
Pro-Arrhythmic Effect of Chronic Stress-Associated Humoral Factors in Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Procedure of CUMS
2.3. Electrocardiographic Recordings
2.4. Serum Preparation and Treatment
2.5. Culturing of hiPSC-CMs and Monitoring Using CardioExcyte96
2.6. Cell Culture
2.7. Patch-Clamp Recordings
2.8. Western Blot Analysis
2.9. LC-MS/MS Analysis and Metabolomics
2.10. Statistical Analysis
3. Results
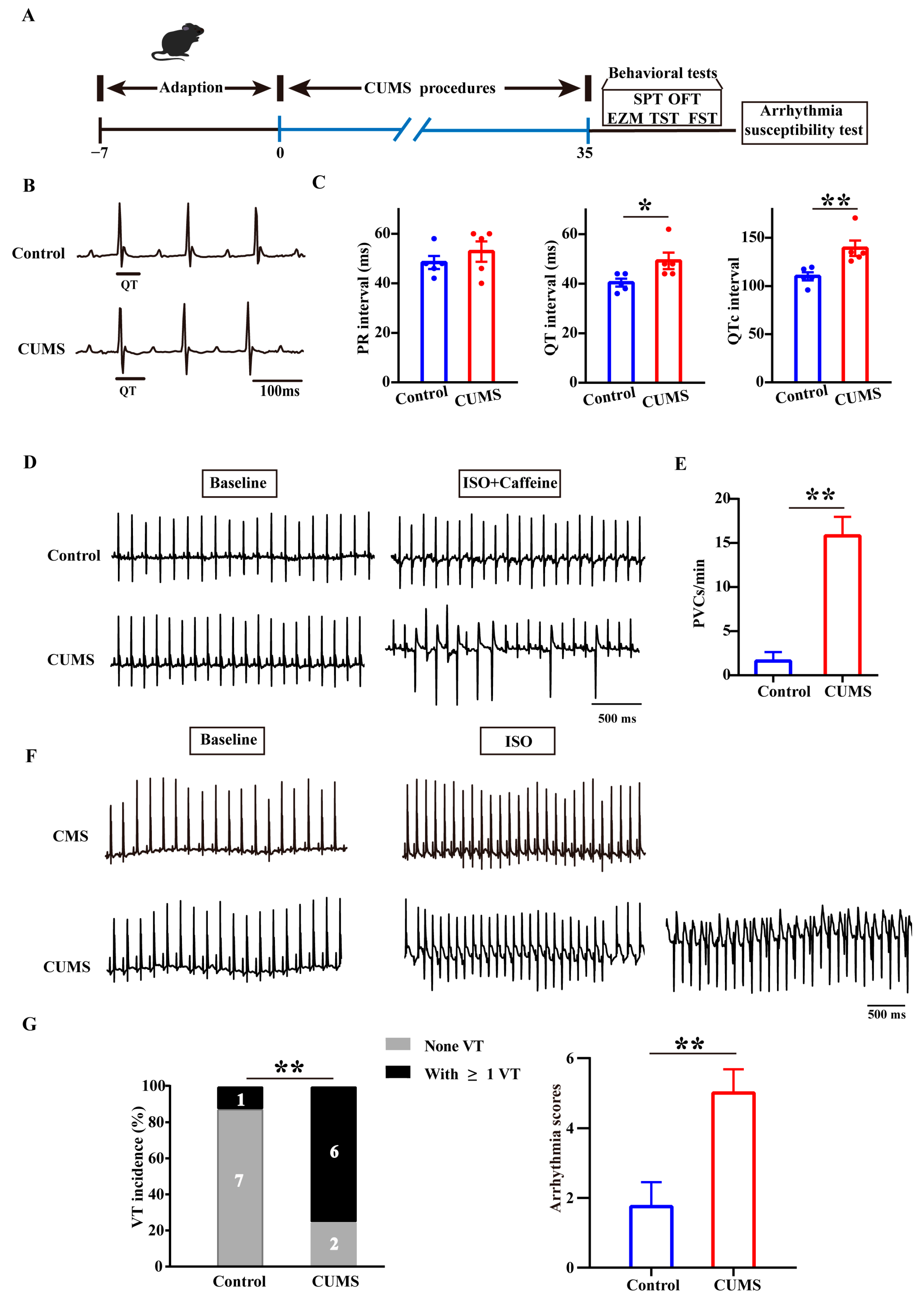
3.1. The CUMS Mice Had an Increased Susceptibility to Arrhythmia
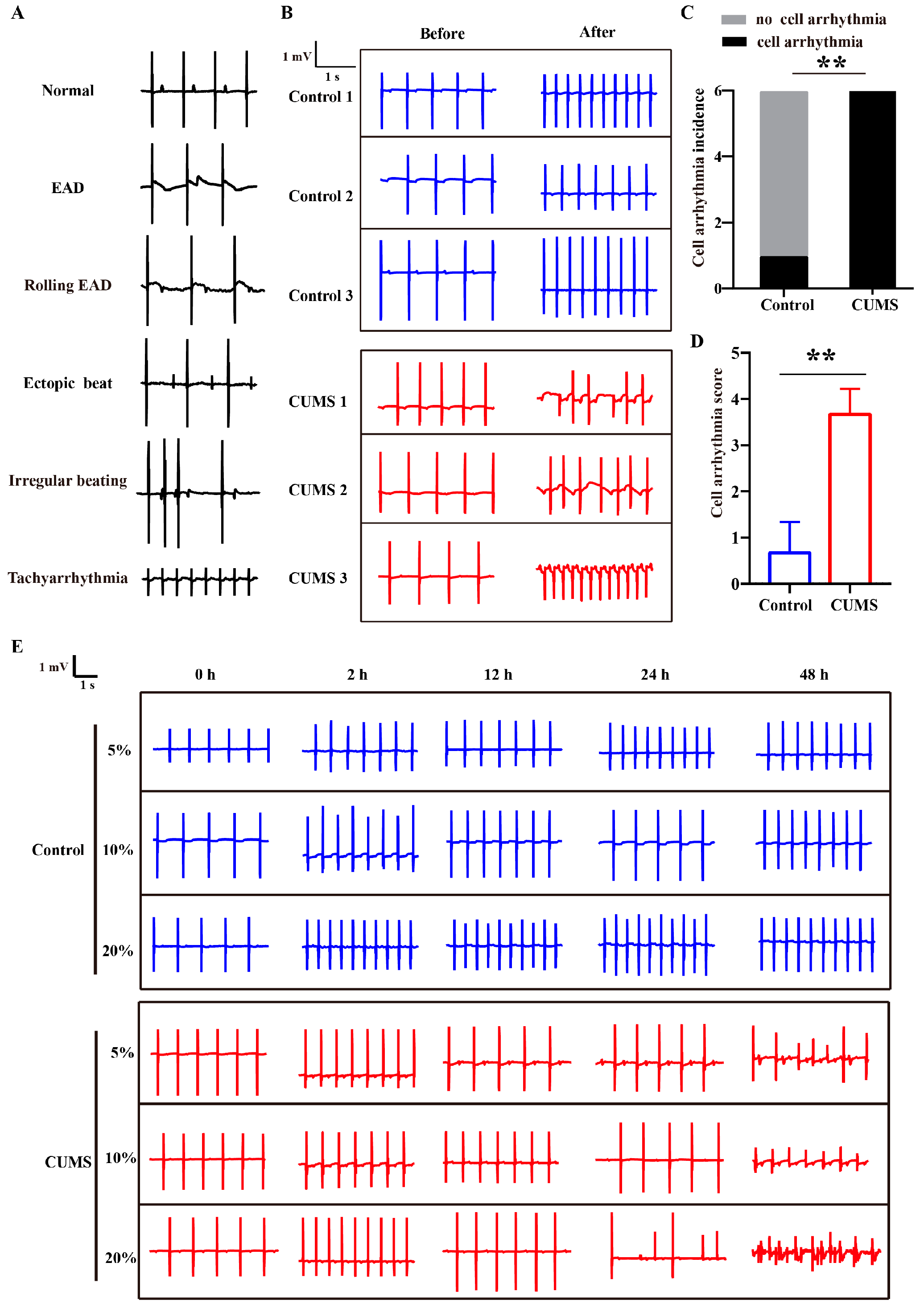
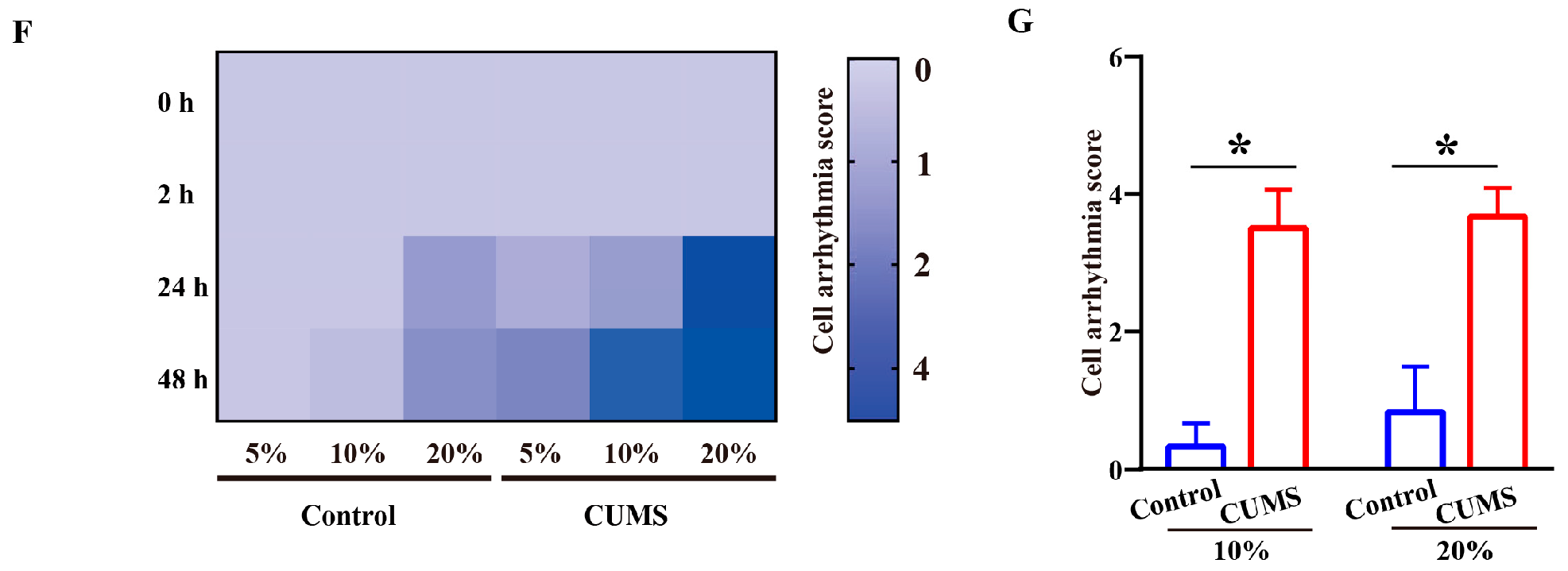
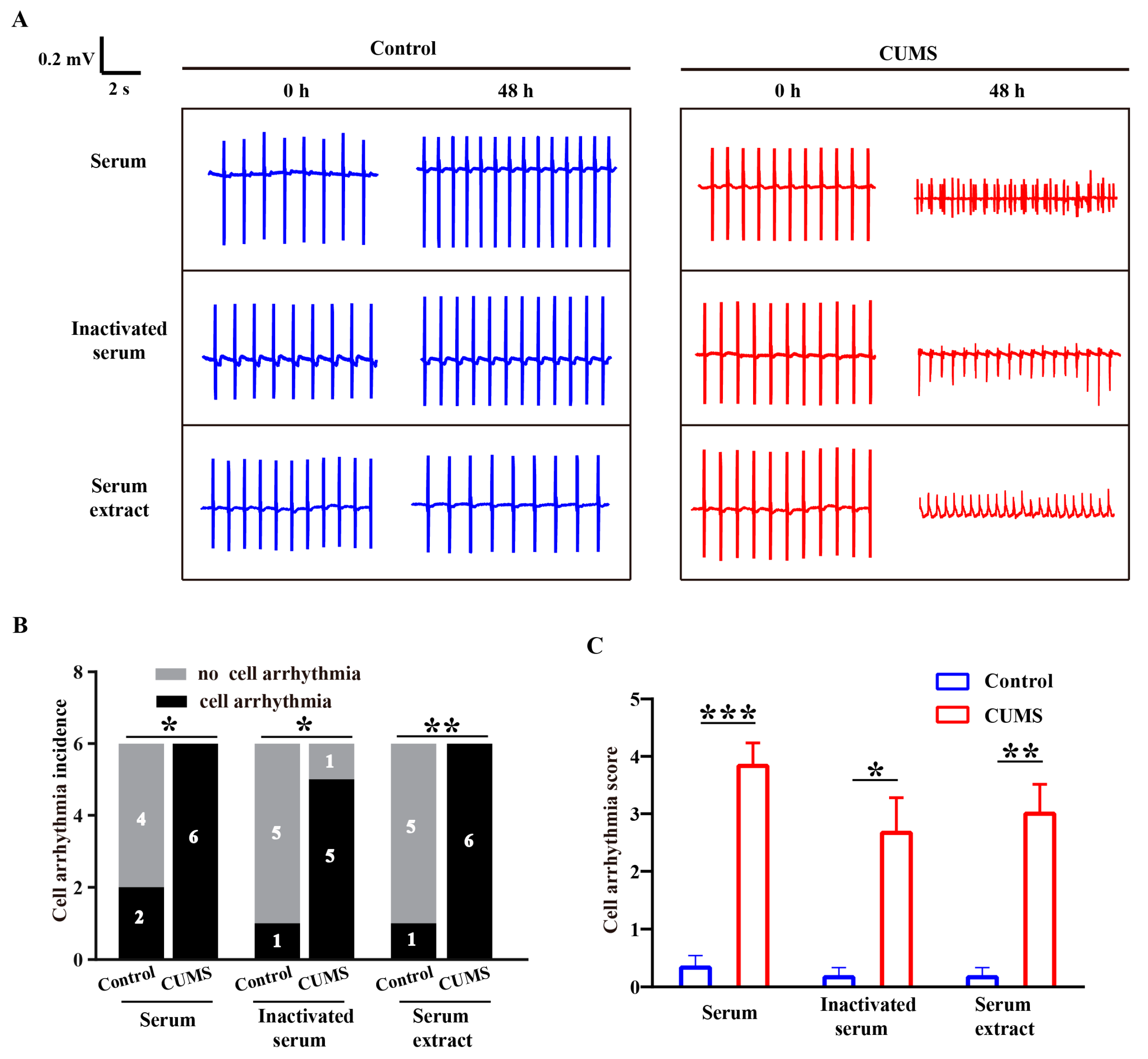
3.2. Pro-Arrhythmic Effect of CUMS Serum on Human Cardiomyocytes
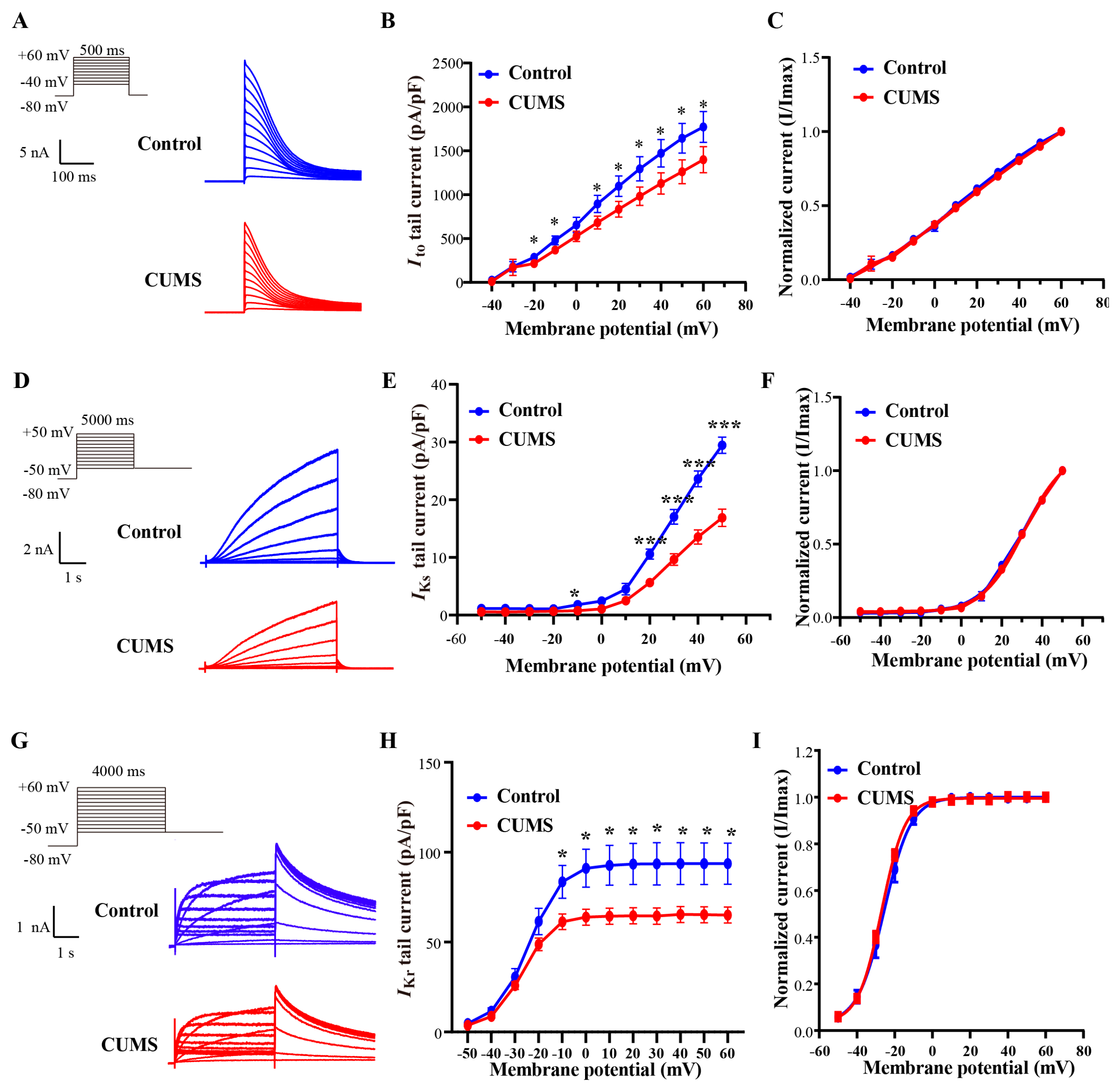
3.3. The CUMS Serum Induced Dysfunction of Cloned Human Potassium Ion Channels
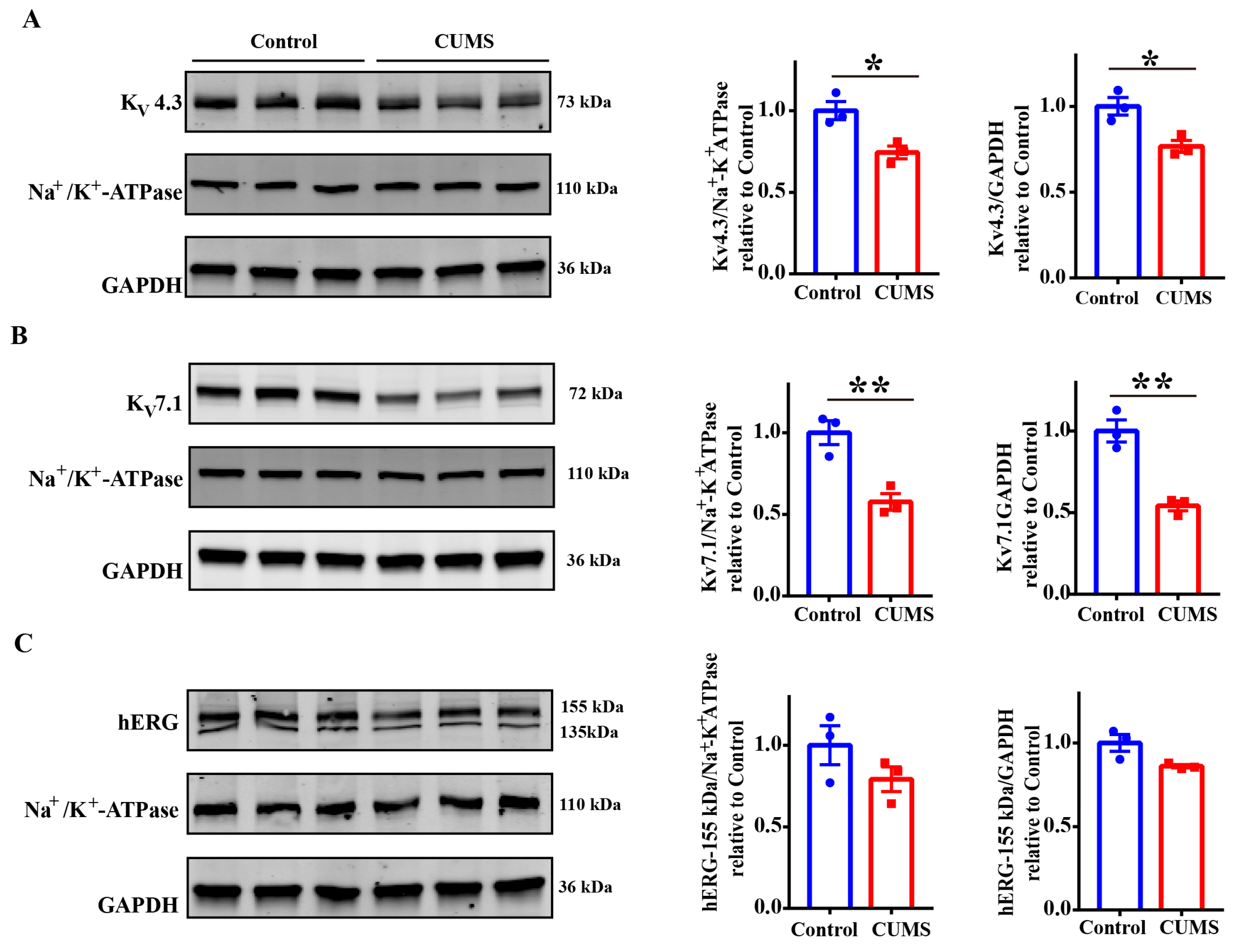
3.4. The CUMS Serum Downregulated the Expression of Potassium Channels
3.5. Small-Molecule Metabolites Mainly Mediated the Pro-Arrhythmic Effects Caused by CUMS
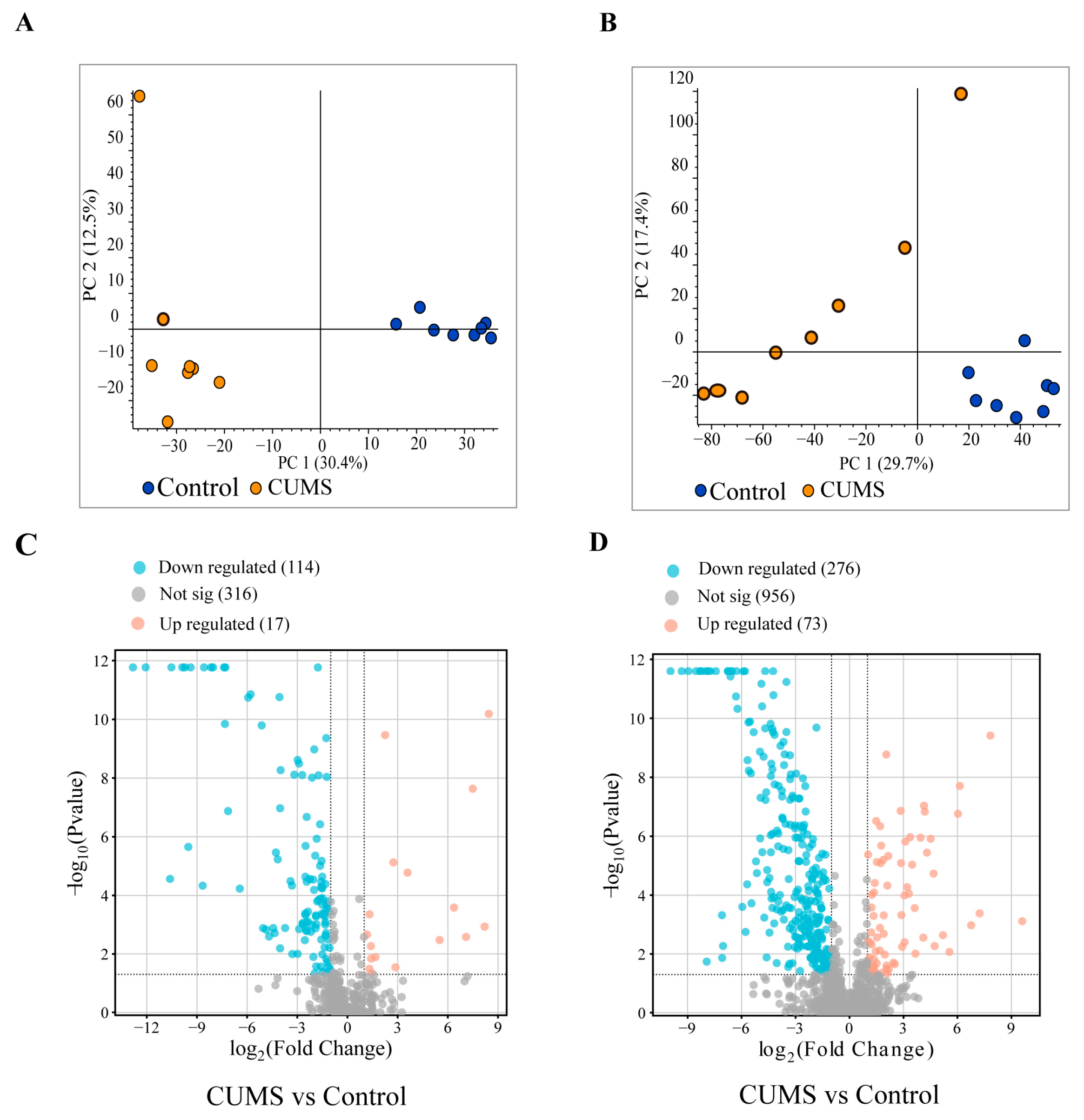
3.6. Analysis of Differential Metabolites in Serum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APD | Action potential duration |
| CUMS | Chronic unpredictable mild stress |
| hiPSC-CMs | Human-induced pluripotent stem cell-derived cardiomyocytes |
| EAD | Early afterdepolarization |
| ECG | Electrocardiogram |
| EFP | Extracellular field potential |
| EPM | Elevated plus-mazetest |
| FST | Forced swimming test |
| HEK | Human embryonic kidney |
| HPA | Hypothalamic–pituitary–adrenal axis |
| ICa-L | L-type calcium current |
| IKr | Rapid inward rectifier potassium current |
| IKs | Slow inward rectifier potassium current |
| iPSC | induced pluripotent stem cell |
| Ito | Electrocardiogram |
| ISO | Isoproterenol |
| OFT | Open field test |
| PCA | Principal component analysis |
| PVCs | Premature ventricular contractions |
| QTc | Corrected QT interval |
| SCD | Sudden cardiac death |
| SPT | Sucrose preference test |
| TST | Tail suspension test |
| VT | Ventricular tachycardia |
References
- Kivimäki, M.; Kawachi, I. Work Stress as a Risk Factor for Cardiovascular Disease. Curr. Cardiol. Rep. 2015, 17, 630. [Google Scholar] [CrossRef] [PubMed]
- Dragano, N.; Siegrist, J.; Nyberg, S.T.; Lunau, T.; Fransson, E.I.; Alfredsson, L.; Bjorner, J.B.; Borritz, M.; Burr, H.; Erbel, R.; et al. Effort-Reward Imbalance at Work and Incident Coronary Heart Disease: A Multicohort Study of 90,164 Individuals. Epidemiology 2017, 28, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Kivimäki, M.; Jokela, M.; Nyberg, S.T.; Singh-Manoux, A.; Fransson, E.I.; Alfredsson, L.; Bjorner, J.B.; Borritz, M.; Burr, H.; Casini, A.; et al. Long working hours and risk of coronary heart disease and stroke: A systematic review and meta-analysis of published and unpublished data for 603,838 individuals. Lancet 2015, 386, 1739–1746. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xu, S.; Hua, J.; Zhu, D.; Liu, C.; Hu, Y.; Liu, T.; Xu, D. Association between job strain and risk of incident stroke: A meta-analysis. Neurology 2015, 85, 1648–1654. [Google Scholar] [CrossRef]
- Sugita, J.; Fujiu, K.; Nakayama, Y.; Matsubara, T.; Matsuda, J.; Oshima, T.; Liu, Y.; Maru, Y.; Hasumi, E.; Kojima, T.; et al. Cardiac macrophages prevent sudden death during heart stress. Nat. Commun. 2021, 12, 1910. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, R.; Wu, C.; Liang, X.; He, L.; Wang, L.; Wang, X. Activation of the sirtuin silent information regulator 1 pathway inhibits pathological myocardial remodeling. Front. Pharmacol. 2023, 14, 1111320. [Google Scholar] [CrossRef]
- Carrive, P. Dual activation of cardiac sympathetic and parasympathetic components during conditioned fear to context in the rat. Clin. Exp. Pharmacol. Physiol. 2006, 33, 1251–1254. [Google Scholar] [CrossRef]
- Chrousos, G.P.; Gold, P.W. The concepts of stress and stress system disorders: Overview of physical and behavioral homeostasis. JAMA 1992, 267, 1244–1252. [Google Scholar] [CrossRef]
- Lampert, R. Mental Stress and Ventricular Arrhythmias. Curr. Cardiol. Rep. 2016, 18, 118. [Google Scholar] [CrossRef]
- Osadchii, O.E. Role of abnormal repolarization in the mechanism of cardiac arrhythmia. Acta Physiol. 2017, 220, 1–71. [Google Scholar] [CrossRef]
- Monitillo, F. Ventricular repolarization measures for arrhythmic risk stratification. World J. Cardiol. 2016, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Ravens, U.; Peyronnet, R. Electrical Remodelling in Cardiac Disease. Cells 2023, 12, 230. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, C.; Chen, X.; Liu, X.; Ye, T.; Fo, Y.; Shi, S.; Qu, C.; Liang, J.; Shen, B.; et al. Sigma-1 receptor ligands improves ventricular repolarization-related ion remodeling in rats with major depression disorder. Psychopharmacology 2021, 238, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Zhou, Y.; Yang, J.; Yu, F.; Song, Z.; Shi, J.; Wang, L.; Huang, Z.; Yang, B.; Wang, X. P2X7 receptor inhibition prevents atrial fibrillation in rodent models of depression. Europace 2024, 26, euae022. [Google Scholar] [CrossRef]
- Liu, X.; Shi, S.; Yang, H.; Qu, C.; Chen, Y.; Liang, J.; Yang, B. The activation of N-methyl-d-aspartate receptors downregulates transient outward potassium and L-type calcium currents in rat models of depression. Am. J. Physiol. Cell Physiol. 2017, 313, C187–C196. [Google Scholar] [CrossRef]
- Cheng, J.H.; Kodama, I. Two components of delayed rectifier K+ current in heart: Molecular basis, functional diversity, and contribution to repolarization. Acta Pharmacol. Sin. 2004, 25, 137–145. [Google Scholar]
- Marchiano, S.; Nakamura, K.; Reinecke, H.; Neidig, L.; Lai, M.; Kadota, S.; Perbellini, F.; Yang, X.; Klaiman, J.M.; Blakely, L.P.; et al. Gene editing to prevent ventricular arrhythmias associated with cardiomyocyte cell therapy. Cell Stem Cell 2023, 30, 396–414. [Google Scholar] [CrossRef]
- Dimai, S.; Semmler, L.; Prabhu, A.; Stachelscheid, H.; Huettemeister, J.; Klaucke, S.C.; Lacour, P.; Blaschke, F.; Kruse, J.; Parwani, A.; et al. COVID19-associated cardiomyocyte dysfunction, arrhythmias and the effect of Canakinumab. PLoS ONE 2021, 16, e0255976. [Google Scholar] [CrossRef]
- Smith, A.S.; Lieberwirth, C.; Wang, Z. Behavioral and physiological responses of female prairie voles (Microtus ochrogaster) to various stressful conditions. Stress 2013, 16, 531–539. [Google Scholar] [CrossRef]
- Jiang, B.; Wang, H.; Wang, J.-L.; Wang, Y.-J.; Zhu, Q.; Wang, C.-N.; Song, L.; Gao, T.-T.; Wang, Y. Hippocampal Salt-Inducible Kinase 2 Plays a Role in Depression via the CREB-Regulated Transcription Coactivator 1–cAMP Response Element Binding–Brain-Derived Neurotrophic Factor Pathway. Biol. Psychiatry 2019, 85, 650–666. [Google Scholar] [CrossRef]
- Handley, S.L.; Mithani, S. Effects of alpha-adrenoceptor agonists and antagonists in a maze-exploration model of ‘fear’-motivated behaviour. Naunyn Schmiedebergs Arch. Pharmacol. 1984, 327, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Strekalova, T.; Spanagel, R.; Bartsch, D.; Henn, F.A.; Gass, P. Stress-Induced Anhedonia in Mice is Associated with Deficits in Forced Swimming and Exploration. Neuropsychopharmacology 2004, 29, 2007–2017. [Google Scholar] [CrossRef] [PubMed]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wu, W.; Xing, J.; Yan, W.; Zhang, J.; Sun, J.; Zhang, Z.; Qiu, S.; Xu, Y.; Wang, X. Berberine attenuates sunitinib-induced cardiac dysfunction by normalizing calcium regulation disorder via SGK1 activation. Food Chem. Toxicol. 2023, 175, 113743. [Google Scholar] [CrossRef]
- Zhang, H.; Fu, T.; Sun, J.; Zou, S.; Qiu, S.; Zhang, J.; Su, S.; Shi, C.; Li, D.-P.; Xu, Y. Pharmacological suppression of Nedd4-2 rescues the reduction of Kv11.1 channels in pathological cardiac hypertrophy. Front. Pharmacol. 2022, 13, 942769. [Google Scholar] [CrossRef]
- Wang, R.; Wang, M.; Wang, S.; Yang, K.; Zhou, P.; Xie, X.; Cheng, Q.; Ye, J.; Sun, G.; Sun, X. An integrated characterization of contractile, electrophysiological, and structural cardiotoxicity of Sophora tonkinensis Gapnep. in human pluripotent stem cell-derived cardiomyocytes. Stem Cell Res. Ther. 2019, 10, 20. [Google Scholar] [CrossRef]
- Wu, W.; Sun, J.; Zhang, J.; Zhao, H.; Qiu, S.; Li, C.; Shi, C.; Xu, Y. Phosphoproteomics reveals a novel mechanism underlying the proarrhythmic effects of nilotinib, vandetanib, and mobocertinib. Toxicology 2024, 505, 153830. [Google Scholar] [CrossRef]
- Cui, X.; Sun, J.; Li, C.; Qiu, S.; Shi, C.; Ma, J.; Xu, Y. Downregulation of hERG channel expression by tyrosine kinase inhibitors nilotinib and vandetanib predominantly contributes to arrhythmogenesis. Toxicol. Lett. 2022, 365, 11–23. [Google Scholar] [CrossRef]
- Doerr, L.; Thomas, U.; Guinot, D.R.; Bot, C.T.; Stoelzle-Feix, S.; Beckler, M.; George, M.; Fertig, N. New easy-to-use hybrid system for extracellular potential and impedance recordings. J. Lab. Autom. 2015, 20, 175–188. [Google Scholar] [CrossRef]
- Blinova, K.; Dang, Q.; Millard, D.; Smith, G.; Pierson, J.; Guo, L.; Brock, M.; Lu, H.R.; Kraushaar, U.; Zeng, H.; et al. International Multisite Study of Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes for Drug Proarrhythmic Potential Assessment. Cell Rep. 2018, 24, 3582–3592. [Google Scholar] [CrossRef]
- Bot, C.T.; Juhasz, K.; Haeusermann, F.; Polonchuk, L.; Traebert, M.; Stoelzle-Feix, S. Cross—Site comparison of excitation-contraction coupling using impedance and field potential recordings in hiPSC cardiomyocytes. J. Pharmacol. Toxicol. Methods 2018, 93, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Faggioni, M.; Hwang, H.S.; van der Werf, C.; Nederend, I.; Kannankeril, P.J.; Wilde, A.A.; Knollmann, B.C. Accelerated sinus rhythm prevents catecholaminergic polymorphic ventricular tachycardia in mice and in patients. Circ. Res. 2013, 112, 689–697. [Google Scholar] [CrossRef]
- van der Werf, C.; Kannankeril, P.J.; Sacher, F.; Krahn, A.D.; Viskin, S.; Leenhardt, A.; Shimizu, W.; Sumitomo, N.; Fish, F.A.; Bhuiyan, Z.A.; et al. Flecainide therapy reduces exercise-induced ventricular arrhythmias in patients with catecholaminergic polymorphic ventricular tachycardia. J. Am. Coll. Cardiol. 2011, 57, 2244–2254. [Google Scholar] [CrossRef] [PubMed]
- Tran, I.; Gellner, A.-K. Long-term effects of chronic stress models in adult mice. J. Neural Transm. 2023, 130, 1133–1151. [Google Scholar] [CrossRef] [PubMed]
- Doron, R.; Burstein, O. The Unpredictable Chronic Mild Stress Protocol for Inducing Anhedonia in Mice. J. Vis. Exp. 2018, 140, 58184. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, C.; Guo, Y.; Liu, X.; Ye, T.; Fo, Y.; Qu, C.; Liang, J.; Shi, S.; Yang, B. Chronic stimulation of the sigma-1 receptor ameliorates ventricular ionic and structural remodeling in a rodent model of depression. Life Sci. 2020, 257, 118047. [Google Scholar] [CrossRef]
- Roden, D.M. Taking the “idio” out of “idiosyncratic”: Predicting torsades de pointes. Pacing Clin. Electrophysiol. PACE 1998, 21, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Varró, A.; Baczkó, I. Cardiac ventricular repolarization reserve: A principle for understanding drug-related proarrhythmic risk. Br. J. Pharmacol. 2011, 164, 14–36. [Google Scholar] [CrossRef]
- Sanguinetti, M.C.; Jurkiewicz, N.K. Two components of cardiac delayed rectifier K+ current. Differential sensitivity to block by class III antiarrhythmic agents. J. Gen. Physiol. 1990, 96, 195–215. [Google Scholar] [CrossRef]
- Zeng, J.; Laurita, K.R.; Rosenbaum, D.S.; Rudy, Y. Two components of the delayed rectifier K+ current in ventricular myocytes of the guinea pig type: Theoretical formulation and their role in repolarization. Circ. Res. 1995, 77, 140–152. [Google Scholar] [CrossRef]
- Bosch, R.F.; Gaspo, R.; Busch, A.E.; Lang, H.J.; Li, G.R.; Nattel, S. Effects of the chromanol 293B, a selective blocker of the slow, component of the delayed rectifier K+ current, on repolarization in human and guinea pig ventricular myocytes. Cardiovasc. Res. 1998, 38, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Kamiya, K.; Opthof, T.; Yasui, K.; Kodama, I. Density and kinetics of I(Kr) and I(Ks) in guinea pig and rabbit ventricular myocytes explain different efficacy of I(Ks) blockade at high heart rate in guinea pig and rabbit: Implications for arrhythmogenesis in humans. Circulation 2001, 104, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Guérard, N.C.; Traebert, M.; Suter, W.; Dumotier, B.M. Selective block of IKs plays a significant role in MAP triangulation induced by IKr block in isolated rabbit heart. J. Pharmacol. Toxicol. Methods 2008, 58, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Varro, A.; Baláti, B.; Iost, N.; Takács, J.; Virág, L.; Lathrop, D.A.; Csaba, L.; Tálosi, L.; Papp, J.G. The role of the delayed rectifier component IKs in dog ventricular muscle and Purkinje fibre repolarization. J. Physiol. 2000, 523 Pt 1, 67–81. [Google Scholar] [CrossRef]
- Stengl, M.; Volders, P.G.; Thomsen, M.B.; Spätjens, R.L.; Sipido, K.R.; Vos, M.A. Accumulation of slowly activating delayed rectifier potassium current (IKs) in canine ventricular myocytes. J. Physiol. 2003, 551, 777–786. [Google Scholar] [CrossRef]
- Jost, N.; Virág, L.; Comtois, P.; Ordög, B.; Szuts, V.; Seprényi, G.; Bitay, M.; Kohajda, Z.; Koncz, I.; Nagy, N.; et al. Ionic mechanisms limiting cardiac repolarization reserve in humans compared to dogs. J. Physiol. 2013, 591, 4189–4206. [Google Scholar] [CrossRef]
- Iravanian, S.; Dudley, S.C., Jr. The renin-angiotensin-aldosterone system (RAAS) and cardiac arrhythmias. Heart Rhythm 2008, 5, S12–S17. [Google Scholar] [CrossRef]
- Prajapati, C.; Ojala, M.; Aalto-Setälä, K. Divergent effects of adrenaline in human induced pluripotent stem cell-derived cardiomyocytes obtained from hypertrophic cardiomyopathy. Dis. Models Mech. 2018, 11, dmm032896. [Google Scholar] [CrossRef]
- Eden, E.; Teirstein, A.; Wiener, I. Ventricular arrhythmia induced by vasopressin: Torsade de pointes related to vasopressin-induced bradycardia. Mt. Sinai J. Med. New York 1983, 50, 49–51. [Google Scholar]
- Herrmann, J. Adverse cardiac effects of cancer therapies: Cardiotoxicity and arrhythmia. Nat. Rev. Cardiol. 2020, 17, 474–502. [Google Scholar] [CrossRef]
- Larsson, S.C.; Lee, W.H.; Burgess, S.; Allara, E. Plasma Cortisol and Risk of Atrial Fibrillation: A Mendelian Randomization Study. J. Clin. Endocrinol. Metab. 2021, 106, e2521–e2526. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, T.; Shufelt, C.; Iribarren, C.; Merz, C.N. Sex hormones and the QT interval: A review. J. Women’s Health 2012, 21, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Billman, G.E.; Carnes, C.A.; Adamson, P.B.; Vanoli, E.; Schwartz, P.J. Dietary omega-3 fatty acids and susceptibility to ventricular fibrillation: Lack of protection and a proarrhythmic effect. Circ. Arrhythmia Electrophysiol. 2012, 5, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, A.O.; van Ginneken, A.C.; Berecki, G.; den Ruijter, H.M.; Schumacher, C.A.; Veldkamp, M.W.; Baartscheer, A.; Casini, S.; Opthof, T.; Hovenier, R.; et al. Incorporated sarcolemmal fish oil fatty acids shorten pig ventricular action potentials. Cardiovasc. Res. 2006, 70, 509–520. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, S.; Sun, J.; Qiu, S.; Wu, W.; Zhang, J.; Wang, Y.; Shi, C.; Xu, Y. Pro-Arrhythmic Effect of Chronic Stress-Associated Humoral Factors in Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Biology 2025, 14, 652. https://doi.org/10.3390/biology14060652
Su S, Sun J, Qiu S, Wu W, Zhang J, Wang Y, Shi C, Xu Y. Pro-Arrhythmic Effect of Chronic Stress-Associated Humoral Factors in Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Biology. 2025; 14(6):652. https://doi.org/10.3390/biology14060652
Chicago/Turabian StyleSu, Shi, Jinglei Sun, Suhua Qiu, Wenting Wu, Jiali Zhang, Yi Wang, Chenxia Shi, and Yanfang Xu. 2025. "Pro-Arrhythmic Effect of Chronic Stress-Associated Humoral Factors in Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes" Biology 14, no. 6: 652. https://doi.org/10.3390/biology14060652
APA StyleSu, S., Sun, J., Qiu, S., Wu, W., Zhang, J., Wang, Y., Shi, C., & Xu, Y. (2025). Pro-Arrhythmic Effect of Chronic Stress-Associated Humoral Factors in Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Biology, 14(6), 652. https://doi.org/10.3390/biology14060652