

Timing and Amplitude of Light Exposure, Not Photoperiod, Predict Blood Lipids in Arctic Residents: A Circadian Light Hypothesis

 , ,
, ,  , ,
, , 
 , , ,
, , , 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Inclusion and Exclusion Criteria
2.2. Actigraphy
2.3. Blood Lipids
2.4. Melatonin
2.5. Chronotype
2.6. Data and Statistical Analyses
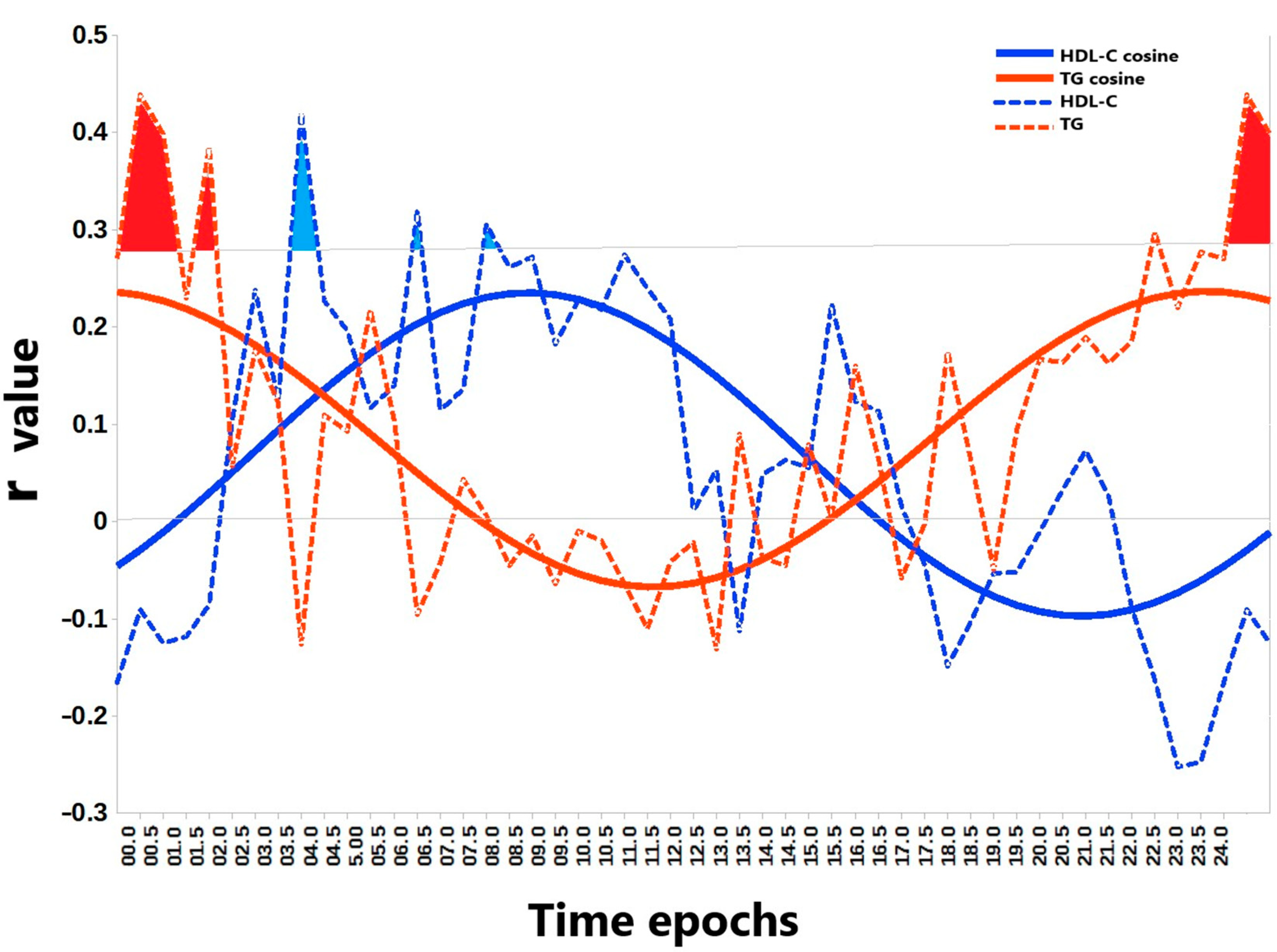
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sahar, S.; Sassone-Corsi, P. Regulation of metabolism: The circadian clock dictates the time. Trends Endocrinol. Metab. 2012, 23, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, K.; Saeki, K.; Iwamoto, J.; Okamoto, N.; Tomioka, K.; Nezu, S.; Ikada, Y.; Kurumatani, N. Exposure to light at night, nocturnal urinary melatonin excretion, and obesity/dyslipidemia in the elderly: A cross-sectional analysis of the HEIJO-KYO study. J. Clin. Endocrinol. Metab. 2013, 98, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Maury, E.; Hong, H.K.; Bass, J. Circadian disruption in the pathogenesis of metabolic syndrome. Diabetes Metab. 2014, 40, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Gnocchi, D.; Pedrelli, M.; Hurt-Camejo, E.; Parini, P. Lipids around the Clock: Focus on Circadian Rhythms and Lipid Metabolism. Biology 2015, 4, 104–132. [Google Scholar] [CrossRef]
- Poggiogalle, E.; Jamshed, H.; Peterson, C.M. Circadian regulation of glucose, lipid, and energy metabolism in humans. Metabolism 2018, 84, 11–27. [Google Scholar] [CrossRef]
- Zimmet, P.; Alberti, K.G.M.M.; Stern, N.; Bilu, C.; El-Osta, A.; Einat, H.; Kronfeld-Schor, N. The Circadian Syndrome: Is the Metabolic Syndrome and much more! J. Intern. Med. 2019, 286, 181–191. [Google Scholar] [CrossRef]
- Cornelissen, G. Circadian disruption and metabolic disease risk. In Functional Foods and Nutraceuticals in Metabolic and Non-Communicable Diseases; Singh, R., Watanabe, S., Isaza, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 513–524. [Google Scholar]
- Okuliarova, M.; Rumanova, V.S.; Stebelova, K.; Zeman, M. Dim Light at Night Disturbs Molecular Pathways of Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6919. [Google Scholar] [CrossRef]
- Rumanova, V.S.; Okuliarova, M.; Foppen, E.; Kalsbeek, A.; Zeman, M. Exposure to dim light at night alters daily rhythms of glucose and lipid metabolism in rats. Front. Physiol. 2022, 13, 973461. [Google Scholar] [CrossRef]
- Gubin, D.; Neroev, V.; Malishevskaya, T.; Kolomeichuk, S.; Weinert, D.; Yuzhakova, N.; Nelaeva, A.; Filippova, Y.; Cornelissen, G. Daytime Lipid Metabolism Modulated by CLOCK Gene Is Linked to Retinal Ganglion Cells Damage in Glaucoma. Appl. Sci. 2022, 12, 6374. [Google Scholar] [CrossRef]
- Gubin, D.; Danilenko, K.; Stefani, O.; Kolomeichuk, S.; Markov, A.; Petrov, I.; Voronin, K.; Mezhakova, M.; Borisenkov, M.; Shigabaeva, A.; et al. Blue Light and Temperature Actigraphy Measures Predicting Metabolic Health Are Linked to Melatonin Receptor Polymorphism. Biology 2024, 13, 22. [Google Scholar] [CrossRef]
- Gubin, D.; Danilenko, K.; Stefani, O.; Kolomeichuk, S.; Markov, A.; Petrov, I.; Voronin, K.; Mezhakova, M.; Borisenkov, M.; Shigabaeva, A.; et al. Light Environment of Arctic Solstices is Coupled With Melatonin Phase-Amplitude Changes and Decline of Metabolic Health. J. Pineal Res. 2025, 77, e70023. [Google Scholar] [CrossRef] [PubMed]
- Kent, B.A.; Rahman, S.A.; St Hilaire, M.A.; Grant, L.K.; Rüger, M.; Czeisler, C.A.; Lockley, S.W. Circadian lipid and hepatic protein rhythms shift with a phase response curve different than melatonin. Nat. Commun. 2022, 13, 681, Erratum in Nat. Commun. 2022, 13, 2241.. [Google Scholar] [CrossRef] [PubMed]
- Kyle, J.E.; Bramer, L.M.; Claborne, D.; Stratton, K.G.; Bloodsworth, K.J.; Teeguarden, J.G.; Gaddameedhi, S.; Metz, T.O.; Van Dongen, H.P.A. Simulated Night-Shift Schedule Disrupts the Plasma Lipidome and Reveals Early Markers of Cardiovascular Disease Risk. Nat. Sci. Sleep 2022, 14, 981–994, Erratum in Nat. Sci. Sleep 2023, 15, 477–478.. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, A.; Courville, A.B.; Chen, K.Y. The Complex Effects of Light on Metabolism in Humans. Nutrients 2023, 15, 1391. [Google Scholar] [CrossRef]
- Han, B.; Deng, J.; Huang, X.; Yin, X.; Liang, Y. Effects of exposure to outdoor light at night on blood lipids in China: A nationwide cross-sectional study. Sci. Rep. 2024, 14, 28263. [Google Scholar] [CrossRef]
- Brown, T.M.; Brainard, G.C.; Cajochen, C.; Czeisler, C.A.; Hanifin, J.P.; Lockley, S.W.; Lucas, R.J.; Münch, M.; O’Hagan, J.B.; Peirson, S.N.; et al. Recommendations for daytime, evening, and nighttime indoor light exposure to best support physiology, sleep, and wakefulness in healthy adults. PLoS Biol. 2022, 20, e3001571. [Google Scholar] [CrossRef]
- Kim, M.; Vu, T.H.; Maas, M.B.; Braun, R.I.; Wolf, M.S.; Roenneberg, T.; Daviglus, M.L.; Reid, K.J.; Zee, P.C. Light at night in older age is associated with obesity, diabetes, and hypertension. Sleep 2023, 46, zsac130. [Google Scholar] [CrossRef]
- Gubin, D.G.; Borisenkov, M.F.; Kolomeichuk, S.N.; Markov, A.; Weinert, D.; Cornelissen, G.; Stefani, O. Evaluating circadian light hygiene: Methodology and health implications. Russ. Open Med. J. 2024, 13, e0415. [Google Scholar] [CrossRef]
- Windred, D.P.; Burns, A.C.; Rutter, M.K.; Ching Yeung, C.H.; Lane, J.M.; Xiao, Q.; Saxena, R.; Cain, S.W.; Phillips, A.J.K. Personal light exposure patterns and incidence of type 2 diabetes: Analysis of 13 million hours of light sensor data and 670,000 person-years of prospective observation. Lancet Reg. Health Eur. 2024, 42, 100943. [Google Scholar] [CrossRef]
- Windred, D.P.; Burns, A.C.; Lane, J.M.; Olivier, P.; Rutter, M.K.; Saxena, R.; Phillips, A.J.K.; Cain, S.W. Brighter nights and darker days predict higher mortality risk: A prospective analysis of personal light exposure in >88,000 individuals. Proc. Natl. Acad. Sci. USA 2024, 121, e2405924121. [Google Scholar] [CrossRef]
- Baek, J.H.; Zhu, Y.; Jackson, C.L.; Park, Y.M. Artificial Light at Night and Type 2 Diabetes Mellitus. Diabetes Metab. J. 2024, 48, 847–863. [Google Scholar] [CrossRef] [PubMed]
- Depner, C.M.; Cheng, P.C.; Devine, J.K.; Khosla, S.; de Zambotti, M.; Robillard, R.; Vakulin, A.; Drummond, S.P.A. Wearable technologies for developing sleep and circadian biomarkers: A summary of workshop discussions. Sleep 2020, 43, zsz254. [Google Scholar] [CrossRef] [PubMed]
- Gubin, D.; Weinert, D.; Stefani, O.; Otsuka, K.; Borisenkov, M.; Cornelissen, G. Wearables in Chronomedicine and Interpretation of Circadian Health. Diagnostics 2025, 15, 327. [Google Scholar] [CrossRef] [PubMed]
- Gubin, D.; Boldyreva, J.; Stefani, O.; Kolomeichuk, S.; Danilova, L.; Shigabaeva, A.; Cornelissen, G.; Weinert, D. Higher vulnerability to poor circadian light hygiene in individuals with a history of COVID-19. Chronobiol. Int. 2025, 42, 133–146. [Google Scholar] [CrossRef]
- Gubin, D.; Kolomeichuk, S.; Danilenko, K.; Stefani, O.; Markov, A.; Petrov, I.; Voronin, K.; Mezhakova, M.; Borisenkov, M.; Shigabaeva, A.; et al. Phase coupling of light exposure and activity differs between Arctic Natives and Non-Natives. Biol. Rhythm. Res. 2025, 1–11. [Google Scholar] [CrossRef]
- Burgess, H.J.; Kagan, D.; Rizvydeen, M.; Swanson, L.M.; Kim, H.M. An independent comparison of the Novolytix salivary melatonin radioimmunoassay with the new Novolytix salivary melatonin enzyme-linked immunosorbent assay. J. Pineal Res. 2024, 76, e12933. [Google Scholar] [CrossRef]
- Horne, J.A.; Ostberg, O. A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int. J. Chronobiol. 1976, 4, 97–110. [Google Scholar]
- Lee, J.S.; Chang, P.Y.; Zhang, Y.; Kizer, J.R.; Best, L.G.; Howard, B.V. Triglyceride and HDL-C Dyslipidemia and Risks of Coronary Heart Disease and Ischemic Stroke by Glycemic Dysregulation Status: The Strong Heart Study. Diabetes Care 2017, 40, 529–537. [Google Scholar] [CrossRef]
- Georgoulis, M.; Chrysohoou, C.; Georgousopoulou, E.; Damigou, E.; Skoumas, I.; Pitsavos, C.; Panagiotakos, D. Long-term prognostic value of LDL-C, HDL-C, lp(a) and TG levels on cardiovascular disease incidence, by body weight status, dietary habits and lipid-lowering treatment: The ATTICA epidemiological cohort study (2002-2012). Lipids Health Dis. 2022, 21, 141. [Google Scholar] [CrossRef]
- Delorme, T.C.; Srikanta, S.B.; Fisk, A.S.; Cloutier, M.È.; Sato, M.; Pothecary, C.A.; Merz, C.; Foster, R.G.; Brown, S.A.; Peirson, S.N.; et al. Chronic Exposure to Dim Light at Night or Irregular Lighting Conditions Impact Circadian Behavior, Motor Coordination, and Neuronal Morphology. Front. Neurosci. 2022, 16, 855154. [Google Scholar] [CrossRef]
- Hand, A.J.; Stone, J.E.; Shen, L.; Vetter, C.; Cain, S.W.; Bei, B.; Phillips, A.J.K. Measuring light regularity: Sleep regularity is associated with regularity of light exposure in adolescents. Sleep 2023, 46, zsad001. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.A.; Qiu, X.; Schwartz, J.; Huang, T.; Scheer, F.A.J.L.; Redline, S.; Sofer, T. Light exposure during sleep is bidirectionally associated with irregular sleep timing: The multi-ethnic study of atherosclerosis (MESA). Environ. Pollut. 2024, 344, 123258. [Google Scholar] [CrossRef] [PubMed]
- Grant, L.K.; St Hilaire, M.A.; Brainard, G.C.; Czeisler, C.A.; Lockley, S.W.; Rahman, S.A. Endogenous circadian regulation and phase resetting of clinical metabolic biomarkers. J. Pineal Res. 2021, 71, e12752. [Google Scholar] [CrossRef]
- Rahman, S.A.; Gathungu, R.M.; Marur, V.R.; St Hilaire, M.A.; Scheuermaier, K.; Belenky, M.; Struble, J.S.; Czeisler, C.A.; Lockley, S.W.; Klerman, E.B.; et al. Age-related changes in circadian regulation of the human plasma lipidome. Commun. Biol. 2023, 6, 756. [Google Scholar] [CrossRef]
- Duffy, J.F.; Zitting, K.M.; Chinoy, E.D. Aging and Circadian Rhythms. Sleep Med. Clin. 2015, 10, 423–434. [Google Scholar] [CrossRef]
- Shen, B.; Ma, C.; Wu, G.; Liu, H.; Chen, L.; Yang, G. Effects of exercise on circadian rhythms in humans. Front. Pharmacol. 2023, 14, 1282357. [Google Scholar] [CrossRef]
- Solt, L.A.; Kojetin, D.J.; Burris, T.P. The REV-ERBs and RORs: Molecular links between circadian rhythms and lipid homeostasis. Future Med. Chem. 2011, 3, 623–638. [Google Scholar] [CrossRef]
- Gubin, D.; Kolomeichuk, S.; Danilenko, K.; Stefani, O.; Markov, A.; Petrov, I.; Voronin, K.; Mezhakova, M.; Borisenkov, M.; Shigabaeva, A.; et al. Light Exposure, Physical Activity, and Indigeneity Modulate Seasonal Variation in NR1D1 (REV-ERBα) Expression. Biology 2025, 14, 231. [Google Scholar] [CrossRef] [PubMed]
- Kosmas, C.E.; Rodriguez Polanco, S.; Bousvarou, M.D.; Papakonstantinou, E.J.; Peña Genao, E.; Guzman, E.; Kostara, C.E. The Triglyceride/High-Density Lipoprotein Cholesterol (TG/HDL-C) Ratio as a Risk Marker for Metabolic Syndrome and Cardiovascular Disease. Diagnostics 2023, 13, 929. [Google Scholar] [CrossRef]
- Gubin, D.; Malishevskaya, T.; Weinert, D.; Zakharova, E.; Astakhov, S.; Cornelissen, G. Circadian Disruption in Glaucoma: Causes, Consequences, and Countermeasures. Front. Biosci. 2024, 29, 410. [Google Scholar] [CrossRef]
- Obara, E.A.; Hannibal, J.; Heegaard, S.; Fahrenkrug, J. Loss of Melanopsin-Expressing Retinal Ganglion Cells in Severely Staged Glaucoma Patients. Investig. Ophthalmol. Vis. Sci. 2016, 57, 4661–4667. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Variable | TC | LDL-C | HDL-C | TG | TG/HDL |
|---|---|---|---|---|---|
| Photoperiod | 0.070 | 0.033 | 0.022 | −0.003 | −0.110 |
| LE MESOR | 0.128 | 0.119 | −0.049 | 0.126 | −0.067 |
| LE Amplitude | 0.039 | 0.038 | −0.087 | 0.036 | −0.115 |
| LE Acrophase | −0.218 | −0.153 | −0.280 * | −0.051 | 0.121 |
| BLE MESOR | 0.116 | 0.111 | −0.052 | 0.096 | −0.071 |
| BLE Amplitude | 0.030 | 0.039 | −0.088 | 0.019 | −0.109 |
| BLE Acrophase | −0.222 | −0.161 | −0.282 * | −0.041 | 0.112 |
| M10 BLE | 0.090 | 0.086 | −0.053 | 0.061 | −0.095 |
| M10 Onset BLE | −0.216 | −0.121 | −0.268 * | −0.126 | 0.007 |
| L5 BLE | 0.312 * | 0.266 | 0.154 | 0.234 | 0.074 |
| L5 BLE Onset | 0.231 | 0.229 | 0.015 | 0.255 | 0.208 |
| RA BLE | −0.098 | −0.074 | −0.036 | −0.162 | −0.154 |
| NA BLE | −0.113 | −0.142 | 0.193 | −0.331 * | −0.405 * |
| Melatonin acrophase | 0.060 | 0.123 | −0.282 | 0.454 * | 0.510 * |
| Variable | TC | LDL-C | HDL-C | TG | TG/HDL-C | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| β R2 | p | β R2 | p | β R2 | p | β R2 | p | β R2 | p | |
| Timing of Light Exposure | ||||||||||
| LE acrophase | --- | --- | --- | −0.280 0.079 | 0.012 | --- | --- | --- | ||
| BLE acrophase | −0.221 0.054 | 0.049 | --- | --- | −0.282 0.080 | 0.012 | --- | --- | --- | |
| M10 BLE Onset | --- | --- | --- | −0.274 0.073 | 0.017 | --- | --- | --- | ||
| L5 BLE Onset | 0.233 0.059 | 0.039 | 0.230 0.054 | 0.042 | --- | --- | 0.255 0.065 | 0.023 | 0.206 0.055 | 0.067 |
| Nocturnal Blue Light Exposure | ||||||||||
| L5 BLE | 0.309 0.098 | 0.006 | 0.267 0.071 | 0.026 | --- | --- | 0.240 0.056 | 0.036 | --- | --- 0.098 |
| Dynamic Range of Light Exposure | ||||||||||
| NA BLE | --- | --- | 0.002 | 0.345 0.114 | 0.197 0.037 | 0.089 | 0.345 0.114 | 0.002 | 0.399 0.164 | <0.001 |
| Melatonin | ||||||||||
| Melatonin acrophase | --- | --- | --- | --- | −0.297 0.087 | 0.057 | 0.473 0.217 | 0.002 | 0.503 0.250 | <0.001 |
| Variable | Winter Solstice (WS) | Spring Equinox (SE) | Summer Solstice (SS) |
|---|---|---|---|
| LE MESOR, lux | 23.68 ± 13.92 | 74.01 ± 46.88 * | 213.67 ± 234.66 **/** |
| LE Amplitude, lux | 25.75 ± 14.78 | 103.93 ± 67.53 * | 276.70 ± 340.14 **/** |
| LE Acrophase, hh:mm | 14:14 ± 1:15 | 12:57 ± 0:58 ** | 14:12 ± 1:36 ns/** |
| BLE MESOR, μw/cm2 | 2.64 ± 1.61 | 11.11 ± 7.55 * | 36.66 ± 43.81 **/** |
| BLE Amplitude, μw/cm2 | 2.93 ± 1.74 | 16.12 ± 11.19 * | 49.91 ± 65.04 **/** |
| BLE Acrophase, hh:mm | 14:12 ± 1:12 | 12:52 ± 0:57 ** | 14:06 ± 1:33 ns/** |
| M10 BLE, μw/cm2 | 4.99 ± 2.92 | 23.48 ± 15.35 * | 67.62 ± 78.70 **/** |
| M10 Onset BLE, hh:mm | 8:54 ± 1:06 | 7:52 ± 0:49 ** | 8:20 ± 1:08 */* |
| L5 BLE, μw/cm2 | 0.128 ± 0.213 | 0.077 ± 0.019 ns | 0.816 ± 2.871 */* |
| L5 BLE Onset, hh:mm | 2:04 ± 1:31 | 1:20 ± 1:49 * | 1:42 ± 1:19 ns/ns |
| RA BLE, a.u. | 0.934 ± 0.110 | 0.983 ± 0.026 ** | 0.960 ± 0.059 ns/ns |
| NA BLE, a.u. | 1.12 ± 0.20 | 1.43 ± 0.16 ** | 1.25 ± 0.29 */** |
| TC | LDL-C | HDL-C | TG | TG/HDL-C | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| β | p-Value | β | p-Value | β | p-Value | β | p-Value | β | p-Value | |
| Age | 0.261 | 0.023 | 0.244 | 0.053 | −0.017 | 0.873 | 0.160 | 0.164 | 0.138 | 0.212 |
| Sex (m) | −0.091 | 0.447 | 0.070 | 0.056 | −0.412 | <0.001 | 0.127 | 0.295 | 0.238 | 0.044 |
| Indigeneity (NN) | 0.004 | 0.969 | 0.065 | 0.578 | −0.167 | 0.124 | 0.333 | 0.003 | 0.288 | 0.010 |
| Photo-period | 0.061 | 0.565 | 0.021 | 0.847 | 0.034 | 0.727 | −0.029 | 0.785 | −0.132 | 0.220 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gubin, D.; Kolomeichuk, S.; Danilenko, K.; Stefani, O.; Markov, A.; Petrov, I.; Voronin, K.; Mezhakova, M.; Borisenkov, M.; Shigabaeva, A.; et al. Timing and Amplitude of Light Exposure, Not Photoperiod, Predict Blood Lipids in Arctic Residents: A Circadian Light Hypothesis. Biology 2025, 14, 799. https://doi.org/10.3390/biology14070799
Gubin D, Kolomeichuk S, Danilenko K, Stefani O, Markov A, Petrov I, Voronin K, Mezhakova M, Borisenkov M, Shigabaeva A, et al. Timing and Amplitude of Light Exposure, Not Photoperiod, Predict Blood Lipids in Arctic Residents: A Circadian Light Hypothesis. Biology. 2025; 14(7):799. https://doi.org/10.3390/biology14070799
Chicago/Turabian StyleGubin, Denis, Sergey Kolomeichuk, Konstantin Danilenko, Oliver Stefani, Alexander Markov, Ivan Petrov, Kirill Voronin, Marina Mezhakova, Mikhail Borisenkov, Aislu Shigabaeva, and et al. 2025. "Timing and Amplitude of Light Exposure, Not Photoperiod, Predict Blood Lipids in Arctic Residents: A Circadian Light Hypothesis" Biology 14, no. 7: 799. https://doi.org/10.3390/biology14070799
APA StyleGubin, D., Kolomeichuk, S., Danilenko, K., Stefani, O., Markov, A., Petrov, I., Voronin, K., Mezhakova, M., Borisenkov, M., Shigabaeva, A., Boldyreva, J., Petrova, J., Alkhimova, L., Weinert, D., & Cornelissen, G. (2025). Timing and Amplitude of Light Exposure, Not Photoperiod, Predict Blood Lipids in Arctic Residents: A Circadian Light Hypothesis. Biology, 14(7), 799. https://doi.org/10.3390/biology14070799