
Decoding Stress Responses in Farmed Crustaceans: Comparative Insights for Sustainable Aquaculture Management

 , , ,
, , ,  and
and 
Simple Summary
Abstract
1. Introduction
2. Scope of Stress in Aquaculture
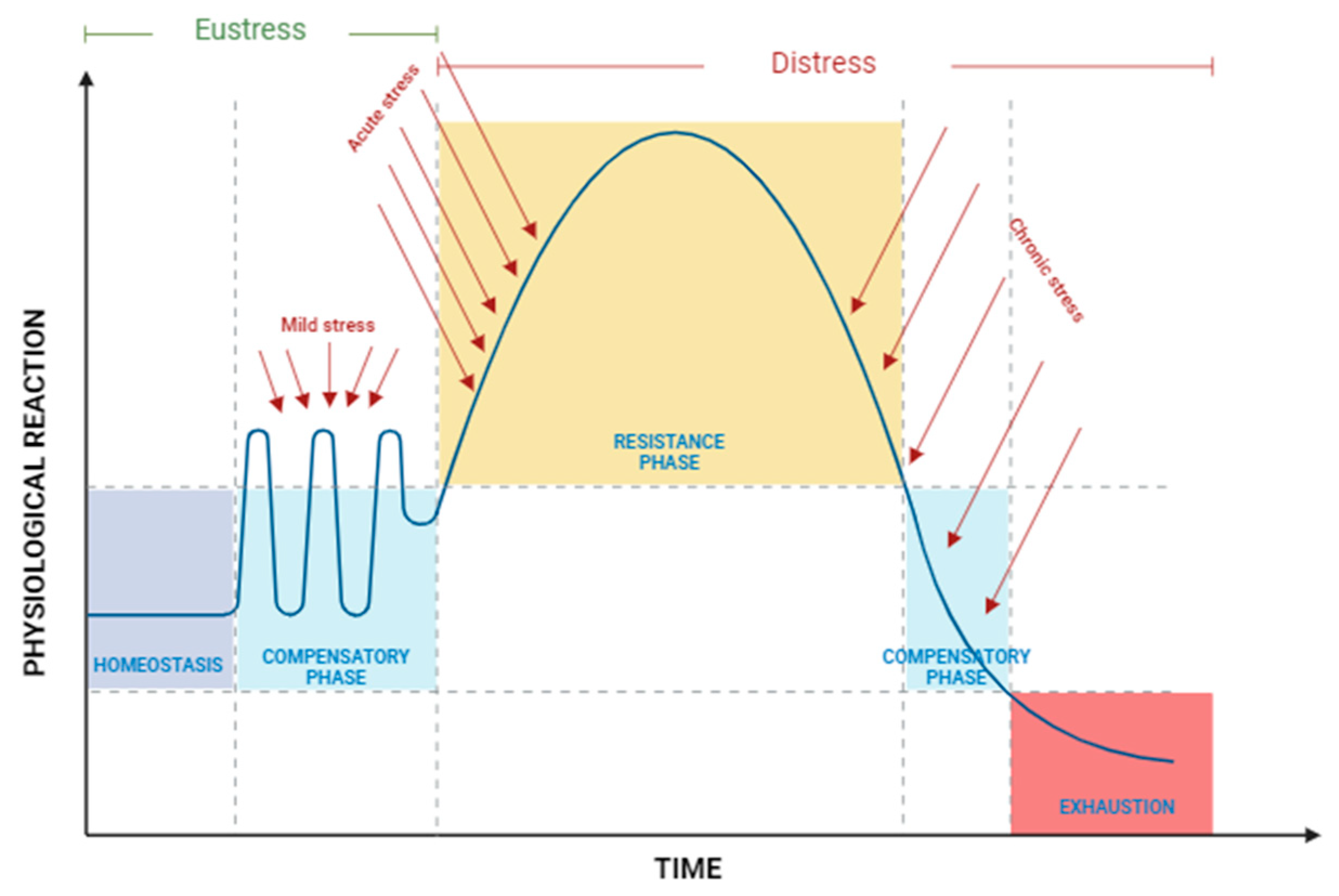
2.1. Definition of Stress
2.2. Types and Sources of Stress
2.2.1. Stressors According to Exposure Duration and Intensity
2.2.2. Stressors According to Source

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Stressor | Stressors | Range | Species | Reference |
|---|---|---|---|---|
| Physical | Temperature | 10 ± 0.5 °C | Marsupenaeus japonicus | [13] |
| Light intensity and photoperiod | 12 L:12 D–24 L:0 D | Eriocheir sinensis | [52] | |
| Sound | 128 dB re 1 µPa | Litopenaeus vannamei | [57] | |
| Turbidity | 30–120 NTU | Litopenaeus vannamei | [58] | |
| Chemical | Dissolved oxygen | 0.8–3.5 mg/L | Litopenaeus vannamei | [11] |
| Ammonia | 60.21 mg/L | Litopenaeus vannamei | [29] | |
| Nitrite | 5–15 mg/L | Scylla paramamosain | [61] | |
| Nitrate | 35–910 ppm | Litopenaeus vannamei | [63] | |
| Hardness | 25–1000 mg/L | Macrobrachium rosenbergii | [85] | |
| pH | 9.5 | Litopenaeus vannamei | [24] | |
| Alkalinity | 10 mmol/L | Exopalaemon carinicauda | [65] | |
| Dissolved CO | 14.5–175.0 mg/L | Litopenaeus vannamei | [66] | |
| Salinity | 3 ppt | Litopenaeus vannamei | [67] | |
| Feed | CBH:P 0.6–2.1; L:P 0.2–0.36 | Litopenaeus vannamei | [73] | |
| Pollutants | 500 μg/L Cd and 500 μg/L Pb | Litopenaeus vannamei | [32] | |
| Toxins | 0 and 4 mg/kg | Eriocheir sinensis | [71] | |
| Biological | Pathogens | 100 µL of a 1 × 1011 CFU/L | Penaeus monodon | [86] |
| Parasites | Not stated | Macrobrachium nipponense | [75] | |
| Predators | Open-flow kairomones | Jasus edwardsii | [76] | |
| Overcrowding | 30–60 PL8/m2 | Litopenaeus vannamei | [78] | |
| Procedural | Handling and hauling | Every morning for 4 weeks | Litopenaeus vannamei | [79] |
| Confinement | Bottom pond for 48 h | Litopenaeus stylirostris | [82] | |
| Disease treatment | 1 mg/L for 1 week | Litopenaeus vannamei | [87] | |
| Feeding | 1–5 times/day | Procambarus clarkia | [88] | |
| Transportation | Water-free for 10 h | Litopenaeus vannamei | [83] |
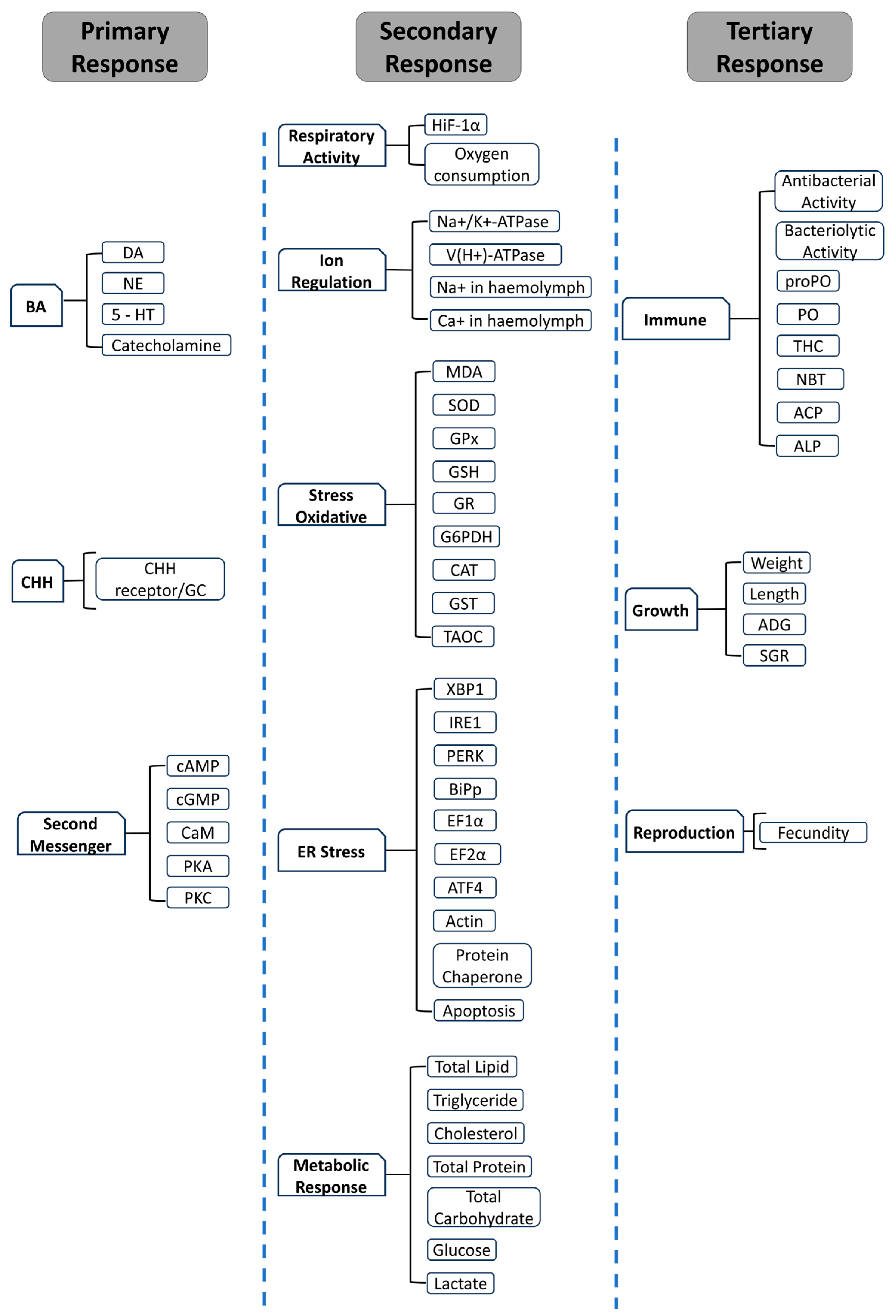
3. Stress Response
3.1. Primary Response
3.2. Secondary Response
3.2.1. Metabolic Alterations
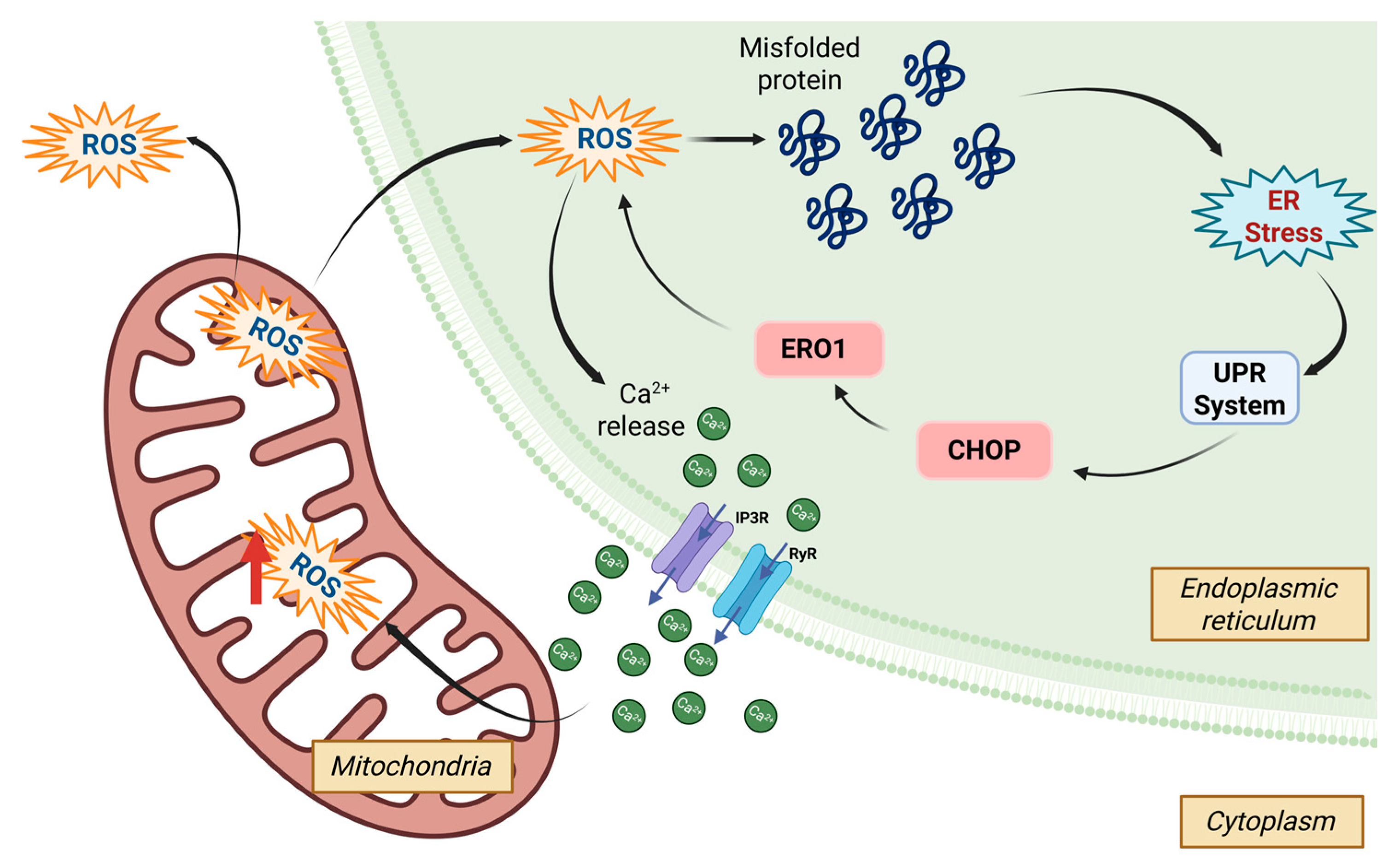
3.2.2. Unfolded Protein Mechanism
3.2.3. Redox Regulation (Antioxidant)
3.2.4. Hydromineral Regulation
3.2.5. Alternation in Immune Responses
3.3. Tertiary Response
4. Bioindicators for Stress
4.1. Hormonal Indicators
4.2. Metabolic and Cellular Indicators
4.2.1. Respiratory Process
4.2.2. Ion Regulation
4.2.3. Cellular Stress
4.2.4. Metabolic Indicators
4.3. Phenotypic Indicators
5. Stress Mitigation Strategy
6. Future Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2024—Blue Transformation in Action; FAO: Rome, Italy, 2024. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- N’Souvi, K.; Sun, C.; Che, B.; Vodounon, A. Shrimp Industry in China: Overview of the Trends in the Production, Imports and Exports during the Last Two Decades, Challenges, and Outlook. Front. Sustain. Food Syst. 2023, 7, 1287034. [Google Scholar] [CrossRef]
- Davis, R.; Abebe, A.; Boyd, C.; McNevin, A. Exploring the Relationship between Production Intensity and Land Use: A Meta-Analytic Approach with Shrimp Aquaculture. J. Environ. Manag. 2021, 300, 113719. [Google Scholar] [CrossRef] [PubMed]
- Maulu, S.; Hasimuna, O.J.; Haambiya, L.H.; Monde, C.; Musuka, C.G.; Makorwa, T.H.; Munganga, B.P.; Phiri, K.J.; Nsekanabo, J.D.M. Climate Change Effects on Aquaculture Production: Sustainability Implications, Mitigation, and Adaptations. Front. Sustain. Food Syst. 2021, 5, 609097. [Google Scholar] [CrossRef]
- Asiedu, B.; Malcolm, D.; Iddrisu, S. Assessing the Economic Impact of Climate Change in the Small-Scale Aquaculture Industry of Ghana, West Africa. AAS Open Res. 2018, 1, 26. [Google Scholar] [CrossRef] [PubMed]
- Hamdan, R.; Othman, A.; Kari, F. Climate Change Effects on Aquaculture Production Performance in Malaysia: An Environmental Performance Analysis. Int. J. Bus. Soc. 2017, 16, 364–385. [Google Scholar] [CrossRef]
- Muhala, V.; Chicombo, T.F.; Macate, I.E.; Guimarães-Costa, A.; Gundana, H.; Malichocho, C.; Hasimuna, O.J.; Remédio, A.; Maulu, S.; Cuamba, L.; et al. Climate Change in Fisheries and Aquaculture: Analysis of the Impact Caused by Idai and Kenneth Cyclones in Mozambique. Front. Sustain. Food Syst. 2021, 5, 714187. [Google Scholar] [CrossRef]
- Fleming, A.; Hobday, A.J.; Farmery, A.; van Putten, E.I.; Pecl, G.T.; Green, B.S.; Lim-Camacho, L. Climate Change Risks and Adaptation Options across Australian Seafood Supply Chains—A Preliminary Assessment. Clim. Risk Manag. 2014, 1, 39–50. [Google Scholar] [CrossRef]
- Aparicio-Simón, B.; Piñón, M.; Racotta, R.; Racotta, I.S. Neuroendocrine and Metabolic Responses of Pacific Whiteleg Shrimp Penaeus Vannamei Exposed to Hypoxia Stress. Lat. Am. J. Aquat. Res. 2018, 46, 364–376. [Google Scholar] [CrossRef]
- Han, S.Y.; Wang, M.Q.; Liu, M.; Wang, B.J.; Jiang, K.Y.; Wang, L. Comparative Sensitivity of the Hepatopancreas and Midgut in the White Shrimp Litopenaeus vannamei to Oxidative Stress under Cyclic Serious/Medium Hypoxia. Aquaculture 2018, 490, 44–52. [Google Scholar] [CrossRef]
- Wang, P.; Liu, H.; Zhao, S.; Yu, S.; Xie, S.; Hua, S.; Yan, B.; Xing, C.; Gao, H. Hypoxia Stress Affects the Physiological Responses, Apoptosis and Innate Immunity of Kuruma shrimp, Marsupenaeus japonicus. Fish Shellfish Immunol. 2022, 122, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Yu, Z.; Xu, Y.; Zhang, Y.; Mu, C.; Liu, P.; Li, J. Integrated Transcriptomic and Metabolomic Responses in the Hepatopancreas of Kuruma shrimp (Marsupenaeus japonicus) under Cold Stress. Ecotoxicol. Environ. Saf. 2020, 206, 111360. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Wang, A.; Miao, Y.; Liao, S.; Ye, C.; Lin, Q. Comparative Proteomic Identification of the Hepatopancreas Response to Cold Stress in White Shrimp, Litopenaeus vannamei. Aquaculture 2016, 454, 27–34. [Google Scholar] [CrossRef]
- Chang, C.C.; Jiang, J.R.; Cheng, W. A First Insight into Temperature Stress-Induced Neuroendocrine and Immunological Changes in Giant Freshwater Prawn, Macrobrachium rosenbergii. Fish Shellfish Immunol. 2015, 47, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, Q.; Song, X.; Pang, Y.; Song, Y.; Cheng, Y.; Yang, X. Dietary L-Tryptophan Modulates the Hematological Immune and Antibacterial Ability of the Chinese Mitten Crab, Eriocheir sinensis, Under Cheliped Autotomy Stress. Front. Immunol. 2018, 9, 2744. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.Y.; Rahman, N.A.; Shazili, N.A.M.; Chen, S.; Lv, A.; Sun, J.; Shi, H.; MacRae, T.H. Non-Lethal Heat Shock Induces Hsp70 Synthesis and Promotes Tolerance against Heat, Ammonia and Metals in Post-Larvae of the White Leg Shrimp Penaeus Vannamei (Boone, 1931). Aquaculture 2018, 483, 21–26. [Google Scholar] [CrossRef]
- Matozzo, V.; Gallo, C.; Marin, M.G. Effects of Temperature on Cellular and Biochemical Parameters in the Crab Carcinus Aestuarii (Crustacea, Decapoda). Mar. Environ. Res. 2011, 71, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Van Horn, J.; Malhoe, V.; Delvina, M.; Thies, M.; Tolley, S.G.; Ueda, T. Molecular Cloning and Expression of a 2-Cys Peroxiredoxin Gene in the Crustacean Eurypanopeus Depressus Induced by Acute Hypo-Osmotic Stress. Comp. Biochem. Physiol.—B Biochem. Mol. Biol. 2010, 155, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Pan, L.; Xu, L.; Si, L. Effects of Ammonia-N Exposure on the Concentrations of Neurotransmitters, Hemocyte Intracellular Signaling Pathways and Immune Responses in White Shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2018, 75, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Pan, L.; Ren, Q.; Wang, L.; Miao, J. Effect of Salinity on Regulation Mechanism of Neuroendocrine-Immunoregulatory Network in Litopenaeus vannamei. Fish Shellfish Immunol. 2016, 49, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Sun, W.W.; Tan, B.P.; Chi, S.Y.; Dong, X.H.; Yang, Q.H. Molecular Cloning and Expression of Hepatopancreas Glutamine Synthetase in the Pacific White Shrimp, Litopenaeus vannamei, Induced by Acute Hypo-Osmotic Stress. Aquaculture 2012, 362–363, 80–87. [Google Scholar] [CrossRef]
- Chang, E.S. Stressed-Out Lobsters: Crustacean Hyperglycemic Hormone and Stress Proteins. Integr. Comp. Biol. 2005, 45, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Xie, J.; Huang, M.; Chen, C.; Qian, D.; Qin, J.G.; Chen, L.; Jia, Y.; Li, E. Growth and Health Responses to a Long-Term PH Stress in Pacific White Shrimp Litopenaeus vannamei. Aquac. Rep. 2020, 16, 100280. [Google Scholar] [CrossRef]
- Wang, W.N.; Li, B.S.; Liu, J.J.; Shi, L.; Alam, M.J.; Su, S.J.; Wu, J.; Wang, L.; Wang, A.L. The Respiratory Burst Activity and Expression of Catalase in White Shrimp, Litopenaeus vannamei, during Long-Term Exposure to PH Stress. Ecotoxicology 2012, 21, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Tayag, C.M.; Huang, C.L.; Tsui, W.C.; Chen, J.C. White Shrimp Litopenaeus vannamei That Had Received the Hot-Water Extract of Spirulina Platensis Showed Earlier Recovery in Immunity and up-Regulation of Gene Expressions after PH Stress. Fish Shellfish Immunol. 2010, 29, 1092–1098. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Liu, J.; Cao, F.; Li, Z.; Chen, T. Transcriptomic Analyses of the Acute Ammonia Stress Response in the Hepatopancreas of the Kuruma shrimp (Marsupenaeus japonicus). Aquaculture 2019, 513, 734328. [Google Scholar] [CrossRef]
- Pinto, M.R.; Lucena, M.N.; Faleiros, R.O.; Almeida, E.A.; McNamara, J.C.; Leone, F.A. Effects of Ammonia Stress in the Amazon River Shrimp Macrobrachium amazonicum (Decapoda, Palaemonidae). Aquat. Toxicol. 2016, 170, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yu, Y.; Gao, Y.; Yin, P.; Tian, L.; Niu, J.; Liu, Y. Exposure to Acute Ammonia Stress Influences Survival, Immune Response and Antioxidant Status of Pacific White Shrimp (Litopenaeus vannamei) Pretreated with Diverse Levels of Inositol. Fish Shellfish Immunol. 2019, 89, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Romano, N.; Zeng, C. Ontogenetic Changes in Tolerance to Acute Ammonia Exposure and Associated Gill Histological Alterations during Early Juvenile Development of the Blue Swimmer Crab, Portunus Pelagicus. Aquaculture 2007, 266, 246–254. [Google Scholar] [CrossRef]
- Ren, Q.; Pan, L. Digital Gene Expression Analysis in the Gills of the Swimming Crab (Portunus trituberculatus) Exposed to Elevated Ambient Ammonia-N. Aquaculture 2014, 434, 108–114. [Google Scholar] [CrossRef]
- Duan, Y.; Wang, Y.; Huang, J.; Li, H.; Dong, H.; Zhang, J. Toxic Effects of Cadmium and Lead Exposure on Intestinal Histology, Oxidative Stress Response, and Microbial Community of Pacific White Shrimp Litopenaeus vannamei. Mar. Pollut. Bull. 2021, 167, 112220. [Google Scholar] [CrossRef] [PubMed]
- Wendelaar Bonga, S.E. The Stress Response in Fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef] [PubMed]
- Schreck, C.B.; Tort, L. The Concept of Stress in Fish. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 1–34. [Google Scholar]
- Eissa, N.; Wang, H.P. Transcriptional Stress Responses to Environmental and Husbandry Stressors in Aquaculture Species. Rev. Aquac. 2016, 8, 61–88. [Google Scholar] [CrossRef]
- Barton, B.A. Stress in Fishes: A Diversity of Responses with Particular Reference to Changes in Circulating Corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P. Stressors, Stress, and Neuroendocrine Integration of the Adaptive Response The 1997 Hans Selye Memorial Lecture. Ann. N. Y. Acad. Sci. 1998, 851, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Manfrin, C.; Pallavicini, A.; Battistella, S.; Lorenzon, S.; Giulianini, P.G. Crustacean Immunity: The Modulation of Stress Responses. In Lessons in Immunity: From Single-Cell Organisms to Mammals; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 107–116. ISBN 9780128032527. [Google Scholar]
- Balasch, J.C.; Tort, L. Netting the Stress Responses in Fish. Front. Endocrinol. 2019, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Tort, L. Stress and Immune Modulation in Fish. Dev. Comp. Immunol. 2011, 35, 1366–1375. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy Homeostasis as an Integrative Tool for Assessing Limits of Environmental Stress Tolerance in Aquatic Invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Adamo, S.A. The Effects of the Stress Response on Immune Function in Invertebrates: An Evolutionary Perspective on an Ancient Connection. Horm. Behav. 2012, 62, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, R. Reality as the Leading Cause of Stress: Rethinking the Impact of Chronic Stress in Nature. Funct. Ecol. 2013, 27, 11–23. [Google Scholar] [CrossRef]
- Toni, M. Variation in Environmental Parameters in Research and Aquaculture: Effects on Behaviour, Physiology and Cell Biology of Teleost Fish. J. Aquac. Mar. Biol. 2017, 5, 1–11. [Google Scholar] [CrossRef]
- Lara, R.A.; Vasconcelos, R.O. Impact of Noise on Development, Physiological Stress and Behavioural Patterns in Larval Zebrafish. Sci. Rep. 2021, 11, 6615. [Google Scholar] [CrossRef] [PubMed]
- Tong, R.; Pan, L.; Zhang, X.; Li, Y. Neuroendocrine-Immune Regulation Mechanism in Crustaceans: A Review. Rev. Aquac. 2022, 14, 378–398. [Google Scholar] [CrossRef]
- Ciji, A.; Akhtar, M.S. Stress Management in Aquaculture: A Review of Dietary Interventions. Rev. Aquac. 2021, 13, 2190–2247. [Google Scholar] [CrossRef]
- von Weissenberg, E.; Jansson, A.; Vuori, K.A.; Engström-Öst, J. Copepod Reproductive Effort and Oxidative Status as Responses to Warming in the Marine Environment. Ecol. Evol. 2022, 12, e8594. [Google Scholar] [CrossRef] [PubMed]
- von Weissenberg, E.; Mottola, G.; Uurasmaa, T.M.; Anttila, K.; Engström-Öst, J. Combined Effect of Salinity and Temperature on Copepod Reproduction and Oxidative Stress in Brackish-Water Environment. Front. Mar. Sci. 2022, 9, 952863. [Google Scholar] [CrossRef]
- She, Q.; Han, Z.; Liang, S.; Xu, W.; Li, X.; Zhao, Y.; Wei, H.; Dong, J.; Li, Y. Impacts of Circadian Rhythm and Melatonin on the Specific Activities of Immune and Antioxidant Enzymes of the Chinese Mitten Crab (Eriocheir sinensis). Fish Shellfish Immunol. 2019, 89, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, K.; Liu, L.; Tanase, C.; Mols, R.; van der Meer, M. Effects of Light Intensity and Photoperiod on the Growth and Stress Response of Juvenile Nile Tilapia (Oreochromis niloticus) in a Recirculating Aquaculture System. Aquac. Fish. 2023, 8, 85–90. [Google Scholar] [CrossRef]
- Zhang, B.; Yu, C.; Xu, Y.; Huang, Z.; Cai, Y.; Li, Y. Hepatopancreas Immune Response during Different Photoperiods in the Chinese Mitten Crab, Eriocheir sinensis. Fish Shellfish Immunol. 2023, 132, 108482. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Rochow, V.B. The Crustacean Eye: Dark/ Light Adaptation, Polarization Sensitivity, Flicker Fusion Frequency, and Photoreceptor Damage. Zool. Sci. 2001, 18, 1175–1197. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Migaud, H.; Shi, C.; Song, C.; Wang, C.; Ye, Y.; Ren, Z.; Wang, H.; Mu, C. Light Intensity Impacts on Growth, Molting and Oxidative Stress of Juvenile Mud Crab Scylla paramamosain. Aquaculture 2021, 545, 737159. [Google Scholar] [CrossRef]
- Xu, H.; Dou, J.; Wu, Q.; Ye, Y.; Song, C.; Mu, C.; Wang, C.; Ren, Z.; Shi, C. Investigation of the Light Intensity Effect on Growth, Molting, Hemolymph Lipid, and Antioxidant Capacity of Juvenile Swimming Crab Portunus trituberculatus. Front. Mar. Sci. 2022, 9, 922021. [Google Scholar] [CrossRef]
- Cartolano, M.C.; Berenshtein, I.; Heuer, R.M.; Pasparakis, C.; Rider, M.; Hammerschlag, N.; Paris, C.B.; Grosell, M.; McDonald, M.D. Impacts of a Local Music Festival on Fish Stress Hormone Levels and the Adjacent Underwater Soundscape. Environ. Pollut. 2020, 265, 114925. [Google Scholar] [CrossRef] [PubMed]
- Slater, M.; Fricke, E.; Weiss, M.; Rebelein, A.; Bögner, M.; Preece, M.; Radford, C. The Impact of Aquaculture Soundscapes on Whiteleg Shrimp Litopenaeus vannamei and Atlantic Salmon Salmo Salar. Aquac. Environ. Interact. 2020, 12, 167–177. [Google Scholar] [CrossRef]
- Kathyayani, S.A.; Muralidhar, M.; Kumar, T.S.; Alavandi, S.V. Stress Quantification in Penaeus vannamei Exposed to Varying Levels of Turbidity. J. Coast. Res. 2019, 86, 177. [Google Scholar] [CrossRef]
- Li, M.; Wang, X.; Qi, C.; Li, E.; Du, Z.; Qin, J.G.; Chen, L. Metabolic Response of Nile Tilapia (Oreochromis niloticus) to Acute and Chronic Hypoxia Stress. Aquaculture 2018, 495, 187–195. [Google Scholar] [CrossRef]
- Jia, R.; Liu, B.L.; Han, C.; Huang, B.; Lei, J.L. Effects of Ammonia Exposure on Stress and Immune Response in Juvenile Turbot (Scophthalmus maximus). Aquac. Res. 2017, 48, 3149–3162. [Google Scholar] [CrossRef]
- Cheng, C.H.; Su, Y.L.; Ma, H.L.; Deng, Y.Q.; Feng, J.; Chen, X.L.; Jie, Y.K.; Guo, Z.X. Effect of Nitrite Exposure on Oxidative Stress, DNA Damage and Apoptosis in Mud Crab (Scylla paramamosain). Chemosphere 2020, 239, 124668. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, H.; Guo, M.; Fang, D.; Mei, J.; Xie, J. Analysis of Acute Nitrite Exposure on Physiological Stress Response, Oxidative Stress, Gill Tissue Morphology and Immune Response of Large Yellow Croaker (Larimichthys crocea). Animals 2022, 12, 1791. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, D.D.; Smith, S.A.; Boardman, G.D.; Angier, M.W.; Marsh, L.; Flick, G.J. Chronic Toxicity of Nitrate to Pacific White Shrimp, Litopenaeus vannamei: Impacts on Survival, Growth, Antennae Length, and Pathology. Aquaculture 2010, 309, 109–114. [Google Scholar] [CrossRef]
- Yu, J.; Xiao, Y.; Wang, Y.; Xu, S.; Zhou, L.; Li, J.; Li, X. Chronic Nitrate Exposure Cause Alteration of Blood Physiological Parameters, Redox Status and Apoptosis of Juvenile Turbot (Scophthalmus maximus). Environ. Pollut. 2021, 283, 117103. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Wang, J.; Li, J.; Li, J. Effect of High Alkalinity on Shrimp Gills: Histopathological Alternations and Cell Specific Responses. Ecotoxicol. Environ. Saf. 2023, 256, 114902. [Google Scholar] [CrossRef] [PubMed]
- Furtado, P.S.; Gaona, C.A.P.; Serra, F.P.; Poersch, L.H.; Wasielesky, W. Acute Toxicity of Carbon Dioxide to Juvenile Marine Shrimp Litopenaeus vannamei (Boone 1931). Mar. Freshw. Behav. Physiol. 2017, 50, 293–301. [Google Scholar] [CrossRef]
- Xu, C.; Li, E.; Liu, Y.; Wang, X.; Qin, J.G.; Chen, L. Comparative Proteome Analysis of the Hepatopancreas from the Pacific White Shrimp Litopenaeus vannamei under Long-Term Low Salinity Stress. J. Proteom. 2017, 162, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ye, Y.; Li, W.; Liu, X.; Zhao, Y.; Jiang, Q.; Che, X. Effects of Salinity Stress on Histological Changes, Glucose Metabolism Index and Transcriptomic Profile in Freshwater Shrimp, Macrobrachium nipponense. Animals 2023, 13, 2884. [Google Scholar] [CrossRef] [PubMed]
- Giffard-Mena, I.; Ponce-Rivas, E.; Sigala-Andrade, H.M.; Uranga-Solís, C.; Re, A.D.; Díaz, F.; Camacho-Jiménez, L. Evaluation of the Osmoregulatory Capacity and Three Stress Biomarkers in White Shrimp Penaeus vannamei Exposed to Different Temperature and Salinity Conditions: Na+/K+ ATPase, Heat Shock Proteins (HSP), and Crustacean Hyperglycemic Hormones (CHHs). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2024, 271, 110942. [Google Scholar] [CrossRef] [PubMed]
- Jaffer, Y.D.; Bhat, I.A.; Mir, I.N.; Bhat, R.A.H.; Sidiq, M.J.; Jana, P. Adaptation of Cultured Decapod Crustaceans to Changing Salinities: Physiological Responses, Molecular Mechanisms and Disease Implications. Rev. Aquac. 2024, 16, 1520–1543. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, C.; Chen, K.; Liu, Y.; Deng, Y.; Liu, W.; Zhang, D.; Jiang, G.; Li, X.; Giri, S.S.; et al. Dietary T-2 Toxin Induces Transcriptomic Changes in Hepatopancreas of Chinese Mitten Crab (Eriocheir sinensis) via Nutrition Metabolism and Apoptosis-Related Pathways. Ecotoxicol. Environ. Saf. 2023, 249, 114397. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Wei, H.; Zhang, Y.; Su, J. Transcriptome Response of a Marine Copepod in Response to Environmentally-Relevant Concentrations of Saxitoxin. Mar. Pollut. Bull. 2024, 205, 116546. [Google Scholar] [CrossRef] [PubMed]
- Ruvalcaba-Márquez, J.C.; Álvarez-Ruíz, P.; Zenteno-Savín, T.; Martínez-Antonio, E.; Goytortúa-Bores, E.; Casillas-Hernández, R.; Mejía-Ruíz, H.; Magallón-Barajas, F.J. Performance, Immune Response, and Oxidative Stress Parameters of Litopenaeus vannamei Fed Diets Containing Varying Carbohydrate/Protein, Lipid/Protein, and Energy/Protein Ratios. Aquac. Rep. 2021, 21, 100771. [Google Scholar] [CrossRef]
- Tarifeño-Saldivia, E.; Aguilar, A.; Contreras, D.; Mercado, L.; Morales-Lange, B.; Márquez, K.; Henríquez, A.; Riquelme-Vidal, C.; Boltana, S. Iron Overload Is Associated with Oxidative Stress and Nutritional Immunity during Viral Infection in Fish. Front. Immunol. 2018, 9, 1296. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, Z.; Xu, W.; Li, X.; Zhao, Y.; Wei, H.; Li, X.; Chen, Q. Antioxidant and Immune Responses of the Oriental River Prawn Macrobrachium Nipponense to the Isopod Parasite Tachaea chinensis. Fish Shellfish Immunol. 2020, 101, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Briceño, F.A.; Polymeropoulos, E.T.; Fitzgibbon, Q.P.; Dambacher, J.M.; Pecl, G.T. Changes in Metabolic Rate of Spiny Lobster under Predation Risk. Mar. Ecol. Prog. Ser. 2018, 598, 71–84. [Google Scholar] [CrossRef]
- Xie, Y.X.; Yang, X.M.; Kaneko, G.; Liang, J.N.; Wen, L.T.; Li, Y.J.; Ao, Q.W.; Huang, L.M.; Li, P.; Min, W.W.; et al. Effects of Different Stocking Densities and Feeding Frequencies on Growth, Physiological and Biochemical Indexes, and Intestinal Microflora of Largemouth Bass (Micropterus salmoides) under Land-Based Round Pond. Aquaculture 2024, 580, 740385. [Google Scholar] [CrossRef]
- Pandey, A.; Pathan, M.A.; Sudhagar, S.A.; Krishnani, K.K.; Sreedharan, K.; Prakash, S.; Jana, P. Influence of Crowding Density Mediated Stress on Haematological, Biochemical Indices and Molecular Changes of Penaeus vannamei Reared in Inland Saline Water (ISW) Sourced Earthen Ponds. Aquac. Int. 2024, 32, 6287–6302. [Google Scholar] [CrossRef]
- Mercier, L.; Palacios, E.; Campa-Córdova, Á.I.; Tovar-Ramírez, D.; Hernández-Herrera, R.; Racotta, I.S. Metabolic and Immune Responses in Pacific Whiteleg Shrimp Litopenaeus vannamei Exposed to a Repeated Handling Stress. Aquaculture 2006, 258, 633–640. [Google Scholar] [CrossRef]
- Aparicio-Simón, B.; Piñón, M.; Racotta, R.; Racotta, I.S. Neuroendocrine and Metabolic Responses of Pacific Whiteleg Shrimp Litopenaeus vannamei Exposed to Acute Handling Stress. Aquaculture 2010, 298, 308–314. [Google Scholar] [CrossRef]
- Nilsson, B.; Jepsen, P.M.; Bucklin, A.; Hansen, B.W. Environmental Stress Responses and Experimental Handling Artifacts of a Model Organism, the Copepod Acartia tonsa (Dana). Front. Mar. Sci. 2018, 5, 156. [Google Scholar] [CrossRef]
- Mugnier, C.; Lemonnier, H.; Legrand, A. Physiological Response of the Blue Shrimp Litopenaeus stylirostris to Short-Term Confinement on a Pond Bottom. Aquaculture 2006, 253, 703–711. [Google Scholar] [CrossRef]
- Guan, W.; Nong, W.; Wei, X.; Zhu, M.; Mao, L. Impacts of a Novel Live Shrimp (Litopenaeus vannamei) Water-Free Transportation Strategy on Flesh Quality: Insights through Stress Response and Oxidation in Lipids and Proteins. Aquaculture 2021, 533, 736168. [Google Scholar] [CrossRef]
- Nie, X.; Chen, Y.; Xu, Z.; Chi, Y.; Zhu, C.; Huang, P.H.; Bai, Q.; Chen, X. Physiological Responses and Quality Assessment of Turbot (Scophthalmus maximus) during Waterless Preservation and Transportation: Implications for Survival and Biochemical Indicators. Aquaculture 2024, 580, 740315. [Google Scholar] [CrossRef]
- González-Vera, C.; Brown, J.H. Effects of Alkalinity and Total Hardness on Growth and Survival of Postlarvae Freshwater Prawns, Macrobrachium rosenbergii (De Man 1879). Aquaculture 2017, 473, 521–527. [Google Scholar] [CrossRef]
- Bu, R.; Wang, P.; Zhao, C.; Bao, W.; Qiu, L. Gene Characteristics, Immune and Stress Responses of PmPrx1 in Black Tiger Shrimp (Penaeus monodon): Insights from Exposure to Pathogenic Bacteria and Toxic Environmental Stressors. Dev. Comp. Immunol. 2017, 77, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sha, H.; Song, T.; Zhan, P.; Zhang, X.; Chen, J.; Xiong, J. Sarafloxacin Hydrochloride Exposure Disrupts Gut Microbiota and Increases Shrimp Susceptibility to Vibrio anguillarum Infection. Aquaculture 2024, 586, 740810. [Google Scholar] [CrossRef]
- Wan, J.J.; Shen, M.F.; Xue, H.; Liu, H.Y.; Zhang, M.Q.; Zhu, X.H.; Wang, C.H. Effects of Feeding Frequency on Growth Performance, Antioxidant Status and Disease Resistance of Juvenile Red Swamp Crayfish, Procambarus clarkia. Pak. J. Zool. 2021, 53, 467–474. [Google Scholar] [CrossRef]
- Winberg, S.; Höglund, E.; Øverli, Ø. Variation in the Neuroendocrine Stress Response. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 35–74. [Google Scholar]
- Selye, H. Stress and The General Adaptation Syndrome. Br. Med. J. 1950, 1, 1383. [Google Scholar] [CrossRef] [PubMed]
- Urbinati, E.C.; Zanuzzo, F.S.; Biller, J.D. Stress and Immune System in Fish. In Biology and Physiology of Freshwater Neotropical Fish; Elsevier: Amsterdam, The Netherlands, 2019; pp. 93–114. ISBN 9780128158722. [Google Scholar]
- Elwood, R.W.; Barr, S.; Patterson, L. Pain and Stress in Crustaceans? Appl. Anim. Behav. Sci. 2009, 118, 128–136. [Google Scholar] [CrossRef]
- Cinel, S.D.; Hahn, D.A.; Kawahara, A.Y. Predator-Induced Stress Responses in Insects: A Review. J. Insect Physiol. 2020, 122, 104039. [Google Scholar] [CrossRef] [PubMed]
- Lorenzon, S. Hyperglycemic Stress Response in Crustacea. ISJ-2 Invertebr. Surviv. J. 2005, 2, 132–141. [Google Scholar]
- Simões, L.A.R.; Normann, R.S.; Chung, J.S.; Vinagre, A.S. A Brief and Updated Introduction to the Neuroendocrine System of Crustaceans. Mol. Cell Endocrinol. 2024, 590, 112265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Pan, L.; Wei, C.; Tong, R.; Li, Y.; Ding, M.; Wang, H. Crustacean Hyperglycemic Hormone (CHH) Regulates the Ammonia Excretion and Metabolism in White Shrimp, Litopenaeus Vannamei under Ammonia-N Stress. Sci. Total Environ. 2020, 723, 138128. [Google Scholar] [CrossRef] [PubMed]
- Miller, S. The Precise Release Time of Crustacean Hyperglycaemic Hormone (CHH) in Response to Environmental Stressors in the Shore Crab Carcinus maenas. Master’s Thesis, Bangor University, Wales, UK, 2023. [Google Scholar]
- Turner, L.M.; Webster, S.G.; Morris, S. Roles of Crustacean Hyperglycaemic Hormone in Ionic and Metabolic Homeostasis in the Christmas Island Blue Crab, Discoplax celeste. J. Exp. Biol. 2013, 216, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, M.; Flik, G. The Endocrinology of the Stress Response in Fish: An Adaptation-Physiological View. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 75–111. [Google Scholar]
- Conde-Sieira, M.; Chivite, M.; Míguez, J.M.; Soengas, J.L. Stress Effects on the Mechanisms Regulating Appetite in Teleost Fish. Front. Endocrinol. 2018, 9, 631. [Google Scholar] [CrossRef] [PubMed]
- Martorell-Ribera, J.; Koczan, D.; Tindara Venuto, M.; Viergutz, T.; Brunner, R.M.; Goldammer, T.; Gimsa, U.; Rebl, A. Experimental Handling Challenges Result in Minor Changes in the Phagocytic Capacity and Transcriptome of Head-Kidney Cells of the Salmonid Fish Coregonus maraena. Front. Vet. Sci. 2022, 9, 889635. [Google Scholar] [CrossRef] [PubMed]
- Faught, E.; Aluru, N.; Vijayan, M.M. The Molecular Stress Response. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 113–166. [Google Scholar]
- Li, Y.; Wei, L.; Cao, J.; Qiu, L.; Jiang, X.; Li, P.; Song, Q.; Zhou, H.; Han, Q.; Diao, X. Oxidative Stress, DNA Damage and Antioxidant Enzyme Activities in the Pacific White Shrimp (Litopenaeus vannamei) When Exposed to Hypoxia and Reoxygenation. Chemosphere 2016, 144, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, A.; Tang, S.; Huang, M.; Yu, Q.; Xu, C.; Li, E. Identification of Key Immune and Stress Related Genes and Pathways by Comparative Analysis of the Gene Expression Profile under Multiple Environmental Stressors in Pacific White Shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2023, 135, 108695. [Google Scholar] [CrossRef] [PubMed]
- Zollinger, S.A.; Goller, F.; Brumm, H. Metabolic and Respiratory Costs of Increasing Song Amplitude in Zebra Finches. PLoS ONE 2011, 6, e23198. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.A.; Keller, R. Effect of Exposure to Atmospheric Air on Blood Glucose and Lactate Concentrations in Two Crustacean Species: A Role of the Crustacean Hyperglycemic Hormone (CHH). Comp. Biochem. Physiol. A Physiol. 1993, 106, 343–347. [Google Scholar] [CrossRef]
- Harris, B.N.; Carr, J.A. The Role of the Hypothalamus-Pituitary-Adrenal/Interrenal Axis in Mediating Predator-Avoidance Trade-Offs. Gen. Comp. Endocrinol. 2016, 230–231, 110–142. [Google Scholar] [CrossRef] [PubMed]
- Sadoul, B.; Vijayan, M.M. Stress and Growth. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 167–205. [Google Scholar]
- Santos, G.A.; Schrama, J.W.; Mamauag, R.E.P.; Rombout, J.H.W.M.; Verreth, J.A.J. Chronic Stress Impairs Performance, Energy Metabolism and Welfare Indicators in European Seabass (Dicentrarchus labrax): The Combined Effects of Fish Crowding and Water Quality Deterioration. Aquaculture 2010, 299, 73–80. [Google Scholar] [CrossRef]
- Sornchuer, P.; Junprung, W.; Yingsunthonwattana, W.; Tassanakajon, A. Heat Shock Factor 1 Regulates Heat Shock Proteins and Immune-Related Genes in Penaeus monodon under Thermal Stress. Dev. Comp. Immunol. 2018, 88, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Mengal, K.; Kor, G.; Kozák, P.; Niksirat, H. Heat Shock Proteins Adaptive Responses to Environmental Stressors and Implications in Health Management of Decapods. Aquac. Rep. 2023, 30, 101564. [Google Scholar] [CrossRef]
- Rahi, M.L.; Mahmud, S.; Dilruba, K.J.; Sabbir, W.; Aziz, D.; Hurwood, D.A. Temperature Induced Changes in Physiological Traits and Expression of Selected Candidate Genes in Black Tiger Shrimp (Penaeus monodon) Larvae. Aquac. Rep. 2021, 19, 100620. [Google Scholar] [CrossRef]
- Sathyanandam, S.; Vasudevan, S.; Natesan, M. Serotonin Modulation of Hemolymph Glucose and Crustacean Hyperglycemic Hormone Titers in Fenneropenaeus indicus. Aquaculture 2008, 281, 106–112. [Google Scholar] [CrossRef]
- Lee, M.C.; Park, J.C.; Lee, J.S. Effects of Environmental Stressors on Lipid Metabolism in Aquatic Invertebrates. Aquat. Toxicol. 2018, 200, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, A.L.; Xian, J.A. Variation of Free Amino Acid and Carbohydrate Concentrations in White Shrimp, Litopenaeus vannamei: Effects of Continuous Cold Stress. Aquaculture 2011, 317, 182–186. [Google Scholar] [CrossRef]
- Zhang, X.; Yuan, J.; Zhang, X.; Yu, Y.; Li, F. Comparative Transcriptomic Analysis Unveils a Network of Energy Reallocation in Litopenaeus vannamei Responsive to Heat-Stress. Ecotoxicol. Environ. Saf. 2022, 238, 113600. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.V.; Alfaro, A.C.; Rodríguez, J.; Bayot, B.; Sonnenholzner, S. Changes in Metabolic Profiling of Whiteleg Shrimp (Penaeus vannamei) under Hypoxic Stress. J. Invertebr. Pathol. 2022, 193, 107798. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhang, X.; Chen, Z.; Guo, X.; Zhou, Q.; Jin, M.; Taj, S.; Zhang, L.; Liu, W.; Jiao, L. Acute Environmental Hypoxia Induced Changes of Tissue Morphology, Antioxidant Capacity, Immune Response and Energy Metabolism in Marsupenaeus japonicus. Aquac. Rep. 2023, 30, 101565. [Google Scholar] [CrossRef]
- Alfaro, A.C.; Nguyen, T.V.; Bayot, B.; Rodriguez Leon, J.A.; Domínguez-Borbor, C.; Sonnenholzner, S. Metabolic Responses of Whiteleg Shrimp to White Spot Syndrome Virus (WSSV). J. Invertebr. Pathol. 2021, 180, 107545. [Google Scholar] [CrossRef] [PubMed]
- Ghemrawi, R.; Khair, M. Endoplasmic Reticulum Stress and Unfolded Protein Response in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 6127. [Google Scholar] [CrossRef] [PubMed]
- Ong, G.; Logue, S.E. Unfolding the Interactions between Endoplasmic Reticulum Stress and Oxidative Stress. Antioxidants 2023, 12, 981. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; He, J.G. Effects of Environmental Stress on Shrimp Innate Immunity and White Spot Syndrome Virus Infection. Fish Shellfish Immunol. 2019, 84, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Murugan, D.D.; Khan, H.; Huang, Y.; Cheang, W.S. Roles and Therapeutic Implications of Endoplasmic Reticulum Stress and Oxidative Stress in Cardiovascular Diseases. Antioxidants 2021, 10, 1167. [Google Scholar] [CrossRef] [PubMed]
- Sirish, P.; Diloretto, D.A.; Thai, P.N.; Chiamvimonvat, N. The Critical Roles of Proteostasis and Endoplasmic Reticulum Stress in Atrial Fibrillation. Front. Physiol. 2022, 12, 793171. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.Y.; MacRae, T.H.; Sorgeloos, P.; Bossier, P. Stress Response for Disease Control in Aquaculture. Rev. Aquac. 2011, 3, 120–137. [Google Scholar] [CrossRef]
- Wang, X.X.; Zhang, H.; Gao, J.; Wang, X.W. Ammonia Stress-Induced Heat Shock Factor 1 Enhances White Spot Syndrome Virus Infection by Targeting the Interferon-like System in Shrimp. mBio 2024, 15, e03136-23. [Google Scholar] [CrossRef] [PubMed]
- Ulaje, S.A.; Lluch-Cota, S.E.; Sicard, M.T.; Ascencio, F.; Cruz-Hernández, P.; Racotta, I.S.; Rojo-Arreola, L. Litopenaeus vannamei Oxygen Consumption and HSP Gene Expression at Cyclic Conditions of Hyperthermia and Hypoxia. J. Therm. Biol. 2020, 92, 102666. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhu, H.; Yuan, J.; Zhang, X.; Xiang, J.; Li, F. Diversity of Heat Shock Proteins in Response to Various Stressors in the Pacific White Shrimp Litopenaeus vannamei. Aquaculture 2024, 584, 740647. [Google Scholar] [CrossRef]
- Lubkowska, A.; Dudzińska, W.; Pluta, W. Antioxidant Enzyme Activity and Serum HSP70 Concentrations in Relation to Insulin Resistance and Lipid Profile in Lean and Overweight Young Men. Antioxidants 2023, 12, 655. [Google Scholar] [CrossRef] [PubMed]
- Hoseinifar, S.H.; Yousefi, S.; Van Doan, H.; Ashouri, G.; Gioacchini, G.; Maradonna, F.; Carnevali, O. Oxidative Stress and Antioxidant Defense in Fish: The Implications of Probiotic, Prebiotic, and Synbiotics. Rev. Fish. Sci. Aquac. 2020, 29, 198–217. [Google Scholar] [CrossRef]
- Song, C.; Sun, C.; Liu, B.; Xu, P. Oxidative Stress in Aquatic Organisms. Antioxidants 2023, 12, 1223. [Google Scholar] [CrossRef] [PubMed]
- Takei, Y.; Hwang, P.P. Homeostatic Responses to Osmotic Stress. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 207–249. [Google Scholar]
- Rivera-Ingraham, G.A.; Lignot, J.-H. Osmoregulation, Bioenergetics and Oxidative Stress in Coastal Marine Invertebrates: Raising the Questions for Future Research. J. Exp. Biol. 2017, 220, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- Bal, A.; Panda, F.; Pati, S.G.; Das, K.; Agrawal, P.K.; Paital, B. Modulation of Physiological Oxidative Stress and Antioxidant Status by Abiotic Factors Especially Salinity in Aquatic Organisms. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 241, 108971. [Google Scholar] [CrossRef] [PubMed]
- Blenau, W.; Baumann, A. Molecular and Pharmacological Properties of Insect Biogenic Amine Receptors: Lessons from Drosophila melanogaster and Apis mellifera; Wiley: Hoboken, NJ, USA, 2001; Volume 48. [Google Scholar]
- Caballero-Herrero, M.J.; Jumilla, E.; Buitrago-Ruiz, M.; Valero-Navarro, G.; Cuevas, S. Role of Damage-Associated Molecular Patterns (DAMPS) in the Postoperative Period after Colorectal Surgery. Int. J. Mol. Sci. 2023, 24, 3862. [Google Scholar] [CrossRef] [PubMed]
- Wanvimonsuk, S.; Somboonwiwat, K. Peroxiredoxin-4 Supplementation Modulates the Immune Response, Shapes the Intestinal Microbiome, and Enhances AHPND Resistance in Penaeus Vannamei. Fish Shellfish Immunol. 2023, 139, 108915. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Li, S.; Tang, D.; Peng, Y.; Meng, J.; Peng, S.; Deng, Z.; Qiu, S.; Liao, X.; Chen, H.; et al. Circulating Peroxiredoxin-1 Is a Novel Damage-Associated Molecular Pattern and Aggravates Acute Liver Injury via Promoting Inflammation. Free Radic. Biol. Med. 2019, 137, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Adamo, S.A. Stress Responses Sculpt the Insect Immune System, Optimizing Defense in an Ever-Changing World. Dev. Comp. Immunol. 2017, 66, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Yada, T.; Tort, L. Stress and Disease Resistance: Immune System and Immunoendocrine Interactions. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 365–403. [Google Scholar]
- Pankhurst, N.W. Reproduction and Development. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 295–331. [Google Scholar]
- Shirly-Lim, Y.L.; Rahmah, S.; Ghaffar, M.A.; Liang, L.Q.; Chang, Y.M.; Chisti, Y.; Lee, M.A.; Liew, H.J. Pacific Whiteleg Shrimps Compromize Their Physiological Needs to Cope with Environmental Stress. Environ. Adv. 2024, 15, 100492. [Google Scholar] [CrossRef]
- Yuslan, A.; Najuwa, S.; Hagiwara, A.; Ghaffar, M.A.; Suhaimi, H.; Rasdi, N.W. Production Performance of Moina macrocopa (Straus 1820) (Crustacea, Cladocera) Cultured in Different Salinities: The Effect on Growth, Survival, Reproduction, and Fatty Acid Composition of the Neonates. Diversity 2021, 13, 105. [Google Scholar] [CrossRef]
- Engström-Öst, J.; Glippa, O.; Feely, R.A.; Kanerva, M.; Keister, J.E.; Alin, S.R.; Carter, B.R.; McLaskey, A.K.; Vuori, K.A.; Bednaršek, N. Eco-Physiological Responses of Copepods and Pteropods to Ocean Warming and Acidification. Sci. Rep. 2019, 9, 4748. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.; Postel, U.; Mrinalini; Turner, L.M.; Palmer, J.; Webster, S.G. The Adaptive Significance of Crustacean Hyperglycaemic Hormone (CHH) in Daily and Seasonal Migratory Activities of the Christmas Island Red Crab Gecarcoidea natalis. J. Exp. Biol. 2010, 213, 3062–3073. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Pan, L.; Tong, R.; Li, Y.; Si, L.; Chen, Y.; Li, D. The Exploration of Neuroendocrine Regulation of Crustacean Hyperglycemic Hormone (CHH) on Innate Immunity of Litopenaeus vannamei under Ammonia-N Stress. Mol. Immunol. 2021, 139, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Pan, L.; Tong, R.; Li, Y.; Si, L.; Chen, Y.; Wu, M.; Wang, Q. Effects of Crustacean Hyperglycaemic Hormone RNA Interference on Regulation of Glucose Metabolism in Litopenaeus vannamei after Ammonia-Nitrogen Exposure. Br. J. Nutr. 2022, 127, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Yang, P.F.; Zou, H.S. Serotonergic Regulation of Crustacean Hyperglycemic Hormone Secretion in the Crayfish, Procambarus clarkii. Physiol. Biochem. Zool. 2001, 74, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, G.; Li, L. Targeted Top-Down Mass Spectrometry for the Characterization and Tissue-Specific Functional Discovery of Crustacean Hyperglycemic Hormones (CHH) and CHH Precursor-Related Peptides in Response to Low PH Stress. J. Am. Soc. Mass. Spectrom. 2021, 32, 1352–1360. [Google Scholar] [CrossRef] [PubMed]
- Raposo de Magalhães, C.S.F.; Cerqueira, M.A.C.; Schrama, D.; Moreira, M.J.V.; Boonanuntanasarn, S.; Rodrigues, P.M.L. A Proteomics and Other Omics Approach in the Context of Farmed Fish Welfare and Biomarker Discovery. Rev. Aquac. 2020, 12, 122–144. [Google Scholar] [CrossRef]
- Sopinka, N.M.; Donaldson, M.R.; O’Connor, C.M.; Suski, C.D.; Cooke, S.J. Stress Indicators in Fish. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 405–462. [Google Scholar]
- Graham, A.M.; Barreto, F.S.; Crandall, K. Independent Losses of the Hypoxia-Inducible Factor (HIF) Pathway within Crustacea. Mol. Biol. Evol. 2020, 37, 1342–1349. [Google Scholar] [CrossRef] [PubMed]
- Terova, G.; Rimoldi, S.; Corà, S.; Bernardini, G.; Gornati, R.; Saroglia, M. Acute and Chronic Hypoxia Affects HIF-1α MRNA Levels in Sea Bass (Dicentrarchus labrax). Aquaculture 2008, 279, 150–159. [Google Scholar] [CrossRef]
- Terova, G.; Rimoldi, S.; Brambilla, F.; Gornati, R.; Bernardini, G.; Saroglia, M. In Vivo Regulation of GLUT2 MRNA in Sea Bass (Dicentrarchus labrax) in Response to Acute and Chronic Hypoxia. Comp. Biochem. Physiol.—B Biochem. Mol. Biol. 2009, 152, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Xuan, F.; Ge, X.; Fu, H.; Zhu, J.; Zhang, S. Identification of Differentially Expressed Genes in Hepatopancreas of Oriental River Prawn, Macrobrachium nipponense Exposed to Environmental Hypoxia. Gene 2014, 534, 298–306. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.D. Endocrine Control of Osmoregulation in Teleost Fish. Am. Zool. 2001, 41, 781–794. [Google Scholar] [CrossRef]
- Lucu, Č.; Towle, D.W. Na++K+-ATPase in Gills of Aquatic Crustacea. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003, 135, 195–214. [Google Scholar] [CrossRef] [PubMed]
- Huong, D.T.T.; Jasmani, S.; Jayasankar, V.; Wilder, M. Na/K-ATPase Activity and Osmo-Ionic Regulation in Adult Whiteleg Shrimp Litopenaeus vannamei Exposed to Low Salinities. Aquaculture 2010, 304, 88–94. [Google Scholar] [CrossRef]
- Hasenbein, M.; Komoroske, L.M.; Connon, R.E.; Geist, J.; Fangue, N.A. Turbidity and Salinity Affect Feeding Performance and Physiological Stress in the Endangered Delta Smelt. Integr. Comp. Biol. 2013, 53, 620–634. [Google Scholar] [CrossRef] [PubMed]
- Tipsmark, C.K.; Madsen, S.S. Regulation of Na+/K+-ATPase Activity by Nitric Oxide in the Kidney and Gill of the Brown Trout (Salmo trutta). J. Exp. Biol. 2003, 206, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.C.; Hwang, P.P. Some Insights into Energy Metabolism for Osmoregulation in Fish. Comp. Biochem. Physiol.—C Toxicol. Pharmacol. 2008, 148, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.C.H.; Wu, S.M.; Tseng, Y.C.; Lee, Y.C.; Baba, O.; Hwang, P.P. Regulation of Glycogen Metabolism in Gills and Liver of the Euryhaline Tilapia (Oreochromis mossambicus) during Acclimation to Seawater. J. Exp. Biol. 2007, 210, 3494–3504. [Google Scholar] [CrossRef] [PubMed]
- Szyller, J.; Bil-Lula, I. Heat Shock Proteins in Oxidative Stress and Ischemia/Reperfusion Injury and Benefits from Physical Exercises: A Review to the Current Knowledge. Oxid. Med. Cell Longev. 2021, 2021, 6678457. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Qu, Y.; Zhuo, X.; Li, J.; Zou, J.; Fan, L. Investigating the Physiological Responses of Pacific White Shrimp Litopenaeus vannamei to Acute Cold-Stress. PeerJ 2019, 7, e7381. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.F.; Luo, Z.; Huang, C.; Chen, Q.L.; Pan, Y.X.; Xu, Y.H. Endoplasmic Reticulum Stress-Related Genes in Yellow Catfish Pelteobagrus fulvidraco: Molecular Characterization, Tissue Expression, and Expression Responses to Dietary Copper Deficiency and Excess. G3 Genes Genomes Genet. 2015, 5, 2091–2104. [Google Scholar] [CrossRef] [PubMed]
- Jeyachandran, S.; Chellapandian, H.; Park, K.; Kwak, I.-S. A Review on the Involvement of Heat Shock Proteins (Extrinsic Chaperones) in Response to Stress Conditions in Aquatic Organisms. Antioxidants 2023, 12, 1444. [Google Scholar] [CrossRef]
- Janewanthanakul, S.; Supungul, P.; Tang, S.; Tassanakajon, A. Heat Shock Protein 70 from Litopenaeus vannamei (LvHSP70) Is Involved in the Innate Immune Response against White Spot Syndrome Virus (WSSV) Infection. Dev. Comp. Immunol. 2020, 102, 103476. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Lin, T.; Li, H.; Lu, T.; Li, J.; Huang, W.; Sun, H.; Jiang, X.; Zhang, J.; Yan, A.; et al. Heat Shock Protein 40 (HSP40) in Pacific White Shrimp (Litopenaeus vannamei): Molecular Cloning, Tissue Distribution and Ontogeny, Response to Temperature, Acidity/Alkalinity and Salinity Stresses, and Potential Role in Ovarian Development. Front. Physiol. 2018, 9, 1784. [Google Scholar] [CrossRef] [PubMed]
- Terrab, L.; Wipf, P. Hsp70 and the Unfolded Protein Response as a Challenging Drug Target and an Inspiration for Probe Molecule Development. ACS Med. Chem. Lett. 2020, 11, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Pretto, A.; Loro, V.L.; Morsch, V.M.; Moraes, B.S.; Menezes, C.; Santi, A.; Toni, C. Alterations in Carbohydrate and Protein Metabolism in Silver Catfish (Rhamdia quelen) Exposed to Cadmium. Ecotoxicol. Environ. Saf. 2014, 100, 188–192. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Qiang, J.; Yang, H.; Xu, P.; Zhu, Z.X.; Yang, R.Q. Changes in the Fatty Acid Composition and Regulation of Antioxidant Enzymes and Physiology of Juvenile Genetically Improved Farmed Tilapia Oreochromis niloticus (L.), Subjected to Short-Term Low Temperature Stress. J. Therm. Biol. 2015, 53, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Conneely, E.A.; Coates, C.J. Meta-Analytic Assessment of Physiological Markers for Decapod Crustacean Welfare. Fish Fish. 2024, 25, 134–150. [Google Scholar] [CrossRef]
- Li, X.; Xu, B.; Shen, P.; Cheng, H.; Fan, Y.; Gao, Q. Regulation and Response Mechanism of Acute Low-Salinity Stress during Larval Stages in Macrobrachium rosenbergii Based on Multi-Omics Analysis. Int. J. Mol. Sci. 2024, 25, 6809. [Google Scholar] [CrossRef] [PubMed]
- Kozłowski, M.; Piotrowska, I. Effect of Stocking Density on Growth, Survival and Cannibalism of Juvenile Pikeperch, Sander Lucioperca (L.), in a Recirculating Aquaculture System. Aquac. Int. 2024, 32, 3587–3595. [Google Scholar] [CrossRef]
- Zidni, I.; Yustiati, A.; Iskandar, I.; Andriani, Y. The Effect of Modified Aquaculture System on Water Quality in Cultivation of Catfish (Pangasius Hypophthalmus. J. Perikan. Dan Kelaut. 2017, 7, 125. [Google Scholar] [CrossRef]
- Nguyen, T.M.; Ha, P.T.; Le, T.T.H.; Phan, K.S.; Le, T.N.C.; Mai, T.T.T.; Hoang, P.H. Modification of Expanded Clay Carrier for Enhancing the Immobilization and Nitrogen Removal Capacity of Nitrifying and Denitrifying Bacteria in the Aquaculture System. J. Biosci. Bioeng. 2022, 134, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Kucharczyk, D.; Jaczewski, J.; Nowosad, J.; Łuczyński, M.K.; Piech, P.; Dietrich, G.; Kucharczyk, D.J.; Drężek, J.; Kotsyumbas, I.Y.; Muzyka, W.P.; et al. Artificial Reproduction of the Indoor-Cultured Brackish Form of Maraena Whitefish (Coregonus Maraena) under Recirculated Aquaculture System (RAS) Conditions. Anim. Reprod. Sci. 2023, 257, 107329. [Google Scholar] [CrossRef] [PubMed]
- Regan, T.; Bean, T.P.; Ellis, T.; Davie, A.; Carboni, S.; Migaud, H.; Houston, R.D. Genetic Improvement Technologies to Support the Sustainable Growth of UK Aquaculture. Rev. Aquac. 2021, 13, 1958–1985. [Google Scholar] [CrossRef]
- Kashyap, N.; Meher, P.K.; Eswaran, S.; Kathirvelpandian, A.; Udit, U.K.; Ramasre, J.R.; Vaishnav, A.; Chandravanshi, S.; Dhruve, D.; Lal, J. A Review on Genetic Improvement in Aquaculture through Selective Breeding. J. Adv. Biol. Biotechnol. 2024, 27, 618–631. [Google Scholar] [CrossRef]
- Anastasiadi, D.; Díaz, N.; Piferrer, F. Small Ocean Temperature Increases Elicit Stage-Dependent Changes in DNA Methylation and Gene Expression in a Fish, the European Sea Bass. Sci. Rep. 2017, 7, 12401. [Google Scholar] [CrossRef] [PubMed]
- Fellous, A.; Wegner, K.M.; John, U.; Mark, F.C.; Shama, L.N.S. Windows of Opportunity: Ocean Warming Shapes Temperature-Sensitive Epigenetic Reprogramming and Gene Expression across Gametogenesis and Embryogenesis in Marine Stickleback. Glob. Change Biol. 2022, 28, 54–71. [Google Scholar] [CrossRef] [PubMed]
- Metzger, D.C.H.; Schulte, P.M. Persistent and Plastic Effects of Temperature on Dna Methylation across the Genome of Threespine Stickleback (Gasterosteus aculeatus). Proc. R. Soc. B Biol. Sci. 2017, 284, 20171667. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, C.W. Aquatic Animal Nutrition Organic Macro-and Micro-Nutrients; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Johnston, W. Effect of Dietary Tryptophan on Plasma and Brain Tryptophan, Brain Serotonin, and Brain 5-Hydroxyindoleacetic Acid in Rainbow Trout. J. Nutr. Biochem. 1990, 1, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Sharf, Y.; Khan, M.A. Dietary Tryptophan Requirement of Fingerling Channa punctatus (Bloch) Based on Growth, Hematological Parameters, Intestinal Enzymes, Non-Specific Immune Response, and Antioxidant Capacity. Aquaculture 2023, 562, 738745. [Google Scholar] [CrossRef]
- Khan, M.S.K.; Salin, K.R.; Yakupitiyage, A.; Tsusaka, T.W.; Nguyen, L.T.; Siddique, M.A.M. L-Tryptophan Mitigates Cannibalism and Improves Growth of Asian Seabass, Lates calcarifer Reared in a RAS System. Aquac. J. 2023, 3, 168–180. [Google Scholar] [CrossRef]
- Martins, C.I.M.; Silva, P.I.M.; Costas, B.; Larsen, B.K.; Santos, G.A.; Conceição, L.E.C.; Dias, J.; Øverli, Ø.; Höglund, E.; Schrama, J.W. The Effect of Tryptophan Supplemented Diets on Brain Serotonergic Activity and Plasma Cortisol under Undisturbed and Stressed Conditions in Grouped-Housed Nile Tilapia Oreochromis niloticus. Aquaculture 2013, 400–401, 129–134. [Google Scholar] [CrossRef]
- Teixeira, C.; Pedrosa, R.; Castro, C.; Magalhães, R.; Matos, E.; Oliva-Teles, A.; Peres, H.; Pérez-Jiménez, A. Dietary Tryptophan Supplementation Implications on Performance, Plasma Metabolites, and Amino Acid Catabolism Enzymes in Meagre (Argyrosomus regius). Fishes 2023, 8, 141. [Google Scholar] [CrossRef]
- Tejpal, C.S.; Pal, A.K.; Sahu, N.P.; Ashish Kumar, J.; Muthappa, N.A.; Vidya, S.; Rajan, M.G. Dietary Supplementation of L-Tryptophan Mitigates Crowding Stress and Augments the Growth in Cirrhinus mrigala Fingerlings. Aquaculture 2009, 293, 272–277. [Google Scholar] [CrossRef]
- Hoglund, E.; Sørensen, C.; Bakke, M.J.; Nilsson, G.E.; Øverli, Ø. Attenuation of Stress-Induced Anorexia in Brown Trout (Salmo trutta) by Pre-Treatment with Dietary L-Tryptophan. Br. J. Nutr. 2007, 97, 786–789. [Google Scholar] [CrossRef] [PubMed]
- Hoseini, S.M.; Pérez-Jiménez, A.; Costas, B.; Azeredo, R.; Gesto, M. Physiological Roles of Tryptophan in Teleosts: Current Knowledge and Perspectives for Future Studies. Rev. Aquac. 2019, 11, 3–24. [Google Scholar] [CrossRef]
- Fossat, P.; Bacqué-Cazenave, J.; De Deurwaerdère, P.; Cattaert, D.; Delbecque, J.P. Serotonin, but Not Dopamine, Controls the Stress Response and Anxiety-like Behavior in the Crayfish Procambarus clarkii. J. Exp. Biol. 2015, 218, 2745–2752. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.P.; Guan, L.Z.; Xiong, J.H.; Xi, Q.Y.; Zhang, Y.L. Effects of L-Tryptophan-Supplemented Dietary on Growth Performance and 5-HT and GABA Levels in Juvenile Litopenaeus vannamei. Aquac. Int. 2015, 23, 235–251. [Google Scholar] [CrossRef]
- Jin, Y.; Liu, F.J.; Liu, Y.J.; Tian, L.X.; Zhang, Z.H. Dietary Tryptophan Requirements of Juvenile Pacific White Shrimp, Litopenaeus vannamei (Boone) Reared in Low-Salinity Water. Aquac. Int. 2017, 25, 955–968. [Google Scholar] [CrossRef]
- Laranja, J.L.Q.; Quinitio, E.T.; Catacutan, M.R.; Coloso, R.M. Effects of Dietary L-Tryptophan on the Agonistic Behavior, Growth and Survival of Juvenile Mud Crab Scylla serrata. Aquaculture 2010, 310, 84–90. [Google Scholar] [CrossRef]
- Rosner, A.; Armengaud, J.; Ballarin, L.; Barnay-Verdier, S.; Cima, F.; Coelho, A.V.; Domart-Coulon, I.; Drobne, D.; Genevière, A.M.; Jemec Kokalj, A.; et al. Stem Cells of Aquatic Invertebrates as an Advanced Tool for Assessing Ecotoxicological Impacts. Sci. Total Environ. 2021, 771, 144565. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Fujii, Y.; Nakano, T.; Arimoto, T.; Murata, M.; Matsumoto, H.; Yoshiura, Y.; Ohnuki, H.; Endo, H. Development of a Novel Enhanced Biosensor System for Real-Time Monitoring of Fish Stress Using a Self-Assembled Monolayer. Sensors 2019, 19, 1518. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, R.; Wu, H.; Murata, M.; Ohnuki, H.; Yoshiura, Y.; Endo, H. Development of an Optical Communication Type Biosensor for Real-Time Monitoring of Fish Stress. Sens. Actuators B Chem. 2017, 247, 765–773. [Google Scholar] [CrossRef]
- Li, D.; Wang, G.; Du, L.; Zheng, Y.; Wang, Z. Recent Advances in Intelligent Recognition Methods for Fish Stress Behavior. Aquac. Eng. 2022, 96, 102222. [Google Scholar] [CrossRef]
- Barreto, M.O.; Rey Planellas, S.; Yang, Y.; Phillips, C.; Descovich, K. Emerging Indicators of Fish Welfare in Aquaculture. Rev. Aquac. 2022, 14, 343–361. [Google Scholar] [CrossRef]
- Duan, Y.; Nan, Y.; Zhu, X.; Yang, Y.; Xing, Y. The Adverse Impacts of Ammonia Stress on the Homeostasis of Intestinal Health in Pacific White Shrimp (Litopenaeus vannamei). Environ. Pollut. 2024, 340, 122762. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shen, Y.; Bao, Y.; Wu, Z.; Yang, B.; Jiao, L.; Zhang, C.; Tocher, D.R.; Zhou, Q.; Jin, M. Physiological Responses and Adaptive Strategies to Acute Low-Salinity Environmental Stress of the Euryhaline Marine Fish Black Seabream (Acanthopagrus schlegelii). Aquaculture 2022, 554, 738117. [Google Scholar] [CrossRef]
- Kumar, N.; Thorat, S.T.; Pradhan, A.; Rane, J.; Reddy, K.S. Significance of Dietary Quinoa Husk (Chenopodium quinoa) in Gene Regulation for Stress Mitigation in Fish. Sci. Rep. 2024, 14, 7647. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.; Buentello, A.; Gatlin, D.; Lightner, D.; Hume, M.; Lawrence, A. Dietary Supplementation of Galactooligosaccharides (GOS) in Pacific White Shrimp, Litopenaeus vannamei, Cultured in a Recirculating System and Its Effects on Gut Microflora, Growth, Stress, and Immune Response. J. Immunoass. Immunochem. 2019, 40, 662–675. [Google Scholar] [CrossRef] [PubMed]
- Luc, Q.C.; Ncho, C.M.; Dhahbi, S.; Olowe, O.S. Mitigation of Cold Stress in Nile Tilapia (Oreochromis niloticus) through Dietary Lipids Supplementation: A Preliminary Network Meta-Analysis. Fish Physiol. Biochem. 2024, 50, 209–223. [Google Scholar] [CrossRef] [PubMed]
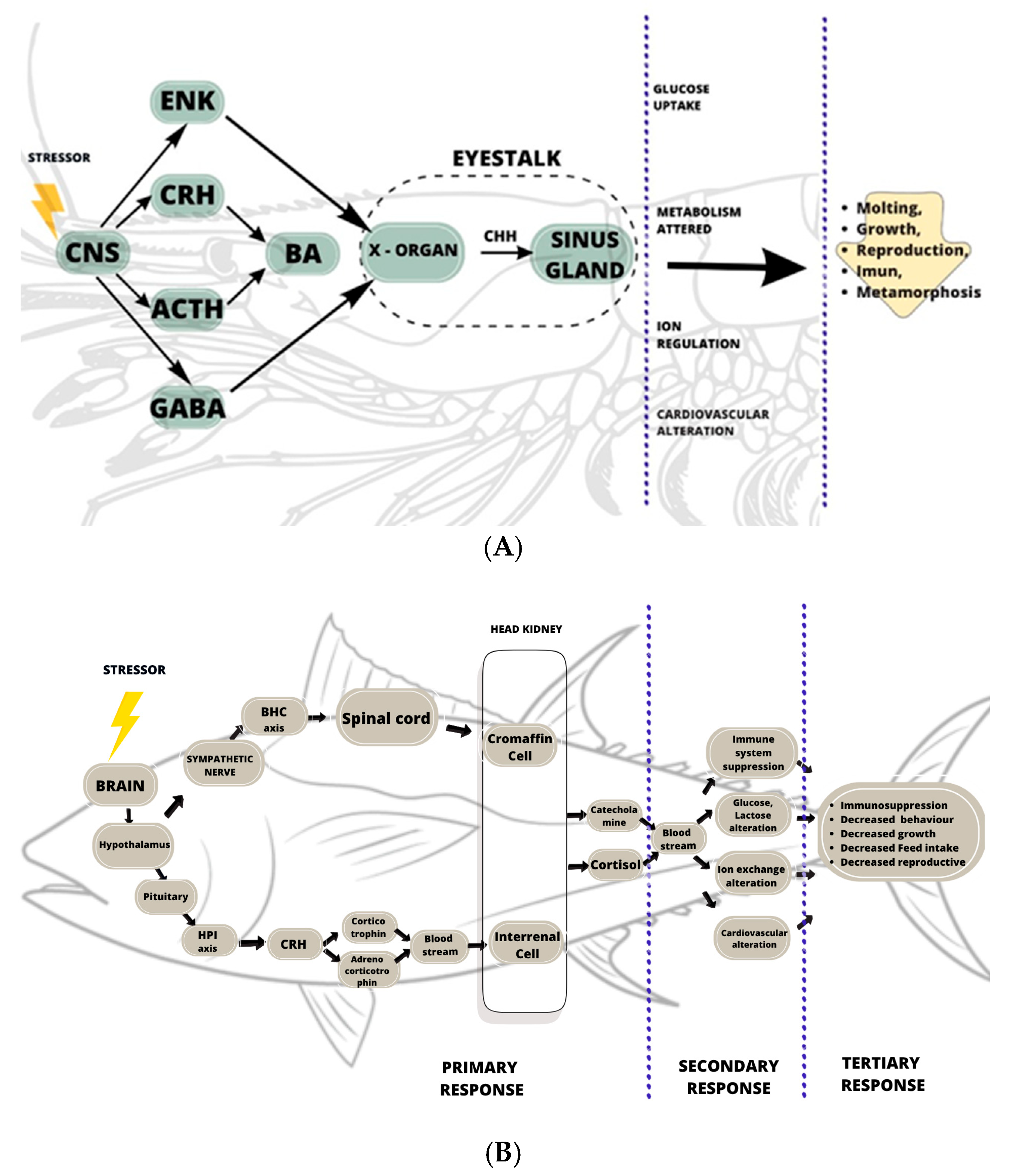

| Invertebrate Crustacean | Vertebrate Fish | |
|---|---|---|
| Signaling system | Neuroendocrine system | Endocrine system |
| Axis | XO-SG axis | BSC axis and HPI axis |
| Hormones | Crustacean hyperglycemic hormone/CHH | Cortisol and Catecholamine |
| Biogenic amines | Norepinephrine, GABA, dopamine, serotonin | Norepinephrine, GABA, dopamine, serotonin |
| Major signaling pathway | Central nervous system | Hypothalamus, spinal cord, and pituitary |
| Stress hormone-producing organ | Eye stalk | Head kidney |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hapsari, F.; Suprayudi, M.A.; Akiyama, D.M.; Ekasari, J.; Norouzitallab, P.; Baruah, K. Decoding Stress Responses in Farmed Crustaceans: Comparative Insights for Sustainable Aquaculture Management. Biology 2025, 14, 920. https://doi.org/10.3390/biology14080920
Hapsari F, Suprayudi MA, Akiyama DM, Ekasari J, Norouzitallab P, Baruah K. Decoding Stress Responses in Farmed Crustaceans: Comparative Insights for Sustainable Aquaculture Management. Biology. 2025; 14(8):920. https://doi.org/10.3390/biology14080920
Chicago/Turabian StyleHapsari, Fitriska, Muhammad Agus Suprayudi, Dean M. Akiyama, Julie Ekasari, Parisa Norouzitallab, and Kartik Baruah. 2025. "Decoding Stress Responses in Farmed Crustaceans: Comparative Insights for Sustainable Aquaculture Management" Biology 14, no. 8: 920. https://doi.org/10.3390/biology14080920
APA StyleHapsari, F., Suprayudi, M. A., Akiyama, D. M., Ekasari, J., Norouzitallab, P., & Baruah, K. (2025). Decoding Stress Responses in Farmed Crustaceans: Comparative Insights for Sustainable Aquaculture Management. Biology, 14(8), 920. https://doi.org/10.3390/biology14080920