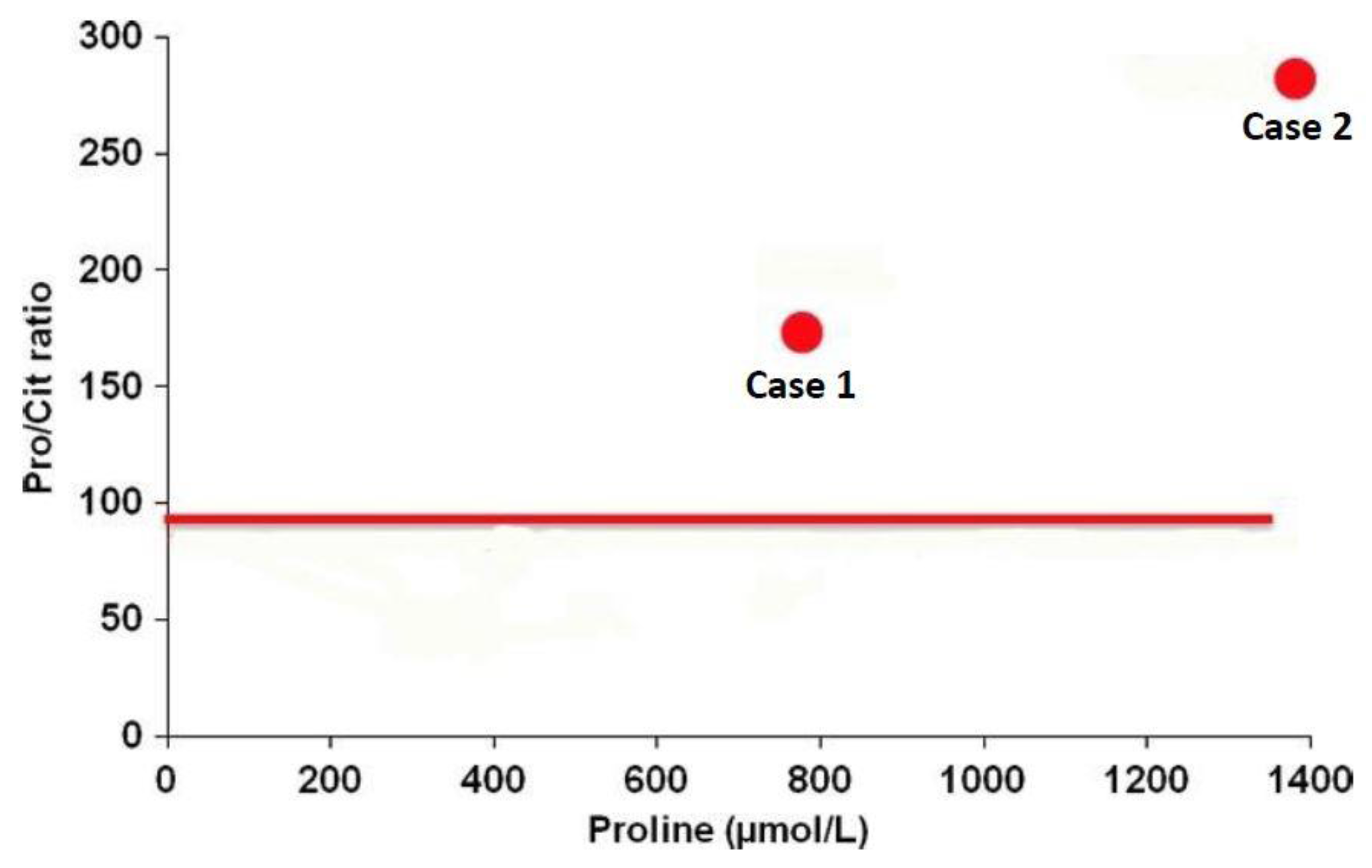
- De Sain-Van Der Velden, M.G.M.; Rinaldo, P.; Elvers, B.; Henderson, M.; Walter, J.H.; Prinsen, B.H.C.M.T.; Verhoeven-Duif, N.M.; De Koning, T.J.; Van Hasselt, P. The proline/citrulline ratio as a biomarker for oat deficiency in early infancy. JIMD Rep. 2012, 6, 95–99. [Google Scholar] [PubMed]
- Kaiser-Kupfer, M.I.; Caruso, R.C.; Valle, D.; Reed, G.F. Use of an arginine-restricted diet to slow progression of visual loss in patients with gyrate atrophy. Arch. Ophthalmol. 2004, 122, 982–984. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, Y.; Ueta, A.; Ito, T.; Sumi, S.; Yamada, M.; Ozawa, K.; Togari, H. Vitamin B6-responsive ornithine aminotransferase deficiency with a novel mutation G237D. Tohoku J. Exp. Med. 2005, 205, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Doimo, M.; Desbats, M.A.; Baldoin, M.C.; Lenzini, E.; Basso, G.; Murphy, E.; Graziano, C.; Seri, M.; Burlina, A.; Sartori, G.; et al. Functional analysis of missense mutations of OAT, causing gyrate atrophy of choroid and retina. Hum. Mutat. 2013, 34, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Nänto-Salonen, K.; Komu, M.; Lundbom, N.; Heinänen, K.; Alanen, A.; Sipilä, I.; Simell, O. Reduced brain creatine in gyrate atrophy of the choroid and retina with hyperornithinemia. Neurology 1999, 53, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Miyasaka, Y.; Enomoto, N.; Nagayama, K.; Izumi, N.; Marumo, F.; Watanabe, M.; Sato, C. Analysis of differentially expressed genes in human hepatocellular carcinoma using suppression subtractive hybridization. Br. J. Cancer 2001, 85, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Zigmond, E.; Ben Ya’acov, A.; Lee, H.; Lichtenstein, Y.; Shalev, Z.; Smith, Y.; Zolotarov, L.; Ziv, E.; Kalman, R.; Le, H.V.; et al. Suppression of hepatocellular carcinoma by inhibition of overexpressed ornithine aminotransferase. ACS Med. Chem. Lett. 2015, 6, 840–844. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Tissue | Species | Optimal pH | Michaelis constant (Km) | Activity | Reference | ||
|---|---|---|---|---|---|---|---|---|
| Orn | aKG | Value | Unit | |||||
| Eye | Retina | Human | 7.6–8.0 | 3.7 | 1.72 | 218 ± 22 | nmol P5C/mg protein/h | [15] |
| Bovine | 5.6 | 1.3 | [16] | |||||
| Calf | 234 ± 26 | nmol P5C/mg protein/h | [15] | |||||
| Rat | 324 ± 43 | nmol P5C/mg protein/h | [15] | |||||
| Rabbit | 240 ± 24 | nmol P5C/mg protein/h | [15] | |||||
| Iris (ciliary body) | Rat | 308 ± 26 | nmol P5C/mg protein/h | [15] | ||||
| Rabbit | 227 ± 28 | nmol P5C/mg protein/h | [15] | |||||
| Calf | 218 ± 26 | nmol P5C/mg protein/h | [15] | |||||
| Human | 165 ± 17 | nmol P5C/mg protein/h | [15] | |||||
| Liver | Rat | 2.8 | [6] | |||||
| 458 ± 56 | nmol P5C/mg protein/h | [15] | ||||||
| 7.8 | 2.7 | 3.0 | 11 | µmol/mg protein/min | [17] | |||
| *** | 8.0 | 0.56 | 0.91 | [18] | ||||
| 8.15 | [19] | |||||||
| 1 | 0.7 | [14] | ||||||
| Mouse | 7.5 | 4,8 | 0.7 | 10,8 | µmol/mg protein/min | [17] | ||
| ** | 1.2 ± 0.11 | 2 ± 0.5 | 0,75 | µmol P5C/mg proteine/h | [20] | |||
| Rabbit | 317 ± 34 | nmol P5C/mg protein/h | [15] | |||||
| Trout ** | 7.3 | 7.5 ± 0.8 | 0.88 ± 0.2 | 0,033 | µmol/min | [21] | ||
| Isolated mitochondria | Rat * | 4 | 0,15 | 25 | nmol/min/mg protein | [22] | ||
| Kidney | Rat | |||||||
| *** | 8.0 | 0.59 | 0.91 | 920 | units/mg protein | [18] | ||
| Rabbit | 674 ± 48 | nmol P5C/mg protein/h | [15] | |||||
| Brain | Rat | 488 ± 42 | nmol P5C/mg protein/h | [15] | ||||
| Rat | 8 | 1,67 | 0,5 | [23] | ||||
| Rabbit | 112 ± 18 | nmol P5C/mg protein/h | [15] | |||||
| 69 ± 18 | nmol P5C/mg protein/h | [15] | ||||||
| Mouse ** | 1.1 ± 0.1 | 2.6 | 1.1 | µmol P5C/mg protein/h | [20] | |||
| Cortical interneurons | Mouse * | 0.8 ± 0.3 | [24] | |||||
| Cerebellar granule cells | Mouse * | 4.7 ± 0.9 | [24] | |||||
| Astrocytes | Mouse * | 4.3 ± 2.2 | [24] | |||||
| Small intestine | Rat *** | 8.0 | 0.60 | 0.95 | [18] | |||
| Modiolus demissus | [25] | |||||||
| Plasmodium falciparum | 3.95 ± 1.04 | 0.65 ± 0.21 | [26] | |||||
| Pea (Pisum sativorum) | 8.8 | 15 | 2 | 60.4 | nmol/mg/s | [27] | ||
| Mothbean (Vignia aconitifolia) | 8 | 2 | 0.75 | [28] | ||||
| Inhibitor | Structure | Nature | Experimental Conditions | Ki or % Inhibition | OAT Origin | Reference |
|---|---|---|---|---|---|---|
| α-Amino acids | ||||||
| l-Canaline |  | Irreversible | 8 U OAT, 37 °C, pH 8, 0.05 mM PO43− buffer 0.1 mM PLP, 175 mM Orn, 35 mM aKG | Ki: 2 µM | Rat (liver) | [39] |
| Irreversible, noncompetitive | pH 8, 0.05 mM PO43− buffer 0.01 mM PLP, 20 mM Orn, 10 mM aKG | Rat (liver) | [40] | |||
| l-Canavanine | 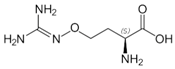 | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 100% at 25 mM | Rat (liver) | [6] | |
| 0.5 U OAT; 50 mM Orn | 23% at 20 mM | Rat (kidney) | [41] | |||
| l-Norvaline | 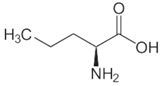 | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 59% at 50 mM | Rat (liver) | [6] | |
| pH 8, 50 mM PO43− buffer 0.1 mM PLP, 10 mM Orn, 5 mM aKG | 50% at 10 mM | Rat (liver) | [14] | |||
| 0.5 U OAT; 50 mM Orn | 44% at 100 mM | Rat (kidney) | [41] | |||
| l-Valine | 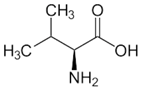 | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 47% at 25 mM | Rat (liver) | [6] | |
| 0.5 U OAT; 50 mM Orn | 33% at 25 mM | Rat (kidney) | [41] | |||
| Competitive for Orn; uncompetitive for aKG | pH 8, 50 mM PO43− buffer 0.1 mM PLP, 10 mM Orn, 5 mM aKG | 2.0 mM (forward reaction) | Rat (liver) | [14] | ||
| Noncompetitive for Glu and P5C | 37 °C, pH 8, 50 mM Tris-HCl buffer 0.1 mM PLP, 5 mM P5C, 40 mM l-Glu | 20 mM (reverse reaction) | Rat (liver) | [14] | ||
| Competitive | pH 8, 12.5 mM Tris-HCl buffer 0.025 mM PLP, 5 mM Orn, 2.5 aKG | 28% at 2.5 mM 41% at 5 mM | Rat (brain) | [23] | ||
| l-Isoleucine | 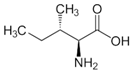 | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 27% at 25 mM | Rat (liver) | [6] | |
| pH 8, 50 mM PO43− buffer 0.1 mM PLP, 10 mM Orn, 5 mM aKG | 30% at 10 mM | Rat (liver) | [14] | |||
| 0.5 U OAT; 50 mM Orn | 19% at 25 mM | Rat (kidney) | [41] | |||
| pH 8, 12.5 mM Tris-HCl buffer 0.025 mM PLP, 5 mM Orn, 2.5 aKG | 29% at 2.5 mM 29% at 5 mM | Rat (brain) | [23] | |||
| l-Leucine | 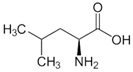 | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 21% at 25 mM | Rat (liver) | [6] | |
| pH 8, 50 mM PO43− buffer 0.1 mM PLP, 10 mM Orn, 5 mM aKG | 20% at 10 mM | Rat (liver) | [14] | |||
| 0.5 U OAT; 50 mM Orn | 17% at 25 mM | Rat (kidney) | [41] | |||
| pH 8, 12.5 mM Tris-HCl buffer 0.025 mM PLP, 5 mM Orn, 2.5 aKG | 8% at 2.5 mM 22% at 5 mM | Rat (brain) | [23] | |||
| Cysteine | pH 8, 12.5 mM Tris-HCl buffer 0.025 mM PLP, 5 mM Orn, 2.5 aKG | 48% at 2.5 mM 66% at 5 mM | Rat (brain) | [23] | ||
| 2,4-Diaminobutyrate | 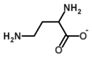 | Competitive | 37 °C, pH 8, 50 mM PO43− buffer 175 mM Orn, 35 mM aKG | 25% at 10 mM 40% at 25 mM | Rat (liver) | [42] |
| γ-Amino acids | ||||||
| γ-Aminobutyrate | 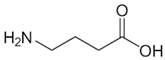 | Competitive | 37 °C, pH 8, 50 mM PO43− buffer 175 mM Orn, 35 mM aKG | 3.4 mM 70% at 10 mM 90% at 25 mM | Rat (liver) | [42] |
| 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 47% at 25 mM | Rat (liver) | [6] | |||
| pH 8, 50 mM PO43− buffer 0.1 mM PLP, 10 mM Orn, 5 mM aKG | 26% at 10 mM | Rat (liver) | [14] | |||
| (GABA) | 0.5 U OAT; 50 mM Orn | 26% at 25 mM | Rat (kidney) | [41] | ||
| 5-Aminovalerate | 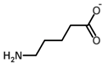 | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 100% at 25 mM | Rat (liver) | [6] | |
| (δ-Aminovalerate) | 0.5 U OAT; 50 mM Orn | 15% at 25 mM | Rat (kidney) | [41] | ||
| Competitive for Orn | pH 8, 0.05 mM PO43− buffer 0.01 mM PLP, 20 mM Orn, 10 mM aKG | 17.7 mM | Rat (liver) | [40] | ||
| ε-Aminocaproate | 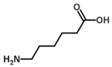 | 0.5 U OAT; 50 mM Orn | 6% at 25 mM | Rat (kidney) | [41] | |
| α-Ketoacids | ||||||
| α-Ketovalerate |  | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 66% at 21 mM | Rat (liver) | [6] | |
| 0.5 U OAT; 50 mM Orn | 38% at 25 mM | Rat (kidney) | [41] | |||
| α-Ketoisocaproate |  | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 60% at 19 mM | Rat (liver) | [6] | |
| (Ketoleucine) | 0.5 U OAT; 50 mM Orn | 41% at 25 mM | Rat (kidney) | [41] | ||
| α-Ketoisovalerate |  | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 45% at 22 mM | Rat (liver) | [6] | |
| 0.5 U OAT; 50 mM Orn | 34% at 25 mM | Rat (kidney) | [41] | |||
| α-Ketocaproate | 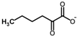 | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 18% at 15 mM | Rat (liver) | [6] | |
| 0.5 U OAT; 50 mM Orn | 34% at 25 mM | Rat (kidney) | [41] | |||
| α-Ketobutyrate | 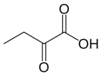 | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 13% at 10 mM | Rat (liver) | [6] | |
| 0.5 U OAT; 50 mM Orn | 27% at 25 mM | Rat (kidney) | [41] | |||
| Polyamines | ||||||
| Cadaverine | H2N-(CH2)5-NH2 | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 14% at 25 mM | Rat (liver) | [6] | |
| 0.5 U OAT; 50 mM Orn | 5.7% at 25 mM | Rat (kidney) | [41] | |||
| Putrescine | H2N-CH2-CH2-CH2-CH2-NH2 | 37 °C, pH 7.1, 50 mM PO43− buffer 35 mM Orn, 3.75 mM aKG,K | 14% at 25 mM | Rat (liver) | [6] | |
| pH 8, 50 mM PO43− buffer 0.1 mM PLP, 10 mM Orn, 5 mM aKG | No inhibition at 10 mM | Rat (liver) | [14] | |||
| 0.5 U OAT; 50 mM Orn | 3% at 25 mM | Rat (kidney) | [41] | |||
| pH 8, 12.5 mM Tris-HCl buffer 0.025 mM PLP, 5 mM Orn, 2.5 aKG | No inhibition at 5 mM 8% at 10 mM | Rat (brain) | [23] | |||
| Spermine | 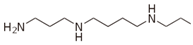 | pH 8, 12.5 mM Tris-HCl buffer 0.025 mM PLP, 5 mM Orn, 2.5 aKG | 33% at 5 mM 56% at 10 mM | Rat (brain) | [23] | |
| pH 8, 50 mM PO43− buffer 0.1 mM PLP, 10 mM Orn, 5 mM aKG | No inhibition at 10 mM | Rat (liver) | [14] | |||
| Spermidine | 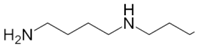 | pH 8, 12.5 mM Tris-HCl buffer 0.025 mM PLP, 5 mM Orn, 2.5 aKG | 30% at 5 mM 48% at 10 mM | Rat (brain) | [23] | |
| Histamine | 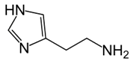 | pH 8, 12.5 mM Tris-HCl buffer 0.025 mM PLP, 5 mM Orn, 2.5 aKG | 42% at 5 mM 50% at 10 mM | Rat (brain) | [23] | |
| Other inhibitors | ||||||
| 5-Fluoromethylornithine | 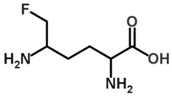 | Irreversible | 37 °C, pH 8, 50 mM PO43− buffer 175 mM Orn, 35 mM aKG | 70 µM | Rat (liver; partially purified) | [37] |
| 8 U OAT, 37°C, pH 8, 0.05 mM PO43− buffer 0.1 mM PLP, 175 mM Orn, 35 mM aKG | 30 µM | Rat (liver) | [39] | |||
| Not given | 11 ± 2 µM | [36] | ||||
| S,S-5-Fluoromethylornithine | Not given | 2.4 ± 0.3 µM | [36] | |||
| N-Acetylornithine | 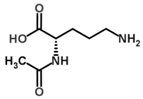 | Competitive with Orn | pH 8, 0.05 mM PO43− buffer 0.01 mM PLP, 20 mM Orn, 10 mM aKG | 8.8 mM | Rat (liver) | [40] |
| pH 8, 12.5 mM Tris-HCl buffer 0.025 mM PLP, 5 mM Orn, 2.5 aKG | 24% at 2.5 mM 27% at 5 mM | Rat (brain) | [23] | |||
| 4-Aminohexynoate | 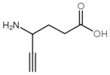 | Irreversible | 0.5 U OAT, 37 °C, 40 mM KPP0,020 mM PLP, 60 mM Orn, 21 mM aKG | Rat (liver) | [43] | |
| Gabaculine | 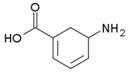 | Irreversible | 0.5 U OAT, 37 °C, 40 mM KPP 0,020 mM PLP, 60 mM Orn, 21 mM aKG | Rat (liver) | [43] | |
| 2-aminooxyacetate | 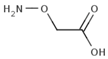 | “almost complete” at 0.2 mM | Rat | [44] |
| Organ | Tissue | Species | Activity | Unit | Reference |
|---|---|---|---|---|---|
| Eye | Neuroretina | Cat | 88.2 | mmol/kg dry wt/h | [70] |
| Ox | 71 ± 17 | nmol/mg protein/30 min | [132] | ||
| Optic nerve | Cat | 6.7 | mmol/kg dry wt/h | [70] | |
| Tapetum | Cat | 26.7 | mmol/kg dry wt/h | [70] | |
| Sclera | Cat | 5.36 | mmol/kg dry wt/h | [70] | |
| Choroid | Cat | 17.4 | mmol/kg dry wt/h | [70] | |
| Ox | 14 ± 5 | nmol/mg protein/30 min | [132] | ||
| Retina | Rat | 174 ± 25 | nmol/mg protein/h | [133] | |
| 97 ± 9 | nmol/mg protein/30 min | [132] | |||
| Ox | 119 ± 34 | nmol/mg protein/h | [133] | ||
| Pig | 409 ± 50 | nmol/mg protein/h | [133] | ||
| Dog | 159 | nmol/mg protein/h | [133] | ||
| Chicken | 124 ± 16 | nmol/mg protein/h | [133] | ||
| 120 ± 20 | nmol/mg/h | [134] | |||
| Turkey | 85 | nmol/mg protein/h | [133] | ||
| Rana pipiens | 302 | nmol/mg protein/h | [133] | ||
| Rana clamitans | 238 | nmol/mg protein/h | [133] | ||
| Xenopus laevis | 392 ± 93 | nmol/mg protein/h | [133] | ||
| Cyprinus carpio | 766 ± 57 | nmol/mg protein/h | [133] | ||
| Pigment epithelium (iris epithelium) | Cat | 138 | mmol/kg dry wt/h | [70] | |
| Rat | 988 ± 130 | nmol/mg protein/h | [133] | ||
| Ox | 497 ± 89 | nmol/mg protein/h | [133] | ||
| 113 ± 24 | nmol/mg protein/30 min | [132] | |||
| Pig | 2260 ± 630 | nmol/mg protein/h | [133] | ||
| Dog | 602 ± 159 | nmol/mg protein/h | [133] | ||
| Chicken | 1431 ± 535 | nmol/mg protein/h | [133] | ||
| 1670 ± 55 | nmol/mg/h | [134] | |||
| Turkey | 1150 | nmol/mg protein/h | [133] | ||
| Rana pipiens | 795 | nmol/mg protein/h | [133] | ||
| Rana clamitans | 217 | nmol/mg protein/h | [133] | ||
| Xenopus laevis | 171 ± 87 | nmol/mg protein/h | [133] | ||
| Cyprinus carpio | 203 ± 51 | nmol/mg protein/h | [133] | ||
| Vascular layer of the ciliary body | Cat | 63.0 | mmol/kg dry wt/h | [70] | |
| Cornea | Cat | 8.11 | mmol/kg dry wt/h | [70] | |
| Ox | 14 ± 9 | nmol/mg protein/30 min | [132] | ||
| Lens | Cat | No | [70] | ||
| Ox | No | [132] | |||
| Brain | Frontal cortex | Rat | 1.45 ± 0.29 | mU/mg protein | [135] |
| Occipital cortex | Rat | 1.38 ± 0.08 | mU/mg protein | [135] | |
| Olfactory bulb | Rat | 1.23 ± 0.07 | mU/mg protein | [135] | |
| Hippocampus | Rat | 1.33 ± 0.16 | mU/mg protein | [135] | |
| Striatum | Rat | 0.95 ± 0.06 | mU/mg protein | [135] | |
| Midbrain, thalamus | Rat | 1.17 ± 0.12 | mU/mg protein | [135] | |
| Hypothalamus | Rat | 1.34 ± 0.10 | mU/mg protein | [135] | |
| Pons, medulla oblongata | Rat | 0.85 ± 0.04 | mU/mg protein | [135] | |
| Cerebellum | Rat | 0.90 ± 0.19 | mU/mg protein | [135] | |
| Tractus opticus | Rat | 0.75 ± 0.10 | mU/mg protein | [135] | |
| Spinal cord | Rat | 0.60 ± 0.07 | mU/mg protein | [135] | |
| Liver | Rat | 179 | nmol/mg protein/h | [133] | |
| 242 ± 41 | nmol/mg protein/30 min | [132] | |||
| Pig | 164 | nmol/mg protein/h | [133] | ||
| Ox | 12 | nmol/mg protein/30 min | [132] | ||
| Chicken | 174 ± 22 | nmol/mg protein/h | [133] | ||
| 150 ± 40 | nmol/mg/h | [134] | |||
| Rana clamitans | 137 | nmol/mg protein/h | [133] | ||
| Xenopus laevis | 161 ± 101 | nmol/mg protein/h | [133] | ||
| Cyprinus carpio | 590 | nmol/mg protein/h | [133] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ginguay, A.; Cynober, L.; Curis, E.; Nicolis, I. Ornithine Aminotransferase, an Important Glutamate-Metabolizing Enzyme at the Crossroads of Multiple Metabolic Pathways. Biology 2017, 6, 18. https://doi.org/10.3390/biology6010018
Ginguay A, Cynober L, Curis E, Nicolis I. Ornithine Aminotransferase, an Important Glutamate-Metabolizing Enzyme at the Crossroads of Multiple Metabolic Pathways. Biology. 2017; 6(1):18. https://doi.org/10.3390/biology6010018
Chicago/Turabian StyleGinguay, Antonin, Luc Cynober, Emmanuel Curis, and Ioannis Nicolis. 2017. "Ornithine Aminotransferase, an Important Glutamate-Metabolizing Enzyme at the Crossroads of Multiple Metabolic Pathways" Biology 6, no. 1: 18. https://doi.org/10.3390/biology6010018
APA StyleGinguay, A., Cynober, L., Curis, E., & Nicolis, I. (2017). Ornithine Aminotransferase, an Important Glutamate-Metabolizing Enzyme at the Crossroads of Multiple Metabolic Pathways. Biology, 6(1), 18. https://doi.org/10.3390/biology6010018