
The Emergence of Physiology and Form: Natural Selection Revisited

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Biologic Structures and Processes in an Evolutionary Context
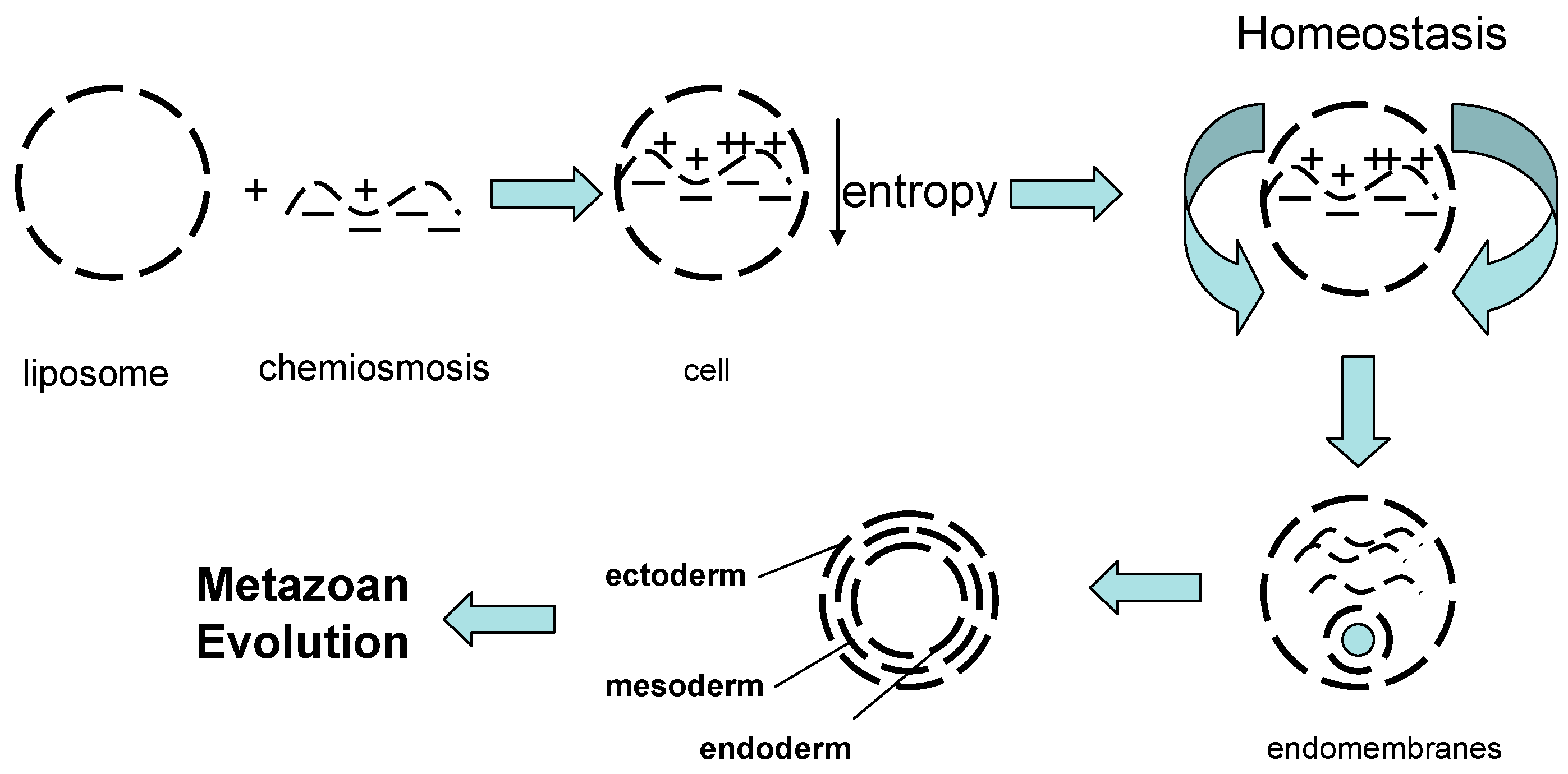
2.1. The Cell as the Fundament for Natural Selection
2.2. Force of Gravity and Development
2.3. The Glomerulus: A Mechanotransductive Homolog of the Alveolus
2.4. Skin as a Vertical Integration of Vertebrate Physiology
3. Model Organisms as Functional Homologs Revealed by the Molecular Physiologic Effects of Gravity
3.1. Yeast
3.2. Dictyostelium Discoideum
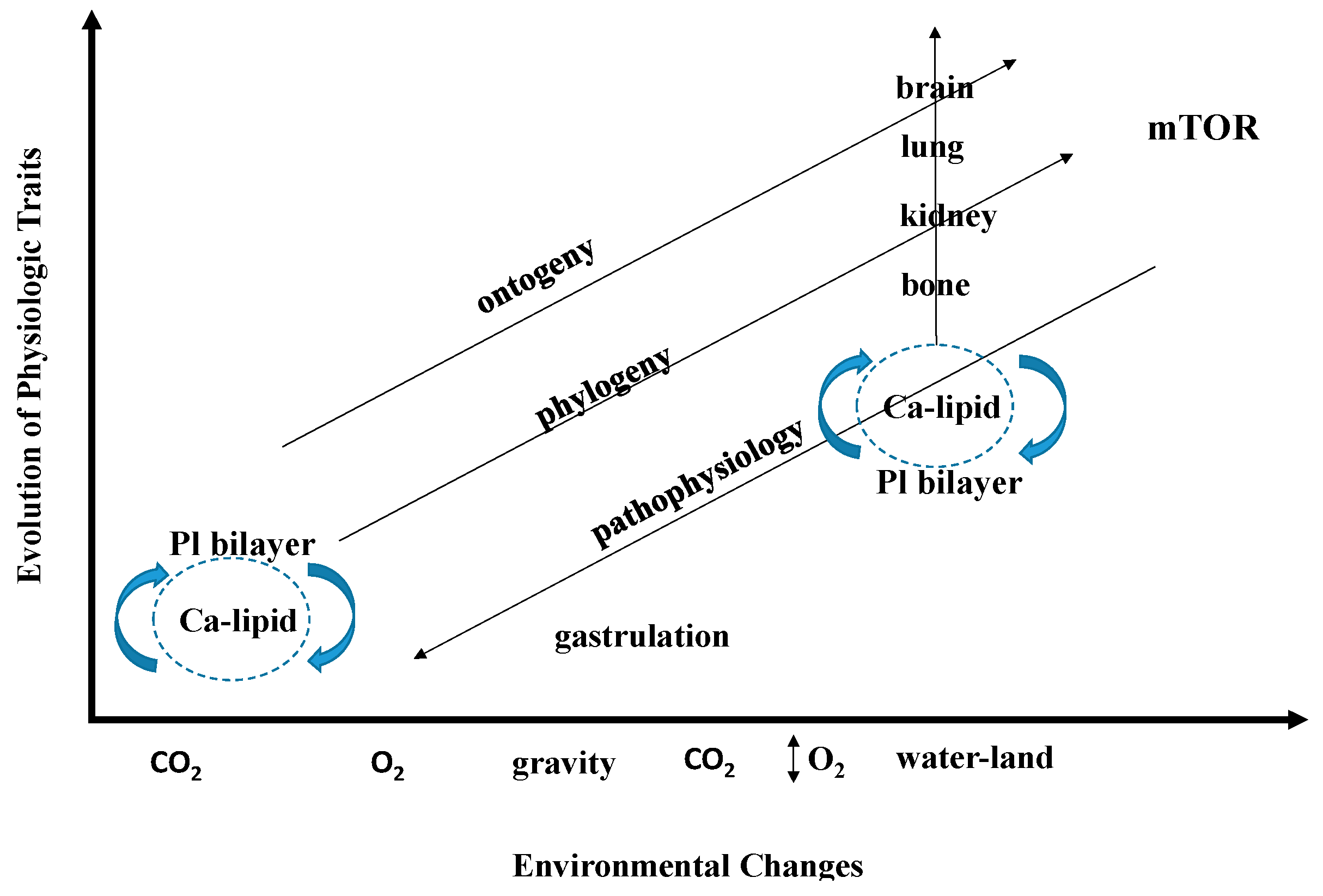
4. Fractal Properties of Physiology
4.1. Fractal Physiology—How and Why
4.2. Fractal Physiology—In the Beginning
4.3. Fractal Properties of Physiology and Chemiosmosis
4.4. Fractal Properties of Physiology and the Evolution of Calcium Homeostasis: From the First Cell to a Ubiquitous Intracellular Signaling System
5. Life has Authored Itself
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- Calisher, C.H. Taxonomy: What’s in a name? Doesn’t a rose by any other name smell as sweet? Croat. Med. J. 2007, 48, 268–270. [Google Scholar] [PubMed]
- Torday, J.S. A central theory of biology. Med. Hypotheses 2015, 85, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. Up-regulation of fetal rat lung parathyroid hormone-related protein gene regulatory network down-regulates the Sonic Hedgehog/Wnt/betacatenin gene regulatory network. Pediatr. Res. 2006, 60, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. The evolutionary continuum from lung development to homeostasis and repair. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 292, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. Lung evolution as a cipher for physiology. Physiol. Genomics 2009, 38, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.K.; Trisnadi, N.; Kadam, S.; Stathopoulos, A. The role of FGF signaling in guiding coordinate movement of cell groups: Guidance cue and cell adhesion regulator? Cell Adh. Migr. 2012, 6, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Barker, N. The canonical Wnt/beta-catenin signalling pathway. Methods Mol. Biol. 2008, 468, 5–15. [Google Scholar] [PubMed]
- Kohsokabe, T.; Kaneko, K. Evolution-development congruence in pattern formation dynamics: Bifurcations in gene expression and regulation of networks structures. J. Exp. Zool. B Mol. Dev. Evol. 2016, 326, 61–84. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S. The cell as the mechanistic basis for evolution. Wiley Interdiscip. Rev. Syst. Biol. Med. 2015, 7, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Moessinger, A.C.; Harding, R.; Adamson, T.M.; Singh, M.; Kiu, G.T. Role of lung fluid volume in growth and maturation of the fetal sheep lung. J. Clin. Invest. 1990, 86, 1270–1277. [Google Scholar] [CrossRef] [PubMed]
- Wolpert, L.; Vicente, C. An interview with Lewis Wolpert. Development 2015, 142, 2547–2548. [Google Scholar] [PubMed]
- Hastings, R.H. Parathyroid hormone-related protein and lung biology. Respir. Physiol. Neurobiol. 2004, 142, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Deschaseaux, F.; Sensébé, L.; Heymann, D. Mechanisms of bone repair and regeneration. Trends Mol. Med. 2009, 15, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Endlich, N.; Endlich, K. cAMP pathway in podocytes. Microsc. Res. Tech. 2002, 57, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Raj, J.U. Parathyroid hormone-related protein-mediated responses in pulmonary arteries and veins of newborn lambs. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 289, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.P.; Kovacs, C.S.; De Paepe, M.E.; Tsai, S.W.; Torday, J.S.; Kronenberg, H.M. Arrested pulmonary alveolar cytodifferentiation and defective surfactant synthesis in mice missing the gene for parathyroid hormone-related protein. Dev. Dyn. 2004, 230, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Hochane, M.; Raison, D.; Coquard, C.; Imhoff, O.; Massfelder, T.; Moulin, B.; Helwig, J.J.; Barthelmebs, M. Parathyroid hormone-related protein is a mitogenic and a survival factor of mesangial cells from male mice: Role of intracrine and paracrine pathways. Endocrinology 2013, 154, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. Developmental cell/molecular biologic approach to the etiology and treatment of bronchopulmonary dysplasia. Pediatr. Res. 2007, 62, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.P.; Kifor, O.; Hua, J.; Brown, E.M.; Torday, J.S. Parathyroid hormone (PTH) and PTH-related protein stimulate surfactant phospholipid synthesis in rat fetal lung, apparently by a mesenchymal-epithelial mechanism. Biochim. Biophys. Acta 1994, 23, 91–100. [Google Scholar] [CrossRef]
- Schultz, C.J.; Torres, E.; Londos, C.; Torday, J.S. Role of adipocyte differentiation-related protein in surfactant phospholipid synthesis by type II cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 283, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, A.R.; Brasaemle, D.L.; McAndrews-Hill, M.; Sztalryd, C.; Londos, C. Adoption of PERILIPIN as a unifying nomenclature for the mammalian PAT-family of intracellular lipid storage droplet proteins. J. Lipid Res. 2010, 51, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Griffith, E.C.; Rapf, R.J.; Shoemaker, R.K.; Carpenter, B.K.; Vaida, V. Photoinitiated synthesis of self-assembled vesicles. J. Amer. Chem. Soc. 2014, 136, 3784–3787. [Google Scholar] [CrossRef] [PubMed]
- Griffith, E.C.; Vaida, V. In situ observation of peptide bond formation at the water-air interface. Proc. Nat. Acad. Sci. USA 2012, 109, 15697–15701. [Google Scholar] [CrossRef] [PubMed]
- Tuck, A. The role of atmospheric aerosols in the origin of life. Surv. Geophys. 2002, 23, 379–409. [Google Scholar] [CrossRef]
- Kazmierczak, J.; Kempe, S.; Kremer, B. Calcium in the early evolution of living systems: A biohistorical approach. Curr. Org. Chem. 2013, 17, 1738–1750. [Google Scholar] [CrossRef]
- De Duve, C. Evolution of the peroxisome. Ann. NY Acad. Sci. 1969, 168, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Csete, M.; Walikonis, J.; Slawny, N.; Wei, Y.; Korsnes, S.; Doyle, J.C.; Wold, B. Oxygen-mediated regulation of skeletal muscle satellite cell proliferation and adipogenesis in culture. J. Cell Physiol. 2001, 189, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Boros, L.G.; Torday, J.S.; Paul Lee, W.N.; Rehan, V.K. Oxygen-induced metabolic changes and transdifferentiation in immature fetal rat lung lipofibroblasts. Mol. Genet. Metab. 2002, 77, 230–236. [Google Scholar] [CrossRef]
- Rehan, V.; Torday, J. Hyperoxia augments pulmonary lipofibroblast-to-myofibroblast transdifferentiation. Cell Biochem. Biophys. 2003, 38, 239–250. [Google Scholar] [CrossRef]
- Rehan, V.K.; Wang, Y.; Patel, S.; Santos, J.; Torday, J.S. Rosiglitazone, a peroxisome proliferator-activated receptor-gamma agonist, prevents hyperoxia-induced neonatal rat lung injury in vivo. Pediatr. Pulmonol. 2006, 41, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.; Hua, J.; Slavin, R. Metabolism and fate of neutral lipids of fetal lung fibroblast origin. Biochim. Biophys. Acta 1995, 1254, 198–206. [Google Scholar] [CrossRef]
- McGowan, S.E.; Torday, J.S. The pulmonary lipofibroblast (lipid interstitial cell) and its contributions to alveolar development. Annu. Rev. Physiol. 1997, 59, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. Deconvoluting lung evolution using functional/comparative genomics. Am. J. Respir. Cell Mol. Biol. 2004, 31, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Torres, E.; Rehan, V.K. The role of fibroblast transdifferentiation in lung epithelial cell proliferation, differentiation, and repair in vitro. Pediatr. Pathol. Mol. Med. 2003, 22, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Sun, H.; Qin, J. Prostaglandin E2 integrates the effects of fluid distension and glucocorticoid on lung maturation. Am. J. Physiol. 1998, 274, 106–111. [Google Scholar]
- West, J.B. Respiratory Physiology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012. [Google Scholar]
- Nielsen, T.S.; Jessen, N.; Jørgensen, J.O.; Møller, N.; Lund, S. Dissecting adipose tissue lipolysis: Molecular regulation and implications for metabolic disease. J. Mol. Endocrinol. 2014, 52, 199–222. [Google Scholar] [CrossRef] [PubMed]
- Kolditz, C.I.; Langin, D. Adipose tissue lipolysis. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Frayn, K.N. Fat as a fuel: Emerging understanding of the adipose tissue-skeletal muscle axis. Acta Physiol. 2010, 199, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Torday, J. Cellular timing of fetal lung development. Semin. Perinatol. 1992, 16, 130–139. [Google Scholar] [PubMed]
- Torday, J.S.; Sun, H.; Wang, L.; Torres, E.; Sunday, M.E.; Rubin, L.P. Leptin mediates the parathyroid hormone-related protein paracrine stimulation of fetal lung maturation. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 282, 405–410. [Google Scholar]
- Pan, H.; Guo, J.; Su, Z. Advances in understanding the interrelations between leptin resistance and obesity. Physiol. Behav. 2014, 130, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. Stretch-stimulated surfactant synthesis is coordinated by the paracrine actions of PTHrP and leptin. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 283, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wang, Z.; Collins, J.E.; Andrews, R.M.; Stemple, D.; Gong, Z. Comparative transcriptome analyses indicate molecular homology of zebrafish swimbladder and mammalian lung. PLoS ONE 2011, 6, e24019. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Ihida-Stansbury, K.; Rehan, V.K. Leptin stimulates Xenopus lung development: Evolution in a dish. Evol. Dev. 2009, 11, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. A cell-molecular approach predicts vertebrate evolution. Mol. Biol. Evol. 2011, 28, 2973–2981. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S. Evolutionary biology redux. Perspect. Biol. Med. 2013, 56, 455–484. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S. Heterochrony as diachronically modified cell-cell interactions. Biology 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S. What we talk about when we talk about evolution. Cell Commun. Insights. 2015, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Endlich, N.; Nobiling, R.; Kriz, W.; Endlich, K. Expression and signaling of parathyroid hormone-related protein in cultured podocytes. Exp. Nephrol. 2001, 9, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Bosch, R.J.; Rodríguez-Puyol, D.; Bover, J.; Rodríguez-Puyol, M. Parathyroid hormone-related protein: Roles in the glomerulus. Exp. Nephrol. 1999, 7, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Husain-Syed, F.; McCullough, P.A.; Birk, H.W.; Renker, M.; Brocca, A.; Seeger, W.; Ronco, C. Cardio-pulmonary-renal interactions: A Multidisciplinary approach. J. Am. Coll. Cardiol. 2015, 65, 2433–2448. [Google Scholar] [CrossRef] [PubMed]
- Holland, N.D. Early central nervous system evolution: An era of skin brains? Nat. Rev. Neurosci. 2003, 4, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Aberg, K.M.; Man, M.Q.; Gallo, R.L.; Ganz, T.; Crumrine, D.; Brown, B.E.; Choi, E.H.; Kim, D.K.; Schröder, J.M.; Feingold, K.R.; et al. Co-regulation and interdependence of the mammalian epidermal permeability and antimicrobial barriers. J. Invest. Dermatol. 2008, 128, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Palaniyar, N.; Ikegami, M.; Korfhagen, T.; Whitsett, J.; McCormack, F.X. Domains of surfactant protein A that affect protein oligomerization, lipid structure and surface tension. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 129, 109–127. [Google Scholar] [CrossRef]
- Palaniyar, N.; Ridsdale, R.A.; Holterman, C.E.; Inchley, K.; Possmayer, F.; Harauz, G. Structural changes of surfactant protein A induced by cations reorient the protein on lipid bilayers. J. Struct. Biol. 1998, 122, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Santa Maria, C.; Cheng, Z.; Li, A.; Wang, J.; Shoback, D.; Tu, C.L.; Chang, W. Interplay between CaSR and PTH1R signaling in skeletal development and osteoanabolism. Semin. Cell Dev. Biol. 2016, 49, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Purevdorj-Gage, B.; Sheehan, K.B.; Hyman, L.E. Effects of low-shear modeled microgravity on cell function, gene expression, and phenotype in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2006, 72, 4569–4575. [Google Scholar] [CrossRef] [PubMed]
- Pincheira-Donoso, D.; Hunt, J. Fecundity selection theory: Concepts and evidence. Biol. Rev. Camb. Philos. Soc. 2015. [Google Scholar] [CrossRef] [PubMed]
- Hardin, J.; Bertoni, G.; Kleinsmith, L.J. Becker’s World of the Cell; Pearson: New York, NY, USA, 2015. [Google Scholar]
- Gefen, A.; Weihs, D. Mechanical cytoprotection: A review of cytoskeleton-protection approaches for cells. J. Biomech. 2015. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.; Li, R. Actin cytoskeleton in cell polarity and asymmetric division during mouse oocyte maturation. Cytoskeleton 2012, 69, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Conduit, P.T.; Wainman, A.; Raff, J.W. Centrosome function and assembly in animal cells. Nat. Rev. Mol. Cell Biol. 2015, 16, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.T.; Firtel, R.A. Regulation of chemotaxis by the orchestrated activation of Ras, PI3K, and TOR. Eur. J. Cell Biol. 2006, 85, 873–895. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Fingar, D.C. Growing knowledge of the mTOR signaling network. Semin. Cell Dev. Biol. 2014, 36, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S. Homeostasis as the mechanism of evolution. Biology 2015, 4, 573–590. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. Evolutionary Biology, Cell-Cell Communication and Complex Disease; Wiley: Hoboken, NJ, USA, 2012. [Google Scholar]
- Gould, S.J.; Lewontin, R.C. The spandrels of San Marco and the Panglossian paradigm: A critique of the adaptationist programme. Proc. R. Soc. Lond. B Biol. Sci. 1979, 205, 581–598. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. The Evolution of cell communication: The road not taken. Cell Commun. Insights 2009, 2, 17–25. [Google Scholar] [PubMed]
- Deamer, D.W. Role of amphiphilic compounds in the evolution of membrane structure on the early earth. Orig. Life Evol. Biosph. 1986, 17, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature 1961, 191, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Case, R.M.; Eisner, D.; Gurney, A.; Jones, O.; Muallem, S.; Verkhratsky, A. Evolution of calcium homeostasis: From birth of the first cell to an omnipresent signaling system. Cell Calcium 2007, 42, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Orgeig, S.; Daniels, C.B. The roles of cholesterol in pulmonary surfactant: Insights from comparative and evolutionary studies. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 129, 75–89. [Google Scholar] [CrossRef]
- Saher, G.; Stumpf, S.K. Cholesterol in myelin biogenesis and hypomyelinating disorders. Biochim. Biophys. Acta 2015, 1851, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, K. Making biological theory more down to Earth. Prog. Biophys. Mol. Biol. 2013, 113, 46–56. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torday, J.S. The Emergence of Physiology and Form: Natural Selection Revisited. Biology 2016, 5, 15. https://doi.org/10.3390/biology5020015
Torday JS. The Emergence of Physiology and Form: Natural Selection Revisited. Biology. 2016; 5(2):15. https://doi.org/10.3390/biology5020015
Chicago/Turabian StyleTorday, John S. 2016. "The Emergence of Physiology and Form: Natural Selection Revisited" Biology 5, no. 2: 15. https://doi.org/10.3390/biology5020015
APA StyleTorday, J. S. (2016). The Emergence of Physiology and Form: Natural Selection Revisited. Biology, 5(2), 15. https://doi.org/10.3390/biology5020015