
Improvement in Salt Tolerance Ability of Pseudomonas putida KT2440



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Transcriptome Analysis
2.3. Quantitative Real-Time PCR (qRT-PCR) for Verification of Differentially Expressed Genes
2.4. Gene Overexpression
2.5. Scanning Electron Micrograph
2.6. Biodegradation and Detection of Degradants
3. Results
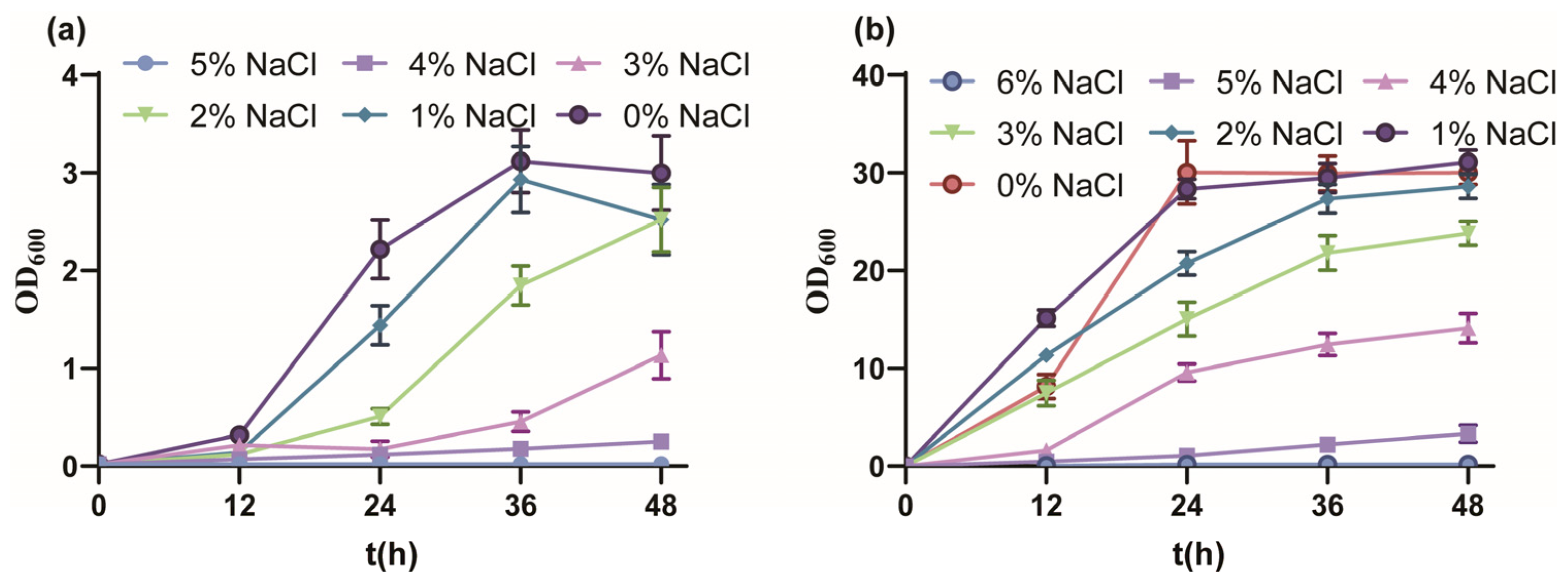
3.1. The Salt Tolerance Performance of P. putida KT2440
3.2. Transcriptomic Analysis of P. putida KT2440 under NaCl Stress Conditions
3.2.1. DEGs Analysis
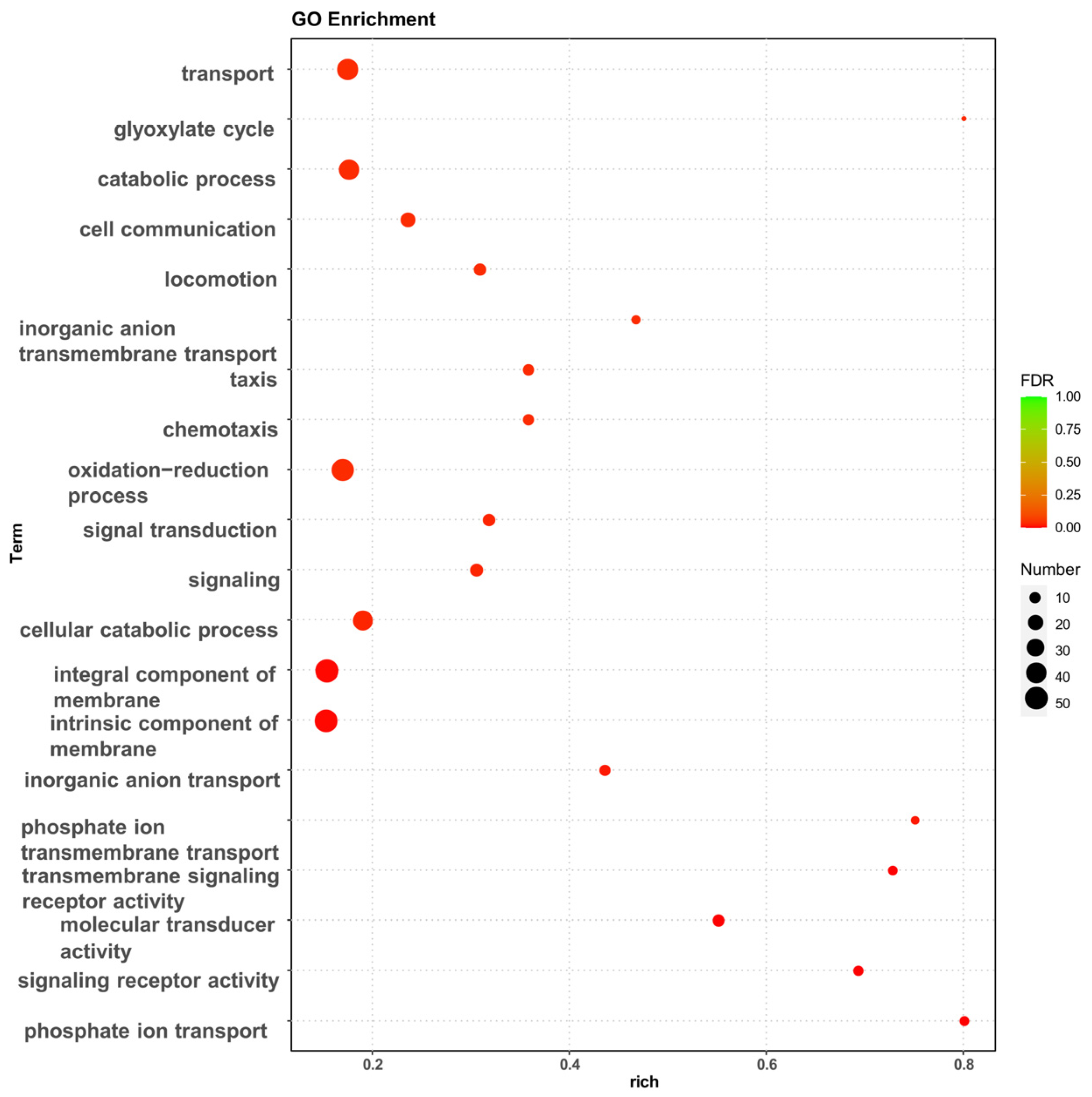
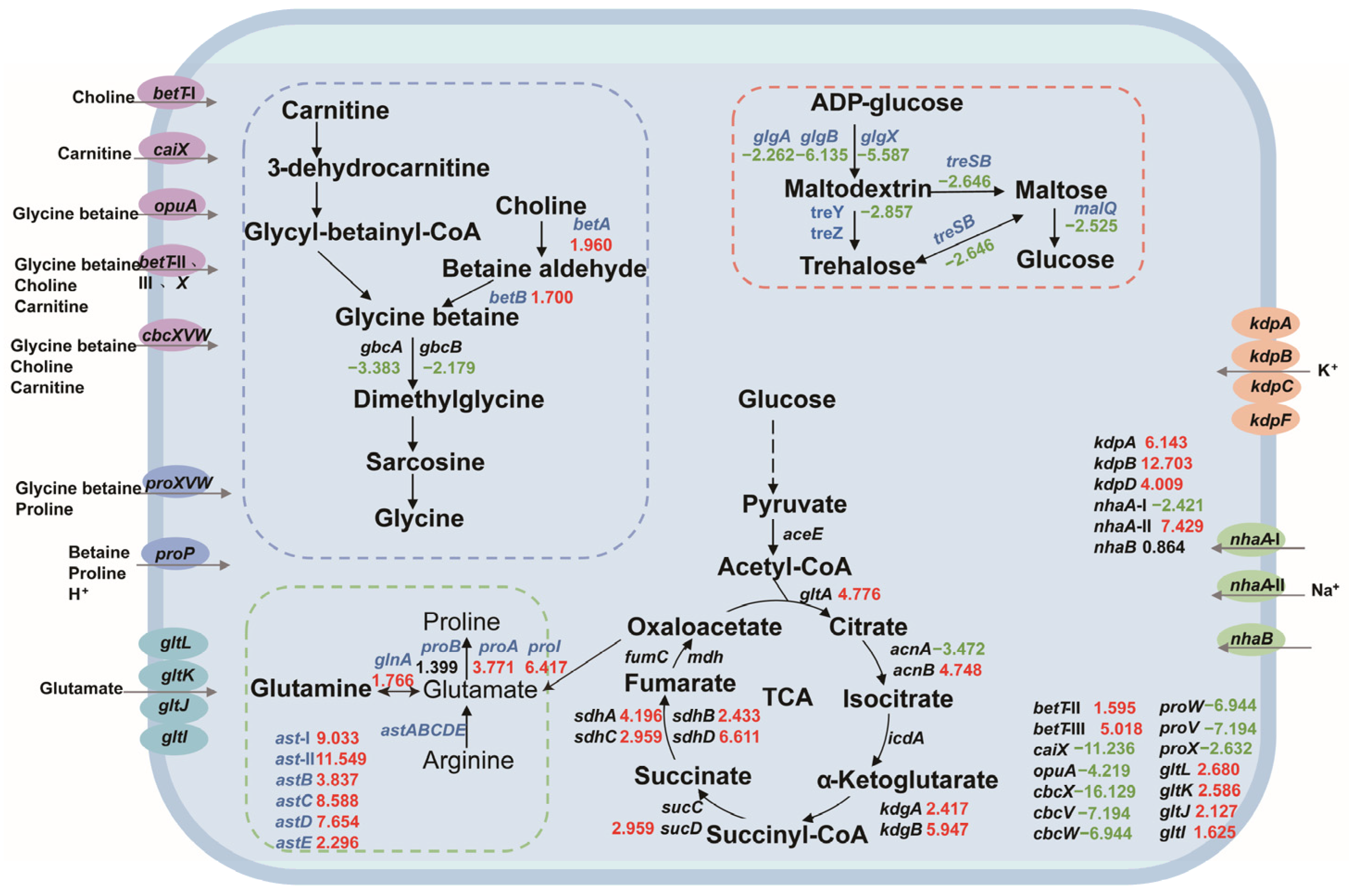
3.2.2. Functional Annotation and Verification of DEGs
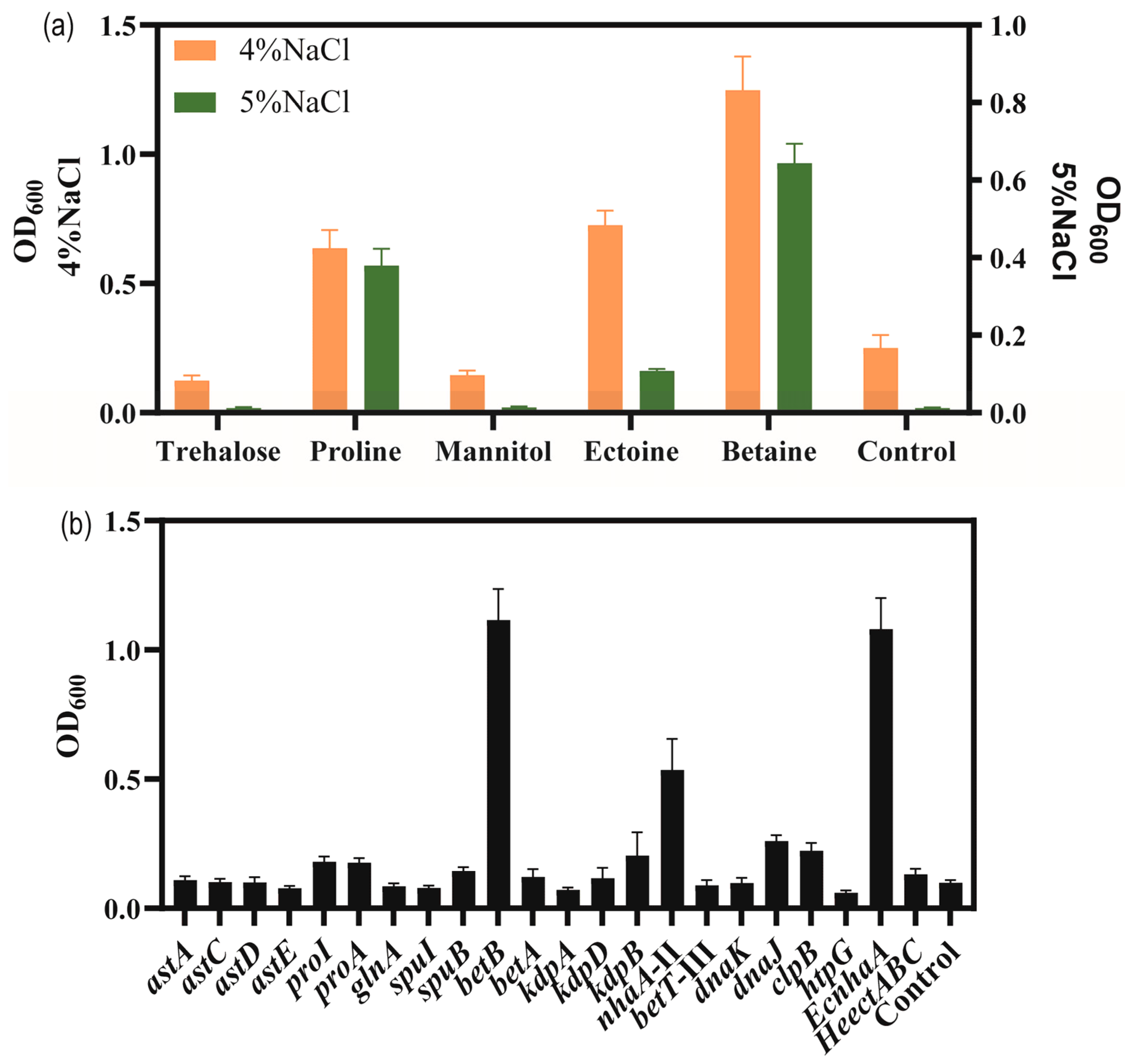
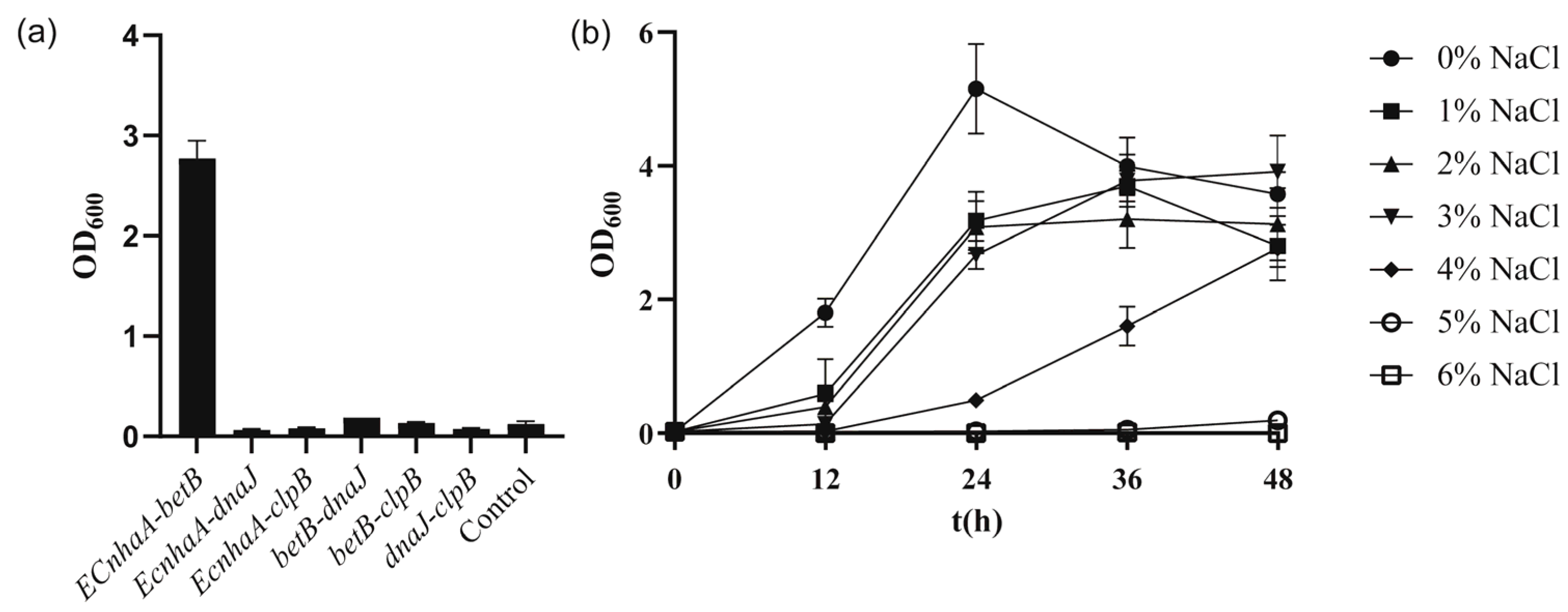
3.3. Improve the Salt Tolerance of Strain KT2440
3.4. The Cell Morphology of the Salt-Tolerant Strain
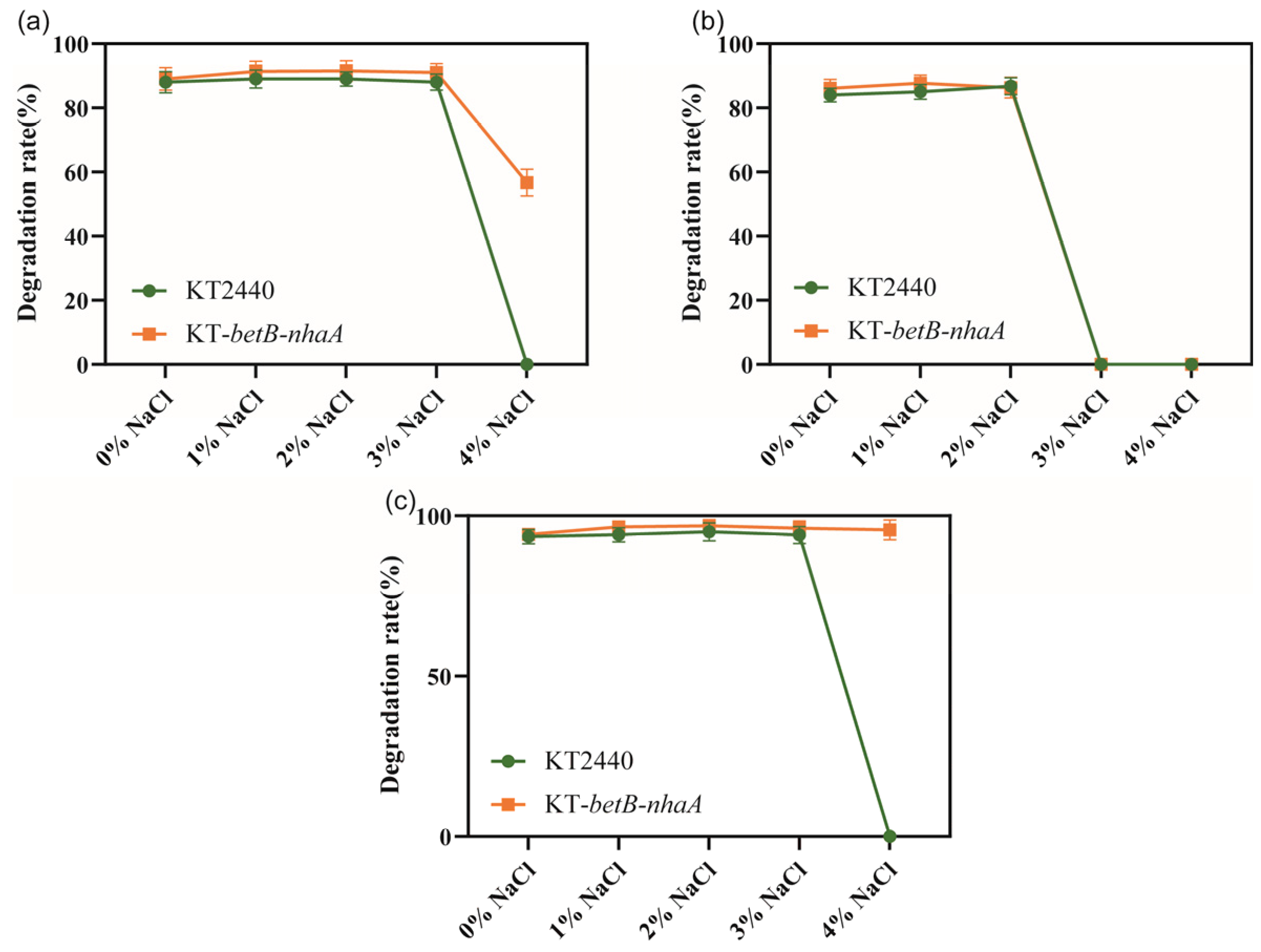
3.5. The Degradation Properties of Engineered Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vreeland, R.H.; Hochstein, L.I. The Biology of Halophilic Bacteria; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Oren, A. Microbial life at high salt concentrations: Phylogenetic and metabolic diversity. Saline Syst. 2008, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Padan, E.; Venturi, M.; Gerchman, Y.; Dover, N. Na+/H+ antiporters. Biochim. Biophs. Acta. 2001, 1505, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Wani, A.K.; Akhtar, N.; Sher, F.; Navarrete, A.A.; Américo-Pinheiro, J.H.P. Microbial adaptation to different environmental conditions: Molecular perspective of evolved genetic and cellular systems. Arch. Microbiol. 2022, 204, 144. [Google Scholar] [CrossRef]
- Kumar, S.; Paul, D.; Bhushan, B.; Wakchaure, G.C.; Meena, K.K.; Shouche, Y. Traversing the “Omic” landscape of microbial halotolerance for key molecular processes and new insights. Crit. Rev. Microbiol. 2020, 46, 631–653. [Google Scholar] [CrossRef] [PubMed]
- Gunde-Cimerman, N.; Plemenitas, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. Fems. Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, E.; Sharma, S.K.; Kashyap, A.S.; Manzar, N.; Sahu, P.K.; Singh, U.B.; Singh, H.V.; Sharma, P.K. Evaluation of osmotolerant potential of Halomonas sulfidaeris MV-19 isolated from a mud volcano. Curr. Microbiol. 2023, 80, 102. [Google Scholar] [CrossRef] [PubMed]
- Goswami, S.K.; Kashyap, A.S.; Kumar, R.; Gujjar, R.S.; Singh, A.; Manzar, N. Harnessing rhizospheric microbes for eco-friendly and sustainable crop production in saline environments. Curr. Microbiol. 2023, 81, 14. [Google Scholar] [CrossRef] [PubMed]
- Dimroth, P.; Jockel, P.; Schmid, M. Coupling mechanism of the oxaloacetate decarboxylase Na+ pump. Biochim. Biophs. Acta. 2001, 1505, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lietzan, A.D.; St Maurice, M. Functionally diverse biotin-dependent enzymes with oxaloacetate decarboxylase activity. Arch. Biochem. Biophys. 2014, 544, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Vitt, S.; Prinz, S.; Hellwig, N.; Morgner, N.; Ermler, U.; Buckel, W. Molecular and Low-Resolution Structural Characterization of the Na+-Translocating Glutaconyl-CoA Decarboxylase From. Front. Microbiol. 2020, 11, 480. [Google Scholar] [CrossRef]
- Becher, B.; Muller, V.; Gottschalk, G. The Methyl-Tetrahydromethanopterin-Coenzyme-M Methyltransferase of Methanosarcina Strain Go1 Is a Primary Sodium-Pump. Fems. Microbiol. Lett. 1992, 91, 239–244. [Google Scholar] [CrossRef]
- Takase, K.; Kakinuma, S.; Yamato, I.; Konishi, K.; Igarashi, K.; Kakinuma, Y. Sequencing and characterization of the ntp gene cluster for vacuolar-type Na(+)-translocating ATPase of Enterococcus hirae. J. Biol. Chem. 1994, 269, 11037–11044. [Google Scholar] [CrossRef] [PubMed]
- Verkhovsky, M.I.; Bogachev, A.V. Sodium-translocating NADH:quinone oxidoreductase as a redox-driven ion pump. Biochim. Biophs. Acta 2010, 1797, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.F.; Jiang, J.Q.; Zhang, B.; Zhao, B.S.; Wang, L.; Yang, S.S. A primary sodium pump gene of the moderate halophile exhibits secondary antiporter properties. Biochem. Biophys. Res. Commun. 2006, 346, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Mager, T.; Braner, M.; Kubsch, B.; Hatahet, L.; Alkoby, D.; Rimon, A.; Padan, E.; Fendler, K. Differential effects of mutations on the transport properties of the Na+/H+ antiporter NhaA from Escherichia coli. J. Biol. Chem. 2013, 288, 24666–24675. [Google Scholar] [CrossRef] [PubMed]
- Padan, E. Functional and structural dynamics of NhaA, a prototype for Na+ and H+ antiporters, which are responsible for Na+ and H+ homeostasis in cells. Biochim. Biophs. Acta. 2014, 1837, 1047–1062. [Google Scholar] [CrossRef] [PubMed]
- Pinner, E.; Kotler, Y.; Padan, E.; Schuldiner, S. Physiological role of nhaB, a specific Na+/H+ antiporter in Escherichia coli. J. Biol. Chem. 1993, 268, 1729–1734. [Google Scholar] [CrossRef] [PubMed]
- Prágai, Z.; Eschevins, C.; Bron, S.; Harwood, C.R. Bacillus subtilis NhaC, an Na+/H+ antiporter, influences expression of the phoPR operon and production of alkaline phosphatases. J. Bacteriol. 2001, 183, 2505–2515. [Google Scholar] [CrossRef] [PubMed]
- Dzioba, J.; Ostroumov, E.; Winogrodzki, A.; Dibrov, P. Cloning, functional expression in Escherichia coli and primary characterization of a new Na+/H+ antiporter, NhaD, of Vibrio cholerae. Mol. Cell. Biochem. 2002, 229, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Patiño-Ruiz, M.; Ganea, C.; Calinescu, O. Prokaryotic Na+/H+ exchangers—Transport mechanism and essential residues. Int. J. Mol. Sci. 2022, 23, 9156. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Cheng, H.; Liu, Q.; Liu, J.; Ma, Y. Research progress of the Na+/H+ antiporters in bacteria. Microbiol. China 2015, 42, 2002–2011. [Google Scholar]
- Wang, W.; Tang, H.; Xu, P. Salt-tolerance related genes in halophilic bacteria and archaea. Microbiology China 2015, 42, 550–558. [Google Scholar]
- Edbeib, M.F.; Wahab, R.A.; Huyop, F. Halophiles: Biology, adaptation, and their role in decontamination of hypersaline environments. World J. Microb. Biotechnol. 2016, 32, 135–158. [Google Scholar] [CrossRef]
- Roberts, M.F. Organic compatible solutes of halotolerant and halophilic microorganisms. Saline Syst. 2005, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Waditee, R.; Tanaka, Y.; Aoki, K.; Hibino, T.; Jikuya, H.; Takano, J.; Takabe, T.; Takabe, T. Isolation and Functional Characterization of N-Methyltransferases That Catalyze Betaine Synthesis from Glycine in a Halotolerant Photosynthetic Organism Aphanothece halophytica. J. Biol. Chem. 2003, 278, 4932–4942. [Google Scholar] [CrossRef]
- Brill, J.; Hoffmann, T.; Bleisteiner, M.; Bremer, E. Osmotically Controlled Synthesis of the Compatible Solute Proline Is Critical for Cellular Defense of against High Osmolarity. J. Bacteriol. 2011, 193, 5335–5346. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M.; Bremer, E.; Csonka, L.N.; Kraemer, R.; Poolman, B.; van der Heide, T.; Smith, L.T. Osmosensing and osmoregulatory compatible solute accumulation by bacteria. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 130, 437–460. [Google Scholar] [CrossRef]
- Zahid, N.; Schweiger, P.; Galinski, E.; Deppenmeier, U. Identification of mannitol as compatible solute in Gluconobacter oxydans. Appl. Microbiol. Biot. 2015, 99, 5511–5521. [Google Scholar] [CrossRef] [PubMed]
- Galleguillos, P.A.; Grail, B.M.; Hallberg, K.B.; Demergasso, C.S.; Johnson, D.B. Identification of trehalose as a compatible solute in different species of acidophilic bacteria. J. Microbiol. 2018, 56, 727–733. [Google Scholar] [CrossRef]
- Kadam, P.; Khisti, M.; Ravishankar, V.; Barvkar, V.; Dhotre, D.; Sharma, A.; Shouche, Y.; Zinjarde, S. Recent advances in production and applications of ectoine, a compatible solute of industrial relevance. Bioresour. Technol. 2024, 393, 130016. [Google Scholar] [CrossRef] [PubMed]
- Fenizia, S.; Thume, K.; Wirgenings, M.; Pohnert, G. Ectoine from Bacterial and Algal Origin Is a Compatible Solute in Microalgae. Mar. Drugs 2020, 18, 42. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A.; Nieto, J.J.; Oren, A. Biology of moderately halophilic aerobic bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 504–544. [Google Scholar] [CrossRef] [PubMed]
- Papoušková, K.; Sychrová, H. Production of Yarrowia lipolytica Nha2 Na+/H+ antiporter improves the salt tolerance of Saccharomyces cerevisiae. Folia Microbiol. 2007, 52, 600–602. [Google Scholar] [CrossRef] [PubMed]
- Sanadhya, P.; Agarwal, P.; Khedia, J.; Agarwal, P.K. A low-affinity K+ transporter AlHKT2;1 from recretohalophyte aeluropus lagopoides confers salt tolerance in yeast. Mol. Biotechnol. 2015, 57, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Willscher, S.; Jablonski, L.; Fona, Z.; Rahmi, R.; Wittig, J. Phytoremediation experiments with Helianthus tuberosus under different pH and heavy metal soil concentrations. Hydrometallurgy 2017, 168, 153–158. [Google Scholar] [CrossRef]
- Tang, L.; Hamid, Y.; Zehra, A.; Sahito, Z.A.; He, Z.; Hussain, B.; Gurajala, H.K.; Yang, X. Characterization of fava bean (Vicia faba L.) genotypes for phytoremediation of cadmium and lead co-contaminated soils coupled with agro-production. Ecotoxicol. Environ. Saf. 2019, 171, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Lin, Z.; Pang, S.; Zhang, W.; Bhatt, P.; Chen, S. Recent advanced technologies for the characterization of xenobiotic-degrading microorganisms and microbial communities. Front. Bioeng. Biotechnol. 2021, 9, 632059. [Google Scholar] [CrossRef] [PubMed]
- Huo, K.Y.; Wang, S.Q.; Zhao, W.W.; Guo, H.F.; Xiong, W.N.; Liu, R.H.; Yang, C. Creating an efficient 1,2-dichloroethane-mineralizing bacterium by a combination of pathway engineering and promoter engineering. Sci. Total. Environ. 2023, 878, 163140. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, E.; de Lorenzo, V. Molecular tools and emerging strategies for deep genetic/genomic refactoring of Pseudomonas. Curr. Opin. Biotechnol. 2017, 47, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Garcia, E.; de Lorenzo, V. Pseudomonas putida as a synthetic biology chassis and a metabolic engineering platform. Curr. Opin. Biotechnol. 2023, 85, 103025. [Google Scholar] [CrossRef] [PubMed]
- Nikel, P.I.; de Lorenzo, V. Pseudomonas putida as a functional chassis for industrial biocatalysis: From native biochemistry to trans-metabolism. Metab. Eng. 2018, 50, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Mu, Q.X.; Xue, Y.B.; Xue, Y.F.; Yu, B.; Ma, Y.H. One major facilitator superfamily transporter is responsible for propionic acid tolerance in Pseudomonas putida KT2440. Microb. Biotechnol. 2021, 14, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.H.; Jin, X.Z.; Tao, Y.M.; Zheng, Z.J.; Ouyang, J. Unraveling the mechanism of furfural tolerance in engineered Pseudomonas putida by genomics. Front. Microbiol. 2022, 13, 1035263. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, E.T.; Werner, A.Z.; Salvachúa, D.; Singer, C.A.; Szostkiewicz, K.; Jiménez-Díaz, M.R.; Eng, T.; Radi, M.S.; Simmons, B.A.; Mukhopadhyay, A.; et al. Adaptive laboratory evolution of Pseudomonas putida KT2440 improves p-coumaric and ferulic acid catabolism and tolerance. Metab. Eng. Commun. 2020, 11, e00143. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.; Niqui-Arroyo, J.L.; Conde, S.; Ramos, J.L.; Duque, E. Enhanced tolerance to naphthalene and enhanced rhizoremediation performance for Pseudomonas putida KT2440 via the NAH7 catabolic plasmid. Appl. Environ. Microb. 2012, 78, 5104–5110. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Liu, R.H.; Che, Y.; Xu, X.Q.; Zhao, F.J.; Yu, H.L.; Song, C.J.; Liu, Y.P.; Yang, C. Engineering Pseudomonas putida KT2440 for simultaneous degradation of carbofuran and chlorpyrifos. Microb. Biotechnol. 2016, 9, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Zhuang, X.M.; Ahmad, S.; Sung, S.; Ni, S.Q. Biotreatment of high-salinity wastewater: Current methods and future directions. World J. Microbiol. Biotechnol. 2020, 36, 37. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.J.; Chen, L.; Ding, Y.C.; Ma, X.J.; How, S.W.; Wu, D. Mechanism on the microbial salt tolerance enhancement by electrical stimulation. Bioelectrochemistry 2022, 147, 108206. [Google Scholar] [CrossRef] [PubMed]
- Belda, E.; van Heck, R.G.A.; Lopez-Sanchez, M.J.; Cruveiller, S.; Barbe, V.; Fraser, C.; Klenk, H.P.; Petersen, J.; Morgat, A.; Nikel, P.I.; et al. The revisited genome of Pseudomonas putida KT2440 enlightens its value as a robust metabolic chassis. Environ. Microbiol. 2016, 18, 3403–3424. [Google Scholar] [CrossRef] [PubMed]
- Borchert, A.J.; Bleem, A.; Beckham, G.T. RB-TnSeq identifies genetic targets for improved tolerance of Pseudomonas putida towards compounds relevant to lignin conversion. Metab. Eng. 2023, 77, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Bojanovic, K.; D’Arrigo, I.; Long, K.S. Global Transcriptional Responses to Osmotic, Oxidative, and lnnipenem Stress Conditions in. Appl. Environ. Microbiol. 2017, 83, e03236-16. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.G.; Wang, X.Y.; Yin, G.B.; Gai, Z.H.; Tang, H.Z.; Ma, C.Q.; Deng, Z.X.; Xu, P. New metabolites in dibenzofuran cometabolicdegradation by a biphenyl-cultivated Pseudomonas putida srain B6-2. Environ. Sci. Technol. 2009, 43, 8635–8642. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W.; Bilal, M.; Zong, Y.N.; Hu, H.B.; Wang, W.; Zhang, X.H. Development of a Plasmid-Free Biosynthetic Pathway for Enhanced Muconic Acid Production in Pseudomonas chlororaphis HT66. Acs Synth. Biol. 2018, 7, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schwibbert, K.; Marin-Sanguino, A.; Bagyan, I.; Heidrich, G.; Lentzen, G.; Seitz, H.; Rampp, M.; Schuster, S.C.; Klenk, H.P.; Pfeiffer, F.; et al. A blueprint of ectoine metabolism from the genome of the industrial producer Halomonas elongata DSM 2581T. Environ. Microbiol. 2011, 13, 1973–1994. [Google Scholar] [CrossRef] [PubMed]
- Padan, E.; Tzubery, T.; Herz, K.; Kozachkov, L.; Rimon, A.; Galili, L. NhaA of Escherichia coli, as a model of a pH-regulated Na+/H+antiporter. Biochim. Biophs. Acta 2004, 1658, 2–13. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Jiang, J.; Jia, K.; Huang, X.; Li, S. Glycine betaine supplied exogenously enhance salinity tolerance of Pseudomonas putida DLL-1. Wei Sheng Wu Xue Bao 2006, 46, 154–157. [Google Scholar] [PubMed]
- Gralla, J.D.; Vargas, D.R. Potassium glutamate as a transcriptional inhibitor during bacterial osmoregulation. EMBO J. 2006, 25, 1515–1521. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Fold Change (T/C) | p-Value | Description |
|---|---|---|---|---|
| PP_4158 | kdpD | 4.009 | 0.0003 | Sensor protein KdpD |
| PP_4160 | kdpB | 12.703 | 0.0000 | K+ transporting ATPase, KdpB subunit |
| PP_4161 | kdpA | 6.143 | 0.0087 | Potassium-transporting ATPase A chain |
| PP_3958 | nhaA-II | 7.429 | 0.0000 | Na+/H+ antiporter NhaA-II |
| PP_3957 | betB-III | 5.018 | 0.0000 | Choline/carnitine/betaine transporter |
| PP_5064 | betA | 1.960 | 0.0348 | Choline dehydrogenase |
| PP_5063 | betB | 1.700 | 0.0878 | NAD-dependent betaine aldehyde dehydrogenase |
| PP_4479 | astA | 9.033 | 0.0000 | Arginine N-succinyltransferase |
| PP_4477 | astB | 3.837 | 0.0000 | N-succinylarginine dihydrolase |
| PP_4481 | astC | 8.588 | 0.0000 | Succinylornithine transaminase/acetylornithine aminotransferase |
| PP_4478 | astD | 7.654 | 0.0000 | N-succinylglutamate 5-semialdehyde |
| PP_4475 | astE | 2.296 | 0.0044 | Succinylglutamate desuccinylase |
| PP_5095 | proI | 6.417 | 0.0012 | Pyrroline-5-carboxylate reductase |
| PP_4811 | proA | 3.770 | 0.0000 | Gamma-glutamyl phosphate reductase |
| PP_5184 | spuI | 2.212 | 0.0072 | Glutamylpolyamine synthetase |
| PP_5046 | glnA | 1.776 | 0.0386 | Glutamine synthetase |
| PP_4727 | dnaK | 3.217 | 0.0000 | Molecular chaperone DnaK |
| PP_0625 | clpB | 2.507 | 0.0564 | ATP-dependent chaperone ClpB |
| PP_4726 | dnaJ | 4.255 | 0.0000 | Molecular chaperone DnaJ |
| PP_4179 | htpG | 5.356 | 0.0000 | Molecular chaperone HtpG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, M.; Tan, S.; Wang, W.; Zhang, X. Improvement in Salt Tolerance Ability of Pseudomonas putida KT2440. Biology 2024, 13, 404. https://doi.org/10.3390/biology13060404
Fan M, Tan S, Wang W, Zhang X. Improvement in Salt Tolerance Ability of Pseudomonas putida KT2440. Biology. 2024; 13(6):404. https://doi.org/10.3390/biology13060404
Chicago/Turabian StyleFan, Min, Shuyu Tan, Wei Wang, and Xuehong Zhang. 2024. "Improvement in Salt Tolerance Ability of Pseudomonas putida KT2440" Biology 13, no. 6: 404. https://doi.org/10.3390/biology13060404
APA StyleFan, M., Tan, S., Wang, W., & Zhang, X. (2024). Improvement in Salt Tolerance Ability of Pseudomonas putida KT2440. Biology, 13(6), 404. https://doi.org/10.3390/biology13060404