
Analysis of the Distribution Pattern of Phenacoccus manihoti in China under Climate Change Based on the Biomod2 Model

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
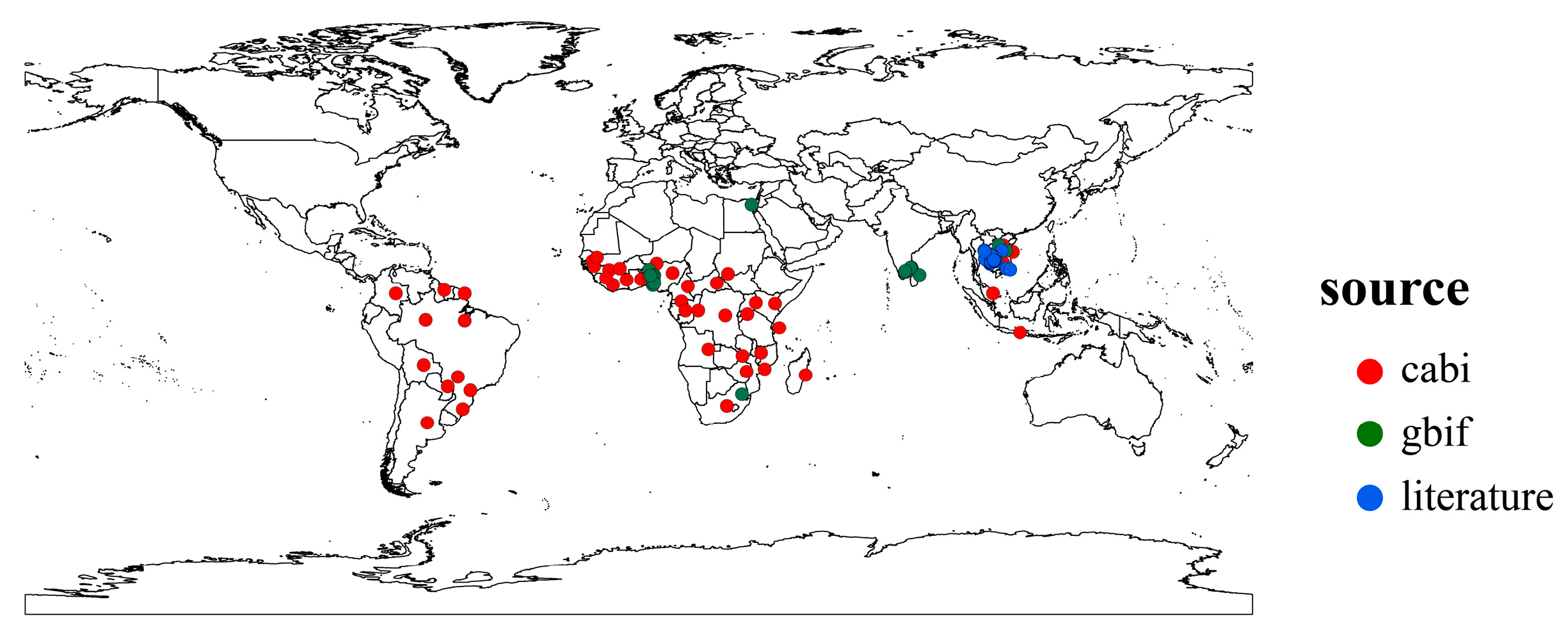
2.1. Distribution Records of Phenacoccus manihoti
2.2. Environmental Variables
2.3. Model Settings and Assessment
2.4. Analysis and Mapping
2.5. Ecological Niche Analysis
3. Results
3.1. Model Validation
3.2. Environmental Variables
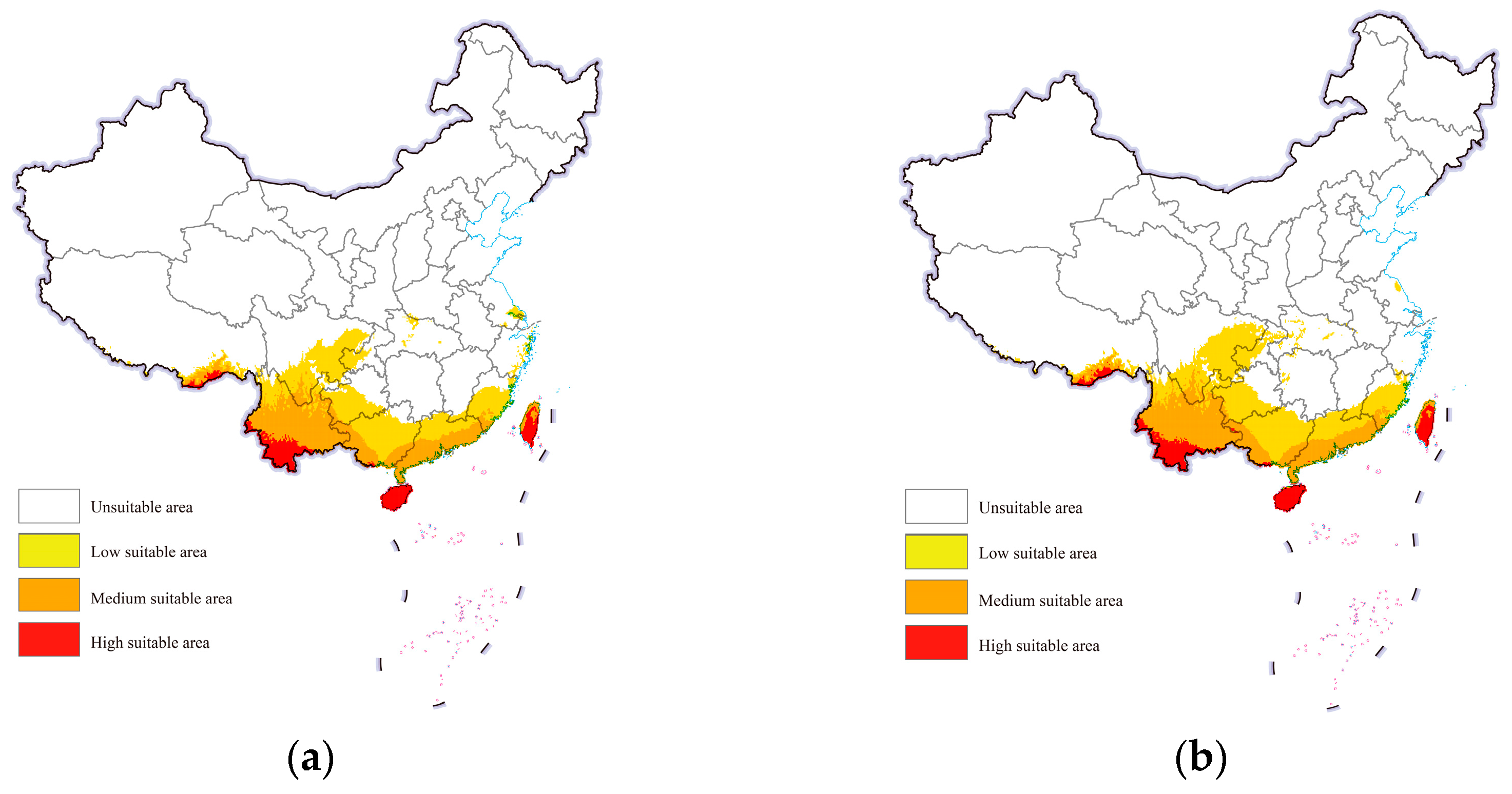
3.3. Potential Geographical Distribution in China
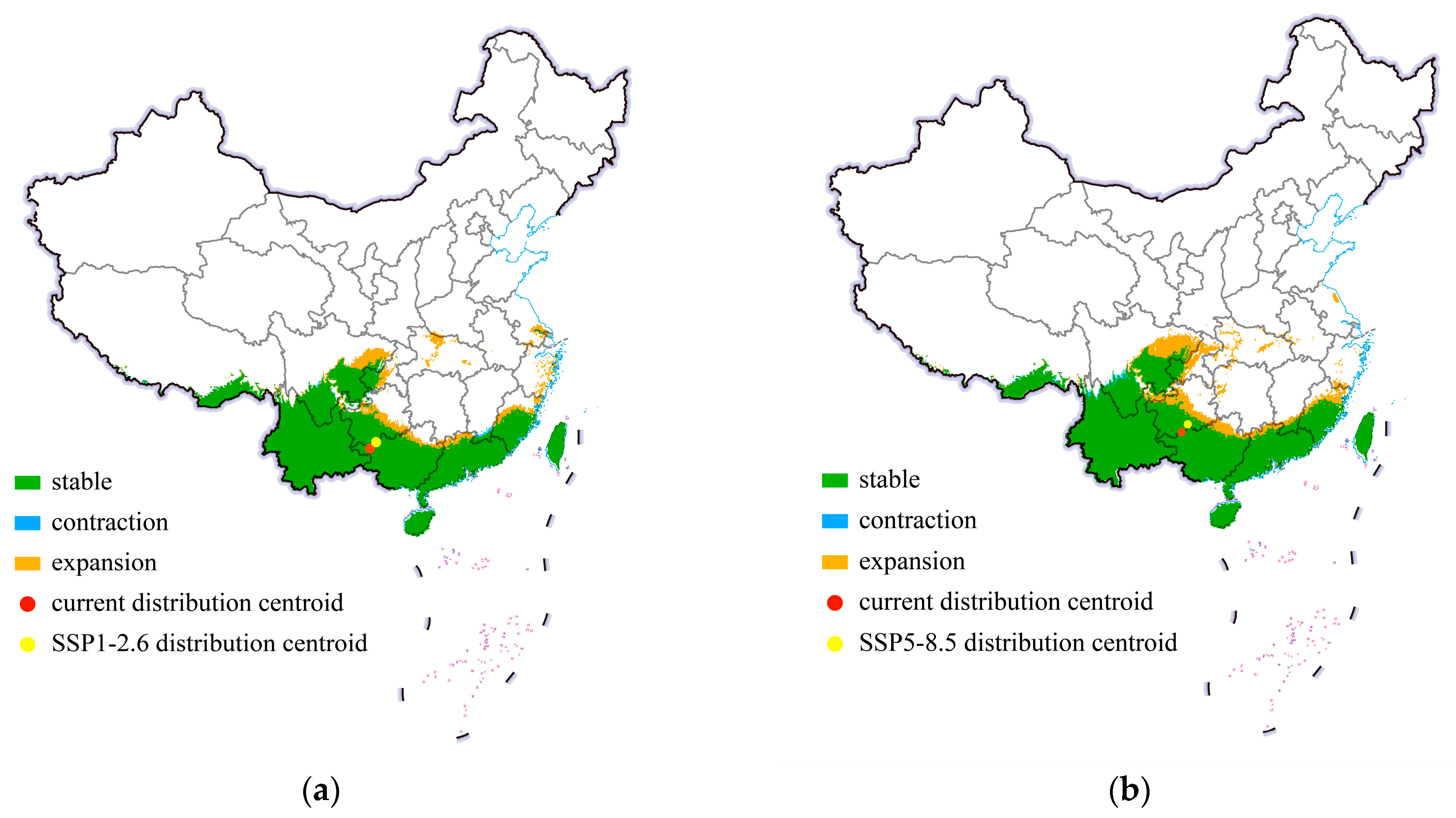
3.4. Shifts in the Distribution Centroid and Distribution Pattern
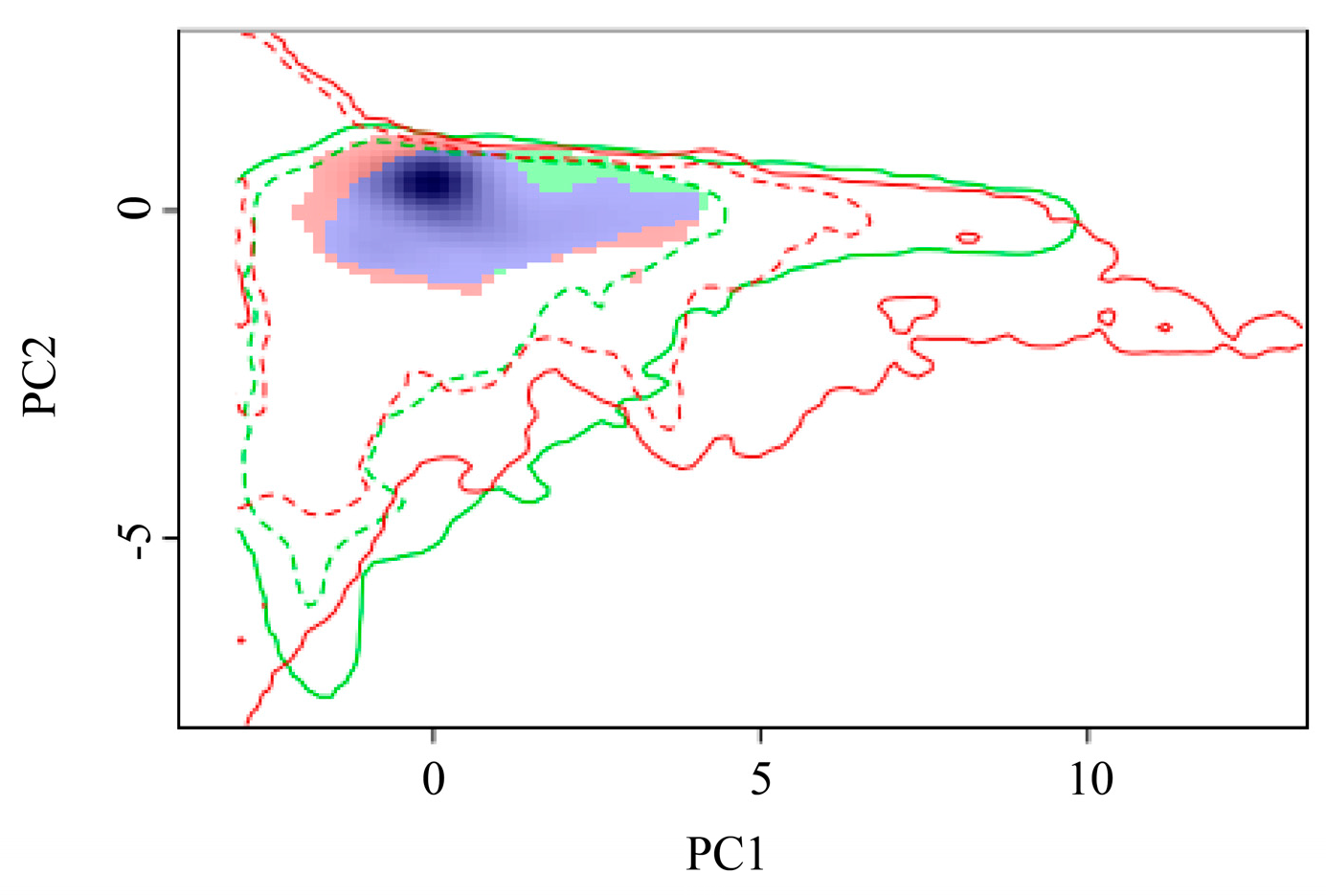
3.5. Ecological Niche of Phenacoccus manihoti
4. Discussion
4.1. Optimization and Selection of the Biomod2 Model
4.2. Significant Environmental Variables
4.3. Distribution Pattern and Ecological Niches
4.4. Management and Monitoring
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of Variables | Environmental Variables | Code |
|---|---|---|
| Climatic factors | Annual mean temperature | bio1 |
| Mean diurnal range | bio2 | |
| Isothermality | bio3 | |
| Temperature seasonality | bio4 | |
| Max temperature of warmest month | bio5 | |
| Min temperature of coldest month | bio6 | |
| Temperature annual range | bio7 | |
| Mean temperature of wettest quarter | bio8 | |
| Mean temperature of driest quarter | bio9 | |
| Mean temperature of warmest quarter | bio10 | |
| Mean temperature of coldest quarter | bio11 | |
| Annual precipitation | bio12 | |
| Precipitation of wettest month | bio13 | |
| Precipitation of driest month | bio14 | |
| Precipitation seasonality | bio15 | |
| Precipitation of wettest quarter | bio16 | |
| Precipitation of driest quarter | bio17 | |
| Precipitation of warmest quarter | bio18 | |
| Precipitation of coldest quarter | bio19 | |
| Terrain factors | Elevation | elev |
References
- Clavijo, M.A.; Effah, E.; Najar-Rodriguez, A. Ecological aspects of volatile organic compounds emitted by exotic invasive plants. Front. Ecol. Evol. 2023, 11, 1059125. [Google Scholar] [CrossRef]
- Turbelin, A.J.; Malamud, B.D.; Francis, R.A. Mapping the global state of invasive alien species: Patterns of invasion and policy responses. Glob. Ecol. Biogeogr. 2017, 26, 78–92. [Google Scholar] [CrossRef]
- Cornelissen, B.; Neumann, P.; Schweiger, O. Global warming promotes biological invasion of a honey bee pest. Glob Chang. Biol. 2019, 25, 3642–3655. [Google Scholar] [CrossRef] [PubMed]
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Estay, S.; Lima, M.; Labra, F. Predicting insect pest status under climate change scenarios: Combining experimental data and population dynamics modelling. J. Appl. Entomol. 2009, 133, 491–499. [Google Scholar] [CrossRef]
- Ziska, L.H.; Blumenthal, D.M.; Runion, G.B.; Hunt, E.R., Jr.; Diaz-Soltero, H. Invasive species and climate change: An agronomic perspective. Clim. Chang. 2011, 105, 13–42. [Google Scholar] [CrossRef]
- Thomson, L.J.; Macfadyen, S.; Hoffmann, A.A. Predicting the effects of climate change on natural enemies of agricultural pests. Biol. Control 2010, 52, 296–306. [Google Scholar] [CrossRef]
- Zhao, Z.C.; Luo, Y.; Huang, J.B. The detection of the CMIP5 climate model to see the development of the CMIP6 earth system models. Clim. Chang. Res. 2018, 14, 643–648. [Google Scholar]
- Fan, Z.F.; Zhou, B.J.; Ma, C.L.; Gao, C.; Han, D.N.; Chai, Y. Impacts of climate change on species distribution patterns of Polyspora sweet in China. Ecol. Evol. 2022, 12, e9516. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, H.L.; Zhang, H.X.; Chen, Y.F.; Zhang, L.W.; Kawashaer, K.; Dilxadam, T.; Zhang, Y.M. Potential distribution of three types of ephemeral plants under climate changes. Front. Plant Sci. 2022, 13, 1035684. [Google Scholar] [CrossRef]
- Bellotti, A.C.; Smith, L.; Lapointe, S.L. Recent advances in cassava pest management. Annu. Rev. Entomol. 1999, 44, 343–370. [Google Scholar] [CrossRef] [PubMed]
- Bellotti, A.C.; Braun, A.R.; Arias, B.; Castillo, J.A.; Guerrero, J.M. Origin and management of neotropical cassava arthropod pests. Afr. Crop Sci. J. 1994, 2, 407–417. [Google Scholar]
- Parsa, S.; Kondo, T.; Winotai, A. The cassava mealybug (Phenacoccus manihoti) in Asia: First records, potential distribution, and an identification key. PLoS ONE 2012, 7, e47675. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, Q.E.; Pan, X.B.; He, Y.Y.; Huang, Y.; Yan, J. Study on the risk analysis of Phenacoccus manihoti Matile-Ferrero, on imported host products. J. Environ. Entomol. 2014, 36, 298–304. [Google Scholar]
- Löhr, B.; Varela, A.M.; Santos, B. Exploration for natural enemies of the cassava mealybug, Phenacoccus manihoti (Homoptera: Pseudococcidae), in South America for the biological control of this introduced pest in Africa. Bull. Entomol. Res. 1990, 80, 417–425. [Google Scholar] [CrossRef]
- Neuenschwander, P. Biological control of the cassava mealybug in Africa: A review. Biol. Control 2001, 21, 214–229. [Google Scholar] [CrossRef]
- Gupta, A.; Sampathkumar, M.; Mohan, M.; Shylesha, A.N.; Venkatesan, T.; Shashank, P.R.; Dhanyakumar, O.; Ramkumara, P. Assessing adverse impact of the native biological control disruptors in the colonies of the recent invasive pest Phenacoccus manihoti Matile-Ferrero (Hemiptera: Pseudococcidae) in India. Glob. Ecol. Conserv. 2021, 32, e01878. [Google Scholar] [CrossRef]
- Iheagwam, E.U. The influence of temperature on increase rates of the cassava mealybug Phenacoccus manihoti Mat.-Ferr. (Homoptera, Pseudococcidae). Rev. Zool. Afr. 1981, 95, 959–967. [Google Scholar]
- Lema, K.M.; Herren, H.R. The influence of constant temperature on population growth rates of the cassava mealybug, Phenacoccus manihoti. Entomol. Exp. Appl. 1985, 38, 165–169. [Google Scholar] [CrossRef]
- Nwanze, K.F. Relationships between cassava root yields and crop infestations by the mealybug, Phenacoccus manihoti. Int. J. Pest Manag. 1982, 28, 27–32. [Google Scholar] [CrossRef]
- Muniappan, R.; Shepard, B.M.; Watson, G.W.; Carner, G.R.; Rauf, A.; Sartiami, D.; Hidayat, P.; Afun, J.V.K.; Goergen, G.; Rahman, A.Z. New records of invasive insects (hemiptera: Sternorrhyncha) in Southeast Asia and West Africa. J. Agric. Urban Entomol. 2009, 26, 167–174. [Google Scholar] [CrossRef]
- Wang, Y.S.; Tian, H.; Wam, F.H.; Zhang, G.F. Species-specific COI primers for rapid identification of a globally significant invasive pest, the cassava mealybug Phenacoccus manihoti Matile-Ferrero. J. Integr. Agric. 2019, 18, 1042–1049. [Google Scholar] [CrossRef]
- Geng, Y.; Chen, Q.; Liang, X.; Liu, Y.; Wu, C.L.; Chen, Y.H. Flavonoid synthesis pathway-mediate defense mechanism of Cassava Against Phenacoccus manihoti. Chin. J. Trop. Crop. 2023, 44, 2449–2460. [Google Scholar]
- CABI Database. Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.40173 (accessed on 1 May 2024).
- Wu, S.A.; Wang, Y.P. Precaution of cassava mealybug, Phenacoccus manihoti Matile-Ferrero in China. J. Environ. Entomol. 2011, 33, 122–125. [Google Scholar] [CrossRef]
- Kahn, R.P. A concept of pest risk analysis. EPPO Bull. 1979, 9, 119–130. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, S.Q.; Ma, C. Discussion on risk analysis of agricultural plant pests in China. China Plant Prot. 2023, 43, 84–86, 43. [Google Scholar]
- Eyre, A.C.; Briscoe, N.J.; Harley, D.K.; Lumsden, L.F.; McComb, L.B.; Lentini, P.E. Using species distribution models and decision tools to direct surveys and identify potential translocation sites for a critically endangered species. Divers. Distrib. 2022, 28, 700–711. [Google Scholar] [CrossRef]
- Villero, D.; Pla, M.; Camps, D.; Ruiz-Olmo, J.; Brotons, L. Integrating species distribution modelling into decision-making to inform conservation actions. Biodivers. Conserv. 2017, 26, 251–271. [Google Scholar] [CrossRef]
- Rodríguez-Rey, M.; Consuegra, S.; Börger, L.; Garcia de Leaniz, C. Improving Species Distribution Modelling of freshwater invasive species for management applications. PLoS ONE 2019, 14, e0217896. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.M.; Zhang, G.L.; Fu, W.D.; Zhang, Y.; Zhao, Z.H.; Li, Z.H.; Qin, Y.J. Impacts of climate change on climatically suitable regions of two invasive Erigeron weeds in China. Front. Plant Sci. 2023, 14, 1238656. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W. Biomod: Optimizing predictions of species distributions and projecting potential future shifts under global change. Glob. Chang. Biol. 2003, 9, 1353–1362. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Luo, M.; Wang, H.; Lv, Z. Evaluating the performance of species distribution models Biomod2 and MaxEnt using the giant panda distribution data. Chin. J. Appl. Ecol. 2017, 28, 4001–4006. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Lahoz-Monfort, J.J.; Guillera-Arroita, G. Testing whether ensemble modelling is advantageous for maximising predictive performance of species distribution models. Ecography 2020, 43, 549–558. [Google Scholar] [CrossRef]
- Dhami, B.; Bhusal, A.; Adhikari, B.; Miya, M.S.; Maharjan, S.K.; Neupane, D.; Adhikari, H. Habitat Suitability and Conflict Zone Mapping for the Blue Bull (Boselaphus tragocamelus) across Nepal. Animals 2023, 13, 937. [Google Scholar] [CrossRef] [PubMed]
- Čengić, M.; Rost, J.; Remenska, D.; Janse, J.H.; Huijbregts, M.A.; Schipper, A.M. On the importance of predictor choice, modelling technique, and number of pseudo-absences for bioclimatic envelope model performance. Ecol. Evol. 2020, 10, 12307–12317. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, X.; Zong, S. Prediction of the Potential Distribution of Teinopalpus aureus Mell, 1923 (Lepidoptera, Papilionidae) in China Using Habitat Suitability Models. Forests 2024, 15, 828. [Google Scholar] [CrossRef]
- De Oliveira, L.R.; Brito, G.; Gama, M.; Ovando, X.M.C.; Anastácio, P.; Cardoso, S.J. Non-Native Decapods in South America: Risk Assessment and Potential Impacts. Diversity 2023, 15, 841. [Google Scholar] [CrossRef]
- Shabani, F.; Kumar, L.; Al Shidi, R.H.S. Impacts of climate change on infestations of Dubas bug (Ommatissus lybicus Bergevin) on date palms in Oman. PeerJ 2018, 6, e5545. [Google Scholar] [CrossRef] [PubMed]
- GBIF Database. Available online: https://www.gbif.org/.
- Lu, H.; Lu, B.Q.; Liu, H.; Guo, A.P.; Tang, J.H.; Su, H. Risk analysis of regional pests in Hainan. Chin. J. Trop. Agric. 2020, 40, 38–42. [Google Scholar]
- Zhang, Y.; Qin, Y.J.; Zhang, Y.; Fu, W.D.; Zhang, G.L.; Li, Z.H. Integration-based modelling for predicting the habitability zones of the invasive alien plant, Solanum carolinense, in North America. J. Plant Prot. 2022, 49, 1434–1439. [Google Scholar] [CrossRef]
- Vignali, S.; Barras, A.G.; Arlettaz, R.; Braunisch, V. SDMtune: An R package to tune and evaluate species distribution models. Ecol. Evol. 2020, 20, 11488–11506. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Wang, C.; Zhao, Z.; Pan, X.; Li, Z. Climate change impacts on the global potential geographical distribution of the agricultural invasive pest, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Clim. Chang. 2019, 155, 145–156. [Google Scholar] [CrossRef]
- Nelder, J.A.; Wedderburn, R.W. Generalized Linea Models. J. R. Stat. Soc. Ser. A Stat. Soc. 1972, 135, 370–384. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R. Generalized additive models: Some applications. J. Am. Stat. Assoc. 1987, 82, 371–386. [Google Scholar] [CrossRef]
- Ridgeway, G. The state of boosting. Comput. Sci. Stat. 1999, 31, 172–181. [Google Scholar]
- Breiman, L.; Friedman, J.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees; Routledge: New York, NY, USA, 1984; pp. 103–105. [Google Scholar] [CrossRef]
- Friedman, J.; Hastie, T.; Tibshirani, R. Additive logistic regression: A statistical view of boosting (With discussion andare joinder by the authors). Ann. Statist. 2000, 28, 337–407. [Google Scholar] [CrossRef]
- Busby, J.R. Bioclim: A bioclimate analysis and prediction system. Plant Prot. Q. 1991, 6, 8–9. [Google Scholar]
- Hastie, T.; Tibshirani, R.; Buja, A. Flexible Discriminant Analysis by Optimal Scoring. J. Am. Stat. Assoc. 1994, 89, 1255–1270. [Google Scholar] [CrossRef]
- Friedman, J.H. Multivariate adaptive regression splines. Ann. Statist. 1991, 19, 1–67. [Google Scholar] [CrossRef]
- Ripley, B.D. Pattern Recognition and Neural Networks; Cambridge University Press: Cambridge, UK, 2007; pp. 1–15. [Google Scholar]
- Yang, L.Y.; Song, M.L.; Wang, Y.Q.; Wang, H.S.; Zhou, R. Distribution of habitable zones of plateau pika and its response to climate change in Guoluo Prefecture, Qinghai Province. Acta Agrestia Sin. 2024; in press. [Google Scholar]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4 July 2004. [Google Scholar] [CrossRef]
- Li, M.; Zhao, H.; Xian, X.; Zhu, J.; Chen, B.; Jia, T.; Wang, R.; Liu, W. Geographical Distribution Pattern and Ecological Niche of Solenopsis invicta Buren in China under Climate Change. Diversity 2023, 15, 607. [Google Scholar] [CrossRef]
- Resquin, F.; Duque-Lazo, J.; Acosta-Muñoz, C.; Rachid-Casnati, C.; Carrasco-Letelier, L.; Navarro-Cerrillo, R.M. Modelling Current and Future Potential Habitats for Plantations of Eucalyptus grandis Hill ex Maiden and E. dunnii Maiden in Uruguay. Forests 2020, 11, 948. [Google Scholar] [CrossRef]
- Sharma, M.K.; Ram, B.; Chawla, A. Ensemble modelling under multiple climate change scenarios predicts reduction in highly suitable range of habitats of Dactylorhiza hatagirea (D. Don) Soo in Himachal Pradesh, western Himalaya. S. Afr. J. Bot. 2023, 154, 203–218. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, Kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Gao, T.; Shi, J. The Potential Global Distribution of Sirex juvencus (Hymenoptera: Siricidae) under Near Current and Future Climatic Conditions as Predicted by the Maximum EntropyModel. Insects 2021, 12, 222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, S.; Liu, P.; Ming, Z.; Yin, X.; Gao, X. Study on the growth suitability of Nauclea officinalis based on ArcGIS and MaxEnt model. Wild Plant Resour. China 2022, 41, 76–80, 88. (In Chinese) [Google Scholar] [CrossRef]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; d’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. Ecospat: An R package to support spatial analyses and modelling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.-J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef]
- Chan, P.T.; Arroyo-Cabrales, J.; Prieto-Torres, D.A.; Sánchez-González, L.A. The role of ecological niche conservatism in the evolution of bird distributional patterns in Mesoamerican seasonally dry forests. J. Biogeogr. 2024, 51, 1213–1229. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, Z.; Li, C.; Yu, H.; Wang, H. Geographical distribution and ecological niche dynamics of Crassostrea sikamea (Amemiya, 1928) in China’s coastal regions under climate change. Sci. Total Environ. 2024, 920, 171061. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, T.; An, W.; Wang, Z.; Li, C. Predicting the Potential Geographic Distribution of Invasive Freshwater Apple Snail Pomacea canaliculate (Lamarck, 1819) under Climate Change Based on Biomod2. Agronomy 2024, 14, 650. [Google Scholar] [CrossRef]
- Guo, K.Q.; Jiang, X.L.; Xu, G.B. Potential habitat areas of the Quercus lamellosa Sm. and the impact of climate change on their distribution. Chin. J. Ecol. 2021, 40, 2563–2574. [Google Scholar] [CrossRef]
- Guisan, A.; Petitpierre, B.; Broennimann, O.; Daehler, C.; Kuefer, C. Unifying niche shift st-udies: Insights from biological invasions. Trends Ecol. Evol. 2014, 29, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Yonow, T.; Kriticos, D.J.; Ota, N. Thepotential distribution of cassava mealybug (Phenacoccus manihoti), a threat to food security for the poor. PLoS ONE 2017, 12, e0173265. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A review of evidence about use and performance of species distribution modelling ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Wisz, M.S.; Guisan, A. Do pseudo-absence selection strategies influence species distribution models and their predictions? An information-theoretic approach based on simulated data. BMC Ecol. 2009, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Chefaoui, R.M.; Lobo, J.M. Assessing the effects of pseudo-absences on predictive distribution model performance. Ecol. Model. 2008, 210, 478–486. [Google Scholar] [CrossRef]
- Lu, H.; Lu, F.P.; Xu, X.L.; Chen, Q. Environmental suitability of the cassava mealybug assessments using ecological niche modelling. Appl. Mech. Mater. 2014, 651, 1432–1435. [Google Scholar] [CrossRef]
- Ou, W.J.; Luo, X.Q.; An, F.F.; Li, K.M. Analysis of climate change and the possibility of cassava moving northward in China. China Trop. Agric. 2014, 59, 4–8. [Google Scholar]
- Bellotti, A.C. Arthropod Pests. In Cassava: Biology, Production and Utilization; Hillocks, R.J., Thresh, J.M., Bellotti, A.C., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 209–235. [Google Scholar]



| Climate Scenario | Area (×104 km2) | ||
|---|---|---|---|
| Low-Suitable Area | Medium-Suitable Area | High-Suitable Area | |
| Historical climatic scenario | 645,489.1268 | 411,300.4540 | 186,008.8396 |
| SSP1-2.6 (2050) | 719,153.7544 | 510,826.8700 | 178,921.3524 |
| SSP5-8.5 (2050) | 766,730.3972 | 506,906.1324 | 192,417.7376 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Li, T.; Chen, W.; Zhang, Y.; Xu, Y.; Guo, T.; Wang, S.; Liu, J.; Qin, Y. Analysis of the Distribution Pattern of Phenacoccus manihoti in China under Climate Change Based on the Biomod2 Model. Biology 2024, 13, 538. https://doi.org/10.3390/biology13070538
Huang Y, Li T, Chen W, Zhang Y, Xu Y, Guo T, Wang S, Liu J, Qin Y. Analysis of the Distribution Pattern of Phenacoccus manihoti in China under Climate Change Based on the Biomod2 Model. Biology. 2024; 13(7):538. https://doi.org/10.3390/biology13070538
Chicago/Turabian StyleHuang, Yumeng, Tong Li, Weijia Chen, Yuan Zhang, Yanling Xu, Tengda Guo, Shuping Wang, Jingyuan Liu, and Yujia Qin. 2024. "Analysis of the Distribution Pattern of Phenacoccus manihoti in China under Climate Change Based on the Biomod2 Model" Biology 13, no. 7: 538. https://doi.org/10.3390/biology13070538
APA StyleHuang, Y., Li, T., Chen, W., Zhang, Y., Xu, Y., Guo, T., Wang, S., Liu, J., & Qin, Y. (2024). Analysis of the Distribution Pattern of Phenacoccus manihoti in China under Climate Change Based on the Biomod2 Model. Biology, 13(7), 538. https://doi.org/10.3390/biology13070538