
Fire and Drought Affect Multiple Aspects of Diversity in a Migratory Bird Stopover Community
 , ,
, , {kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Bird Captures
2.3. Vegetation Data
2.4. Statistical Analyses
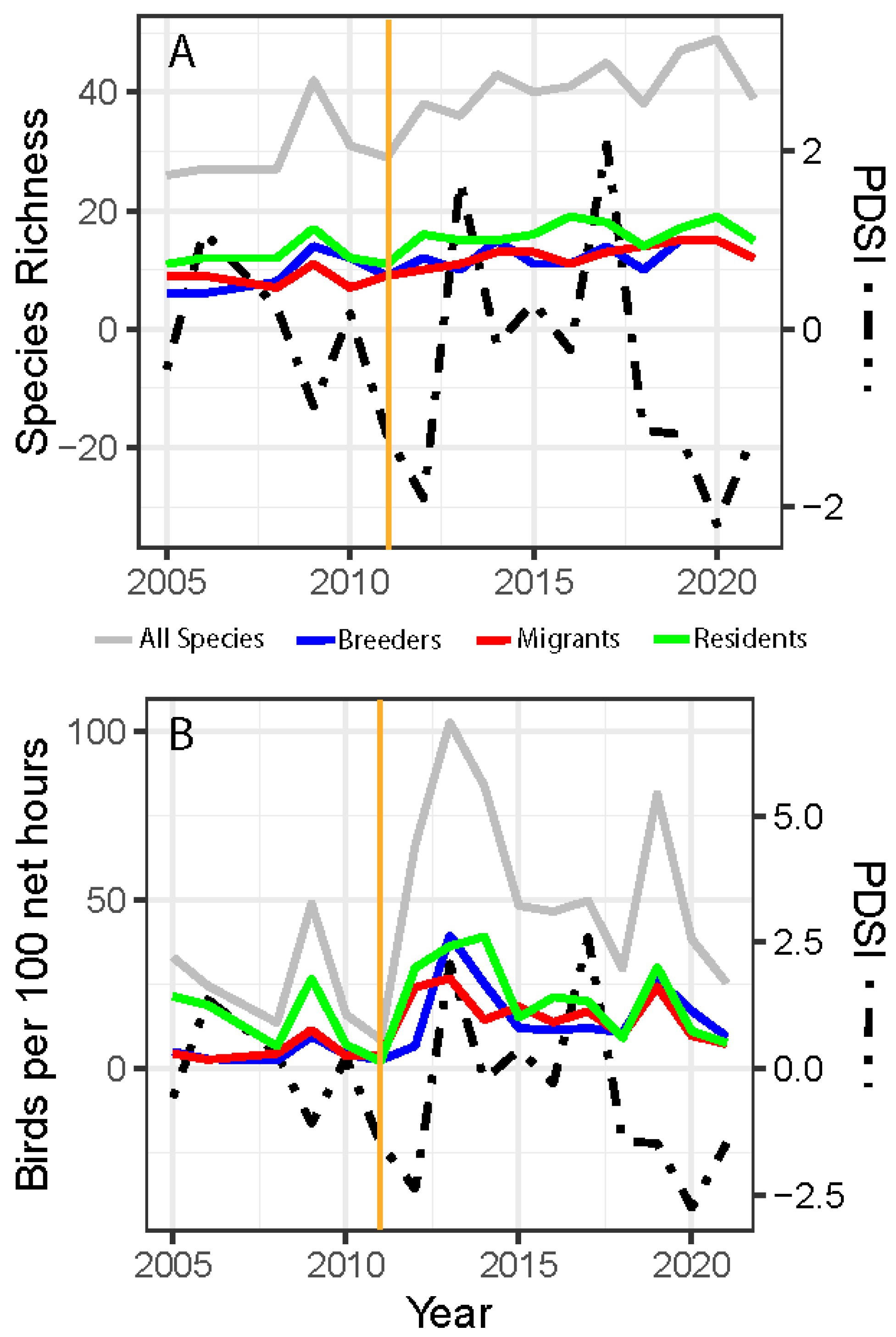
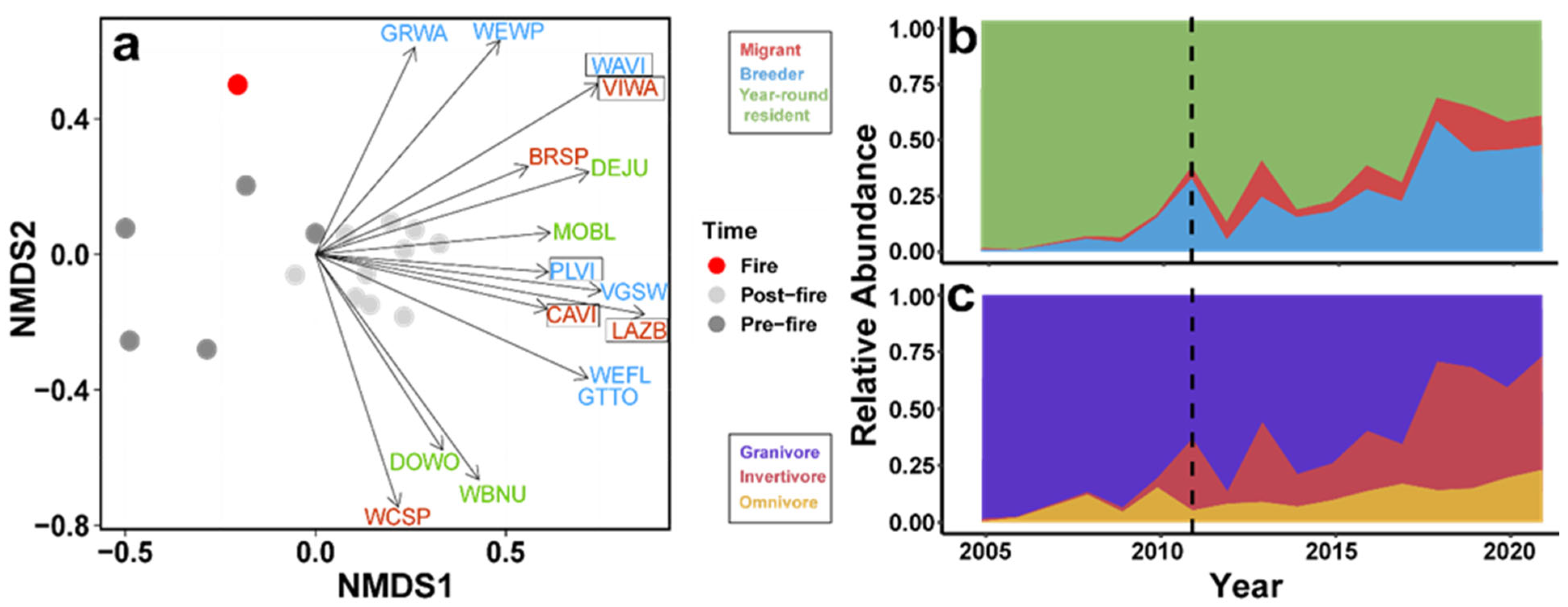
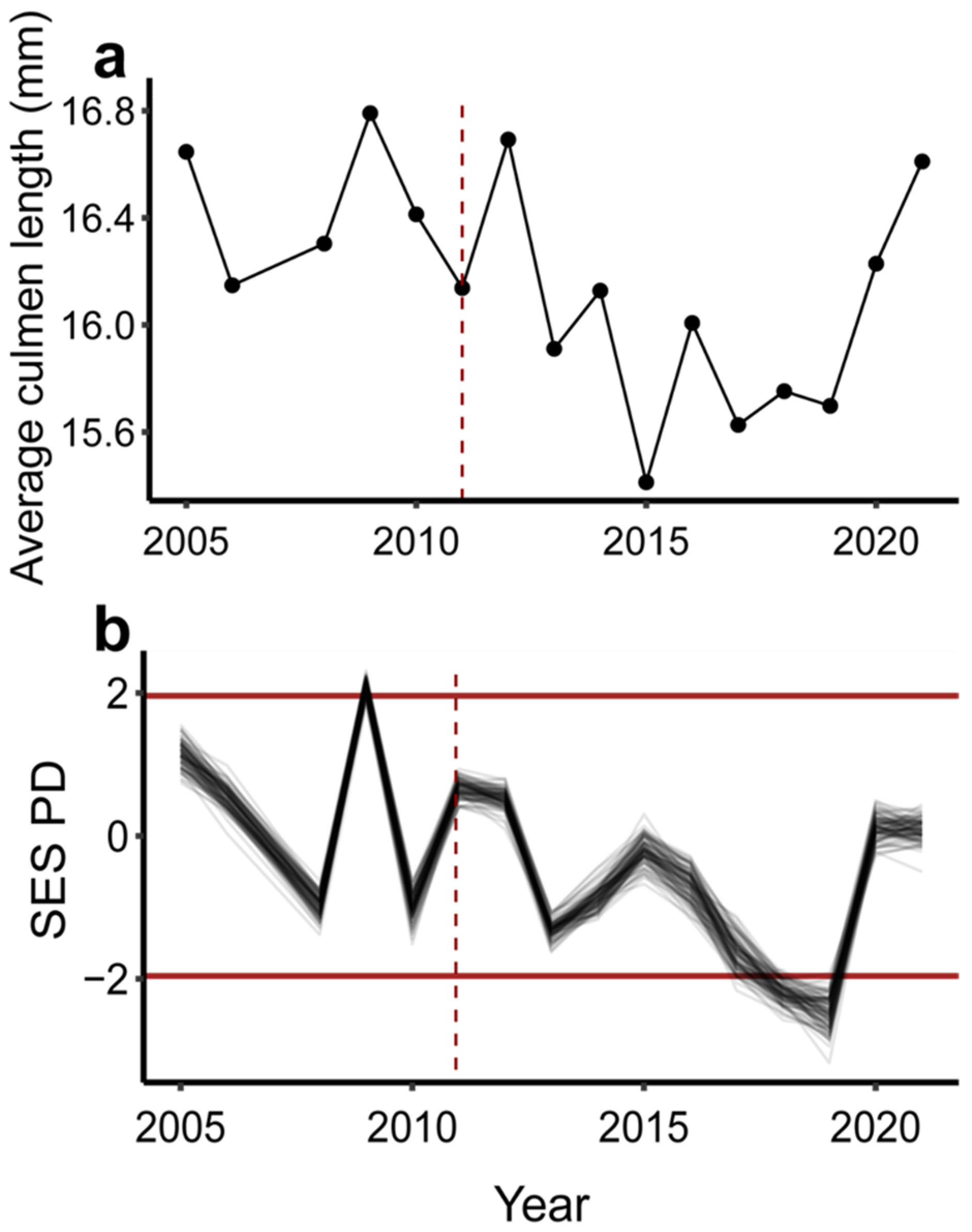
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOSIM | Analysis of Similarities |
| DBH | diameter at breast height |
| MAD | mean absolute deviation |
| NMDS | nonmetric multidimensional scaling |
| PCoA | Principal Coordinate Analysis |
| PD | phylogenetic diversity |
| PDSI | Palmer Drought Severity Index |
| SES | standard effect sizes |
References
- Chen, I.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.; Crick, H.; Learmonth, J.; Maclean, I.; Thomas, C.; Bairlein, F.; Forchhammer, M.; Francis, C.; Gill, J.; Godley, B.; et al. Travelling through a warming world: Climate change and migratory species. Endanger. Species Res. 2009, 7, 87–99. [Google Scholar] [CrossRef]
- Swetnam, T.W.; Farella, J.; Roos, C.I.; Liebmann, M.J.; Falk, D.A.; Allen, C.D. Multiscale perspectives of fire, climate and humans in western North America and the Jemez Mountains, USA. Phil. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150168. [Google Scholar] [CrossRef]
- Crockett, J.L.; Westerling, A.L. Greater temperature and precipitation extremes intensify western U.S. droughts, wildfire severity, and Sierra Nevada tree mortality. J. Clim. 2018, 31, 341–354. [Google Scholar] [CrossRef]
- Wasserman, T.N.; Mueller, S.E. Climate influences on future fire severity: A synthesis of climate-fire interactions and impacts on fire regimes, high-severity fire, and forests in the western United States. Fire Ecol. 2023, 19, 1–22. [Google Scholar] [CrossRef]
- Jones, G.M.; Shirk, A.J.; Yang, Z.; Davis, R.J.; Ganey, J.L.; Gutiérrez, R.J.; Healey, S.; Hedwall, S.; Hoagland, S.; Maes, R.; et al. Spatial and temporal dynamics of Mexican spotted owl habitat in the southwestern US. Landsc. Ecol. 2023, 38, 7–22. [Google Scholar]
- Albright, T.P.; Pidgeon, A.M.; Rittenhouse, C.D.; Clayton, M.K.; Flather, C.H.; Culbert, P.D.; Wardlow, B.D.; Radeloff, V.C. Effects of drought on avian community structure. Glob. Chang. Biol. 2010, 16, 2158–2170. [Google Scholar] [CrossRef]
- Albright, T.P.; Pidgeon, A.M.; Rittenhouse, C.D.; Clayton, M.K.; Wardlow, B.D.; Flather, C.H.; Culbert, P.D.; Radeloff, V.C. Combined effects of heat waves and droughts on avian communities across the conterminous United States. Ecosphere 2010, 1, 1–22. [Google Scholar] [CrossRef]
- Saracco, J.F.; Fettig, S.M.; San Miguel, G.L.; Mehlman, D.W.; Thompson, B.E.; Albert, S.K. Avian demographic responses to drought and fire: A community-level perspective. Ecol. Appl. 2018, 28, 1773–1781. [Google Scholar] [CrossRef]
- Selwood, K.E.; McGeoch, M.A.; Clarke, R.H.; Mac Nally, R. High-productivity vegetation is important for lessening bird declines during prolonged drought. J. Appl. Ecol. 2018, 55, 641–650. [Google Scholar] [CrossRef]
- Smucker, K.M.; Hutto, R.L.; Steele, B.M. Changes in bird abundance after wildfire: Importance of fire severity and time since fire. Ecol. Appl. 2005, 15, 1535–1549. [Google Scholar] [CrossRef]
- Steel, Z.L.; Fogg, A.M.; Burnett, R.; Roberts, L.J.; Safford, H.D. When bigger isn’t better—Implications of large high-severity wildfire patches for avian diversity and community composition. Divers. Distrib. 2021, 28, 439–453. [Google Scholar] [CrossRef]
- Earnst, S.L.; Newsome, H.L.; LaFramboise, W.L.; LaFramboise, N. Avian response to wildfire in interior Columbian basin shrubsteppe. Condor Ornithol. Appl. 2009, 111, 370–376. [Google Scholar]
- Taillie, P.J.; Burnett, R.D.; Roberts, L.J.; Campos, B.R.; Peterson, M.N.; Moorman, C.E. Interacting and non-linear avian responses to mixed-severity wildfire and time since fire. Ecosphere 2018, 9, e02291. [Google Scholar] [CrossRef]
- Burbidge, A.A.; Fuller, P.J. Gibson Desert birds: Responses to drought and plenty. Emu Austral Ornithol. 2007, 107, 126–134. [Google Scholar] [CrossRef]
- Stanek, J.E.; Thompson, B.E.; Milligan, S.E.; Tranquillo, K.A.; Fettig, S.M.; Hathcock, C.D. Does age, residency, or feeding guild coupled with a drought index predict avian health during fall migration? Animals 2022, 12, 454. [Google Scholar] [CrossRef]
- Stevens, H.C.; Watson, D.M. Interacting impacts of drought and fire on bird populations—Insights from a long-term study in the Warrumbungles. Aust. Zool. 2022, 42, 608–630. [Google Scholar] [CrossRef]
- Crockett, J.L.; Hurteau, M.D. Post-fire early successional vegetation buffers surface microclimate and increases survival of planted conifer seedlings in the southwestern United States. Can. J. For. Res. 2021, 52, 416–425. [Google Scholar] [CrossRef]
- Zlonis, E.J.; Walton, N.G.; Sturtevant, B.R.; Wolter, P.T.; Niemi, G.J. Burn severity and heterogeneity mediate avian response to wildfire in a hemiboreal forest. For. Ecol. Manage. 2019, 439, 70–80. [Google Scholar] [CrossRef]
- McWilliams, S.R.; Guglielmo, C.; Pierce, B.; Klaassen, M. Flying, fasting, and feeding in birds during migration: A nutritional and physiological ecology perspective. J. Avian Biol. 2004, 35, 377–393. [Google Scholar] [CrossRef]
- Klaassen, R.H.G.; Hake, M.; Strandberg, R.; Koks, B.J.; Trierweiler, C.; Exo, K.; Bairlein, F.; Alerstam, T. When and where does mortality occur in migratory birds? Direct evidence from long-term satellite tracking of raptors. J. Anim. Ecol. 2014, 83, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Sergio, F.; Tavecchia, G.; Tanferna, A.; Blas, J.; Blanco, G.; Hiraldo, F. When and where mortality occurs throughout the annual cycle changes with age in a migratory bird: Individual vs. population implications. Sci. Rep. 2019, 9, 17352. [Google Scholar] [CrossRef] [PubMed]
- Gordo, O. Why are bird migration dates shifting? A review of weather and climate effects on avian migratory phenology. Clim. Res. 2007, 35, 37–58. [Google Scholar] [CrossRef]
- Haest, B.; Hüppop, O.; van de Pol, M.; Bairlein, F. Autumn bird migration phenology: A potpourri of wind, precipitation and temperature effects. Glob. Change Biol. 2019, 25, 4064–4080. [Google Scholar] [CrossRef] [PubMed]
- Kendeigh, S.C.; Dol’Nik, V.R.; Gavrilov, V.M.; Pinowski, J. Avian energetics. In Granivorous Birds in Ecosystems: Their Evolution, Populations, Energetics, Adaptations, Impacts and Control (International Biological Programme 12), 1st ed.; Pinowski, J., Kendeigh, S.C., Eds.; Cambridge University Press: New York, NY, USA, 1977; pp. 127–203. [Google Scholar]
- Fretwell, S.D. Evolution of migration in relation to factors regulating bird numbers. In Migrant Birds in the Neotropics: Ecology, Behavior, Distribution, and Conservation, 1st ed.; Keast, A., Morton, E.S., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1980; pp. 517–527. [Google Scholar]
- Levey, D.J.; Stiles, F.G. Evolutionary precursors of long-distance migration: Resource availability and movement patterns in Neotropical landbirds. Am. Nat. 1992, 140, 447–476. [Google Scholar] [CrossRef]
- Wiener, P.; Tuljapurkar, S. Migration in variable environments: Exploring life-history evolution using structured population models. J. Theor. Biol. 1994, 166, 75–90. [Google Scholar] [CrossRef]
- Kittelberger, K.D.; Miller, M.K.; Şekercioğlu, C.H. Fall bird migration in western North America during a period of heightened wildfire activity. Avian Conserv. Ecol. 2022, 17, 43. [Google Scholar] [CrossRef]
- Boucek, R.E.; Rehage, J.S. Climate extremes drive changes in functional community structure. Glob. Change Biol. 2014, 20, 1821–1831. [Google Scholar] [CrossRef]
- Barnagaud, J.Y.; Gaüzère, P.; Zuckerberg, B.; Princé, K.; Svenning, J.C. Temporal changes in bird functional diversity across the United States. Oecologia 2017, 185, 737–748. [Google Scholar] [CrossRef]
- Docherty, T.D.S.; Hethcoat, M.G.; MacTavish, L.M.; MacTavish, D.; Dell, S.; Stephens, P.A.; Willis, S.G. Burning savanna for avian species richness and functional diversity. Ecol. Appl. 2020, 30, e02091. [Google Scholar] [CrossRef]
- Knaggs, M.; Haché, S.; Nielsen, S.E.; Pankratz, R.F.; Bayne, E. Avian response to wildfire severity in a northern boreal region. Forests 2020, 11, 1330. [Google Scholar] [CrossRef]
- Ocampo-Zuleta, K.; Parrado-Rosselli, Á. Functional diversity in an Andean subpáramo affected by wildfire in Colombia. Plant Divers. 2023, 45, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Hussell, D.J.; Ralph, C.J. Recommended methods for monitoring change in landbird populations by counting and capturing migrants. North Am. Bird Bander 2005, 30, 2. [Google Scholar]
- Fair, J.M.; Paul, E.; Jones, J.; Bies, L. (Eds.) Guidelines to the Use of Wild Birds in Research, 4th ed.; The Ornithological Council: Washington, DC, USA, 2023; pp. 42–54. [Google Scholar]
- Pyle, P. Identification Guide to North American Passerines; Slate Creek Press: Bolinas, CA, USA, 1987; p. 273. Volume 1. [Google Scholar]
- Karr, J.R. Surveying birds with mist nets. Stud. Avian Biol. 1981, 6, 62–67. [Google Scholar]
- Billerman, S.M.; Keeney, B.K.; Rodewald, P.G.; Schulenberg, T.S. (Eds.) Birds of the World; Cornell Laboratory of Ornithology: Ithaca, NY, USA, 2022. [Google Scholar]
- NOAA National Centers for Environmental Information, Climate at a Glance: National Time Series. National Oceanic and Atmospheric Administration, Published March 2025. Available online: https://www.ncei.noaa.gov/access/monitoring/climate-at-a-glance/national/time-series (accessed on 20 March 2025).
- Clarke, K.R.; Green, R.H. Statistical design and analysis for a ‘biological effects’ study. Mar. Ecol. Prog. Ser. 1988, 41, 213–226. [Google Scholar] [CrossRef]
- Clarke, K.R. Nonparametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. vegan: Community Ecology Package. R Package Version 2.6-8. 2025. Available online: https://CRAN.R-project.org/package=vegan (accessed on 20 March 2025).
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- McCune, B.P.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Tobias, J.A.; Sheard, C.; Pigot, A.L.; Devenish, A.J.M.; Yang, J.; Sayol, F.; Neate-Clegg, M.H.C.; Alioravainen, N.; Weeks, T.L.; Barber, R.A.; et al. AVONET: Morphological, ecological and geographical data for all birds. Ecol. Lett. 2022, 25, 581–597. [Google Scholar] [CrossRef]
- Magneville, C.; Loiseau, N.; Albouy, C.; Casajus, N.; Claverie, T.; Escalas, A.; Leprieur, F.; Maire, E.; Mouillot, D.; Villéger, S. mFD: An R package to compute and illustrate the multiple facets of functional diversity. Ecography 2022, 2022, 1–15. [Google Scholar] [CrossRef]
- Swenson, N.G. Functional and Phylogenetic Ecology in R; Springer: New York, NY, USA, 2014; Volume 639. [Google Scholar]
- Ortega-Martínez, I.J.; Moreno, C.E.; Rios-Díaz, C.L.; Arellano, L.; Rosas, F.; Castellanos, I. Assembly mechanisms of dung beetles in temperate forests and grazing pastures. Sci. Rep. 2020, 10, 391. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org/ (accessed on 20 March 2025).
- Komsta, L. Package ‘mblm’ Median-Based Linear Models. R Package Version 0.12.1. 2019. Available online: https://cran.r-project.org/web/packages/mblm/mblm.pdf (accessed on 20 March 2025).
- Robertson, E.P.; La Sorte, F.A.; Mays, J.D.; Taillie, P.J.; Robinson, O.J.; Ansley, R.J.; O’connell, T.J.; Davis, C.A.; Loss, S.R. Decoupling of bird migration from the changing phenology of spring green-up. Proc. Natl. Acad. Sci. USA 2024, 121, e2308433121. [Google Scholar] [CrossRef]
- Wan, S.; Hui, D.; Luo, Y. Fire effects on nitrogen pools and dynamics in terrestrial ecosystems: A meta-analysis. Ecol. Appl. 2001, 11, 1349–1365. [Google Scholar] [CrossRef]
- Smith, E.A.; O’Loughlin, D.; Buck, J.R.; St Clair, S.B. The influences of conifer succession, physiographic conditions and herbivory on quaking aspen regeneration after fire. For. Ecol. Manage. 2011, 262, 325–330. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Reich, P.B. Shocks to the system: Community assembly of the oak savanna in a 40-year fire frequency experiment. Ecology 2012, 93, S52–S69. [Google Scholar] [CrossRef]
- Bender, E.A.; Case, T.J.; Gilpin, M.E. Perturbation experiments in community ecology: Theory and practice. Ecology 1984, 65, 1–13. [Google Scholar] [CrossRef]
- Ives, A.R. Predicting the response of populations to environmental change. Ecology 1995, 76, 926–941. [Google Scholar] [CrossRef]
- Puig-Gironès, R.; Brotons, L.; Pons, P. Aridity influences the recovery of vegetation and shrubland birds after wildfire. PLoS ONE 2017, 12, e0173599. [Google Scholar] [CrossRef]
- Song, C.; Woodcock, C.E. Monitoring forest succession with multitemporal Landsat images: Factors of uncertainty. IEEE Trans. Geosci. Remote Sens. 2003, 41, 2557–2567. [Google Scholar] [CrossRef]
- Lentile, L.B.; Holden, Z.A.; Smith, A.M.S.; Falkowski, M.J.; Hudak, A.T.; Morgan, P.; Lewis, S.A.; Gessler, P.E.; Benson, N.C. Remote sensing techniques to assess active fire characteristics and post-fire effects. Int. J. Wildland Fire 2006, 15, 319–345. [Google Scholar] [CrossRef]
- Rainsford, F.W.; Giljohann, K.M.; Bennett, A.F.; Clarke, M.F.; MacHunter, J.; Senior, K.; Sitters, H.; Watson, S.; Kelly, L.T. Ecosystem type and species’ traits help explain bird responses to spatial patterns of fire. Fire Ecol. 2023, 19, 59. [Google Scholar] [CrossRef]
- Bailey, J.K.; Whitman, T.G. Interactions among fire, aspen, and elk affect insect diversity: Reversal of a community response. Ecology 2002, 83, 1701–1712. [Google Scholar] [CrossRef]
- Wenninger, A.; Hollingsworth, T.; Wagner, D. Predatory hymenopteran assemblages in boreal Alaska: Associations with forest composition and post-fire succession. Ecoscience 2019, 26, 205–220. [Google Scholar] [CrossRef]
- Rainsford, F.W.; Kelly, L.T.; Leonard, S.W.J.; Bennett, A.F. Fire and functional traits: Using functional groups of birds and plants to guide management in a fire-prone, heathy woodland ecosystem. Divers. Distrib. 2021, 28, 372–385. [Google Scholar] [CrossRef]
- Rainsford, F.W.; Kelly, L.T.; Leonard, S.W.J.; Bennett, A.F. Post-fire habitat relationships for birds differ among ecosystems. Biol. Conserv. 2021, 260, 109218. [Google Scholar] [CrossRef]
- Veit, R.R. Do vagrant birds in Massachusetts reflect population growth and dispersal rather than weather patterns? Bird Obs. 1990, 18, 5. [Google Scholar]
- Simpson, E.H. The interpretation of interaction in contingency tables. J. R. Stat. Soc. Ser. B 1951, 13, 238–241. [Google Scholar] [CrossRef]
- Remsen, J.V., Jr.; Good, D.A. Misuse of data from mist-net captures to assess relative abundance in bird populations. Auk 1996, 113, 381–398. [Google Scholar] [CrossRef]
- Gómez, C.; Tenorio, E.A.; Montoya, P.; Cadena, C.D. Niche-tracking migrants and niche-switching residents: Evolution of climatic niches in New World warblers (Parulidae). Proc. R. Soc. B Biol. Sci. 2016, 283, 2015–2458. [Google Scholar] [CrossRef]
- Zurell, D.; Gallien, L.; Graham, C.H.; Zimmermann, N.E. Do long-distance migratory birds track their niche through seasons? J. Biogeogr. 2018, 45, 1459–1468. [Google Scholar] [CrossRef]
- Fandos, G.; Tellería, J.L. Seasonal niche-tracking behaviour of two partially migratory passerines. Ibis 2020, 162, 307–317. [Google Scholar] [CrossRef]
- Lindstrom, Å. The role of predation risk in stopover habitat selection in migrating bramblings, Fringilla montifringilla. Behav. Ecol. 1990, 1, 102–106. [Google Scholar] [CrossRef]
- Moore, F.R.; Aborn, D.A. Mechanisms of en route habitat selection: How do migrants make habitat decisions during stopover? Stud. Avian Biol. 2000, 20, 34–42. [Google Scholar]
- Cohen, E.B.; Horton, K.G.; Marra, P.P.; Clipp, H.L.; Farnsworth, A.; Smolinsky, J.A.; Sheldon, D.; Buler, J.J. A place to land: Spatiotemporal drivers of stopover habitat use by migrating birds. Ecol. Lett. 2021, 24, 38–49. [Google Scholar] [CrossRef]
- Cady, S.M.; O’Connell, T.J.; Loss, S.R.; Jaffe, N.E.; Davis, C.A. Species-specific and temporal scale-dependent responses of birds to drought. Glob. Change Biol. 2019, 25, 2691–2702. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanek, J.E.; Gadek, C.R.; Milligan, S.E.; Thompson, B.E.; Tranquillo, K.A.; Trader, L.L.; Hathcock, C.D.; Fettig, S.M. Fire and Drought Affect Multiple Aspects of Diversity in a Migratory Bird Stopover Community. Biology 2025, 14, 597. https://doi.org/10.3390/biology14060597
Stanek JE, Gadek CR, Milligan SE, Thompson BE, Tranquillo KA, Trader LL, Hathcock CD, Fettig SM. Fire and Drought Affect Multiple Aspects of Diversity in a Migratory Bird Stopover Community. Biology. 2025; 14(6):597. https://doi.org/10.3390/biology14060597
Chicago/Turabian StyleStanek, Jenna E., Chauncey R. Gadek, Sarah E. Milligan, Brent E. Thompson, Keegan A. Tranquillo, Laura L. Trader, Charles D. Hathcock, and Stephen M. Fettig. 2025. "Fire and Drought Affect Multiple Aspects of Diversity in a Migratory Bird Stopover Community" Biology 14, no. 6: 597. https://doi.org/10.3390/biology14060597
APA StyleStanek, J. E., Gadek, C. R., Milligan, S. E., Thompson, B. E., Tranquillo, K. A., Trader, L. L., Hathcock, C. D., & Fettig, S. M. (2025). Fire and Drought Affect Multiple Aspects of Diversity in a Migratory Bird Stopover Community. Biology, 14(6), 597. https://doi.org/10.3390/biology14060597