

Resveratrol and (-)-Epigallocatechin-3-gallate Regulate Lipid Metabolism by Activating the AMPK Pathway in Hepatocytes


Abstract
Simple Summary
Abstract
1. Introduction
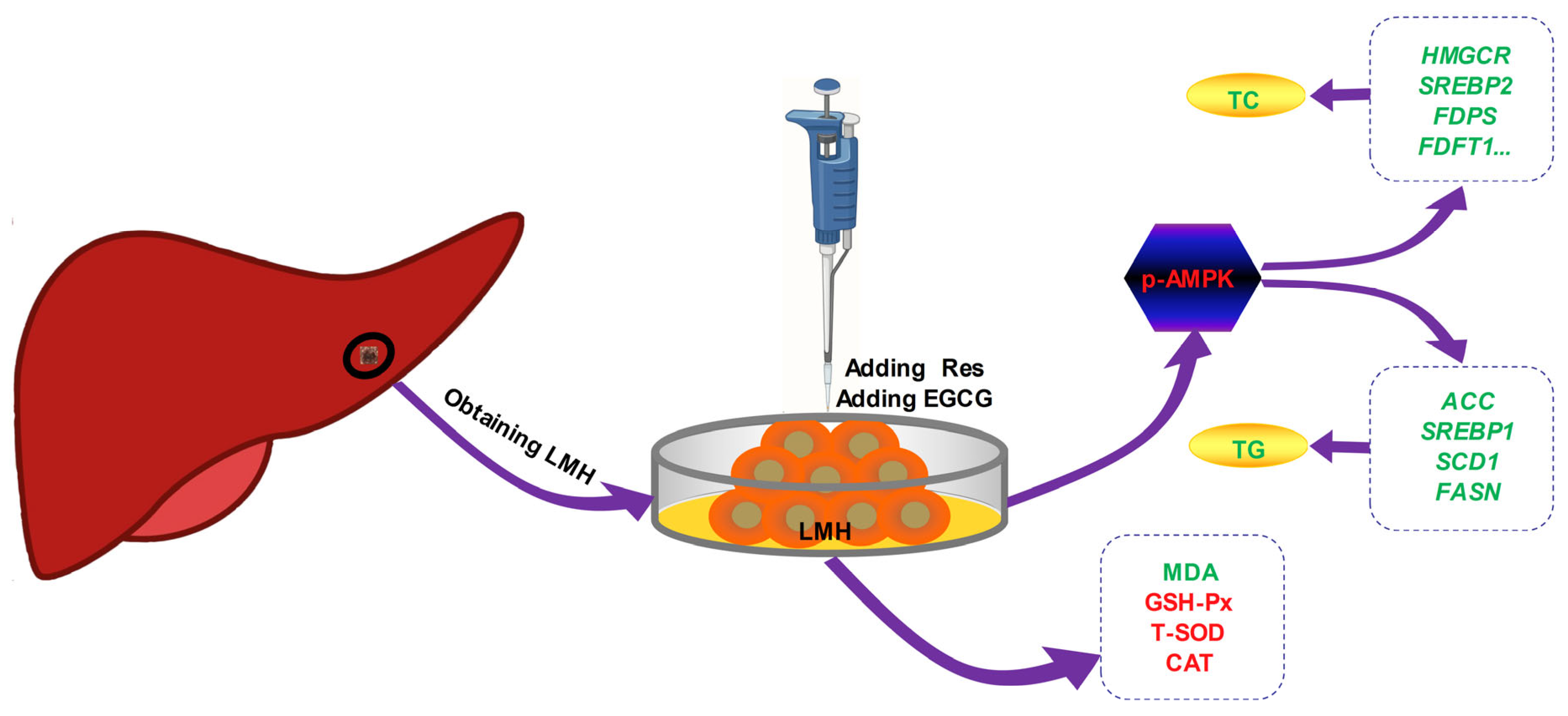
2. Materials and Methods
2.1. Materials and Reagents
2.2. Cell Culture
2.3. Cell Viability Assay (CCK-8)
2.4. Cell Growth Trends
2.5. Determination of Cellular TC and TG Content
2.6. Determination of Cellular Antioxidant Levels
2.7. RNA Extraction and cDNA Synthesis
2.8. Real-Time q-PCR
2.9. Western Blotting Analysis
2.10. Statistic Analysis
3. Results
3.1. Effects of Res and EGCG on Cell Viability and Cell Growth
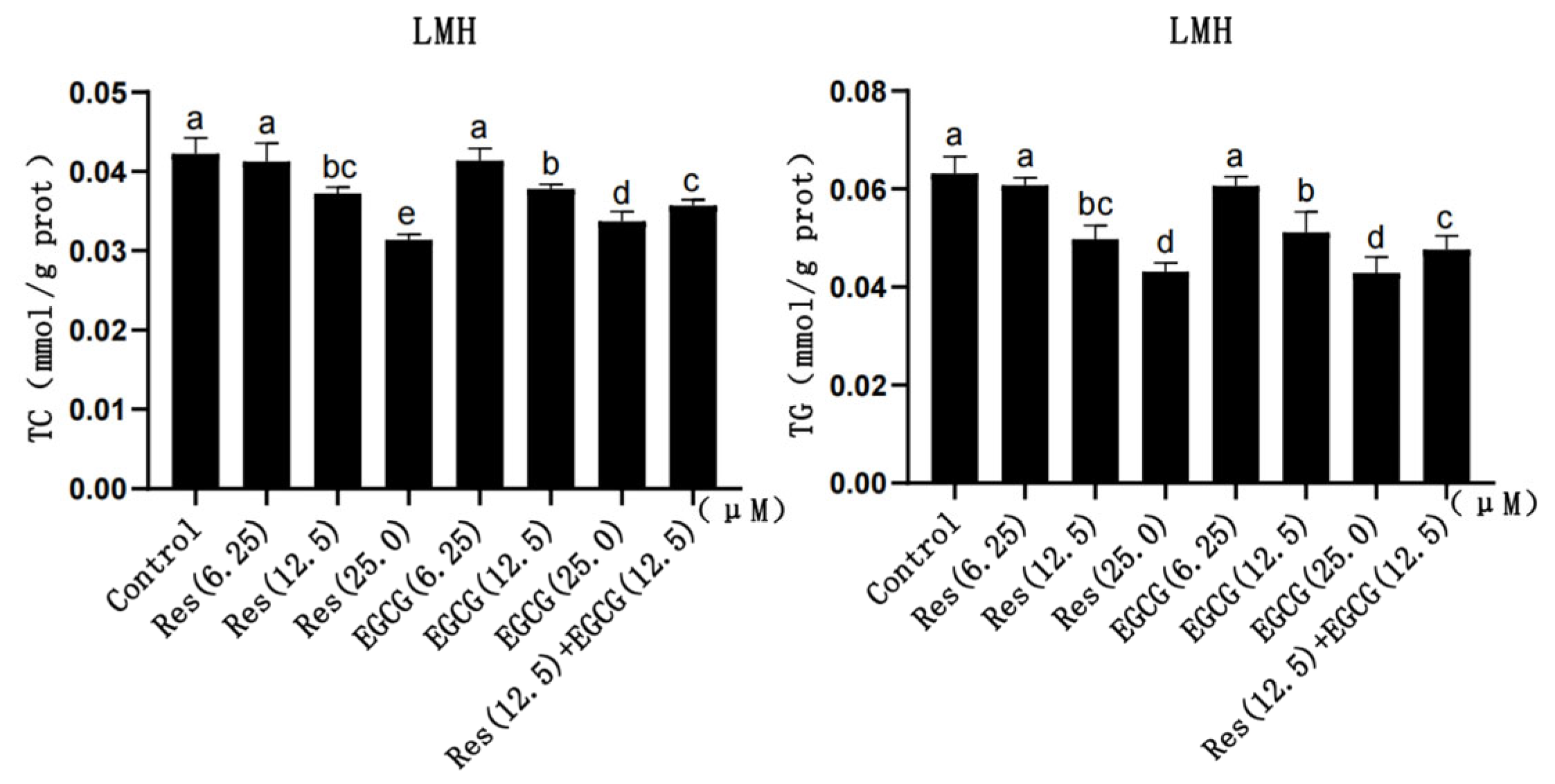
3.2. TC and TG Content
3.3. Changes in Antioxidant Capacity
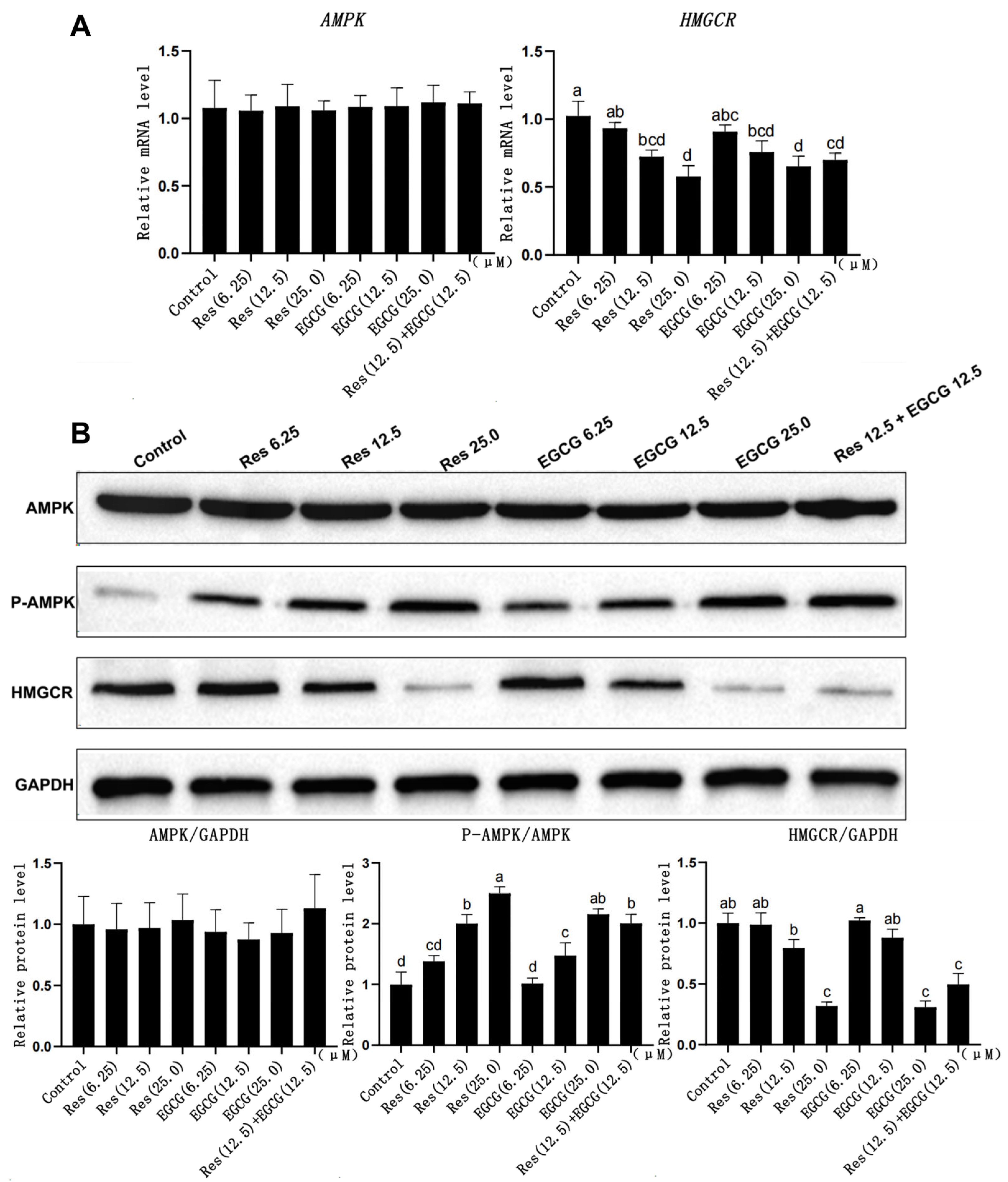
3.4. Changes in Gene and Protein Expression Levels of the AMPK/HMGCR Pathway
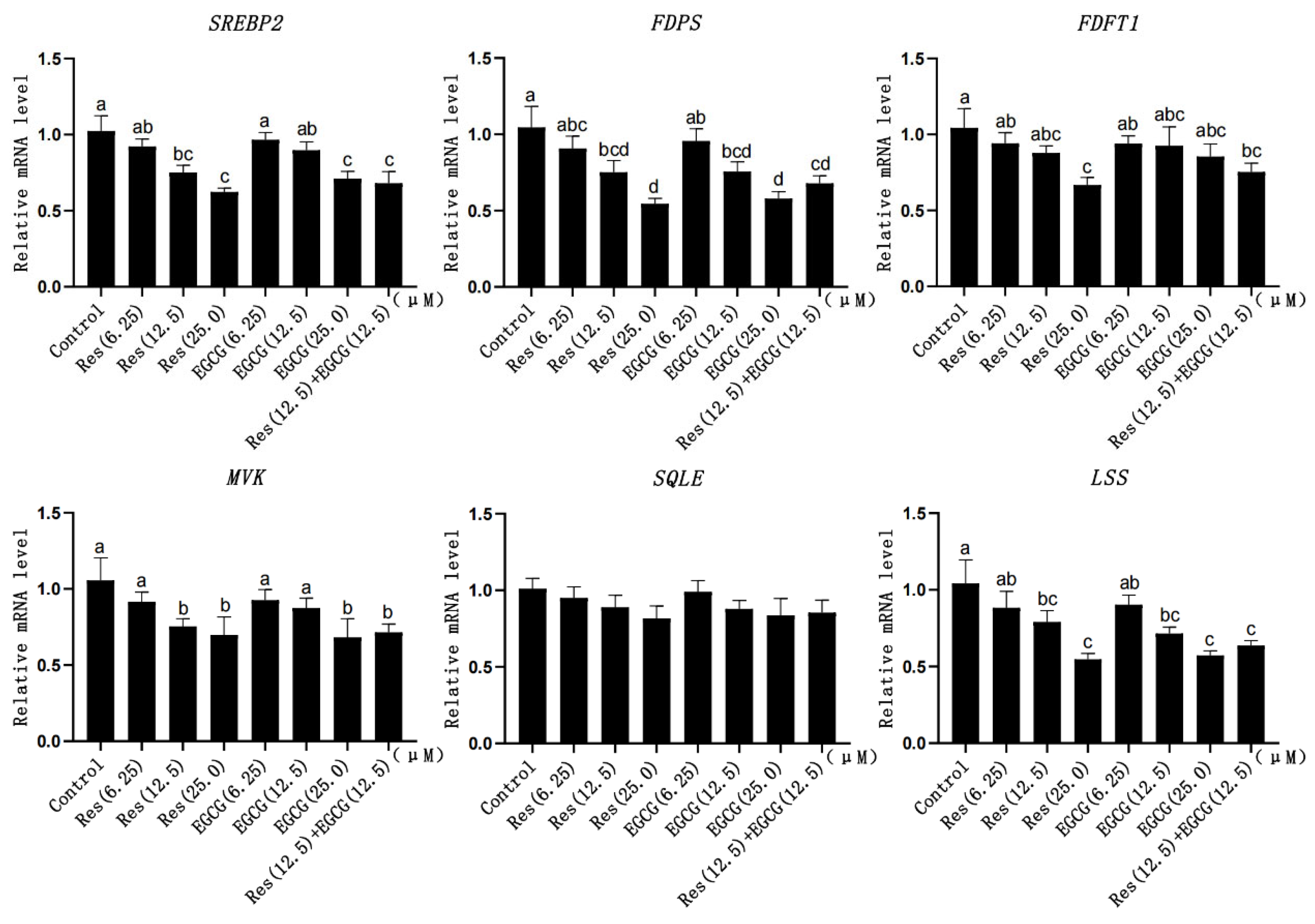
3.5. Expression Changes of Cholesterol Synthesis-Related Genes
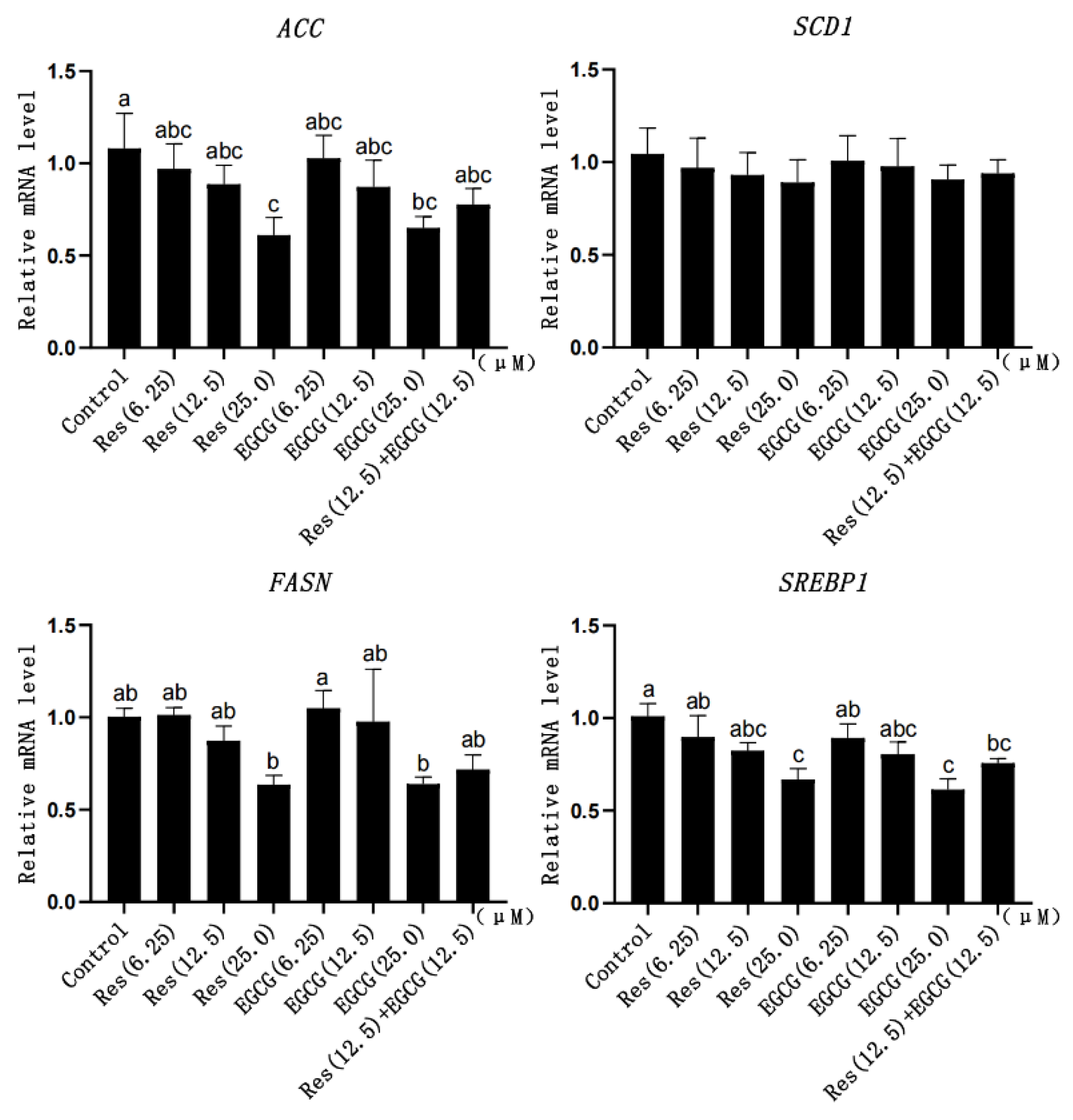
3.6. Expression Changes of Fatty Acid Synthesis-Related Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagarajan, S.R.; Paul-Heng, M.; Krycer, J.R.; Fazakerley, D.J.; Sharland, A.F.; Hoy, A.J. Lipid and glucose metabolism in hepatocyte cell lines and primary mouse hepatocytes: A comprehensive resource for in vitro studies of hepatic metabolism. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E578–E589. [Google Scholar] [CrossRef]
- Bian, X.; Liu, R.; Meng, Y.; Xing, D.; Xu, D.; Lu, Z. Lipid metabolism and cancer. J. Exp. Med. 2021, 218, e20201606. [Google Scholar] [CrossRef] [PubMed]
- Walhin, J.P.; Gonzalez, J.T.; Betts, J.A. Physiological responses to carbohydrate overfeeding. Curr. Opin. Clin. Nutr. Metab. Care 2021, 24, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Ishigaki, Y.; Katagiri, H.; Yamada, T.; Ogihara, T.; Imai, J.; Uno, K.; Hasegawa, Y.; Gao, J.; Ishihara, H.; Shimosegawa, T.; et al. Dissipating excess energy stored in the liver is a potential treatment strategy for diabetes associated with obesity. Diabetes 2005, 54, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Repa, J.J.; Mangelsdorf, D.J. The role of orphan nuclear receptors in the regulation of cholesterol homeostasis. Annu. Rev. Cell Dev. Biol. 2000, 16, 459–481. [Google Scholar] [CrossRef]
- Luo, J.; Yang, H.; Song, B.L. Mechanisms and regulation of cholesterol homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wu, K.; Mi, X.; Rajput, S.A.; Qi, D. Effects of 3-Hydroxy-3-methylglutaryl-CoA reductase inhibitors on cholesterol metabolism in laying hens. Animals 2023, 13, 1868. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, S.; Zhai, A.; Zhang, B.; Tian, G. AMPK-Mediated regulation of lipid metabolism by phosphorylation. Biol. Pharm. Bull. 2018, 41, 985–993. [Google Scholar] [CrossRef]
- Hwang, Y.P.; Kim, H.G.; Choi, J.H.; Do, M.T.; Chung, Y.C.; Jeong, T.C.; Jeong, H.G. S-allyl cysteine attenuates free fatty acid-induced lipogenesis in human HepG2 cells through activation of the AMP-activated protein kinase-dependent pathway. J. Nutr. Biochem. 2013, 24, 1469–1478. [Google Scholar] [CrossRef]
- Wang, M.Y.; Zhang, S.S.; An, M.F.; Xia, Y.F.; Fan, M.S.; Sun, Z.R.; Zhang, L.J.; Zhao, Y.L.; Sheng, J.; Wang, X.J. Neferine ameliorates nonalcoholic steatohepatitis through regulating AMPK pathway. Phytomedicine 2023, 114, 154798. [Google Scholar] [CrossRef]
- Dolinsky, V.W.; Chakrabarti, S.; Pereira, T.J.; Oka, T.; Levasseur, J.; Beker, D.; Zordoky, B.N.; Morton, J.S.; Nagendran, J.; Lopaschuk, G.D.; et al. Resveratrol prevents hypertension and cardiac hypertrophy in hypertensive rats and mice. Biochim. Biophys. Acta 2013, 1832, 1723–1733. [Google Scholar] [CrossRef] [PubMed]
- Price, N.L.; Gomes, A.P.; Ling, A.J.; Duarte, F.V.; Martin-Montalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. SIRT1 is required for AMPK activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab. 2012, 15, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Chen, G.; Hu, T.; Mo, X.; Hou, X.; Cao, K.; Wang, L.; Pan, Z.; Wu, Q.; Li, X.; et al. Resveratrol ameliorates lipid accumulation and inflammation in human SZ95 sebocytes via the AMPK signaling pathways in vitro. J. Dermatol. Sci. 2021, 103, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K. Protective Effect of Epigallocatechin Gallate on Endothelial Disorders in Atherosclerosis. J. Cardiovasc. Pharmacol. 2020, 75, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Gao, C.; Yan, P.; Zhang, M.; Wang, Y.; Hu, Y.; Wu, X.; Wang, X.; Sheng, J. EGCG reduces obesity and white adipose tissue gain partly through AMPK activation in mice. Front. Pharmacol. 2018, 9, 1366. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Tsai, S.J.; Wang, Y.J.; Pan, M.H.; Kao, J.Y.; Way, T.D. EGCG inhibits protein synthesis, lipogenesis, and cell cycle progression through activation of AMPK in p53 positive and negative human hepatoma cells. Mol. Nutr. Food Res. 2009, 53, 1156–1165. [Google Scholar] [CrossRef]
- Song, H.; Yang, R.; Zhang, J.; Sun, P.; Xing, X.; Wang, L.; Sairijima, T.; Hu, Y.; Liu, Y.; Cheng, H.; et al. Oleic acid-induced steatosis model establishment in LMH cells and its effect on lipid metabolism. Poult. Sci. 2023, 102, 102297. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Kim, T.H.; Hong, M.W.; Park, T.S.; Lee, H.; Lee, S.J. Transcriptomic alterations induced by aflatoxin B1 and ochratoxin a in LMH cell line. Poult. Sci. 2020, 99, 5265–5274. [Google Scholar] [CrossRef]
- Adan, A.; Kiraz, Y.; Baran, Y. Cell proliferation and cytotoxicity assays. Curr. Pharm. Biotechnol. 2016, 17, 1213–1221. [Google Scholar] [CrossRef]
- Chang, H.T.; Chou, C.T.; Chen, I.L.; Liang, W.Z.; Kuo, D.H.; Huang, J.K.; Shieh, P.; Jan, C. Mechanisms of resveratrol-induced changes in [Ca(2+)]i and cell viability in PC3 human prostate cancer cells. J. Recept. Signal Transduct. Res. 2013, 33, 298–303. [Google Scholar] [CrossRef]
- Yang, H.Z.; Zhang, J.; Zeng, J.; Liu, S.; Zhou, F.; Zhang, F.; Giampieri, F.; Cianciosi, D.; Forbes-Hernandez, T.Y.; Ansary, J.; et al. Resveratrol inhibits the proliferation of melanoma cells by modulating cell cycle. Int. J. Food Sci. Nutr. 2019, 71, 84–93. [Google Scholar] [CrossRef]
- Yu, P.W.; Fu, P.F.; Zeng, L.; Qi, Y.L.; Li, X.Q.; Wang, Q.; Yang, G.Y.; Li, H.W.; Wang, J.; Chu, B.B.; et al. EGCG restricts PRRSV proliferation by disturbing lipid metabolism. Microbiol. Spectr. 2022, 10, e227621. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Della-Fera, M.A.; Baile, C.A. Green tea polyphenol epigallocatechin gallate inhibits adipogenesis and induces apoptosis in 3T3-L1 adipocytes. Obes. Res. 2005, 13, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.N.; Shen, S.R.; Yin, J.J. Effects of interactions of EGCG and Cd(2+) on the growth of PC-3 cells and their mechanisms. Food Chem. Toxicol. 2007, 45, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P. Hormesis defined. Ageing Res. Rev. 2008, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kasper, P.; Martin, A.; Lang, S.; Kütting, F.; Goeser, T.; Demir, M.; Steffen, H.M. NAFLD and cardiovascular diseases: A clinical review. Clin. Res. Cardiol. 2021, 110, 921–937. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.H.; Gong, J.G.; Zhao, G.X.; Lin, X.; Liu, Y.C.; Ma, K.W. Effects of dietary supplementation of resveratrol on performance, egg quality, yolk cholesterol and antioxidant enzyme activity of laying hens. Br. Poult. Sci. 2017, 58, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, X.; Ren, B.; Zhao, Q.; Zhang, F. The effect of resveratrol on blood glucose and blood lipids in rats with gestational diabetes mellitus. Evid.-Based Complement. Altern. Med. 2021, 2021, 2956795. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Cao, Y.; Yuan, J. Effects of (+)-catechin and epigallocatechin gallate on alcoholic fatty liver in mice models. Zhonghua Yu Fang Yi Xue Za Zhi 2021, 55, 1305–1310. [Google Scholar] [CrossRef]
- Li, Y.; Wu, S. Epigallocatechin gallate suppresses hepatic cholesterol synthesis by targeting SREBP-2 through SIRT1/FOXO1 signaling pathway. Mol. Cell. Biochem. 2018, 448, 175–185. [Google Scholar] [CrossRef]
- Sun, X.; Li, X.; Jia, H.; Wang, H.; Shui, G.; Qin, Y.; Shu, X.; Wang, Y.; Dong, J.; Liu, G.; et al. Nuclear factor E2-Related factor 2 mediates oxidative Stress-Induced lipid accumulation in adipocytes by increasing adipogenesis and decreasing lipolysis. Antioxid. Redox Signal. 2020, 32, 173–192. [Google Scholar] [CrossRef]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free. Radic. Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef] [PubMed]
- Magalingam, K.B.; Radhakrishnan, A.; Haleagrahara, N. Rutin, a bioflavonoid antioxidant protects rat pheochromocytoma (PC-12) cells against 6-hydroxydopamine (6-OHDA)-induced neurotoxicity. Int. J. Mol. Med. 2013, 32, 235–240. [Google Scholar] [CrossRef]
- Erdogan, E.; Ilgaz, Y.; Gurgor, P.N.; Oztas, Y.; Topal, T.; Oztas, E. Rutin ameliorates methotrexate induced hepatic injury in rats. ACTA Cir. Bras. 2015, 30, 778–784. [Google Scholar] [CrossRef]
- Wu, H.; Xue, J.; Di, H.; Lv, C.; Hao, Y.; Nie, Z. Resveratrol improves ovarian function in aged rat by inhibiting oxidative stress and activating the Sirt1. Gen. Physiol. Biophys. 2022, 41, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Karamese, M.; Guvendi, B.; Karamese, S.A.; Cinar, I.; Can, S.; Erol, H.S.; Aydin, H.; Gelen, V.; Karakus, E. The protective effects of epigallocatechin gallate on lipopolysa ccharide-induced hepatotoxicity: An in vitro study on Hep3B cells. Iran. J. Basic Med. Sci. 2016, 19, 483–489. [Google Scholar] [PubMed] [PubMed Central]
- Li, Y.; Zhao, S.; Zhang, W.; Zhao, P.; He, B.; Wu, N.; Han, P. Epigallocatechin-3-O-gallate (EGCG) attenuates FFAs-induced peripheral insulin resistance through AMPK pathway and insulin signaling pathway in vivo. Diabetes Res. Clin. Pract. 2011, 93, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Pokhrel, R.H.; Acharya, S.; Ahn, J.H.; Gu, Y.; Pandit, M.; Kim, J.O.; Park, Y.Y.; Kang, B.; Ko, H.J.; Chang, J.H. AMPK promotes antitumor immunity by downregulating PD-1 in regulatory T cells via the HMGCR/p38 signaling pathway. Mol. Cancer 2021, 20, 133. [Google Scholar] [CrossRef]
- Liu, S.; Jing, F.; Yu, C.; Gao, L.; Qin, Y.; Zhao, J. AICAR-Induced activation of AMPK inhibits TSH/SREBP-2/HMGCR pathway in liver. PLoS ONE 2015, 10, e124951. [Google Scholar] [CrossRef]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; van de Weijer, T.; Goossens, G.H.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S.; et al. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab. 2011, 14, 612–622. [Google Scholar] [CrossRef]
- Yang, C.S.; Zhang, J.; Zhang, L.; Huang, J.; Wang, Y. Mechanisms of body weight reduction and metabolic syndrome alleviation by tea. Mol. Nutr. Food Res. 2016, 60, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Baselga-Escudero, L.; Blade, C.; Ribas-Latre, A.; Casanova, E.; Suarez, M.; Torres, J.L.; Salvado, M.J.; Arola, L.; Arola-Arnal, A. Resveratrol and EGCG bind directly and distinctively to miR-33a and miR-122 and modulate divergently their levels in hepatic cells. Nucleic Acids Res. 2014, 42, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Yu, R.; Liu, S.; Huwatibieke, B.; Li, Z.; Zhang, W. Irisin Inhibits Hepatic Cholesterol Synthesis via AMPK-SREBP2 Signaling. EBioMedicine 2016, 6, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Cui, L.; Chen, Z.; Zhang, L. Gene expression profiling of DMU-212-induced apoptosis and anti-angiogenesis in vascular endothelial cells. Pharm. Biol. 2016, 54, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Liu, J.; Zhao, W.; Wang, Y.; He, Q.; Wu, R.; Li, D.; Xu, J. Mechanistic studies for tri-targeted inhibition of enzymes involved in cholesterol biosynthesis by green tea polyphenols. Org. Biomol. Chem. 2014, 12, 4941–4951. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Pan, J.; Qu, N.; Lei, Y.; Han, J.; Zhang, J.; Han, D. The AMPK pathway in fatty liver disease. Front. Physiol. 2022, 13, 970292. [Google Scholar] [CrossRef]
- Sun, W.; Yu, S.; Han, H.; Yuan, Q.; Chen, J.; Xu, G. Resveratrol Inhibits Human Visceral Preadipocyte Proliferation and Differentiation in vitro. Lipids 2019, 54, 679–686. [Google Scholar] [CrossRef]
- Gracia, A.; Miranda, J.; Fernandez-Quintela, A.; Eseberri, I.; Garcia-Lacarte, M.; Milagro, F.I.; Martínez, A.; Aguirre, L.; Portillo, M.P. Involvement of miR-539-5p in the inhibition of de novo lipogenesis induced by resveratrol in white adipose tissue. Food Funct. 2016, 7, 1680–1688. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Primer Sequence (5′-3′) | Product Size (bp) |
|---|---|---|---|
| AMPK | 427185 | F: CAAGCCCGCCAGATTCTTTT | 184 |
| R: CGACTCTGACTCCGTATCCC | |||
| HMGCR | 395145 | F: AGGAGCTTGCTGTGAAAACG | 254 |
| R: AAGCTTTCACTTCTGCAGCC | |||
| SREBP2 | 395304 | F: AAGAGAAGATCCCACCCAGC | 259 |
| R: TTCCTCTGGGTGCAGTACAG | |||
| FDPS | 425061 | F: GGTTGGTGCATCGAGTTGTT | 181 |
| R: GCCTGCAGTACTTCTTCAGC | |||
| FDFT1 | 422038 | F: GCCTTTCCCGACTCTTCTCT | 152 |
| R: GGGCCAGAACTCTCTTCCTT | |||
| MVK | 768555 | F: TCGGGTGTGGATAATGCTGT | 159 |
| R: CTTCTCCTTAACCCCAGCCA | |||
| SQLE | 420335 | F: TTGTGGGTTCAGGTGTCCTT | 234 |
| R: CCTCCGACTTGCTCTCTAGG | |||
| LSS | 424037 | F: CCAAGCAGATGACAGATGGC | 280 |
| R: CAGATGGGGAAGACATTGCG | |||
| ACC | 396504 | F: AAACTGATGGGGACGTGGAT | 261 |
| R: ATGGAATGGCAGTGAGGTCA | |||
| SCD1 | 395706 | F: TTGCAAACTCCATGGCCTTC | 193 |
| R: TCACTCAGGTCCAGCTTCTG | |||
| FASN | 396061 | F: AGCTGGACTACATTGCCACT | 238 |
| R: GTGTGCAGCAAAACAAACCC | |||
| SREBP1 | 373915 | F: GACCTCCAGCATCACCTCTT | 290 |
| R: CCGACTTGTTGAGCTTAGCC | |||
| β-actin | 396526 | F: AGTACCCCATTGAACACGGT | 197 |
| R: ATACATGGCTGGGGTGTTGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; An, Y.; Rajput, S.A.; Qi, D. Resveratrol and (-)-Epigallocatechin-3-gallate Regulate Lipid Metabolism by Activating the AMPK Pathway in Hepatocytes. Biology 2024, 13, 368. https://doi.org/10.3390/biology13060368
Wang H, An Y, Rajput SA, Qi D. Resveratrol and (-)-Epigallocatechin-3-gallate Regulate Lipid Metabolism by Activating the AMPK Pathway in Hepatocytes. Biology. 2024; 13(6):368. https://doi.org/10.3390/biology13060368
Chicago/Turabian StyleWang, Huanbin, Yu An, Shahid Ali Rajput, and Desheng Qi. 2024. "Resveratrol and (-)-Epigallocatechin-3-gallate Regulate Lipid Metabolism by Activating the AMPK Pathway in Hepatocytes" Biology 13, no. 6: 368. https://doi.org/10.3390/biology13060368
APA StyleWang, H., An, Y., Rajput, S. A., & Qi, D. (2024). Resveratrol and (-)-Epigallocatechin-3-gallate Regulate Lipid Metabolism by Activating the AMPK Pathway in Hepatocytes. Biology, 13(6), 368. https://doi.org/10.3390/biology13060368