
Reducing Epistasis and Pleiotropy Can Avoid the Survival of the Flattest Tragedy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Indirect Encoding
2.2. Epistasis and Pleiotropy
2.3. Two-Peak Model
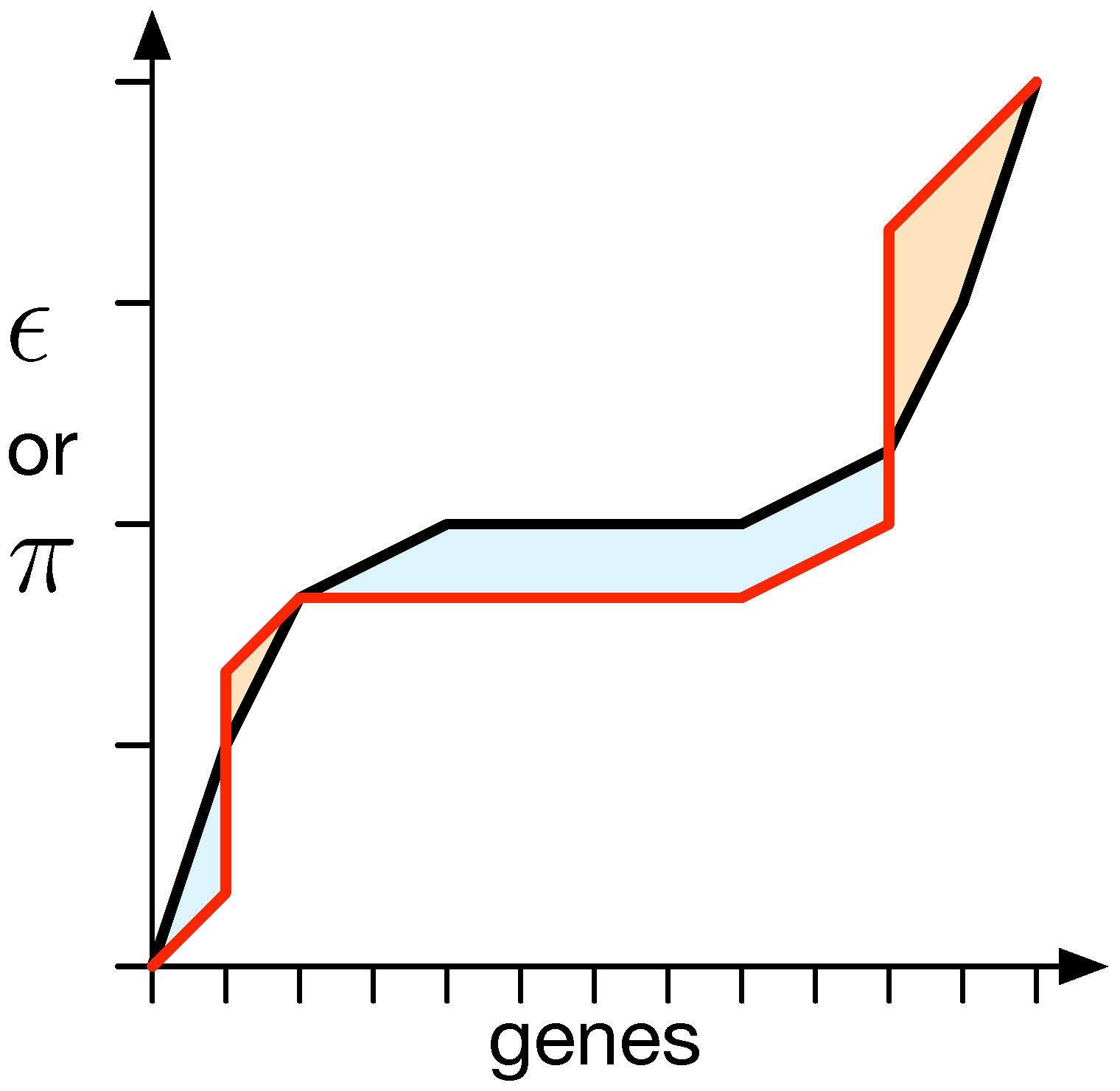
2.4. Changing Epistasis and Pleiotropy while Keeping the Phenotype Identical
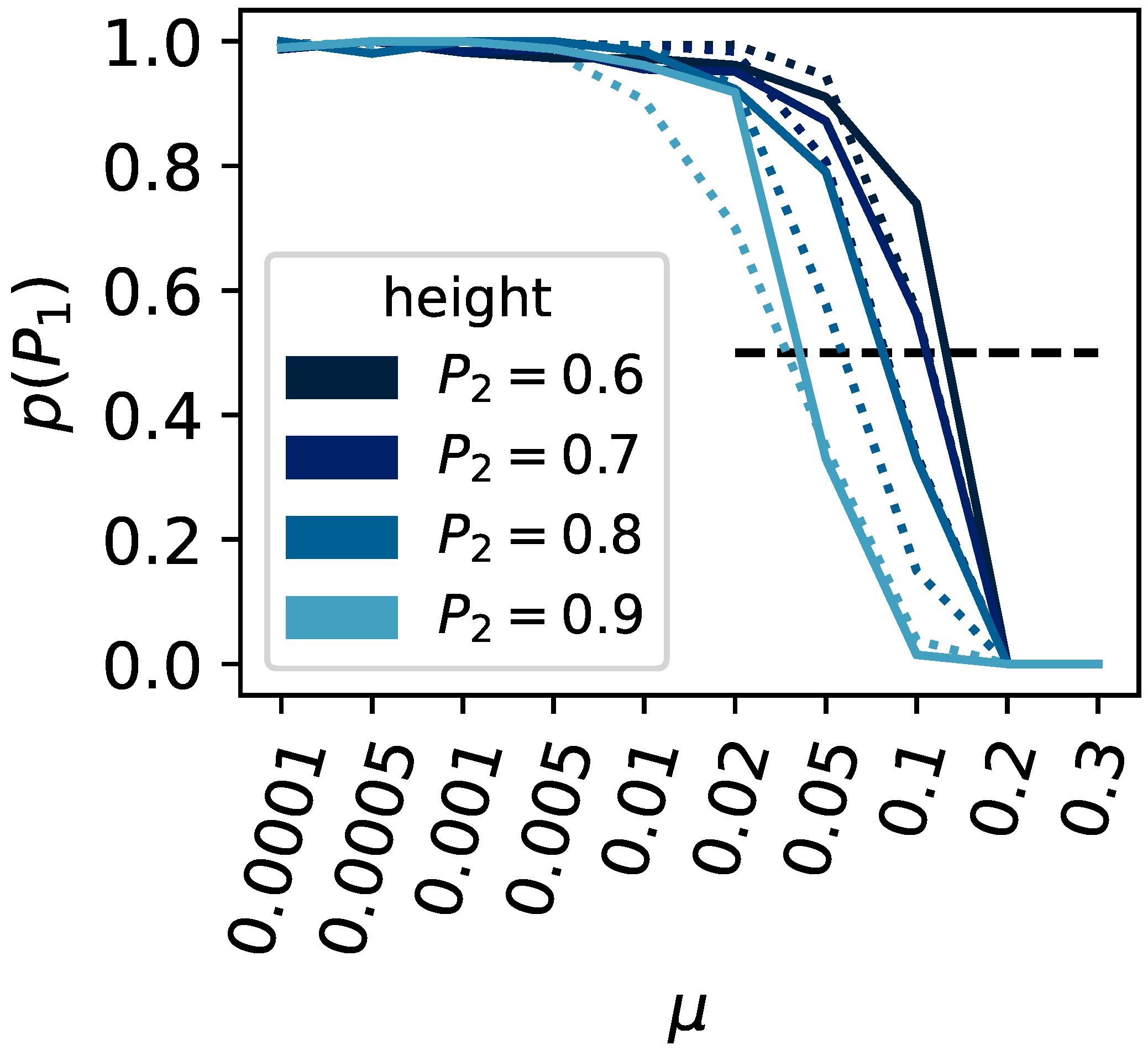
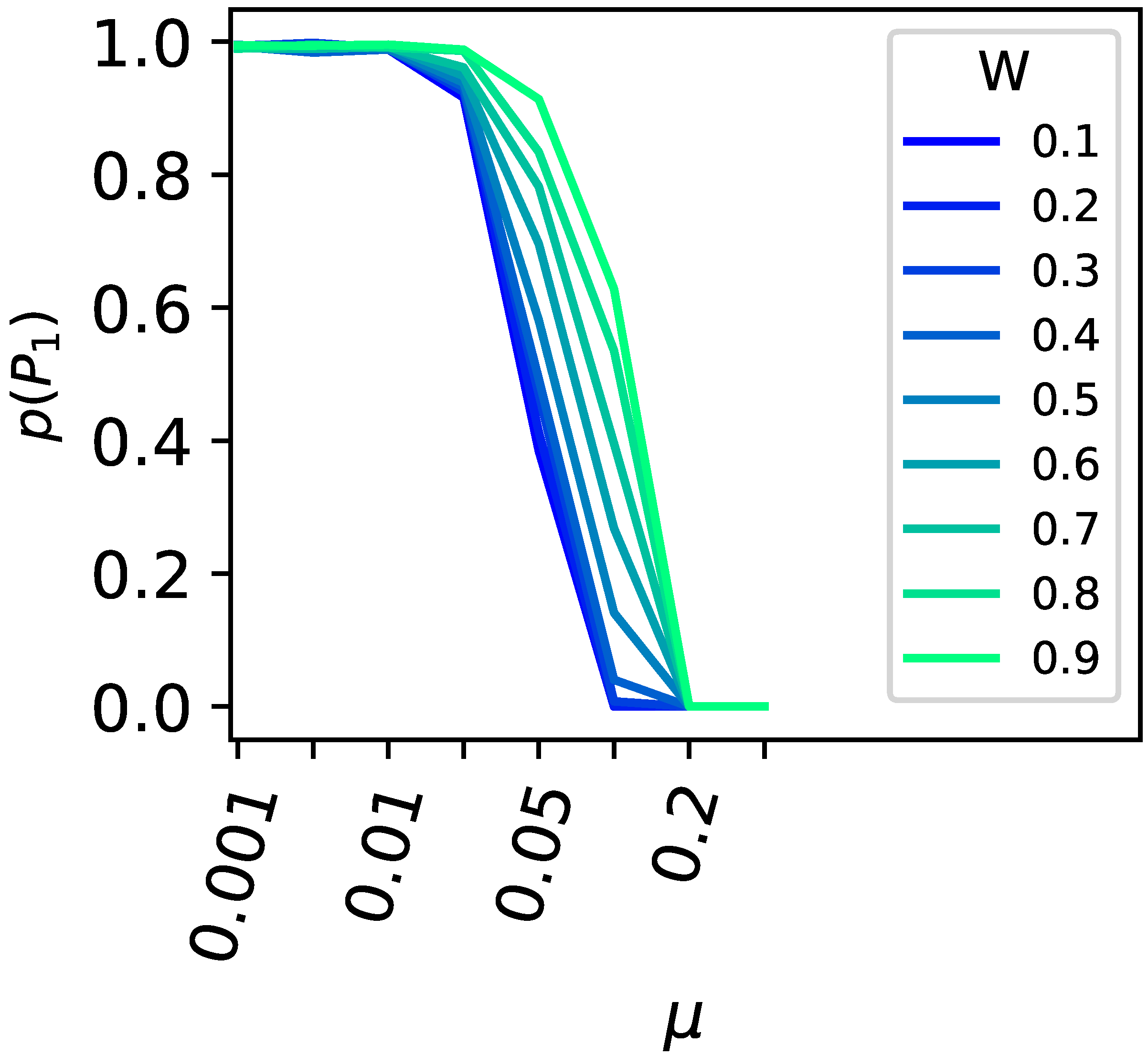
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SoF | Survival of the Flattest |
| Epistasis | |
| Pleiotropy |
References
- Smith, J.M.; Szathmary, E. The Major Transitions in Evolution; OUP Oxford: Oxford, UK, 1997. [Google Scholar]
- Adami, C. Information theory in molecular biology. Phys. Life Rev. 2004, 1, 3–22. [Google Scholar] [CrossRef]
- Thompson, E.G.; Galitski, T. Quantifying and analyzing the network basis of genetic complexity. PLoS Comput. Biol. 2012, 8, e1002583. [Google Scholar] [CrossRef]
- Hwang, S.; Park, S.C.; Krug, J. Genotypic complexity of Fisher’s geometric model. Genetics 2017, 206, 1049–1079. [Google Scholar] [CrossRef]
- Lehre, P.K.; Haddow, P.C. Developmental mappings and phenotypic complexity. In Proceedings of the The 2003 Congress on Evolutionary Computation, CEC’03, Canberra, Australia, 8–12 December 2003; Volume 1, pp. 62–68. [Google Scholar]
- Ryan Gregory, T. Genome size and developmental complexity. Genetica 2002, 115, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Alberch, P. From genes to phenotype: Dynamical systems and evolvability. Genetica 1991, 84, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Fortuna, M.A.; Zaman, L.; Ofria, C.; Wagner, A. The genotype-phenotype map of an evolving digital organism. PLoS Comput. Biol. 2017, 13, e1005414. [Google Scholar] [CrossRef] [PubMed]
- Manrubia, S.; Cuesta, J.A.; Aguirre, J.; Ahnert, S.E.; Altenberg, L.; Cano, A.V.; Catalán, P.; Diaz-Uriarte, R.; Elena, S.F.; García-Martín, J.A.; et al. From genotypes to organisms: State-of-the-art and perspectives of a cornerstone in evolutionary dynamics. Phys. Life Rev. 2021, 38, 55–106. [Google Scholar] [CrossRef]
- Adami, C.; Ofria, C.; Collier, T.C. Evolution of biological complexity. Proc. Natl. Acad. Sci. USA 2000, 97, 4463–4468. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M. Complexity: A Guided Tour; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Gould, S.J.; Eldredge, N. Punctuated equilibria: An alternative to phyletic gradualism. Model. Paleobiol. 1972, 1972, 82–115. [Google Scholar]
- Gould, S.J. The Structure of Evolutionary Theory; Harvard University Press: Cambridge, MA, UAS, 2002. [Google Scholar]
- LaBar, T.; Adami, C. Different evolutionary paths to complexity for small and large populations of digital organisms. PLoS Comput. Biol. 2016, 12, e1005066. [Google Scholar] [CrossRef]
- Hagolani, P.F.; Zimm, R.; Vroomans, R.; Salazar-Ciudad, I. On the evolution and development of morphological complexity: A view from gene regulatory networks. PLoS Comput. Biol. 2021, 17, e1008570. [Google Scholar]
- Behe, M.J. Experimental evolution, loss-of-function mutations, and “the first rule of adaptive evolution”. Q. Rev. Biol. 2010, 85, 419–445. [Google Scholar] [CrossRef] [PubMed]
- Bomblies, K.; Peichel, C.L. Gregor Johann Mendel and Modern Evolutionary Biology: Genetics of adaptation. Proc. Natl. Acad. Sci. USA 2022, 119, e2122152119. [Google Scholar] [CrossRef] [PubMed]
- Mackay, T.F. Epistasis and quantitative traits: Using model organisms to study gene–gene interactions. Nat. Rev. Genet. 2014, 15, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, M.; Baryshnikova, A.; Bellay, J.; Kim, Y.; Spear, E.D.; Sevier, C.S.; Ding, H.; Koh, J.L.; Toufighi, K.; Mostafavi, S.; et al. The genetic landscape of a cell. Science 2010, 327, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.C.; Kishony, R. Understanding, predicting and manipulating the genotypic evolution of antibiotic resistance. Nat. Rev. Genet. 2013, 14, 243–248. [Google Scholar] [CrossRef]
- Kaelin, W.G., Jr. The concept of synthetic lethality in the context of anticancer therapy. Nat. Rev. Cancer 2005, 5, 689–698. [Google Scholar] [CrossRef]
- Mackay, T.F.; Stone, E.A.; Ayroles, J.F. The genetics of quantitative traits: Challenges and prospects. Nat. Rev. Genet. 2009, 10, 565–577. [Google Scholar] [CrossRef]
- Whitlock, M.C. Variance-induced peak shifts. Evolution 1995, 49, 252–259. [Google Scholar] [CrossRef]
- Phillips, P.C. Epistasis—the essential role of gene interactions in the structure and evolution of genetic systems. Nat. Rev. Genet. 2008, 9, 855–867. [Google Scholar] [CrossRef]
- Paaby, A.B.; Rockman, M.V. The many faces of pleiotropy. Trends Genet. 2013, 29, 66–73. [Google Scholar] [CrossRef]
- Sivakumaran, S.; Agakov, F.; Theodoratou, E.; Prendergast, J.G.; Zgaga, L.; Manolio, T.; Rudan, I.; McKeigue, P.; Wilson, J.F.; Campbell, H. Abundant pleiotropy in human complex diseases and traits. Am. J. Hum. Genet. 2011, 89, 607–618. [Google Scholar] [CrossRef]
- Wagner, G.P.; Zhang, J. The pleiotropic structure of the genotype–phenotype map: The evolvability of complex organisms. Nat. Rev. Genet. 2011, 12, 204–213. [Google Scholar] [CrossRef]
- Kitano, H. Biological robustness. Nat. Rev. Genet. 2004, 5, 826–837. [Google Scholar] [CrossRef]
- Palumbi, S.R. Humans as the world’s greatest evolutionary force. Science 2001, 293, 1786–1790. [Google Scholar] [CrossRef] [PubMed]
- Stearns, S.C. The evolutionary significance of phenotypic plasticity. Bioscience 1989, 39, 436–445. [Google Scholar] [CrossRef]
- Hodgkin, J. Seven types of pleiotropy. Int. J. Dev. Biol. 2002, 42, 501–505. [Google Scholar]
- Wagner, G.P. Homologues, natural kinds and the evolution of modularity. Am. Zool. 1996, 36, 36–43. [Google Scholar] [CrossRef]
- Elena Fito, S.F.; Sanjuán Verdeguer, R. The effect of genetic robustness on evolvability in digital organisms. Bmc Evol. Biol. 2008, 8, 284. [Google Scholar] [CrossRef]
- Wolff, A.; Krug, J. Robustness and epistasis in mutation-selection models. Phys. Biol. 2009, 6, 036007. [Google Scholar] [CrossRef]
- Barton, N.H. How does epistasis influence the response to selection? Heredity 2017, 118, 96–109. [Google Scholar] [CrossRef]
- Johnson, M.S.; Martsul, A.; Kryazhimskiy, S.; Desai, M.M. Higher-fitness yeast genotypes are less robust to deleterious mutations. Science 2019, 366, 490–493. [Google Scholar] [CrossRef]
- Sydykova, D.K.; LaBar, T.; Adami, C.; Wilke, C.O. Moderate amounts of epistasis are not evolutionarily stable in small populations. J. Mol. Evol. 2020, 88, 435–444. [Google Scholar] [CrossRef]
- Johnson, M.S.; Reddy, G.; Desai, M.M. Epistasis and evolution: Recent advances and an outlook for prediction. BMC Biol. 2023, 21, 120. [Google Scholar] [CrossRef] [PubMed]
- Wilke, C.O.; Wang, J.L.; Ofria, C.; Lenski, R.E.; Adami, C. Evolution of digital organisms at high mutation rates leads to survival of the flattest. Nature 2001, 412, 331–333. [Google Scholar] [CrossRef]
- Adami, C. Digital genetics: Unravelling the genetic basis of evolution. Nat. Rev. Genet. 2006, 7, 109–118. [Google Scholar] [CrossRef]
- Meli, C.; Nezval, V.; Oplatkova, Z.K.; Buttigieg, V.; Staines, A.S. A study of direct and indirect encoding in phenotype-genotype relationships. In Artificial Intelligence and Soft Computing, Proceedings of the 20th International Conference, ICAISC 2021, Virtual Event, 21–23 June 2021; Proceedings, Part II 20; Springer: Berlin/Heidelberg, Germany, 2021; pp. 290–301. [Google Scholar]
- Mehra, P.; Hintze, A. An extension to the NK fitness landscape model to study pleiotropy, epistasis, and ruggedness independently. In Proceedings of the 2022 IEEE Symposium Series on Computational Intelligence (SSCI), Singapore, 4–7 December 2022; pp. 1259–1267. [Google Scholar]
- Clune, J.; Stanley, K.O.; Pennock, R.T.; Ofria, C. On the performance of indirect encoding across the continuum of regularity. IEEE Trans. Evol. Comput. 2011, 15, 346–367. [Google Scholar] [CrossRef]
- Nichele, S.; Tufte, G. Evolutionary growth of genomes for the development and replication of multicellular organisms with indirect encoding. In Proceedings of the 2014 IEEE International Conference on Evolvable Systems, Orlando, FL, USA, 9–12 December 2014; pp. 141–148. [Google Scholar]
- Cussat-Blanc, S.; Harrington, K.; Banzhaf, W. Artificial gene regulatory networks—A review. Artif. Life 2019, 24, 296–328. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Ciudad, I.; Marín-Riera, M. Adaptive dynamics under development-based genotype–phenotype maps. Nature 2013, 497, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Finnigan, G.C.; Hanson-Smith, V.; Stevens, T.H.; Thornton, J.W. Evolution of increased complexity in a molecular machine. Nature 2012, 481, 360–364. [Google Scholar] [CrossRef]
- Salazar-Ciudad, I.; Marín-Riera, M.; Brun-Usan, M. Understanding the Genotype-Phenotype Map: Contrasting Mathematical Models. In Evolutionary Systems Biology: Advances, Questions, and Opportunities; Springer: Cham, Switzerland, 2021; pp. 221–244. [Google Scholar]
- Wilke, C.O.; Adami, C. Evolution of mutational robustness. Mutat. Res. Mol. Mech. Mutagen. 2003, 522, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Channon, A.; Aston, E.; Day, C.; Belavkin, R.V.; Knight, C.G. Critical mutation rate has an exponential dependence on population size. In Advances in Artificial Life, ECAL 2011, Proceedings of the Eleventh European Conference on the Synthesis and Simulation of Living, Systems, Paris, France, 8–12 August 2011; MIT Press: Cambridge, MA, USA, 2011; pp. 117–124. [Google Scholar]
- Comas, I.; Moya, A.; González-Candelas, F. Validating viral quasispecies with digital organisms: A re-examination of the critical mutation rate. BMC Evol. Biol. 2005, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Majerus, M.E. Industrial melanism in the peppered moth, Biston betularia: An excellent teaching example of Darwinian evolution in action. Evol. Educ. Outreach 2009, 2, 63–74. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehra, P.; Hintze, A. Reducing Epistasis and Pleiotropy Can Avoid the Survival of the Flattest Tragedy. Biology 2024, 13, 193. https://doi.org/10.3390/biology13030193
Mehra P, Hintze A. Reducing Epistasis and Pleiotropy Can Avoid the Survival of the Flattest Tragedy. Biology. 2024; 13(3):193. https://doi.org/10.3390/biology13030193
Chicago/Turabian StyleMehra, Priyanka, and Arend Hintze. 2024. "Reducing Epistasis and Pleiotropy Can Avoid the Survival of the Flattest Tragedy" Biology 13, no. 3: 193. https://doi.org/10.3390/biology13030193
APA StyleMehra, P., & Hintze, A. (2024). Reducing Epistasis and Pleiotropy Can Avoid the Survival of the Flattest Tragedy. Biology, 13(3), 193. https://doi.org/10.3390/biology13030193