
Returning to the Wilderness: Potential Habitat Suitability of Non-Native Pet Birds in South Africa
 , , , ,
, , , ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
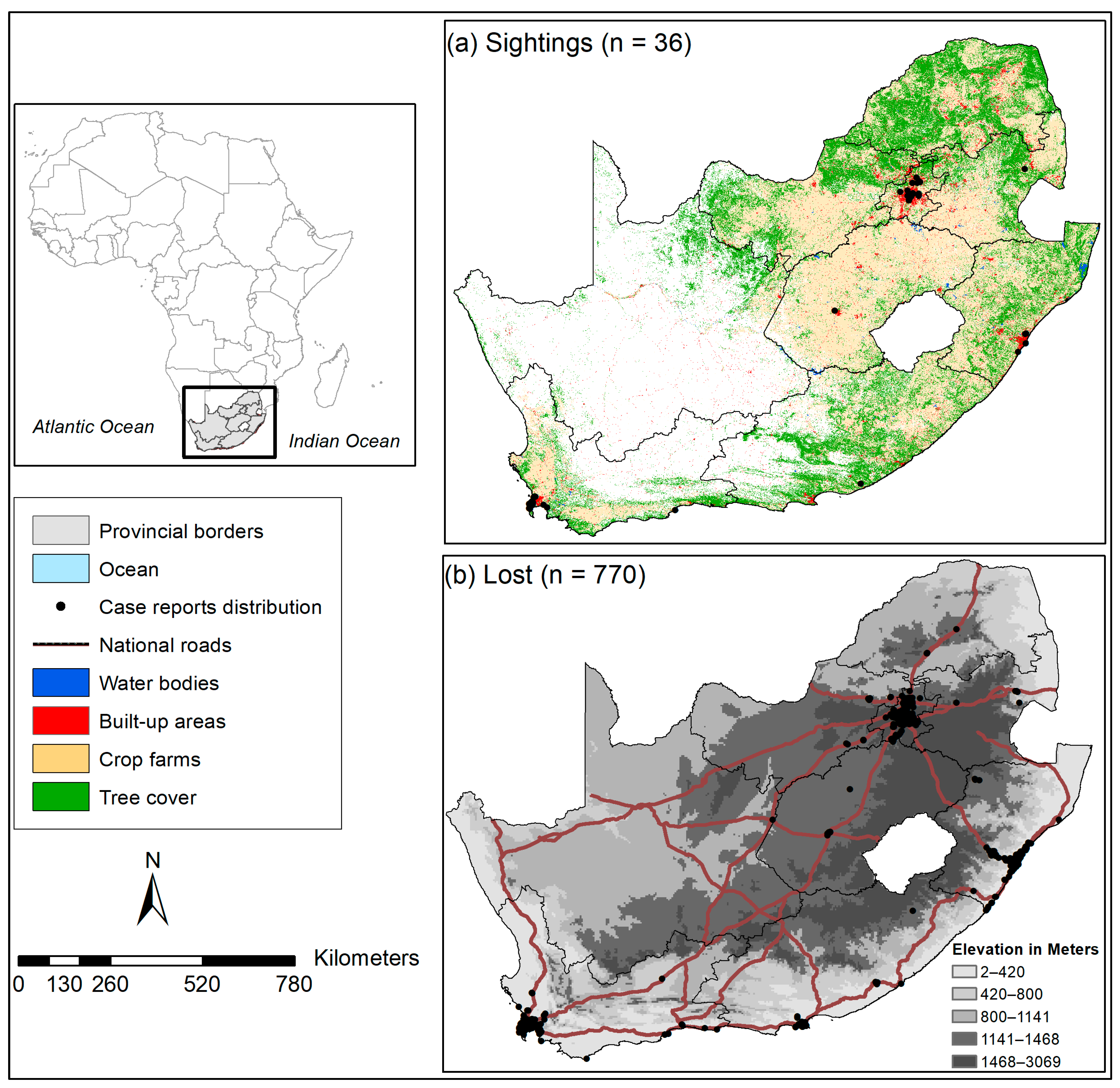
2.1. Geographical Study Areas
2.2. Species Selection Criteria
2.3. Species Occurrence Records Download and Cleaning
2.4. Climatic Data and Environmental Variable Selections

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables Code | Variable Description | Sources |
|---|---|---|
| BIO2 | Mean Diurnal Range (Mean of monthly (max temp − min temp)) | (WorldClim, [82,83]) |
| BIO3 | Isothermality (BIO2/BIO7) (×100) | (WorldClim, [82,83]) |
| BIO5 | Max Temperature of Warmest Month | (WorldClim, [82,83]) |
| BIO8 | Mean Temperature of Wettest Quarter | (WorldClim, [82,83]) |
| Bio9 | Mean Temperature of Driest Quarter | (WorldClim, [82,83]) |
| Bio15 | Precipitation Seasonality (Coefficient of Variation) | (WorldClim, [82,83]) |
| Bio18 | Precipitation of Warmest Quarter | (WorldClim, [82,83]) |
| Bio19 | Precipitation of Coldest Quarter | (WorldClim, [82,83]) |
| WVP | Water Vapor Pressure (kPa) | (WorldClim, [83]) |
| Elv | Elevation | (WorldClim, [83,84]) |
| NDVI | Normalized Difference Vegetation Index | [85] |
| HF3 | Human footprint | [95,96] |
2.5. Model Fitting and Performance
2.6. Establishment Pathway Scenarios
3. Results
3.1. Model Performance
| Species | Lost Case Reports | Variable | Variable of Importance (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AUC | TSS | BIO2 | BIO3 | BIO5 | BIO8 | BIO9 | BIO15 | BIO18 | BIO19 | Humidity | Elv | NDVI | HF3 | ||
| African grey | 314 | 0.968 | 0.826 | 8.30 | 9.94 | 5.67 | 6.22 | 7.85 | 10.52 | 13.81 | 4.63 | 4.73 | 5.34 | 8.3 | 14.69 |
| Budgerigar | 26 | 0.889 | 0.64 | 7.52 | 10.15 | 3.30 | 2.21 | 13.33 | 15.42 | 10.85 | 6.62 | 5.40 | 8.33 | 5.3 | 11.57 |
| Cockatiel | 220 | 0.912 | 0.68 | 5.77 | 9.32 | 3.01 | 2.32 | 14.82 | 8.18 | 10.22 | 9.21 | 7.30 | 7.94 | 4.2 | 17.71 |
| Green-cheeked conure | 41 | 0.977 | 0.891 | 9.01 | 6.41 | 3.73 | 3.41 | 6.87 | 7.93 | 16.71 | 12.62 | 5.04 | 4.15 | 10.33 | 11.79 |
| Monk parakeet | 20 | 0.943 | 0.788 | 9.41 | 7.42 | 3.64 | 2.75 | 8.63 | 10.11 | 8.54 | 11.71 | 13.02 | 4.41 | 8.11 | 12.25 |
| Rose-ringed parakeet | 149 | 0.938 | 0.762 | 5.91 | 7.13 | 3.34 | 1.64 | 12.87 | 11.63 | 9.13 | 14.05 | 7.15 | 3.26 | 9.4 | 14.49 |
3.2. Habitat Suitability and Importance Variables
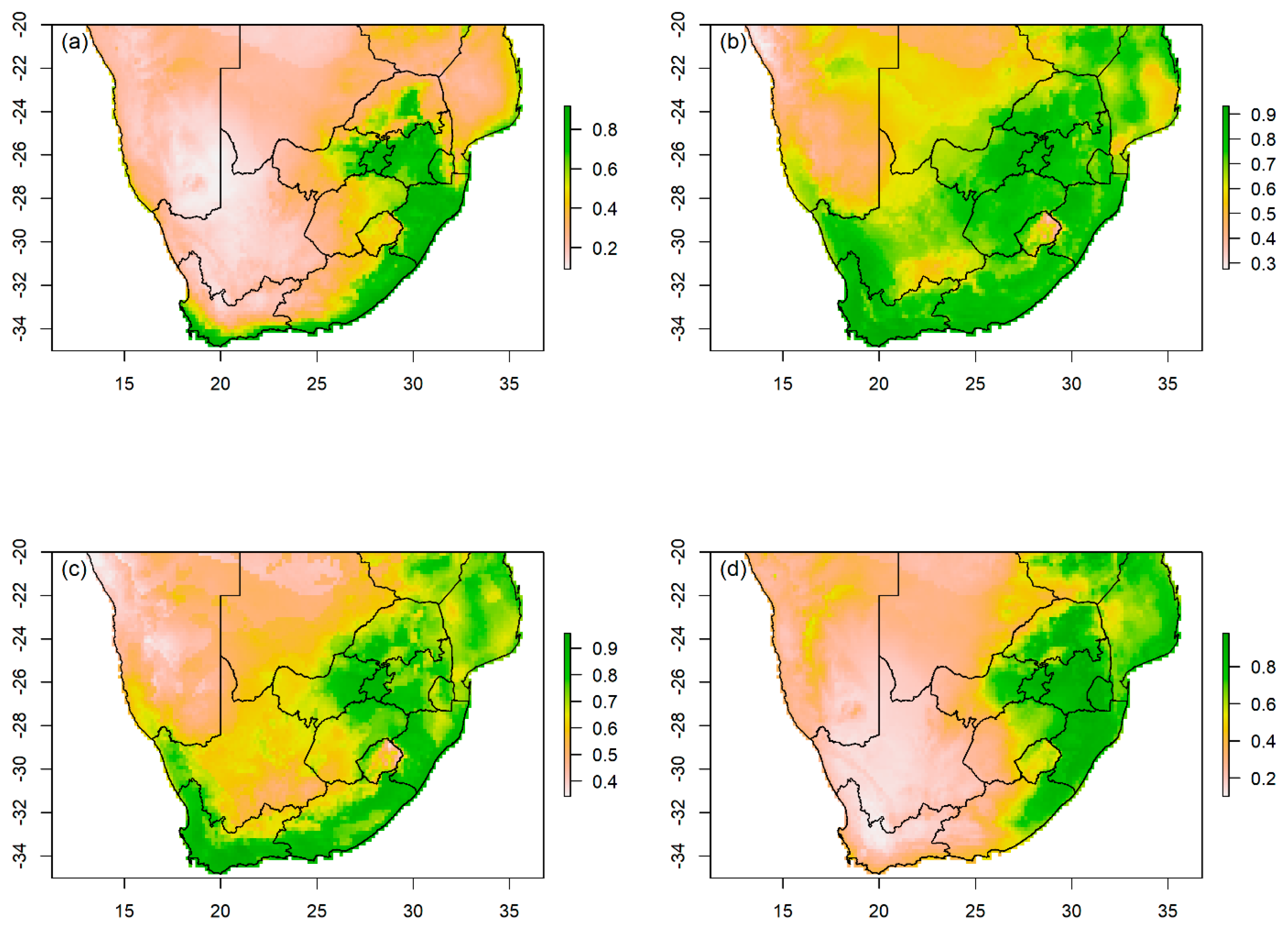

3.3. Pathway Scenario of Lost Pet Bird Leading to Potential Establishment
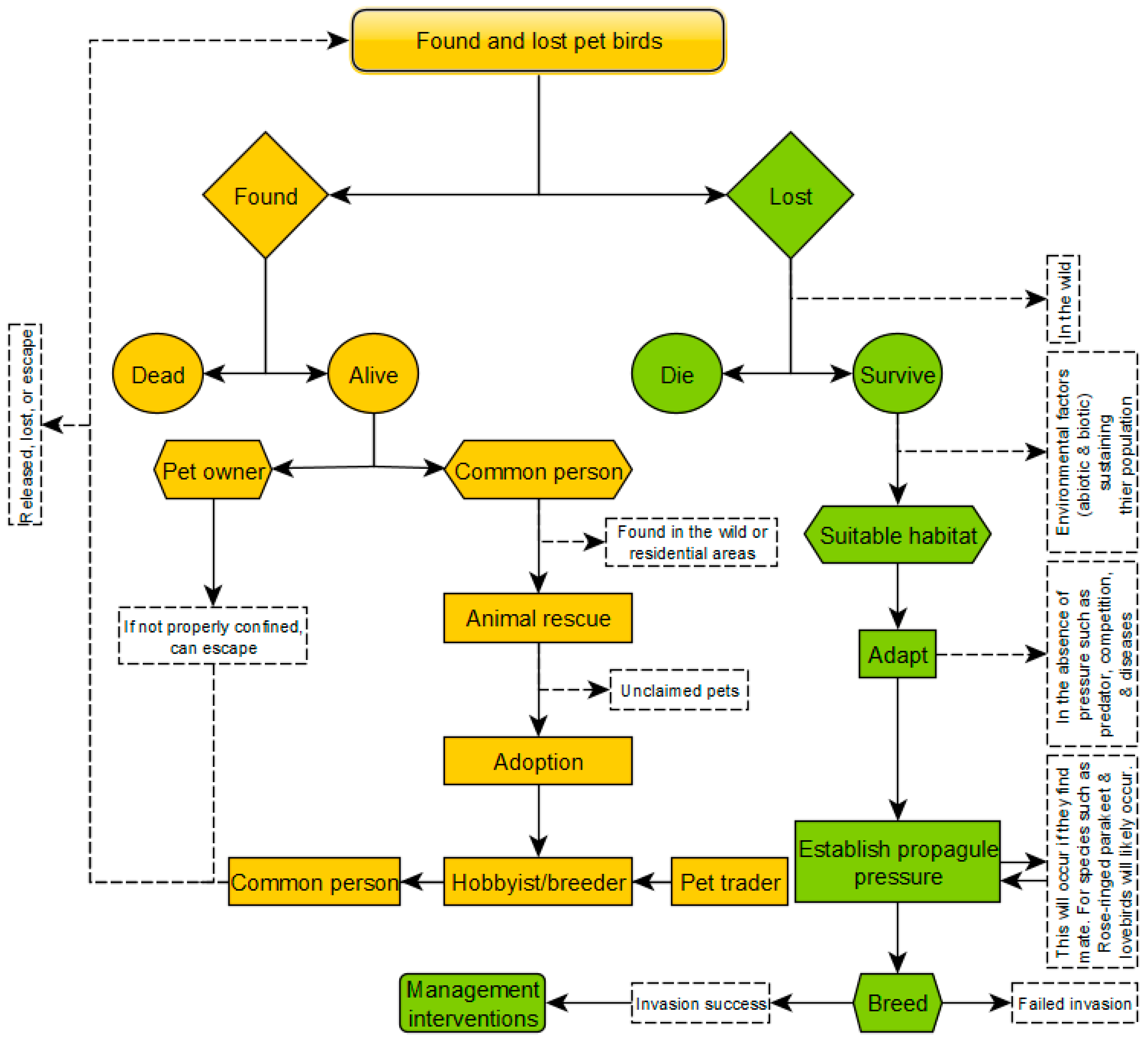
4. Discussion
4.1. Model Performance and Predictive Power
4.2. Environmental Variables Shaping Species Suitability
4.3. Potential Invasion Hotspots and Risk Areas
4.4. Lost Birds Pathways Scenarios to Species Establishment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meyerson, L.A.; Mooney, H.A. Invasive alien species in an era of globalization. Front. Ecol. Environ. 2007, 5, 199–208. [Google Scholar] [CrossRef]
- Ladin, Z.S.; Eggen, D.A.; Trammell, T.L.; D’Amico, V. Human-mediated dispersal drives the spread of the spotted lanternfly (Lycorma delicatula). Sci. Rep. 2023, 13, 1098. [Google Scholar] [CrossRef] [PubMed]
- Searcy, C.A.; Howell, H.J.; David, A.S.; Rumelt, R.B.; Clements, S.L. Patterns of Non-Native Species Introduction, Spread, and Ecological Impact in South Florida, the World’s Most Invaded Continental Ecoregion. Annu. Rev. Ecol. Evol. Syst. 2023, 54, 195–218. [Google Scholar] [CrossRef]
- Williams, S.L.; Grosholz, E.D. The invasive species challenge in estuarine and coastal environments: Marrying management and science. Estuaries Coasts 2008, 31, 3–20. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A global assessment of invasive plant impacts on resident species, communities, and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Chang. Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Chardonnet, P.; Clers, B.D.; Fischer, J.; Gerhold, R.; Jori, F.; Lamarque, F. The value of wildlife. Sci. Tech. J. Int. Off. Epiz. 2002, 21, 15–52. [Google Scholar] [CrossRef] [PubMed]
- Merem, E.C.; Twumasi, Y.; Wesley, J.; Isokpehi, P.; Fageir, S.; Crisler, M.; Romorno, C.; Hines, A.; Ochai, G.S.; Leggett, S.; et al. Assessing the menace of illegal wildlife trade in the sub-Saharan African region. Adv. Life Sci. 2018, 8, 1–25. [Google Scholar] [CrossRef]
- Collard, R.C. Animal Traffic: Lively Capital in the Global Exotic Pet Trade; Duke University Press: Durham, NC, USA, 2020. [Google Scholar]
- Andersson, A.A.; Tilley, H.B.; Lau, W.; Dudgeon, D.; Bonebrake, T.C.; Dingle, C. CITES and beyond: Illuminating 20 years of global, legal wildlife trade. Glob. Ecol. Conserv. 2021, 26, e01455. [Google Scholar] [CrossRef]
- Bush, E.R.; Baker, S.E.; Macdonald, D.W. Global trade in exotic pets 2006–2012. Conserv. Biol. 2014, 28, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Nelufule, T.; Robertson, M.P.; Wilson, J.R.; Faulkner, K.T.; Sole, C.; Kumschick, S. The threats posed by the pet trade in alien terrestrial invertebrates in South Africa. J. Nat. Conserv. 2020, 55, 125831. [Google Scholar] [CrossRef]
- Shivambu, T.C.; Shivambu, N.; Downs, C.T. An assessment of avian species sold in the South African pet trade. Afr. J. Ecol. 2022, 60, 980–995. [Google Scholar] [CrossRef]
- Sinclair, J.S.; Stringham, O.C.; Udell, B.; Mandrak, N.E.; Leung, B.; Romagosa, C.M.; Lockwood, J.L. The international vertebrate pet trade network and insights from US imports of exotic pets. BioScience 2021, 71, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Caldas, A.T.M.; Dias, M.A.; Peres, M. Invertebrate (Araenae: Mygamolomorphae) illegal trade: An ignored side of wildlife trafficking. Am. J. Zool. 2018, 7, 20–23. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Welbourne, D.J.; Romagosa, C.M.; Cassey, P.; Mandrak, N.E.; Strecker, A.; Leung, B.; Stringham, O.C.; Udell, B.; Episcopio-Sturgeon, D.J.; et al. When pets become pests: The role of the exotic pet trade in producing invasive vertebrate animals. Front. Ecol. Environ. 2019, 17, 323–330. [Google Scholar] [CrossRef]
- Mozer, A.; Prost, S. An introduction to illegal wildlife trade and its effects on biodiversity and society. Forensic Sc. Int. Anim. Environ. 2023, 3, 100064. [Google Scholar] [CrossRef]
- Auliya, M.; Altherr, S.; Ariano-Sanchez, D.; Baard, E.H.; Brown, C.; Brown, R.M.; Cantu, J.C.; Gentil, E.G.; Gildenhuys, P.; Henningheim, E.; et al. Trade in live reptiles, its impact on wild populations, and the role of the European market. Biol. Conserv. 2016, 204, 103–119. [Google Scholar] [CrossRef]
- Toland, E.; Bando, M.; Hamers, M.; Cadenas, V.; Laidlaw, R.; Martínez-Silvestre, A.; van der Wielen, P. Turning negatives into positives for pet trading and keeping: A review of positive lists. Animals 2020, 10, 2371. [Google Scholar] [CrossRef] [PubMed]
- Hulme, P.E. Unwelcome exchange: International trade as a direct and indirect driver of biological invasions worldwide. One Earth 2021, 4, 666–679. [Google Scholar] [CrossRef]
- Toomes, A.; Moncayo, S.; Stringham, O.C.; Lassaline, C.; Wood, L.; Millington, M.; Drake, C.; Jense, C.; Allen, A.; Hill, K.G.; et al. A snapshot of online wildlife trade: Australian e-commerce trade of native and non-native pets. Biol. Conserv. 2023, 282, 110040. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, K.T.; Robertson, M.P.; Rouget, M.; Wilson, J.R. Understanding and managing the introduction pathways of alien taxa: South Africa as a case study. Biol. Invasions 2016, 18, 73–87. [Google Scholar] [CrossRef]
- Sagoff, M. What is invasion biology? Ecol. Econ. 2018, 154, 22–30. [Google Scholar] [CrossRef]
- Krishnakumar, K.; Raghavan, R.; Prasad, G.; Bijukumar, A.; Sekharan, M.; Pereira, B.; Ali, A. When pets become pests–exotic aquarium fishes and biological invasions in Kerala, India. Curr. Sci. 2009, 97, 474–476. [Google Scholar]
- Stanley, M.C.; McNaughton, E.J.; Fewster, R.M.; Galbraith, J.A. Cumulative propagule pressure exerted by escaped pet parrots. J. Appl. Ecol. 2023, 60, 384–392. [Google Scholar] [CrossRef]
- Shieh, B.; Lin, Y.; Lee, T.; Chang, C.; Cheng, K. Pet trade as sources of introduced bird species in Taiwan. Taiwania 2006, 51, 81–86. [Google Scholar]
- Vall-Llosera, M.; Cassey, P. Leaky doors: Private captivity as a prominent source of bird introductions in Australia. PLoS ONE 2017, 12, e0172851. [Google Scholar] [CrossRef]
- Strubbe, D.; Matthysen, E. Establishment success of invasive ring-necked and monk parakeets in Europe. J. Biogeogr. 2009, 36, 2264–2278. [Google Scholar] [CrossRef]
- Su, S.; Cassey, P.; Blackburn, T.M. The wildlife pet trade as a driver of introduction and establishment in alien birds in Taiwan. Biol. Invasions 2016, 18, 215–229. [Google Scholar] [CrossRef]
- Postigo, J.L.; Carrillo-Ortiz, J.; Domènech, J.; Tomàs, X.; Arroyo, L.; Senar, J.C. Dietary plasticity in an invasive species and implications for management: The case of the monk parakeet in a Mediterranean city. Anim. Biodivers. Conserv. 2021, 44, 185–194. [Google Scholar] [CrossRef]
- Downs, C.T.; Hart, L.A. Invasive Birds: Global Trends and Impacts; CABI International: Wallingford, UK, 2020; 400p. [Google Scholar] [CrossRef]
- Martin-Albarracin, V.L.; Amico, G.C.; Simberloff, D.; Nuñez, M.A. Impact of non-native birds on native ecosystems: A global analysis. PLoS ONE 2015, 10, e0143070. [Google Scholar] [CrossRef]
- Avery, M.L.; Greiner, E.C.; Lindsay, J.R.; Newman, J.R.; Pruett-Jones, S. Monk parakeet management at electric utility facilities in South Florida. Proc. Vertebr. Pest Conf. 2002, 20, 140–145. [Google Scholar] [CrossRef]
- Hernández-Brito, D.; Carrete, M.; Ibáñez, C.; Juste, J.; Tella, J.L. Nest-site competition and killing by invasive parakeets cause the decline of a threatened bat population. R. Soc. Open Sci. 2018, 5, 172477. [Google Scholar] [CrossRef] [PubMed]
- Battisti, C.; Fraticelli, F. When ‘bullies’ come out of cities: Mobbing among introduced Rose-Ringed Parakeets Psittacula krameri and native species of conservation concern in extra-urban habitats with suggestions for a framework of PSIR indicators. Biodiversity 2023, 24, 96–101. [Google Scholar] [CrossRef]
- Bovet, J. Homing in red squirrels (Tamiasciurus hudsonicus): The importance of going straight. Ethology 1995, 101, 1–9. [Google Scholar] [CrossRef]
- Andreychev, A.V.; Kiyaykina, O.S. Homing of the forest dormouse (Dryomys nitedula, Rodentia, Gliridae). Biol. Bull. Russ. Acad. Sci. 2020, 47, 1227–1234. [Google Scholar] [CrossRef]
- McCartney, K.R.; Kumar, S.; Sing, S.E.; Ward, S.M. Using invaded-range species distribution modeling to estimate the potential distribution of Linaria species and their hybrids in the US northern Rockies. Invas. Plant Sci. Manag. 2019, 12, 97–111. [Google Scholar] [CrossRef]
- Gong, S.; Gao, Y.; Duan, H.; Ge, Y.; Wei, Y. Incorporating physiological data into species distribution models to predict the potential distribution range of the red-eared slider in China. Ecol. Indic. 2023, 154, 110749. [Google Scholar] [CrossRef]
- El-Barougy, R.F.; Dakhil, M.A.; Halmy, M.W.; Gray, S.M.; Abdelaal, M.; Khedr, A.H.A.; Bersier, L.F. Invasion risk assessment using trait-environment and species distribution modelling techniques in an arid protected area: Towards conservation prioritization. Ecol. Indic. 2021, 129, 107951. [Google Scholar] [CrossRef]
- Adhikari, P.; Lee, Y.H.; Poudel, A.; Hong, S.H.; Park, Y.S. Global spatial distribution of Chromolaena odorata habitat under climate change: Random forest modeling of one of the 100 worst invasive alien species. Sci. Rep. 2023, 13, 9745. [Google Scholar] [CrossRef] [PubMed]
- Jarnevich, C.; Engelstad, P.; LaRoe, J.; Hays, B.; Hogan, T.; Jirak, J.; Pearse, I.; Prevéy, J.; Sieracki, J.; Simpson, A.; et al. Invaders at the doorstep: Using species distribution modeling to enhance invasive plant watch lists. Ecol. Inform. 2023, 75, 101997. [Google Scholar] [CrossRef]
- Naimi, B.; Capinha, C.; Ribeiro, J.; Rahbek, C.; Strubbe, D.; Reino, L.; Araújo, M.B. Potential for invasion of traded birds under climate and land-cover change. Glob. Chang. Biol. 2022, 28, 5654–5666. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, J.; Vanmechelen, A.; Holloway, P. Quantifying the distribution and potential biotic interactions between deer and flora using species distribution modelling. Ann. GIS 2023, 29, 553–568. [Google Scholar] [CrossRef]
- Miller, J. Species distribution modeling. Geogr. Comp. 2010, 4, 490–509. [Google Scholar] [CrossRef]
- Naimi, B.; Araújo, M.B. sdm: A reproducible and extensible R platform for species distribution modelling. Ecography 2016, 39, 368–375. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.; Lafond, V.; Griess, V.C. Species distribution models (SDM): Applications, benefits and challenges in invasive species management. CABI Rev. 2019, 1–13. [Google Scholar] [CrossRef]
- Vilizzi, L.; Copp, G.H.; Hill, J.E.; Adamovich, B.; Aislabie, L.; Akin, D.; Al-Faisal, A.J.; Almeida, D.; Azmai, M.A.; Bakiu, R.; et al. A global-scale screening of non-native aquatic organisms to identify potentially invasive species under current and future climate conditions. Sci. Total Environ. 2021, 788, 147868. [Google Scholar] [CrossRef] [PubMed]
- Britton, J.R.; Lynch, A.J.; Bardal, H.; Bradbeer, S.J.; Coetzee, J.A.; Coughlan, N.E.; Dalu, T.; Tricarico, E.; Gallardo, B.; Lintermans, M.; et al. Preventing and controlling nonnative species invasions to bend the curve of global freshwater biodiversity loss. Environ. Rev. 2023, 31, 310–326. [Google Scholar] [CrossRef]
- Le Louarn, M.; Clergeau, P.; Strubbe, D.; Deschamps-Cottin, M. Dynamic species distribution models reveal spatiotemporal habitat shifts in native range-expanding versus non-native invasive birds in an urban area. J. Avian Biol. 2018, 49, jav-01527. [Google Scholar] [CrossRef]
- Shivambu, T.C.; Shivambu, N.; Downs, C.T. Impact assessment of seven alien invasive bird species already introduced to South Africa. Biol. Invasions 2020, 22, 1829–1847. [Google Scholar] [CrossRef]
- Farashi, A.; Alizadeh-Noughani, M. Global distribution modelling, assessment of the spatial invasion risk of Asian pied starling (Sturnus contra Linnaeus, 1758) under climate change. Eur. J. Wildl. Res. 2023, 69, 83. [Google Scholar] [CrossRef]
- Chittenden, H.; Davies, G.; Weiersbye, I. Roberts Bird Guide: Illustrating nearly 1,000 Species in Southern Africa; John Voelcker Bird Book Fund: Cape Town, South Africa, 2016. [Google Scholar]
- Measey, J.; Hui, C.; Somers, M.J. Terrestrial vertebrate invasions in South Africa. In Biological Invasions in South Africa; Van Wilgen, B., Measey, J., Richardson, D., Wilson, J., Zengeya, T., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 115–151. [Google Scholar] [CrossRef]
- Shivambu, T.C.; Shivambu, N.; Downs, C.T. Population estimates of non-native rose-ringed parakeets Psittacula krameri (Scopoli, 1769) in the Durban Metropole, KwaZulu-Natal Province, South Africa. Urban Ecosyst. 2021, 24, 649–659. [Google Scholar] [CrossRef]
- Shivambu, T.C.; Shivambu, N.; Downs, C.T. Breeding status of invasive Rose-ringed Parakeets Psittacula krameri in Durban, South Africa. Ostrich 2022, 93, 257–270. [Google Scholar] [CrossRef]
- Shivambu, T.C.; Shivambu, N.; Nelufule, T.; Moshobane, M.C.; Seoraj-Pillai, N.; Nangammbi, T.C. Evaluating the status of lost, found and sighted non-native pet bird species in South Africa. Diversity 2024, 16, 283. [Google Scholar] [CrossRef]
- Finch, J.M.; Meadows, M.E. South African biomes and their changes over time. In The Geography of South Africa; Knight, J., Rogerson, C., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 57–69. [Google Scholar] [CrossRef]
- IUCN. International Union for Conservation of Nature. 2024. Available online: https://www.iucnredlist.org/ (accessed on 5 December 2023).
- GISD. The Global Invasive Species Database. 2024. Available online: https://www.iucngisd.org/gisd/ (accessed on 12 January 2024).
- Chucholl, C. Invaders for sale: Trade and determinants of introduction of ornamental freshwater crayfish. Biol. Invasions 2013, 15, 125–141. [Google Scholar] [CrossRef]
- Steiner, F.M.; Schlick-Steiner, B.C.; Vanderwal, J.; Reuther, K.D.; Christian, E.; Stauffer, C.; Crozier, R.H. Combined modelling of distribution and niche in invasion biology: A case study of two invasive Tetramorium ant species. Divers. Distrib. 2008, 14, 538–545. [Google Scholar] [CrossRef]
- GBIF.org. GBIF Occurrence of African Grey Psittacus erithacus (Linnaeus, 1758). 2024. [CrossRef]
- GBIF.org. GBIF Occurrence of Budgerigars Melopsittacus undulatus (Shaw, 1805). 2024. [CrossRef]
- GBIF.org. GBIF Occurrence of Cockatiel Nymphicus hollandicus (Kerr, 1792). 2024. [CrossRef]
- GBIF.org. GBIF Occurrence of Green Cheeked Conure Pyrrhura molinae (Massena & Souance, 1854). 2024. [CrossRef]
- GBIF.org. GBIF Occurrence of Monk parakeet Myiopsitta monachus (Boddaert, 1783). 2024. [CrossRef]
- GBIF.org. GBIF Occurrence of Rose-ringed parakeet Psittacula krameria (Scopoli, 1769). 2024. [CrossRef]
- Chamberlain, S.; Boettiger, C.; Ram, K.R.; Barve, V. rgbif: Interface to the Global Biodiversity Information Facility API Methods. R Package Version 0.3.0. 2014. Available online: http://CRAN.R-project.org/package=rgbif (accessed on 12 January 2024).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: http://www.R-project.org/ (accessed on 24 November 2023).
- Li, G.; Xu, G.; Guo, K.; Du, S. Mapping the global potential geographical distribution of black locust (Robinia pseudoacacia L.) using herbarium data and a maximum entropy model. Forests 2014, 5, 2773–2792. [Google Scholar] [CrossRef]
- James, S.A.; Soltis, P.S.; Belbin, L.; Chapman, A.D.; Nelson, G.; Paul, D.L.; Collins, M. Herbarium data: Global biodiversity and societal botanical needs for novel research. Appl. Plant Sci. 2018, 6, e1024. [Google Scholar] [CrossRef] [PubMed]
- Huettmann, F. The forgotten data: A rather short but deep story of museums and libraries in HKH and similar information sources in support of the Global Biodiversity Information System (GBIF.org) and model-predictions for improved conservation management. In Hindu Kush-Himalaya Watersheds Downhill: Landscape Ecology and Conservation Perspectives; Regmi, G., Huettmann, F., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 497–520. [Google Scholar] [CrossRef]
- Robertson, M.P.; Visser, V.; Hui, C. Biogeo: An R package for assessing and improving data quality of occurrence record datasets. Ecography 2016, 39, 394–401. [Google Scholar] [CrossRef]
- Maldonado, C.; Molina, C.I.; Zizka, A.; Persson, C.; Taylor, C.M.; Albán, J.; Chilquillo, E.; Rønsted, N.; Antonelli, A. Estimating species diversity and distribution in the era of Big Data: To what extent can we trust public databases. Glob. Ecol. Biogeogr. 2015, 24, 973–984. [Google Scholar] [CrossRef] [PubMed]
- ESRI. ArcGIS Desktop: Release 10.4.1; Environmental Systems Research Institute: Redlands, CA, USA, 2022. [Google Scholar]
- Hijmans, R. Raster: Geographic Data Analysis and Modeling. R Package Version 3.6-14. 2023. Available online: https://cran.r-project.org/web/packages/raster/raster.pdf (accessed on 11 November 2023).
- Austin, M. Species distribution models and ecological theory: A critical assessment and some possible new approaches. Ecol. Model. 2007, 200, 1–19. [Google Scholar] [CrossRef]
- Rodríguez-Castañeda, G.; Hof, A.R.; Jansson, R.; Harding, L.E. Predicting the fate of biodiversity using species’ distribution models: Enhancing model comparability and repeatability. PLoS ONE 2012, 7, e44402. [Google Scholar] [CrossRef] [PubMed]
- Nishida, S.; Kitamura, W. An influx of non-native bird species into the natural environment owing to the accidental release of pet birds in Japan. Animals 2024, 14, 221. [Google Scholar] [CrossRef] [PubMed]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. BIOCLIM: The first species distribution modelling package, its early applications and relevance to most current MAXENT studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- SRTM. Shuttle Radar Topography Mission 1 Arc-Second Global; USGS: Reston, VA, USA, 2023. [Google Scholar] [CrossRef]
- Copernicus Global Land ServiceProviding Bio-Geophysical Products of Global Land Surface. 2023. Available online: https://land.copernicus.eu/global/products/ndvi (accessed on 29 January 2024).
- Santillán, V.; Quitián, M.; Tinoco, B.A.; Zárate, E.; Schleuning, M.; Böhning-Gaese, K.; Neuschulz, E.L. Direct and indirect effects of elevation, climate and vegetation structure on bird communities on a tropical mountain. Acta Oecol. 2020, 102, 103500. [Google Scholar] [CrossRef]
- Schoell, E.M.; Hille, S.M. Heavy and persistent rainfall leads to brood reduction and nest failure in a passerine bird. J. Avian Biol. 2020, 51, 1–11. [Google Scholar] [CrossRef]
- Veldsman, L.M.; Kylin, H.; Bronkhorst, P.; Engelbrecht, I.; Bouwman, H. A method to determine the combined effects of climate change (temperature and humidity) and eggshell thickness on water loss from bird eggs. Environ. Geochem. Health 2020, 42, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Zwarts, L.; Bijlsma, R.G.; van der Kamp, J.; Sikkema, M. Distribution and numbers of ground-foraging birds between the hyper-arid Sahara and the hyper-humid Guinea forests. Ardea 2023, 111, 7–66. [Google Scholar] [CrossRef]
- Deeming, D.C. Importance of nest type on the regulation of humidity in bird nests. Avian Biol. Res. 2011, 4, 23–31. [Google Scholar] [CrossRef]
- Salaberria, C.; Celis, P.; López-Rull, I.; Gil, D. Effects of temperature and nest heat exposure on nestling growth, dehydration and survival in a Mediterranean hole-nesting passerine. Ibis 2014, 156, 265–275. [Google Scholar] [CrossRef]
- Deeming, D.C.; Gilchrist, R.; Szafraniec, M.; Pollins, J.M. Water vapour conductance of passerine nest walls. Acta Ornithol. 2020, 55, 13–22. [Google Scholar] [CrossRef]
- Marquardt, D.W. Generalized inverses, ridge regression, biased linear estimation, and nonlinear estimation. Technometrics 1970, 12, 591–612. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Sanderson, E.W.; Jaiteh, M.; Levy, M.A.; Redford, K.H.; Wannebo, A.V.; Woolmer, G. The human footprint and the last of the wild: The human footprint is a global map of human influence on the land surface, which suggests that human beings are stewards of nature, whether we like it or not. BioScience 2002, 52, 891–904. [Google Scholar] [CrossRef]
- Sanderson, E.W.; Fisher, K.; Robinson, N.; Sampson, D.; Duncan, A.; Royte, L. The march of the human footprint. EcoEvoRxiv 2022, 1–54. [Google Scholar] [CrossRef]
- Banha, F.; Gama, M.; Anastácio, P.M. The effect of reproductive occurrences and human descriptors on invasive pet distribution modelling: Trachemys scripta elegans in the Iberian Peninsula. Ecol. Model. 2017, 360, 45–52. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Phillips, S.J. Transferability, sample selection bias and background data in presence-only modelling: A response to Peterson et al. (2007). Ecography 2008, 31, 272–278. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- McCullagh, P. Generalized Linear Models; CRC: New York, NY, USA, 2019. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Meyer, D.; Dimitriadou, E.; Hornik, K.; Weingessel, A.; Leisch, F. e1071: Misc Functions of the Department of Statistics, Probability Theory Group (Formerly: E1071), TU Wien. R Package Version 1.6-7. 2015. Available online: https://CRAN.R-package.org?package=e1071 (accessed on 30 January 2024).
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Araújo, M.B.; Anderson, R.P.; Márcia Barbosa, A.; Beale, C.M.; Dormann, C.F.; Early, R.; Garcia, R.A.; Guisan, A.; Maiorano, L.; Naimi, B.; et al. Standards for distribution models in biodiversity assessments. Sci. Adv. 2019, 5, eaat4858. [Google Scholar] [CrossRef] [PubMed]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 2009, 15, 59–69. [Google Scholar] [CrossRef]
- Manel, S.; Williams, H.C.; Ormerod, S.J. Evaluating presence–absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Freeman, E.A.; Moisen, G.G. A comparison of the performance of threshold criteria for binary classification in terms of predicted prevalence and kappa. Ecol. Model. 2008, 217, 48–58. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Lobo, J.M. Threshold criteria for conversion of probability of species presence to either–or presence–absence. Acta Oecol. 2007, 31, 361–369. [Google Scholar] [CrossRef]
- Ed Graph Editor. Works GmbH Software Developer; Version 3.23.2; Works GmbH: Tübingen, Germany, 2023; Available online: https://www.yworks.com/products/yed/download (accessed on 11 November 2023).
- Benemann, V.R.; Araujo, L.D.; Fabbris, A.Z.; Montone, R.C.; Petry, M.V. Nesting distribution of Masked Booby Sula dactylatra at Trindade Island, western South Atlantic Ocean. Mar. Ornithol. 2022, 50, 189–195. [Google Scholar]
- Liu, X.; Liu, H.; Datta, P.; Frey, J.; Koch, B. Mapping an invasive plant Spartina alterniflora by combining an ensemble one-class classification algorithm with a phenological NDVI time-series analysis approach in middle coast of Jiangsu, China. Remote Sens. 2020, 12, 4010. [Google Scholar] [CrossRef]
- Petrosyan, V.; Dinets, V.; Osipov, F.; Dergunova, N.; Khlyap, L. Range dynamics of Striped field mouse (Apodemus agrarius) in Northern Eurasia under Global Climate Change Based on Ensemble Species Distribution Models. Biology 2023, 12, 1034. [Google Scholar] [CrossRef] [PubMed]
- Falcón, W.; Tremblay, R.L. From the cage to the wild: Introductions of Psittaciformes to Puerto Rico. PeerJ 2018, 6, e5669. [Google Scholar] [CrossRef] [PubMed]
- Senar, J.C.; Domènech, J.; Arroyo, L.; Torre, I.; Gordo, O. An evaluation of monk parakeet damage to crops in the metropolitan area of Barcelona. Anim. Biodivers. Conserv. 2016, 39, 141–145. [Google Scholar] [CrossRef]
- Senar, J.C.; Moyà, A.; Pujol, J.; Tomas, X.; Hatchwell, B.J. Sex and age effects on monk parakeet home-range variation in the urban habitat. Diversity 2021, 13, 648. [Google Scholar] [CrossRef]
- Borray-Escalante, N.A.; Baucells, J.; Carrillo-Ortiz, J.; Hatchwell, B.J.; Senar, J.C. Long-distance dispersal of monk parakeets. Anim. Biodivers. Conserv. 2023, 46, 71–78. [Google Scholar] [CrossRef]
- Strubbe, D.; Matthysen, E. A radiotelemetry study of habitat use by the exotic Ring-necked Parakeet Psittacula krameri in Belgium. Ibis 2011, 153, 180–184. [Google Scholar] [CrossRef]
- Reed, J.E.; McCleery, R.A.; Silvy, N.J.; Smeins, F.E.; Brightsmith, D.J. Monk parakeet nest-site selection of electric utility structures in Texas. Landsc. Urban Plan. 2014, 129, 65–72. [Google Scholar] [CrossRef]
- Shivambu, N.; Shivambu, T.C.; Downs, C.T. Predicting the potential distribution of non-native mammalian species sold in the South African pet trade. Diversity 2021, 13, 478. [Google Scholar] [CrossRef]
- Goetz, S.J.; Sun, M.; Zolkos, S.; Hansen, A.; Dubayah, R. The relative importance of climate and vegetation properties on patterns of North American breeding bird species richness. Environ. Res. Lett. 2014, 9, 034013. [Google Scholar] [CrossRef]
- Illán, J.G.; Thomas, C.D.; Jones, J.A.; Wong, W.K.; Shirley, S.M.; Betts, M.G. Precipitation and winter temperature predict long-term range-scale abundance changes in Western North American birds. Glob. Chang. Biol. 2014, 20, 3351–3364. [Google Scholar] [CrossRef] [PubMed]
- Zuckerberg, B.; Ribic, C.A.; McCauley, L.A. Effects of temperature and precipitation on grassland bird nesting success as mediated by patch size. Conserv. Biol. 2018, 32, 872–882. [Google Scholar] [CrossRef]
- Ragusa-Netto, J. Feeding ecology of the Green-cheeked parakeet (Pyrrhura molinae) in dry forests in western Brazil. Braz. J. Biol. 2007, 67, 243–249. [Google Scholar] [CrossRef]
- Butler, C.J.; Cresswell, W.; Gosler, A.; Perrins, C. The breeding biology of Rose-ringed Parakeets Psittacula krameri in England during a period of rapid population expansion. Bird Study 2013, 60, 527–532. [Google Scholar] [CrossRef]
- Postigo, J.L.; Shwartz, A.; Strubbe, D.; Muñoz, A.R. Unrelenting spread of the alien monk parakeet Myiopsitta monachus in Israel. Is it time to sound the alarm? Pest Manag. Sci. 2017, 73, 349–353. [Google Scholar] [CrossRef]
- Chamberlain, D.; Brambilla, M.; Caprio, E.; Pedrini, P.; Rolando, A. Alpine bird distributions along elevation gradients: The consistency of climate and habitat effects across geographic regions. Oecologia 2016, 181, 1139–1150. [Google Scholar] [CrossRef]
- Dyer, E.E.; Cassey, P.; Redding, D.W.; Collen, B.; Franks, V.; Gaston, K.J.; Jones, K.E.; Kark, S.; Orme, C.D.L.; Blackburn, T.M. The global distribution and drivers of alien bird species richness. PLoS Biol. 2017, 15, e2000942. [Google Scholar] [CrossRef] [PubMed]
- Banko, P.C.; Oboyski, P.T.; Slotterback, J.W.; Dougill, S.J.; Goltz, D.M.; Johnson, L.; Laut, M.E.; Murray, T.C. Availability of food resources, distribution of invasive species, and conservation of a Hawaiian bird along a gradient of elevation. J. Biogeogr. 2002, 29, 789–808. [Google Scholar] [CrossRef]
- Sauve, D.; Friesen, V.L.; Charmantier, A. The effects of weather on avian growth and implications for adaptation to climate change. Front. Ecol. Evol. 2021, 9, 5. [Google Scholar] [CrossRef]
- Ritchison, G. Avian reproduction: Clutch sizes, incubation, and hatching. In In a Class of Their Own: A Detailed Examination of Avian Forms and Functions; Ritchison, G., Ed.; Springer International Publishing: Cham, Switzerland, 2023; pp. 2285–2382. [Google Scholar] [CrossRef]
- Department of Environmental Affairs (DEA). Environmental Management: Biodiversity Act 2004 (Act No. 10 of 2004) Alien and Invasive Species Regulations; National Government Gazette: Pretoria, South Africa, 2020; pp. 31–77. [Google Scholar]
- Hernández-Brito, D.; Carrete, M.; Blanco, G.; Romero-Vidal, P.; Senar, J.C.; Mori, E.; White, T.H., Jr.; Luna, Á.; Tella, J.L. The Role of Monk Parakeets as Nest-Site Facilitators in Their Native and Invaded Areas. Biology 2021, 10, 683. [Google Scholar] [CrossRef] [PubMed]
- Elhariri, M.; Hamza, D.; Elhelw, R.; Refai, M. Lovebirds and cockatiels risk reservoir of Cryptococcus neoformans, a potential hazard to human health. J. Vet. Sci. Med. Diagn. 2015, 4, 2. [Google Scholar] [CrossRef]
- Pranty, B. The Budgerigar in Florida: Rise and fall of an exotic psittacid. N. Am. Birds 2001, 55, 389–397. [Google Scholar]
- Pranty, B.; Epps, S. Distribution, population status, and documentation of exotic parrots in Broward County, Florida. Flo. Field Nat. 2002, 30, 111–131. [Google Scholar]
- Toft, C.A.; Wright, T.F. Parrots of the Wild: A Natural History of the World’s Most Captivating Birds; University of California Press: Oakland, CA, USA, 2015; p. 248. [Google Scholar]
- van Wilgen, N.J.; Wilson, J.R.U.; Elith, J.; Wintle, B.A.; Richardson, D.M. Alien invaders and reptile traders: What drives the live animal trade in South Africa? Anim. Conserv. 2010, 13, 24–32. [Google Scholar] [CrossRef]
- Stringham, O.C.; Lockwood, J.L. Pet problems: Biological and economic factors that influence the release of alien reptiles and amphibians by pet owners. J. Appl. Ecol. 2018, 55, 2632–2640. [Google Scholar] [CrossRef]
- Rouget, M.; Robertson, M.P.; Wilson, J.R.U.; Hui, C.; Essl, F.; Rentería, J.L.; Richardson, D.M. Invasion debt—Quantifying future biological invasions. Divers. Distrib. 2016, 22, 445–456. [Google Scholar] [CrossRef]
- Wilson, J.R.; Ivey, P.; Manyama, P.; Nanni, I. A new national unit for invasive species detection, assessment and eradication planning. S. Afr. J. Sci. 2013, 109, 13. [Google Scholar] [CrossRef]
- Faulkner, K.T.; Robertson, M.P.; Rouget, M.; Wilson, J.R. A simple, rapid methodology for developing invasive species watch lists. Biol. Conserv. 2014, 179, 25–32. [Google Scholar] [CrossRef]
- van Wilgen, B.W.; Wannenburgh, A.; Wilson, J.R. A review of two decades of government support for managing alien plant invasions in South Africa. Biol. Conserv. 2022, 274, 109741. [Google Scholar] [CrossRef]
- Moshobane, M.C.; Mukundamago, M.; Adu-Acheampong, S.; Shackleton, R. Development of alien and invasive taxa lists for regulation of biological invasions in South Africa. Bothalia 2019, 49, 11. [Google Scholar] [CrossRef]
- Wilson, J.R.; Faulkner, K.T.; Rahlao, S.J.; Richardson, D.M.; Zengeya, T.A.; Van Wilgen, B.W. Indicators for monitoring biological invasions at a national level. J. Appl. Ecol. 2018, 55, 2612–2620. [Google Scholar] [CrossRef]

| Method Full Names | Code | Dependent Packages | References |
|---|---|---|---|
| Generalized Linear Model | GLM | STATS | [71,101] |
| Random Forest | RF | randomForest | [102,103] |
| Support Vector Machine | SVM | e1071 | [104] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shivambu, T.C.; Shivambu, N.; Nelufule, T.; Moshobane, M.C.; Seoraj-Pillai, N.; Nangammbi, T.C. Returning to the Wilderness: Potential Habitat Suitability of Non-Native Pet Birds in South Africa. Biology 2024, 13, 483. https://doi.org/10.3390/biology13070483
Shivambu TC, Shivambu N, Nelufule T, Moshobane MC, Seoraj-Pillai N, Nangammbi TC. Returning to the Wilderness: Potential Habitat Suitability of Non-Native Pet Birds in South Africa. Biology. 2024; 13(7):483. https://doi.org/10.3390/biology13070483
Chicago/Turabian StyleShivambu, Tinyiko C., Ndivhuwo Shivambu, Takalani Nelufule, Moleseng C. Moshobane, Nimmi Seoraj-Pillai, and Tshifhiwa C. Nangammbi. 2024. "Returning to the Wilderness: Potential Habitat Suitability of Non-Native Pet Birds in South Africa" Biology 13, no. 7: 483. https://doi.org/10.3390/biology13070483
APA StyleShivambu, T. C., Shivambu, N., Nelufule, T., Moshobane, M. C., Seoraj-Pillai, N., & Nangammbi, T. C. (2024). Returning to the Wilderness: Potential Habitat Suitability of Non-Native Pet Birds in South Africa. Biology, 13(7), 483. https://doi.org/10.3390/biology13070483