
Revalidation of DNA Fragmentation Analyses for Human Sperm—Measurement Principles, Comparative Standards, Calibration Curve, Required Sensitivity, and Eligibility Criteria for Test Sperm

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Overview
2. First Step of Qualitative-Method Validation—Establishment of Comparative Standards
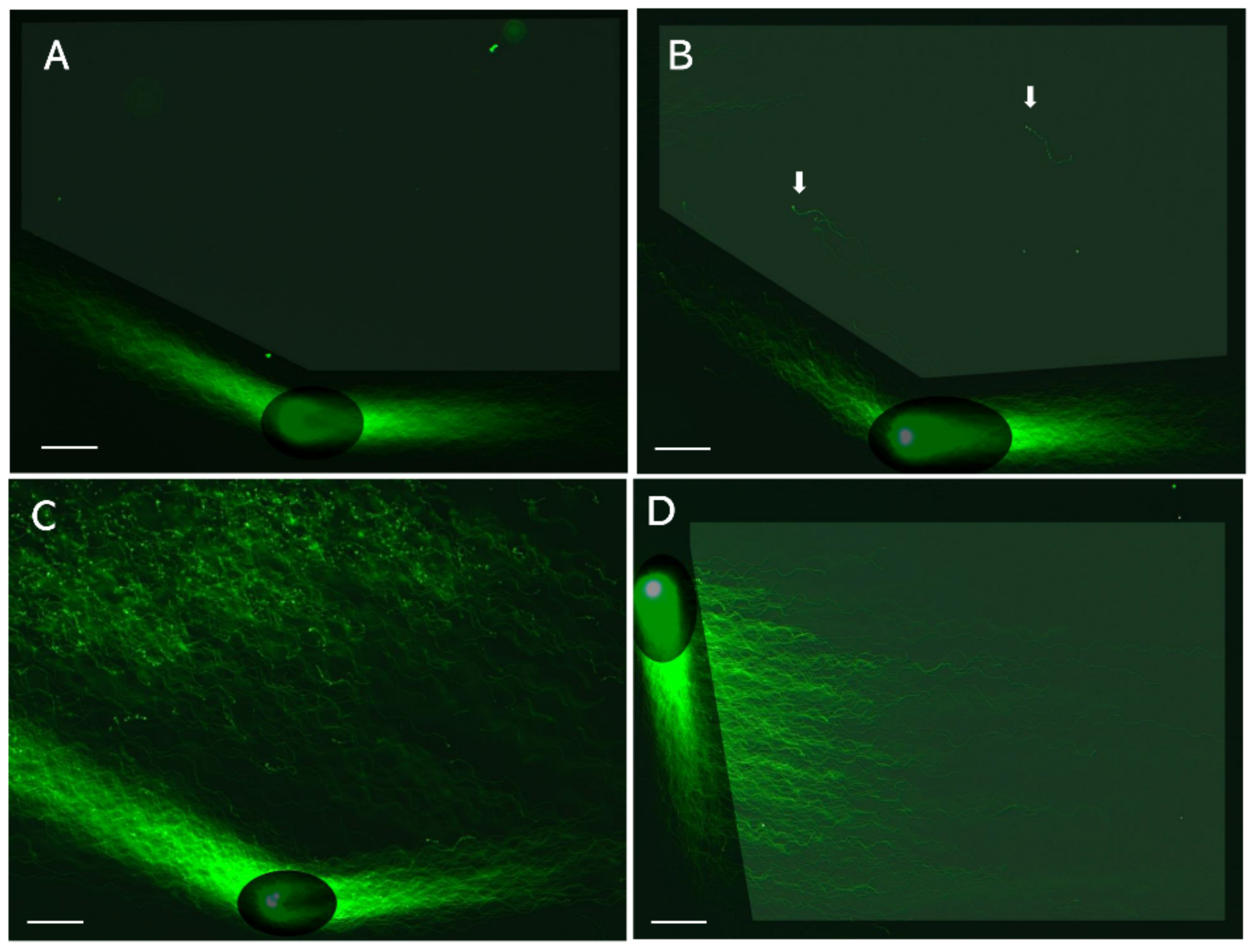
3. 1D-SCPFGE
4. Conventional Analytical Methods for DNA Fragmentation Do Not Pass the First Step of Qualitative Validation
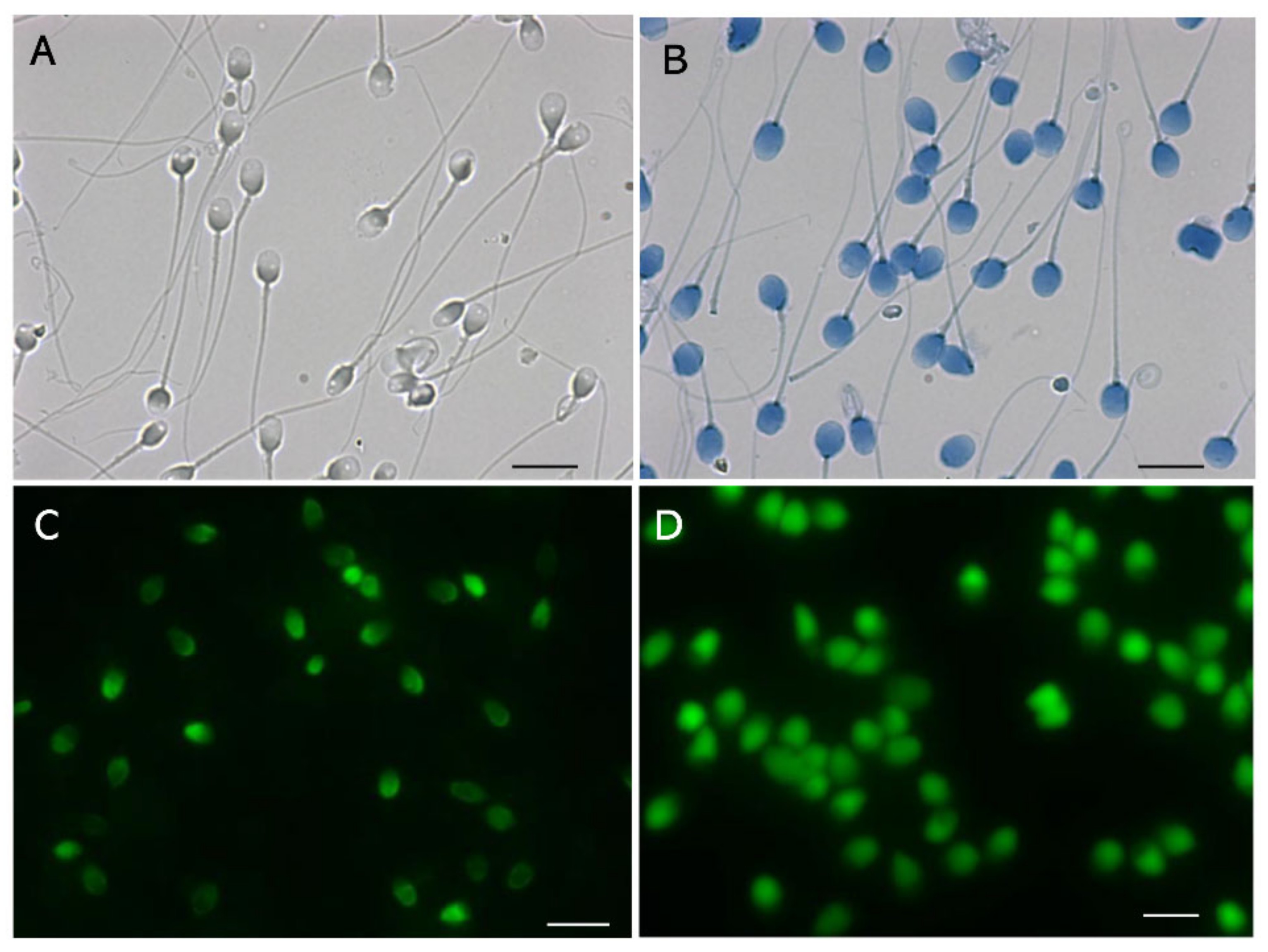
4.1. Lack of Proteolysis Produces False Negative Results in the CA
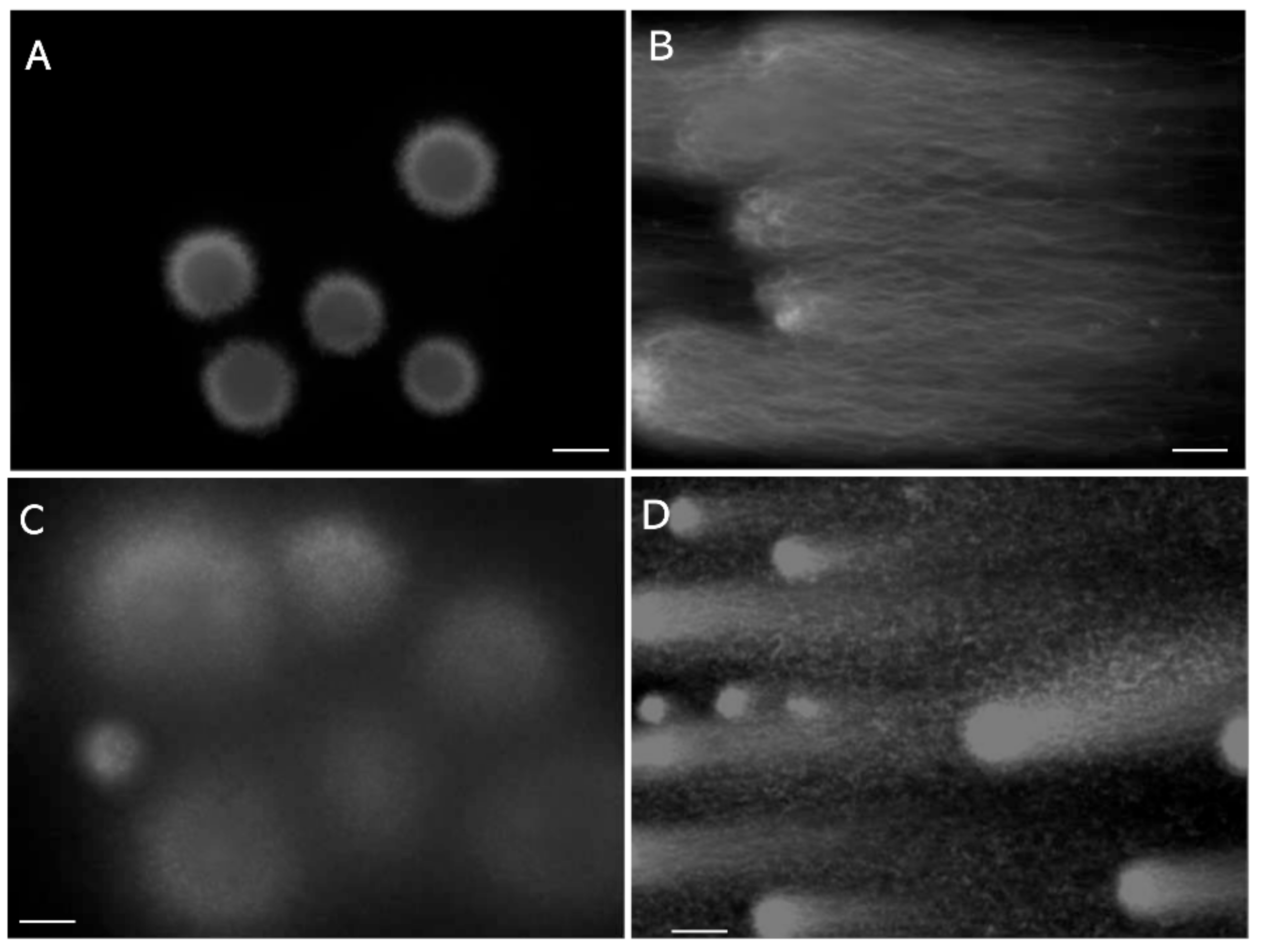
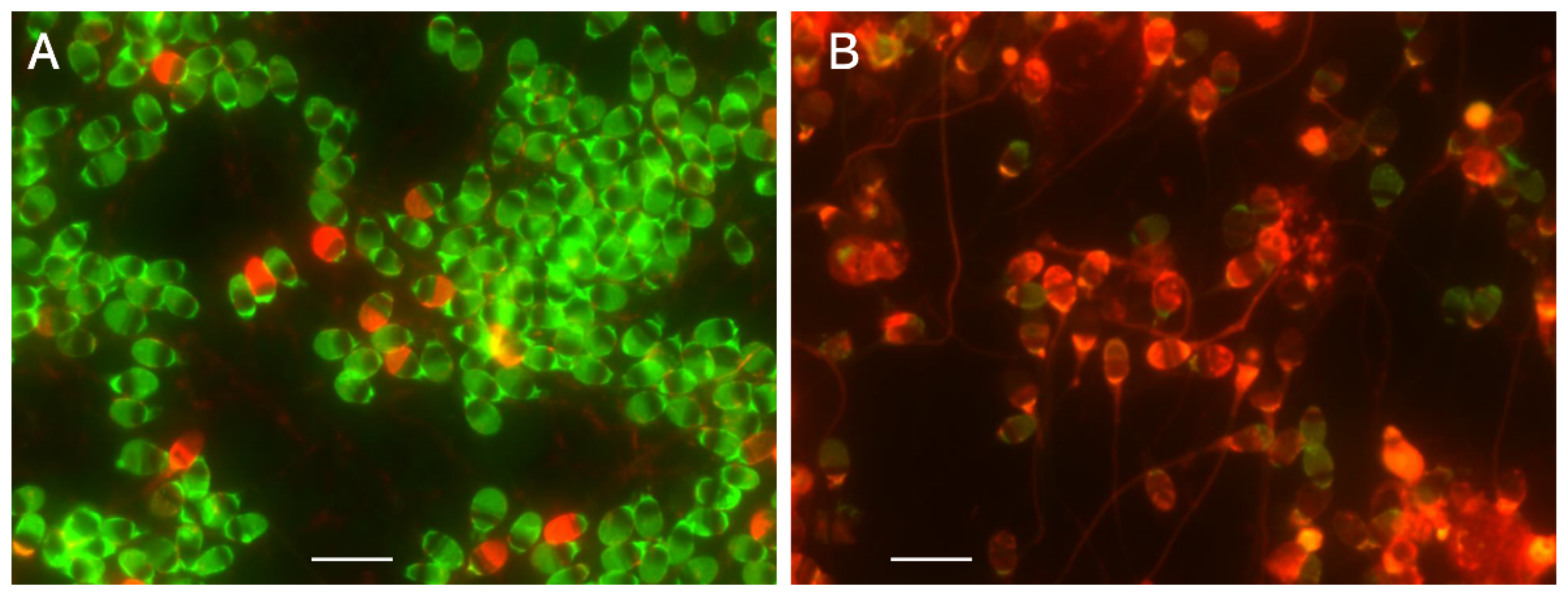
4.2. Red Fluorescence in the SCSA Is Derived from Nucleoproteins but Not from Single-Stranded DNA
4.3. The Halo in the SCD Test Comprises Unextracted Nucleoproteins Adhered to DNA Fibers
4.4. Tightly Packed DNA-Nucleoprotein Complex Physically Blocks Permeation of Certain Dyes and Terminal Deoxynucleotidyl Transferase
5. Angle-Modulated 2D-SCPPFGE
6. How to Develop and Validate Analytical Methods for DNA Fragmentation
7. Separated Motile Sperm Include Various Impairments Besides DNA Fragmentation
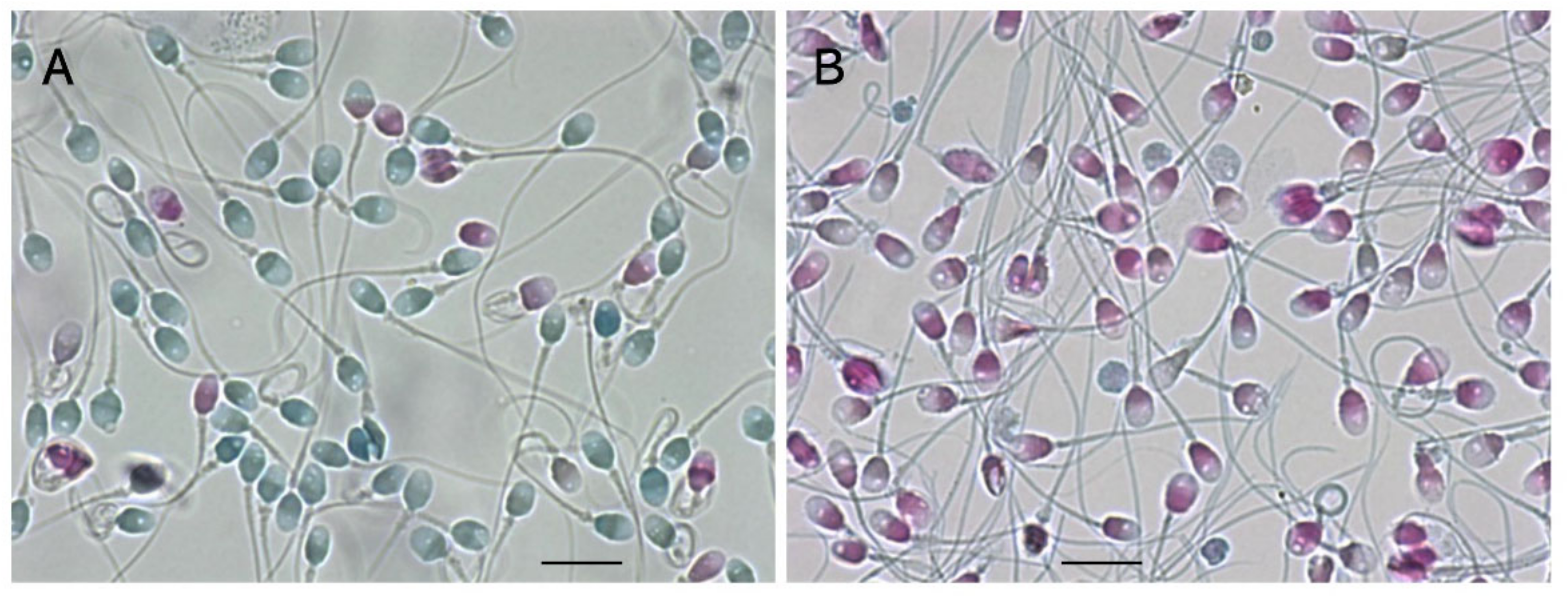
7.1. Sperm-Specific Two-Step Dye-Exclusion Assay to Observe Damage of Plasma Membrane
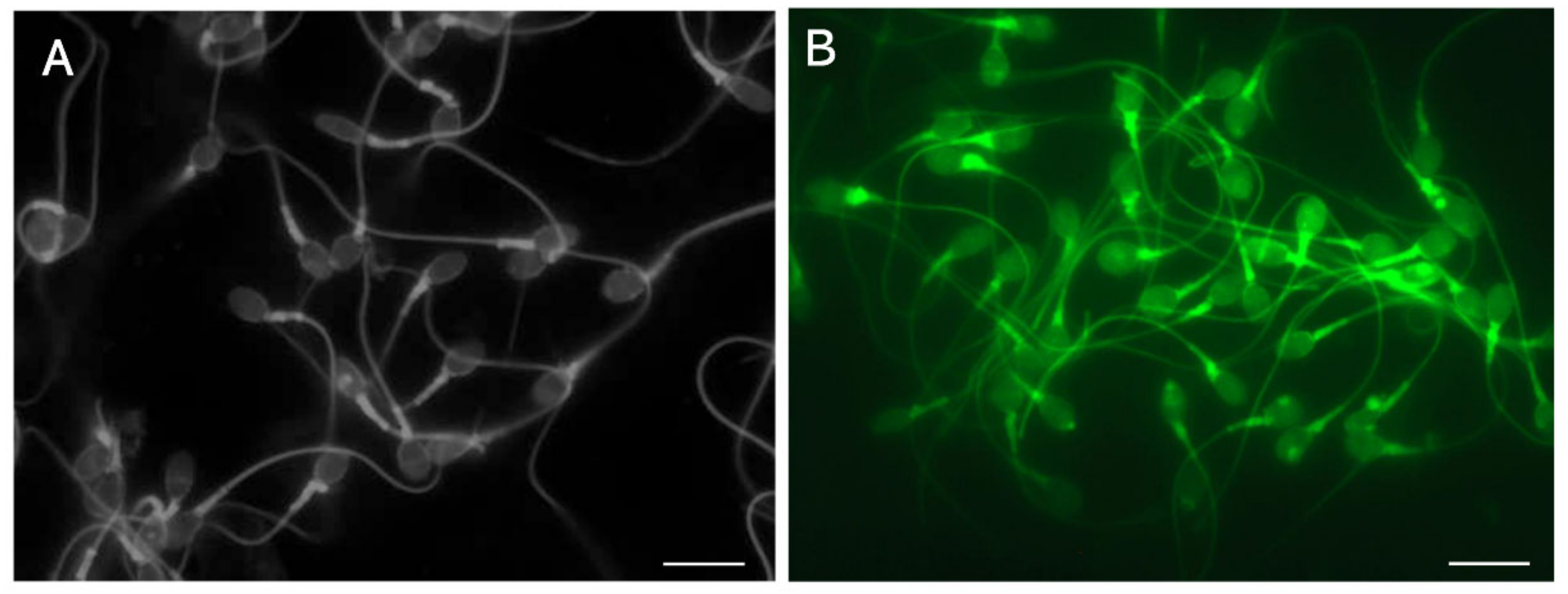
7.2. Observation of the Plasma and Acrosomal Membranes via Two-Step Concanavalin A-Labeling
7.3. Dye-Retention Assay for Endogenous ROS Generated in the Mitochondria
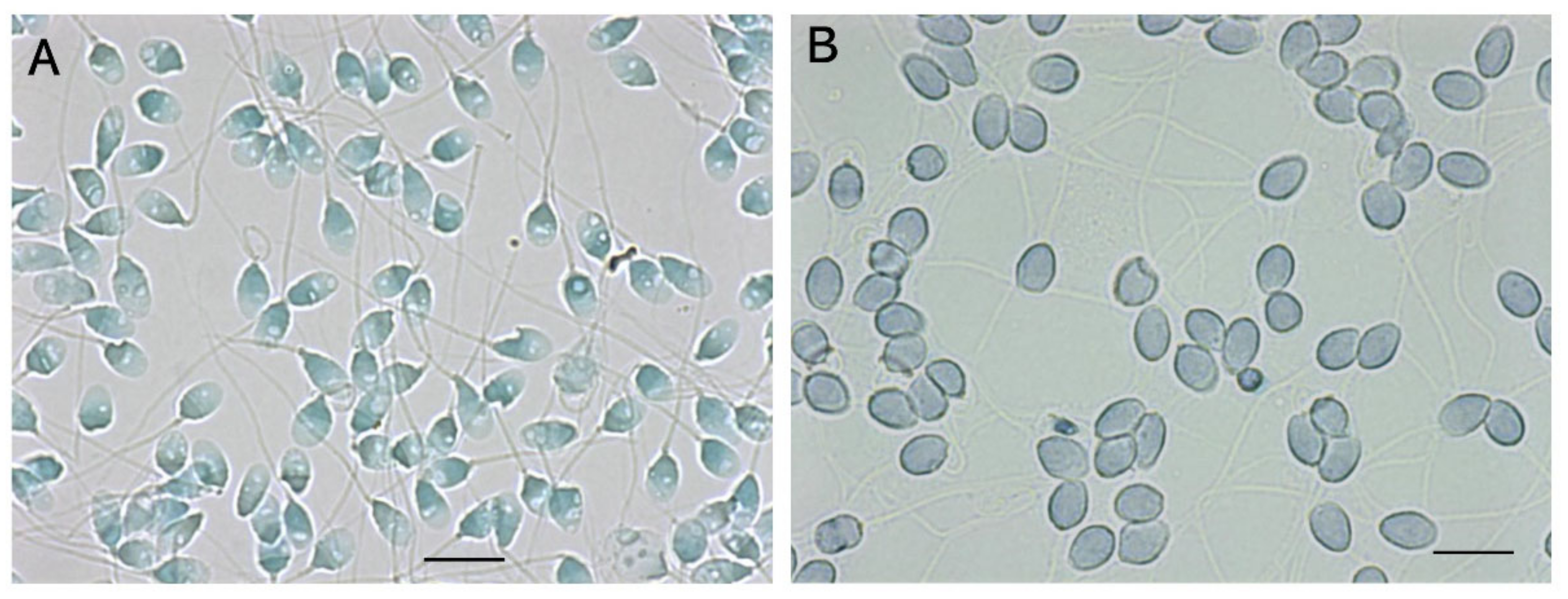
7.4. Observation of Vacuoles in the Heads of Sperm
8. Multivariate Analyses of DNA Damage and Confounding Factors in Cohort Studies
9. Vision for the Future
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Acuna-Hidalgo, R.; Veltman, J.A.; Hoischen, A. New insights into the generation and role of de novo mutations in health and disease. Genome Biol. 2020, 17, 241. [Google Scholar] [CrossRef] [PubMed]
- Middelkamp, S.; Van Tol, H.T.A.; Spierings, D.C.J.; Boymans, S.; Victor Guryev, V. Sperm DNA damage causes genomic instability in early embryonic development. Sci. Adv. 2020, 6, eaaz7602. [Google Scholar] [CrossRef] [PubMed]
- Oud, M.S.; Smits, R.M.; Smith, H.E.; Mastrorosa, F.K.; Holt, G.S. A de novo paradigm for male infertility. Nature 2022, 13, 154. [Google Scholar] [CrossRef] [PubMed]
- Ricci, G.; Perticarari, S.; Fragonas, E.; Giolo, E.; Canova, S. Apoptosis in human sperm: Its correlation with semen quality and the presence of leukocytes. Hum. Reprod. 2002, 17, 2665–2672. [Google Scholar] [CrossRef]
- Nakanishi, Y.; Shiratsuchi, A. Phagocytic Removal of Apoptotic Spermatogenic Cells by Sertoli Cells: Mechanisms and Consequences. Biol. Pharma. Bull. 2004, 27, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, F.; Essers, J.; Kanaar, R.; Wyrobek, A.J. Disruption of maternal DNA repair increases sperm-derived chromosomal aberrations. Proc. Natl. Acad. Sci. USA 2007, 104, 17725–17729. [Google Scholar] [CrossRef] [PubMed]
- Arends, M.J.; Morris, R.G.; Wyllie, A.H. Apoptosis. The role of the endonuclease. Am. J. Pathol. 1990, 136, 593–608. [Google Scholar] [PubMed]
- Borini, A.; Tarozzi, N.; Bizzaro, D.; Bonu, M.A.; Fava, L. Sperm DNA fragmentation: Paternal effect on early post-implantation embryo development in ART. Hum. Reprod. 2006, 21, 2876–2881. [Google Scholar] [CrossRef] [PubMed]
- Wennerholm, U.B.; Bergh, C.; Hamberger, L.; Lundin, K.; Nilsson, L.; Wikland, M.; Källén, B. Incidence of congenital malformations in children born after ICSI. Hum. Reprod. 2000, 15, 944–948. [Google Scholar] [CrossRef]
- Evenson, D.P. The sperm chromatin structure assay (SCSA®) and other sperm DNA fragmentation tests for evaluation of sperm nuclear DNA integrity as related to fertility. Anim. Reprod. Sci. 2016, 169, 56–75. [Google Scholar] [CrossRef]
- Van Brakel, J.; Dinkelman-Smit, M.; de Muinck Keizer-Schrama, S.M.P.F.; Hazebroek, F.W.J.; Dohle, G.R. Sperm DNA damage measured by sperm chromatin structure assay in men with a history of undescended testes. Andrology 2017, 5, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.L.; Muriel, L.; Rivero, M.T.; Goyanes, V.; Vazquez, R. The sperm chromatin dispersion test: A simple method for the determination of sperm DNA fragmentation. J. Androl. 2003, 24, 59–66. [Google Scholar] [CrossRef]
- Fernández, J.L.; Johnston, S.; Gosálvez, J. Sperm chromatin dispersion (SCD) assay. In A Clinician’s Guide to Sperm DNA and Chromatin Damage; Zini, A., Agarwal, A., Eds.; Springer Nature: Barling, AR, USA, 2018; pp. 137–152. [Google Scholar]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environ. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Olive, P.L.; Banáth, J.P. The comet assay: A method to measure DNA damage in individual cells. Nat. Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef]
- Kaneko, S.; Yoshida, J.; Ishikawa, H.; Takamatsu, K. Single-cell pulsed-field gel electrophoresis to detect the early stage of DNA fragmentation in human sperm nuclei. PLoS ONE 2012, 7, e42257. [Google Scholar] [CrossRef]
- Kaneko, S.; Yoshida, J.; Ishikawa, H.; Takamatsu, K. Single-nuclear DNA instability analyses by means of single-cell pulsed-field gel electrophoresis—Technical problems of the comet assay and their solutions for quantitative measurements. J. Mol. Biomark. Diagn. 2013, 5, 5. [Google Scholar] [CrossRef]
- Kaneko, S.; Takamatsu, K. Revalidation of the sperm chromatin dispersion test and the comet assay using intercomparative studies between purified human sperm without and with end-stage DNA fragmentation. J. Med. Diagn. Methods 2023, 12, 406. [Google Scholar]
- Kaneko, S.; Takamatsu, K. Re-Evaluation of Significance of Anti-Sperm Antibodies in Clinical Immune Infertility-Antigenicity of Human Sperm Diminishes during DNA Fragmentation. J. Med. Diagn. Methods 2023, 12, 439. [Google Scholar]
- Kaneko, S.; Okada, Y.; Takamatsu, K. Sperm Specific Two-Step Dye Exclusion Assays to Evaluate Integrity of Plasma and Organelle Membranes-New Approach for Quality Assurance of the Sperm for Intra-Cytoplasmic Sperm Injection. J. Med. Diagn. Methods 2024, 12, 450. [Google Scholar]
- Bortner, C.D.; Cidlowski, J.A. Apoptotic volume decrease and the incredible shrinking cell. Cell Death Differ. 2002, 9, 1307–1310. [Google Scholar] [CrossRef]
- Van Gent, D.C.; Hoeijmakers, J.H.; Kanaar, R. Chromosomal stability and the DNA double-stranded break connection. Nat. Rev. Genet. 2001, 2, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Ceccaldi, R.; Rondinelli, B.; D’Andrea, A.D. Repair pathway choices and consequences at the double-strand break. Trends Cell Biol. 2016, 26, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Kalisky, T.; Quake, S.T. Single-cell genomics. Nat. Methods 2011, 8, 311–314. [Google Scholar]
- Wang, J.; Fan, H.C.; Behr, B.; Quake, S.R. Genome-wide single-cell analysis of recombination activity and de novo mutation rates in human sperm. Cell 2012, 150, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.C.; Cantor, C.R. Separation of yeast chromosome-sized DNAs by pulsed field gradient gel electrophoresis. Cell 1984, 37, 67–75. [Google Scholar] [CrossRef]
- Herschleb, J.; Ananiev, G.; Schwartz, D.C. Pulsed-field gel electrophoresis. Nat. Protoc. 2007, 2, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Restriction endonuclease digestion of DNA in agarose plugs. CSH Protoc. 2006, 2006, pdb.prot4031. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, S.; Takamatsu, K. Angle modulated two-dimensional single cell pulsed-field gel electrophoresis for detecting early symptoms of DNA fragmentation in human sperm nuclei. Sci. Rep. 2024, 14, 840. [Google Scholar] [CrossRef] [PubMed]
- Balhorn, R. The protamine family of sperm nuclear proteins. Genome Biol. 2007, 8, 227. [Google Scholar] [CrossRef]
- Torres-Flores, U.; Hernández-Hernández, A. The interplay between replacement and retention of histones in the sperm genome. Front. Genet. 2020, 11, 780. [Google Scholar] [CrossRef]
- Okada, Y. Sperm chromatin condensation: Epigenetic mechanisms to compact the genome and spatiotemporal regulation from inside and outside the nucleus. Genes Genet. Syst. 2022, 97, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Hagstrom, K.A.; Meyer, B.J. Condensin and cohesin: More than chromosome compactor and glue. Nat. Rev. Genet. 2003, 4, 520–534. [Google Scholar] [CrossRef] [PubMed]
- Nickerson, J. Experimental observations of a nuclear matrix. J. Cell Sci. 2001, 114, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Shaman, J.A.; Yamauchi, Y.; Ward, W.S. Function of the sperm nuclear matrix. Arch. Androl. 2007, 53, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Ada Soler-Ventura, A.; Castillo, J.; de la Iglesia, A.; Jodar, M.; Barrachina, F.; Ballesca, J.L.; Oliva, R. Mammalian sperm protamine extraction and analysis, a step-by-step detailed protocol and brief review of protamine alterations. Protein Pept. Lett. 2018, 25, 424–433. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. Families of serine peptidases. Methods Enzymol. 1994, 244, 19–61. [Google Scholar]
- Jameson, G.W.; Elmore, D.T. Affinity chromatography of bovine trypsin. A rapid separation of bovine α- and β-trypsin. Biochem. J. 1974, 141, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Ebeling, W.; Hennrich, N.; Klockow, M.; Metz, H.; Orth, H.D.; Lang, H. Proteinase K from Tritirachium album Limber. Eur. J. Biochem. 1974, 47, 91–97. [Google Scholar] [CrossRef]
- Kaneko, S.; Takamatsu, K. Re-evaluation of sperm chromatin structure assay (SCSA). J. Med. Diagn. Methods 2023, 12, 399. [Google Scholar]
- Moriyama, Y.; Takano, T.; Ohkuma, S. Acridine orange as a fluorescent probe for lysosomal proton pump. J. Biochem. 1982, 92, 1333–1336. [Google Scholar] [CrossRef]
- Traganos, F.; Darzynkiewicz, Z. Lysosomal proton pump activity: Supravital cell staining with acridine orange differentiates leukocyte subpopulations. Methods Cell Biol. 1994, 41, 185–194. [Google Scholar] [PubMed]
- Han, J.; Burgess, K. Fluorescent indicators for intracellular pH. Chem. Rev. 2010, 110, 2709–2728. [Google Scholar] [CrossRef] [PubMed]
- Darzynkiewicz, Z.; Kapuscinski, J. Acridine orange: A versatile probe of nucleic acids and other cell constituents. In Flow Cytometry and Sorting, 2nd ed.; Wiley: New York, NY, USA, 1990; pp. 291–314. [Google Scholar]
- Uchimoto, T.; Nohara, H.; Kamehara, R.; Iwamura, M.; Watanabe, N. Mechanism of apoptosis induced by a lysosomotropic agent, L-Leucyl-L-Leucine methyl ester. Apoptosis 1999, 4, 357–362. [Google Scholar] [CrossRef]
- Kaneko, S.; Okada, Y.; Yokota, S.; Takamatsu, K. Reactive Blue Dye: Highlights of Vacuoles in Human Sperm. J. Med. Diagn. Methods 2023, 12, 400. [Google Scholar]
- Crowley, L.C.; Brooke, J.; Marfell, B.J.; Waterhouse, N.J. Detection of DNA Fragmentation in Apoptotic Cells by TUNEL. Cold Cold Spring Harb. Protoc. 2016, 2016, pdb.prot087221. [Google Scholar] [CrossRef]
- Chang, L.M.S.; Bollum, F. Deoxynucleotide-polymerizing Enzymes of Calf Thymus Gland. J. Biol. Chem. 1971, 246, 909–916. [Google Scholar] [CrossRef]
- Kaneko, S.; Yoshida, J.; Takamatsu, K. Direct visualization of ascorbic acid inducing double-stranded breaks in single nuclear DNA using single-cell pulsed field gel electrophoresis. Indian J. App. Res. 2015, 5, 248–252. [Google Scholar]
- Anton, K.; Raap, A.K.; Florijn, R.J.; Blonden, L.A.J.; Wiegant, J. Fiber FISH as a DNA Mapping Tool. Methods 1996, 9, 67–73. [Google Scholar]
- Kaneko, S.; Yoshida, J.; Takamatsu, K. Low density regions of DNA in human sperm appear as vacuoles after translucent staining with reactive blue 2. J. Med. Diagn. Methods 2013, 2, 145. [Google Scholar] [CrossRef]
- Patel, K.S. Human Sperm Morphology. In Atlas of Assisted Reproductive Technologies; Ghumman, S., Ed.; Springer Nature: Berlin, Germany, 2023; pp. 179–188. [Google Scholar]
- Yanagimachi, R. Mammalian sperm acrosome reaction: Where does it begin before fertilization? Biol. Reprod. 2011, 85, 4–5. [Google Scholar] [CrossRef]
- Shen, J.; Griffiths, P.T.; Campbell, S.J.; Utinger, B.; Kalberer, M. Ascorbate oxidation by iron, copper and reactive oxygen species: Review, model development, and derivation of key rate constants. Sci. Rep. 2021, 11, 7417. [Google Scholar] [CrossRef] [PubMed]
- Colagar, A.H.; Marzony, E.T. Ascorbic Acid in Human Seminal Plasma: Determination and Its Relationship to Sperm Quality. J. Clin. Biochem. Nutr. 2009, 45, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Clegg, R.M.; Loontiens, F.G.; Van Landschoot, A.; Jovin, T.M. Binding kinetics of methyl alpha-D-mannopyranoside to concanavalin A: Temperature-jump relaxation study with 4-methylumbelliferyl alpha-D-mannopyranoside as a fluorescence indicator ligand. Biochemistry 1981, 20, 4687–4692. [Google Scholar] [CrossRef] [PubMed]
- Celeghini, E.C.; Alves, M.B.; De Arruda, R.P.; de Rezende, G.M.; Florez Rodriguez, S.A. Efficiency of CellROX deep red® and CellROX orange® fluorescent probes in identifying reactive oxygen species in sperm samples from high and low fertility bulls. Anim. Biotechnol. 2021, 32, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Kholmukhamedov, A.; Schwartz, J.M.; Lemasters, J.J. Mitotracker probes and mitochondrial membrane potential. Shock 2013, 39, 543. [Google Scholar] [CrossRef] [PubMed]
- Claes, P.; van Kolen, K.; Roymans, D.; Blero, D.; Vissenberg, K. Reactive blue 2 inhibition of cyclic AMP-dependent differentiation of rat C6 glioma cells by purinergic receptor independent inactivation of phosphatidylinositol 3-kinase. Biochem. Pharmacol. 2004, 67, 1489–1498. [Google Scholar] [CrossRef]
- Atamna, H.; Cheung, I.; Ames, B.N. A method for detecting abasic sites in living cells: Age-dependent changes in base excision repair. Proc. Natl. Acad. Sci. USA 2000, 97, 686–691. [Google Scholar] [CrossRef]
- Talpaert-Borlè, M. Formation, detection and repair of AP sites. Mutat. Res. 1987, 181, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Boiteux, S.; Guillet, M. Abasic sites in DNA: Repair and biological consequences in Saccharomyces cerevisiae. DNA Repair 2004, 3, 1–12. [Google Scholar] [CrossRef]
- Bratton, D.L.; Fadok, V.A.; Richter, D.A.; Kailey, J.M.; Guthrie, L.A. Appearance of Phosphatidylserine on Apoptotic Cells Requires Calcium-mediated Nonspecific Flip-Flop and Is Enhanced by Loss of the Aminophospholipid Translocase. J. Biol. Chem. 1997, 272, 26159–26165. [Google Scholar] [CrossRef]
- García-Peiró, A.; Martínez-Heredia, J.; Oliver-Bonet, M.; Abad, C.; Amengual, M.J.; Navarro, J. Protamine 1 to protamine 2 ratio correlates with dynamic aspects of DNA fragmentation in human sperm. Fertil. Steril. 2011, 95, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Spiess, A.N.; Schuppe, H.C.; Steger, K. The impact of sperm protamine deficiency and sperm DNA damage on human male fertility: A systematic review and meta-analysis. Andrology 2016, 4, 789–799. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaneko, S.; Okada, Y. Revalidation of DNA Fragmentation Analyses for Human Sperm—Measurement Principles, Comparative Standards, Calibration Curve, Required Sensitivity, and Eligibility Criteria for Test Sperm. Biology 2024, 13, 484. https://doi.org/10.3390/biology13070484
Kaneko S, Okada Y. Revalidation of DNA Fragmentation Analyses for Human Sperm—Measurement Principles, Comparative Standards, Calibration Curve, Required Sensitivity, and Eligibility Criteria for Test Sperm. Biology. 2024; 13(7):484. https://doi.org/10.3390/biology13070484
Chicago/Turabian StyleKaneko, Satoru, and Yuki Okada. 2024. "Revalidation of DNA Fragmentation Analyses for Human Sperm—Measurement Principles, Comparative Standards, Calibration Curve, Required Sensitivity, and Eligibility Criteria for Test Sperm" Biology 13, no. 7: 484. https://doi.org/10.3390/biology13070484
APA StyleKaneko, S., & Okada, Y. (2024). Revalidation of DNA Fragmentation Analyses for Human Sperm—Measurement Principles, Comparative Standards, Calibration Curve, Required Sensitivity, and Eligibility Criteria for Test Sperm. Biology, 13(7), 484. https://doi.org/10.3390/biology13070484