
Integrative Transcriptome Analysis of mRNA and miRNA in Pepper’s Response to Phytophthora capsici Infection


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Pathogen Culture and Inoculation Assay
2.3. mRNA-Seq and miRNA-Seq Analysis
2.4. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR) Analysis
3. Results
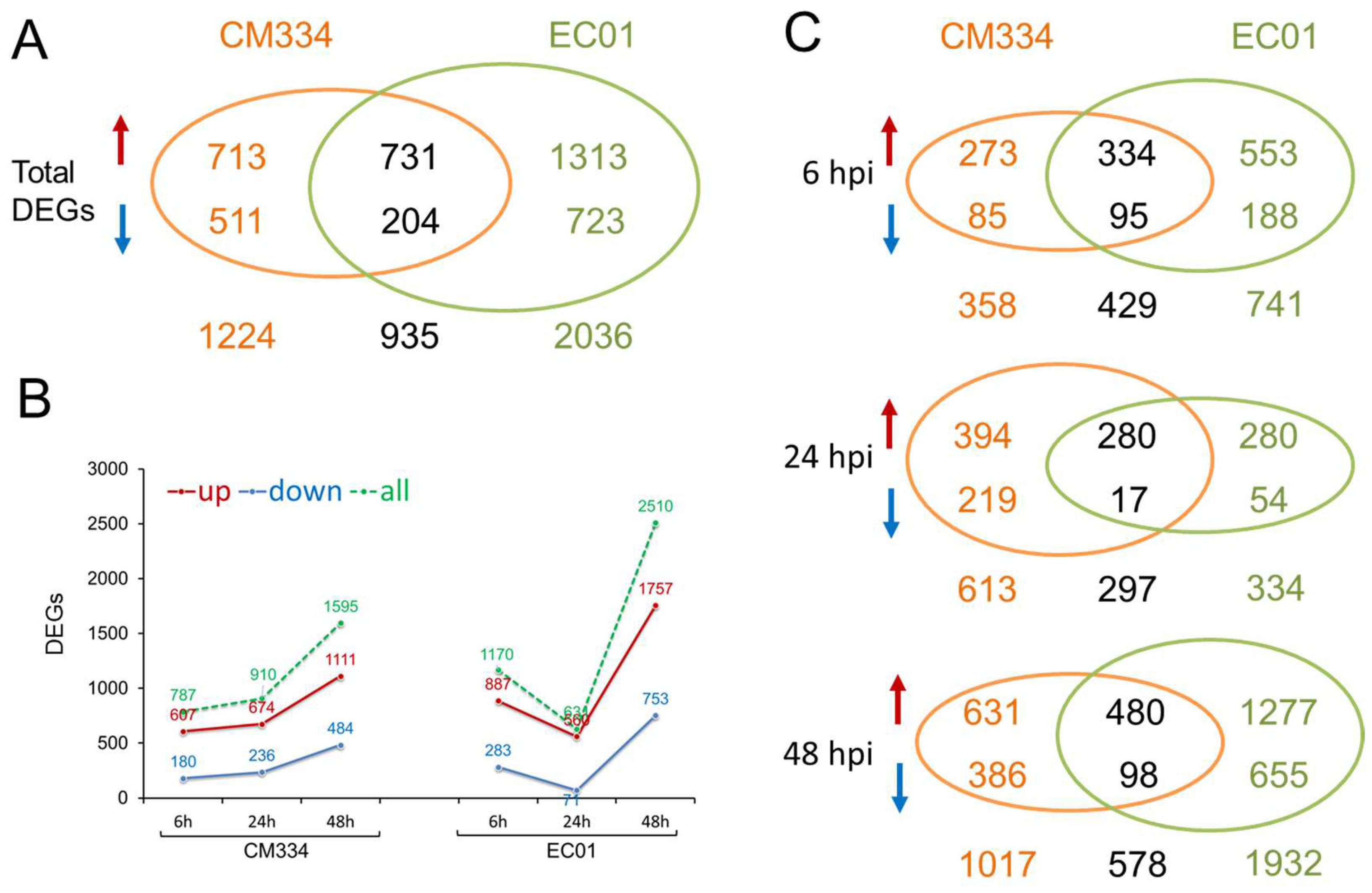
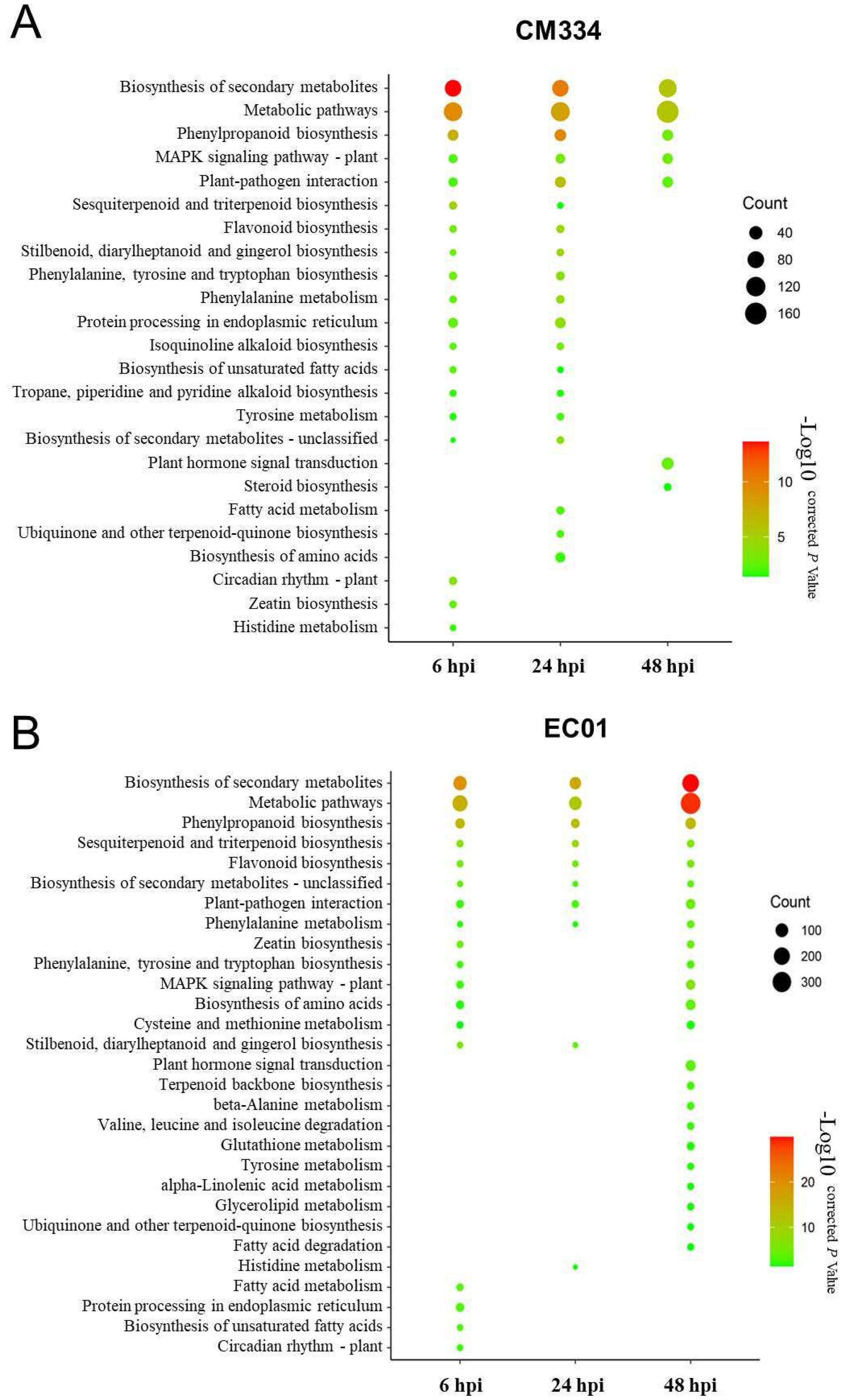
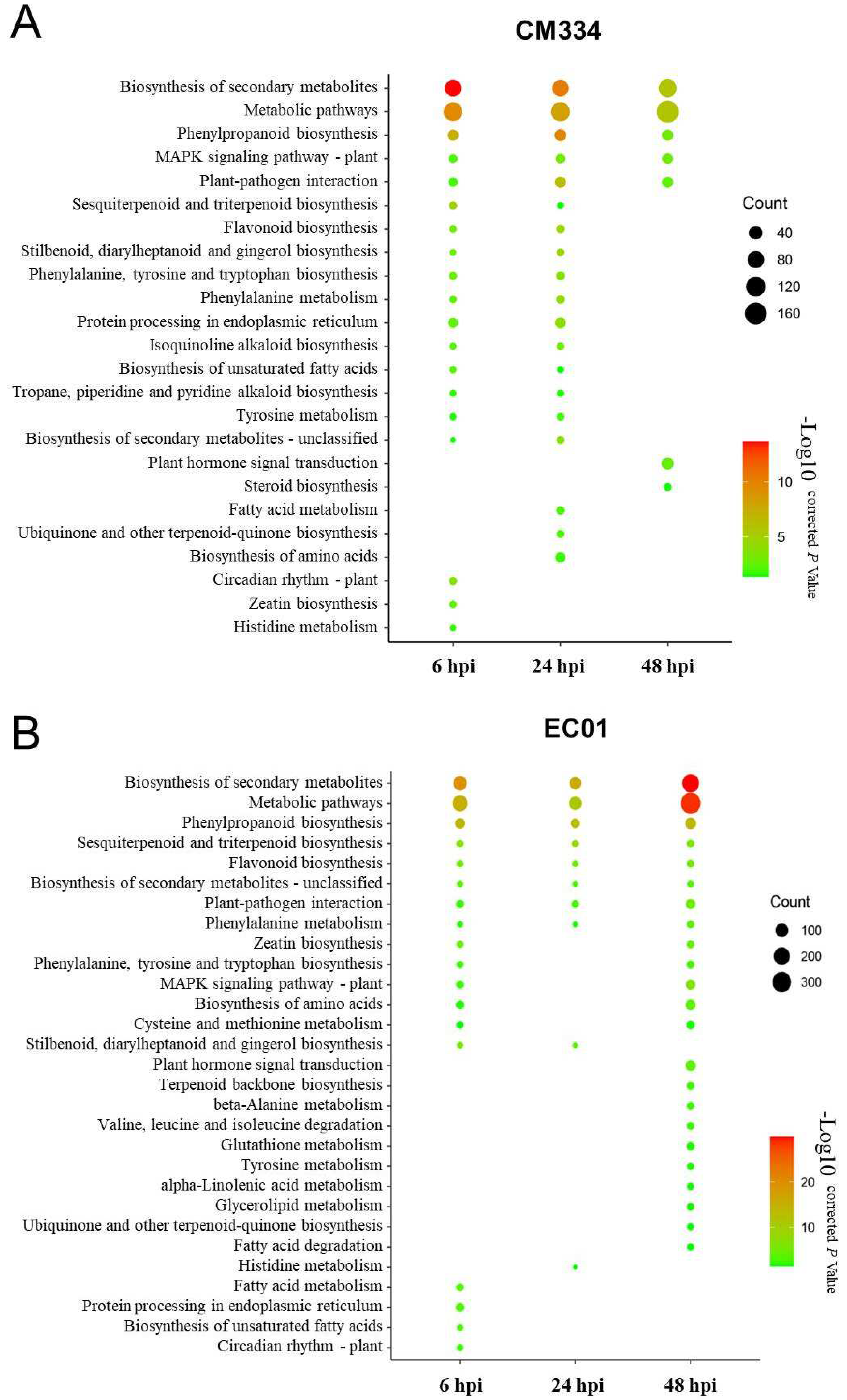
3.1. High-Throughput mRNA Sequencing and Differentially Expressed Gene (DEG) Analysis
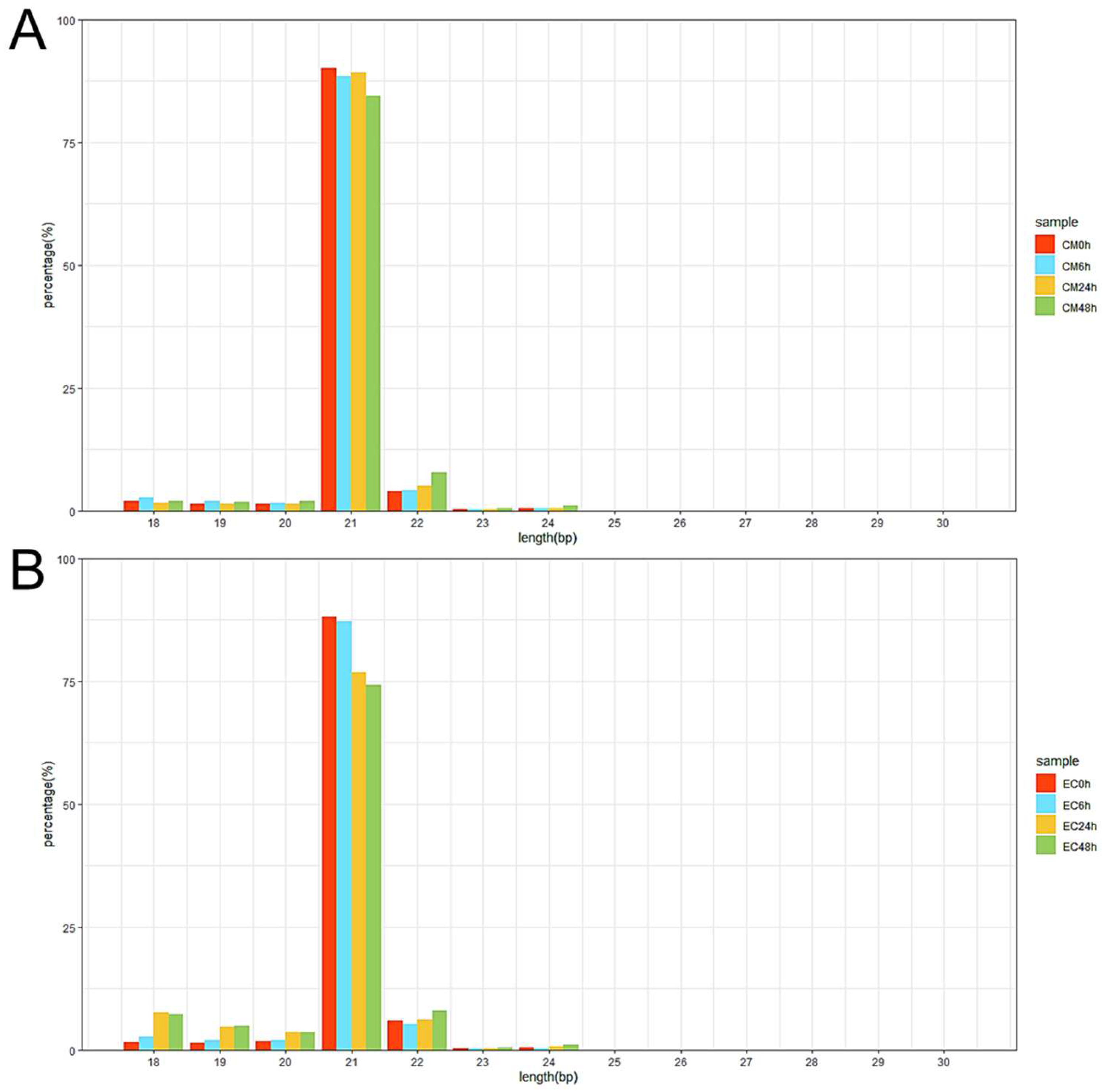
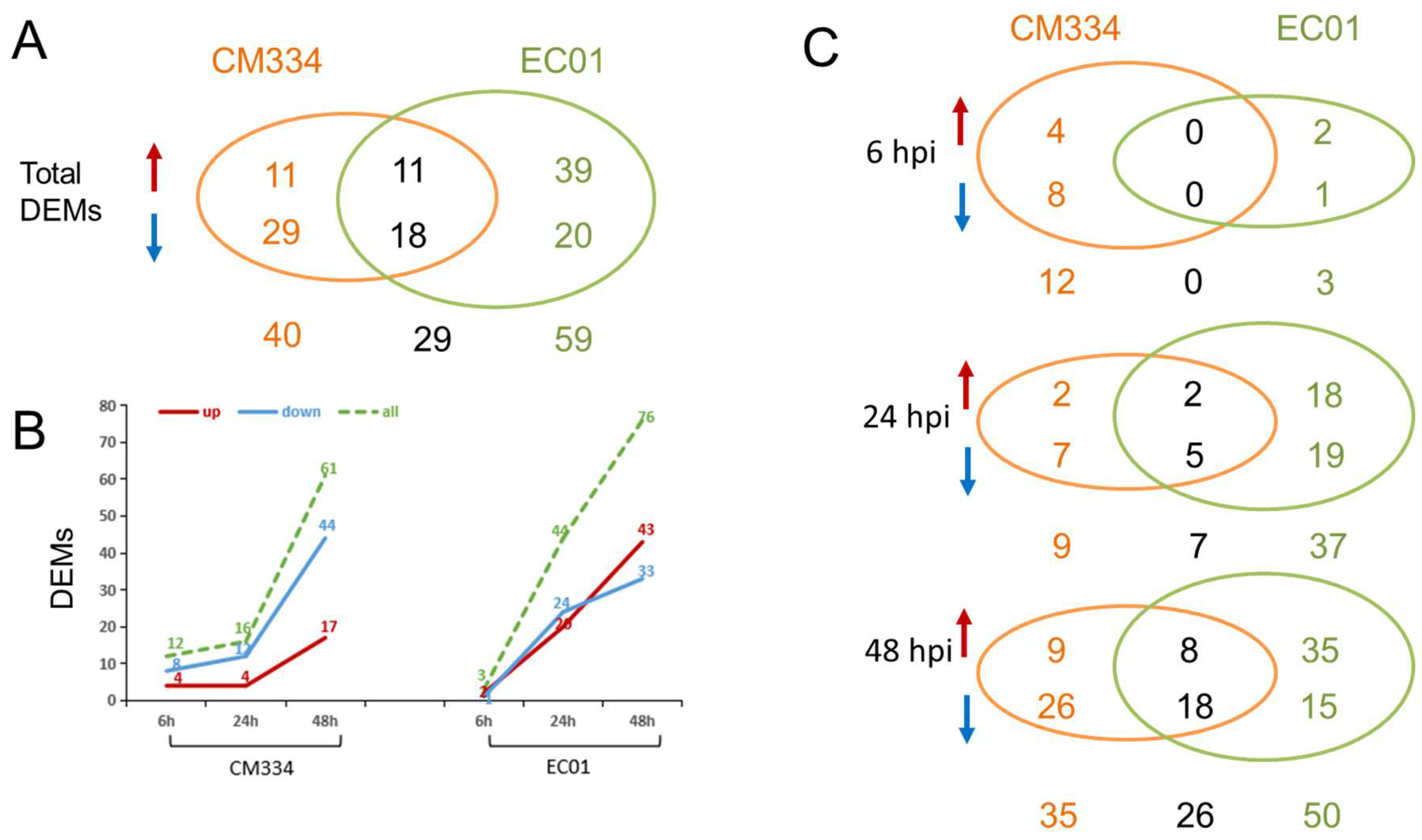
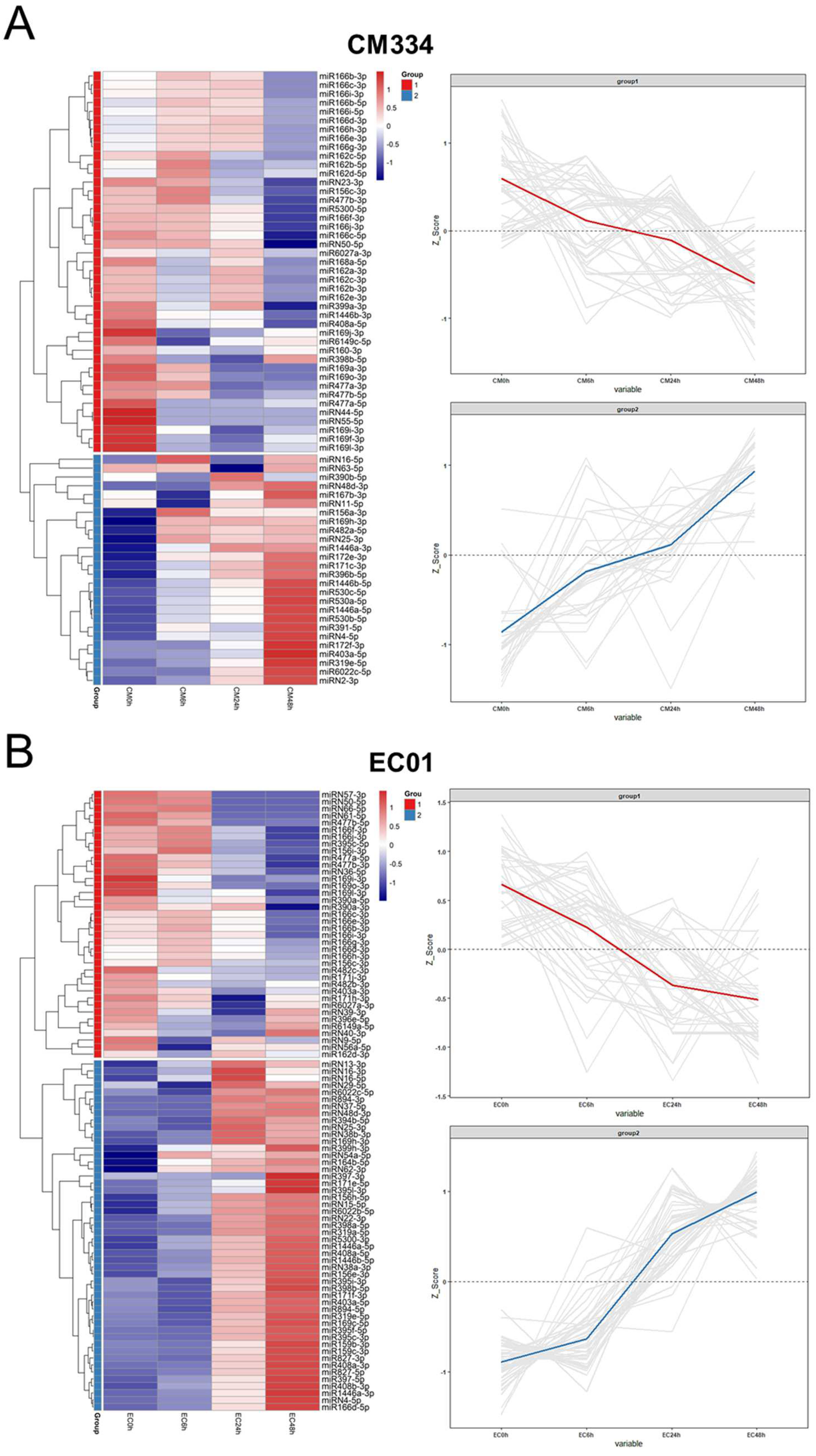
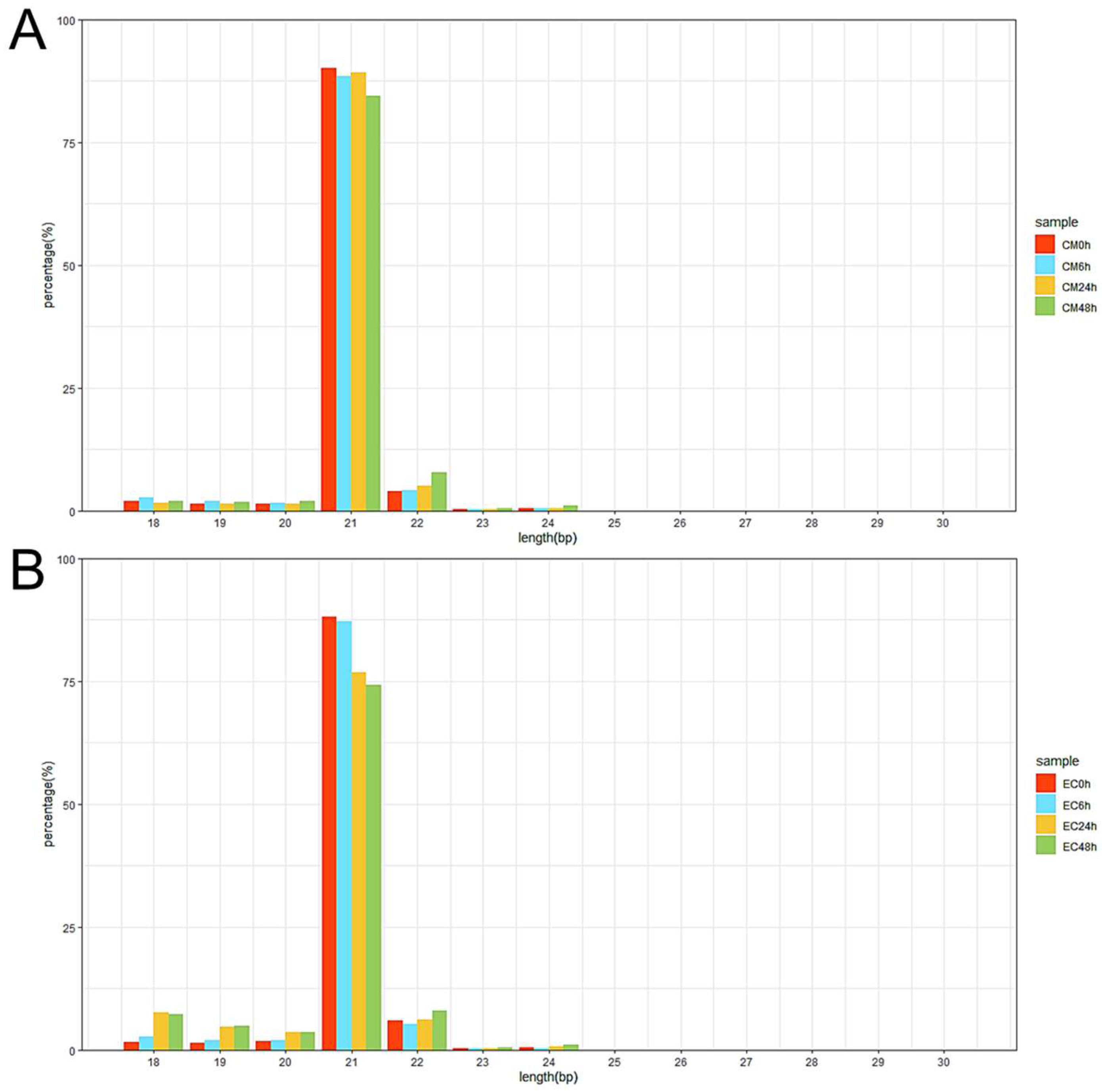
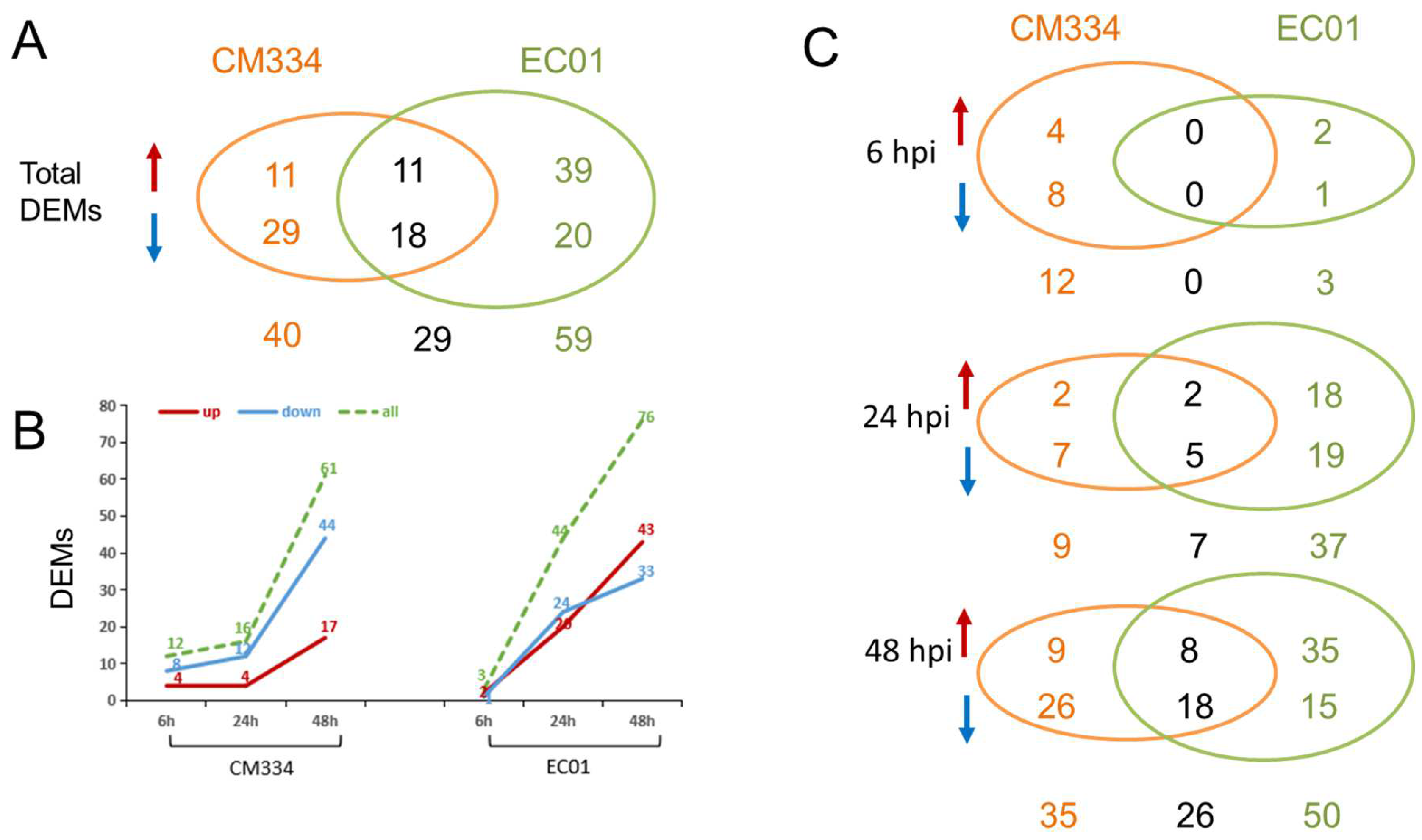
3.2. High-Throughput Small RNA Sequencing and Differentially Expressed miRNA (DEM) Analysis
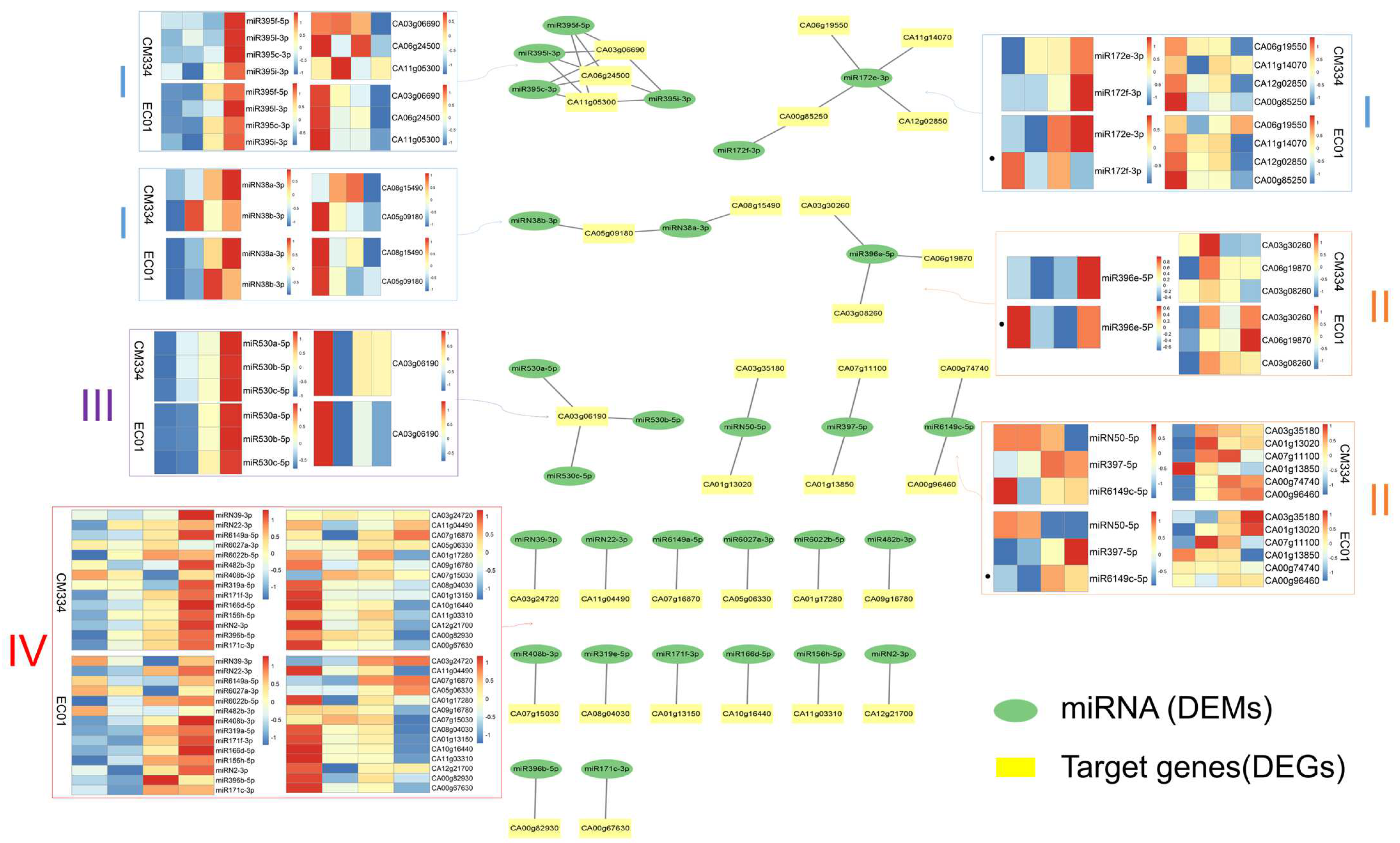
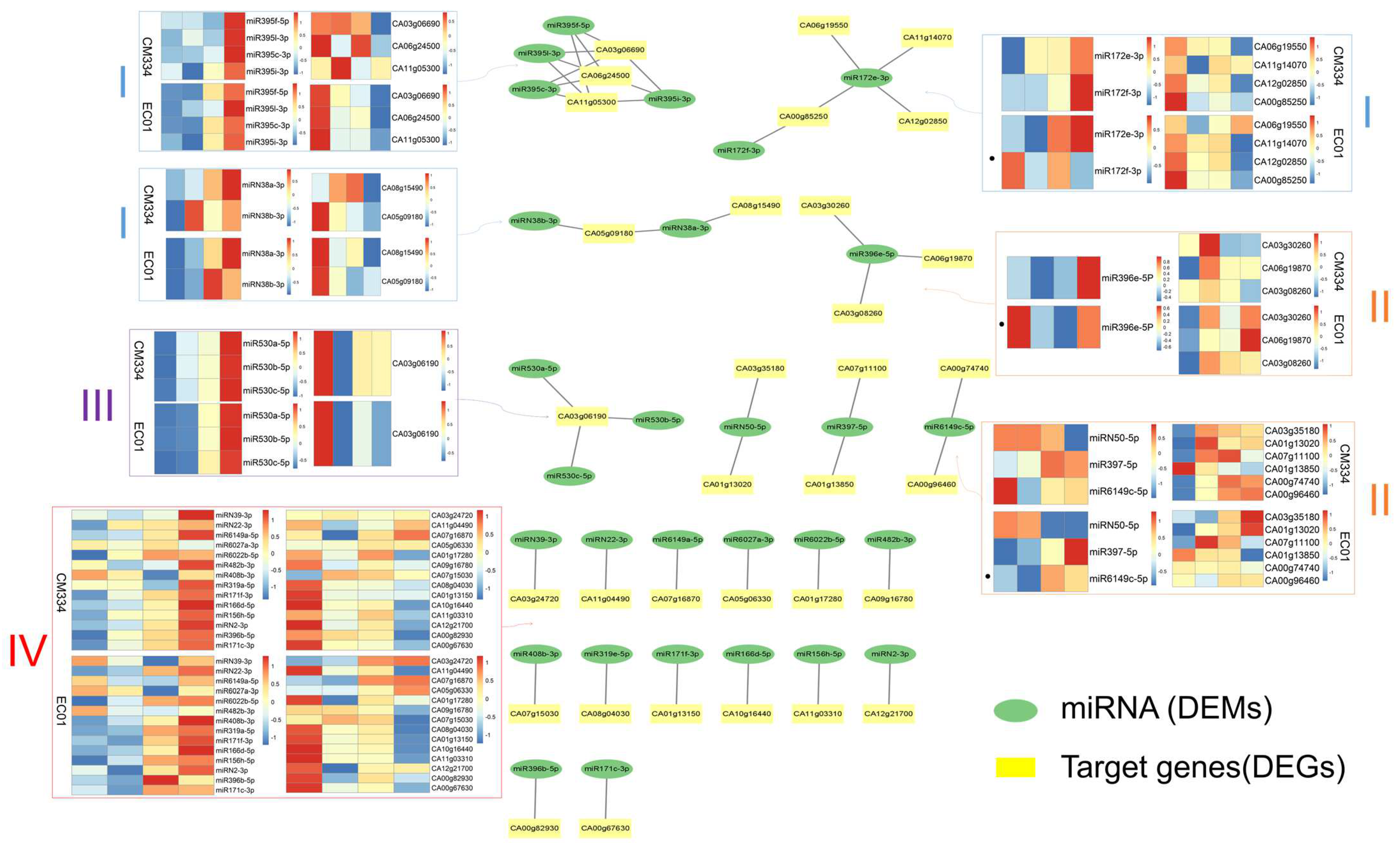
3.3. Combined Analysis of miRNA and mRNA Transcriptome
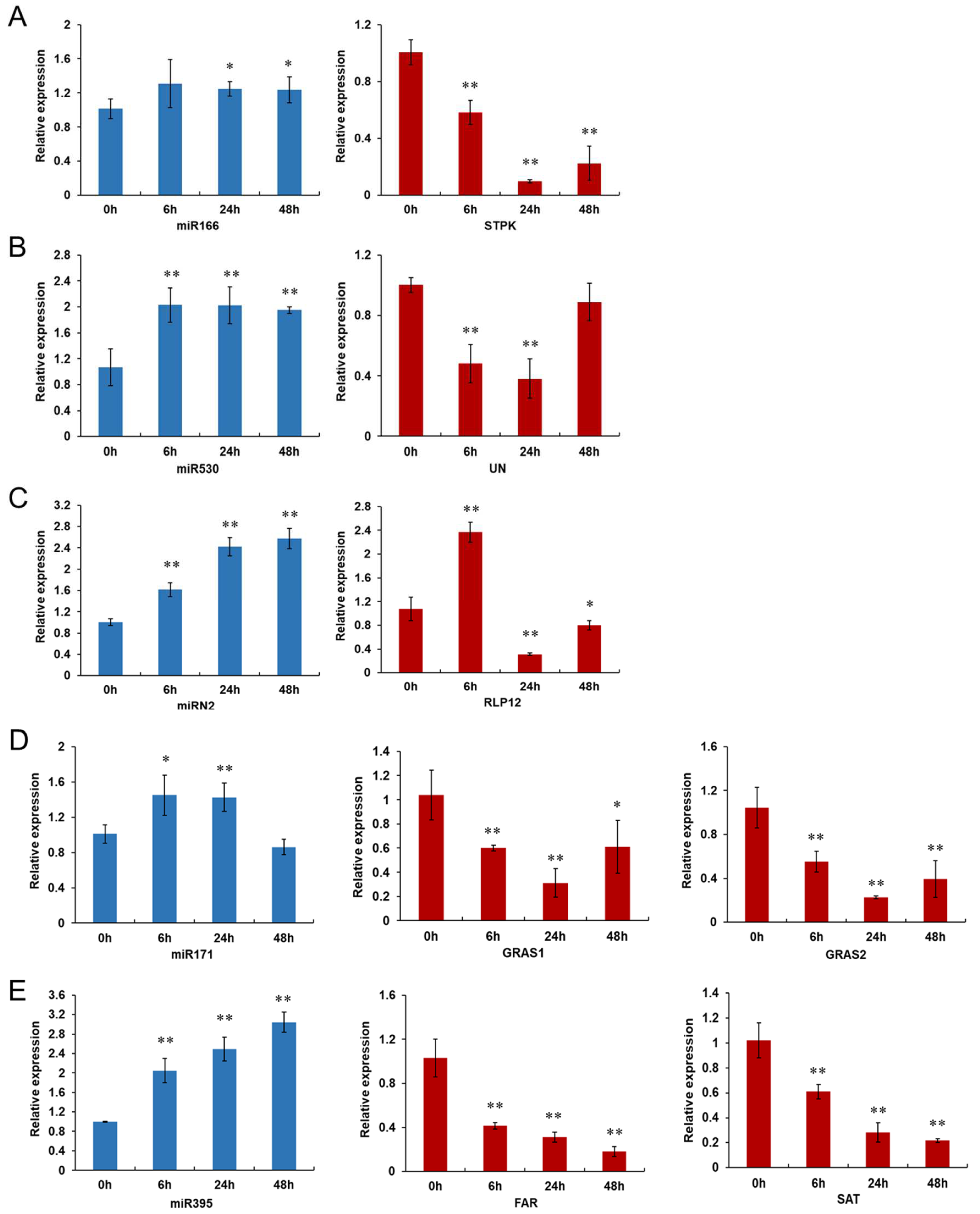
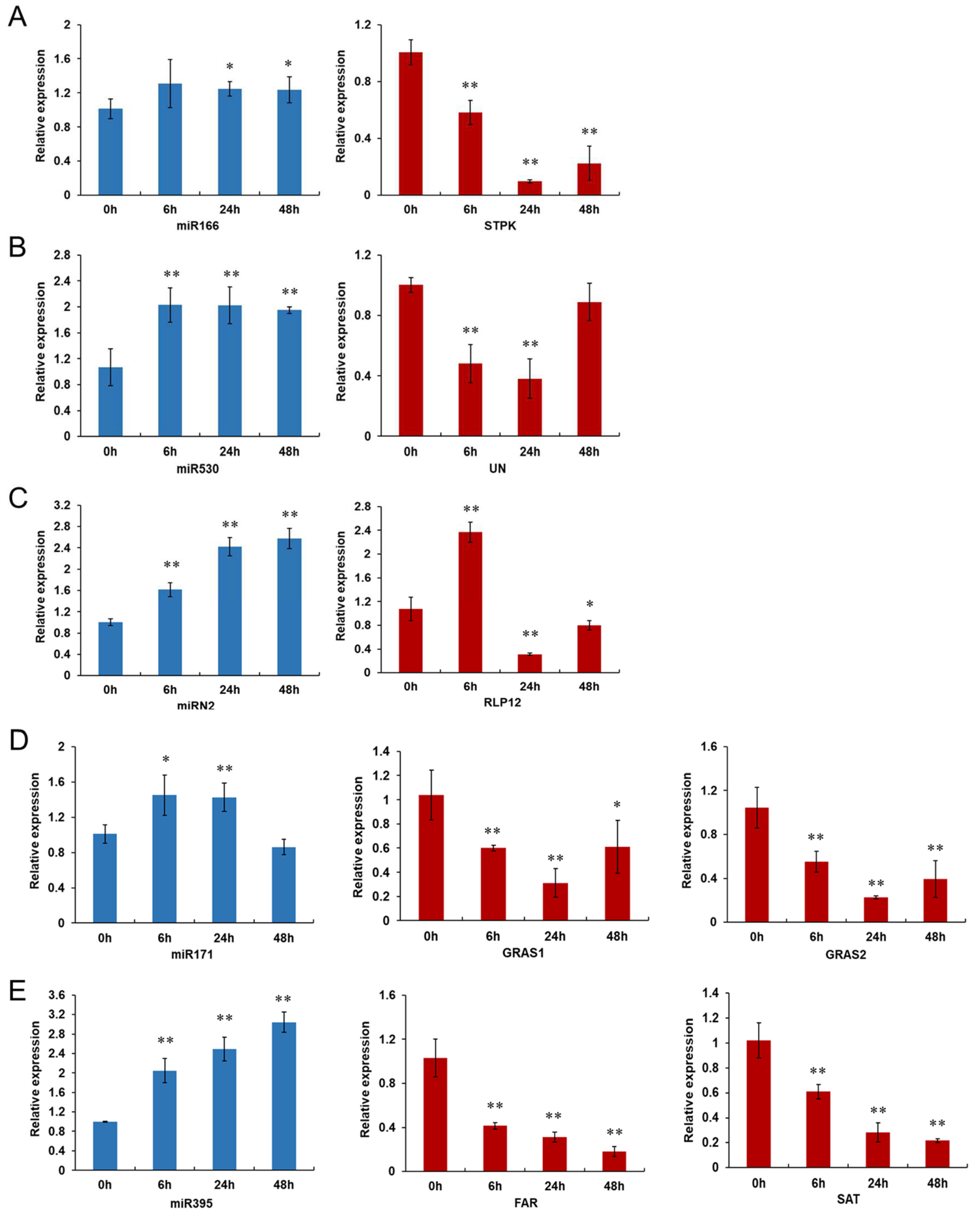
3.4. Expression Profiles of miRNAs and Their Target Genes Examined by qRT-PCR Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barchenger, D.W.; Lamour, K.H.; Bosland, P.W. Challenges and strategies for breeding resistance in Capsicum annuum to the multifarious pathogen, Phytophthora capsici. Front. Plant Sci. 2018, 9, 628. [Google Scholar] [CrossRef] [PubMed]
- Lamour, K.H.; Stam, R.; Jupe, J.; Huitema, E. The oomycete broad-host-range pathogen Phytophthora capsici. Mol. Plant Pathol. 2012, 13, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Ocampo, L.M.; Parada-Rojas, C.H.; Hansen, Z.; Vogel, G.; Smart, C.; Hausbeck, M.K.; Carmo, R.M.; Huitema, E.; Naegele, R.P.; Kousik, C.S.; et al. Phytophthora capsici: Recent progress on fundamental biology and disease management 100 years after its description. Annu. Rev. Phytopathol. 2023, 61, 185–208. [Google Scholar] [CrossRef] [PubMed]
- Tyler, B.M.; Tripathy, S.; Zhang, X.M.; Dehal, P.; Jiang, R.H.Y.; Aerts, A.; Arredondo, F.D.; Baxter, L.; Bensasson, D.; Beynon, J.L.; et al. Phytophthora genome sequences uncover evolutionary origins and mechanisms of pathogenesis. Science 2006, 313, 1261–1266. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, S.; Furzer, O.; Jones, J.D.G.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.D.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabieres, F.; et al. The Top 10 oomycete pathogens in molecular plant pathology. Mol. Plant Pathol. 2015, 16, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Khraiwesh, B.; Zhu, J.K.; Zhu, J.H. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta-Gene Regul. Mech. 2012, 1819, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Q.; Zhang, L.C.; Yang, Y.Z.; Schmid, M.; Wang, Y.W. miRNA mediated regulation and interaction between plants and pathogens. Int. J. Mol. Sci. 2021, 22, 2913. [Google Scholar] [CrossRef] [PubMed]
- Secic, E.; Kogel, K.H.; Ladera-Carmona, M.J. Biotic stress-associated microRNA families in plants. J. Plant Physiol. 2021, 263, 153451. [Google Scholar] [CrossRef]
- Zhu, X.G.; He, S.D.; Fang, D.; Guo, L.; Zhou, X.Y.; Guo, Y.S.; Gao, L.; Qiao, Y.L. High-throughput sequencing-based identification of Arabidopsis miRNAs induced by Phytophthora capsici infection. Front. Microbiol. 2020, 11, 1094. [Google Scholar] [CrossRef]
- Luan, Y.S.; Cui, J.; Zhai, J.M.; Li, J.; Han, L.; Meng, J. High-throughput sequencing reveals differential expression of miRNAs in tomato inoculated with Phytophthora infestans. Planta 2015, 241, 1405–1416. [Google Scholar] [CrossRef]
- Guo, N.; Tahir, A.; Cui, X.X.; Xu, J.Y.; Sun, J.T.; Zhang, N.N.; Sun, R.D.; Deng, S.S.; Xing, H.; Zhao, J.M. Genome-wide identification of Phytophthora sojae-associated microRNAs and network in a resistant and a susceptible soybean germplasm. Agronomy 2022, 12, 2922. [Google Scholar] [CrossRef]
- Rabuma, T.; Gupta, O.P.; Chhokar, V. Genome-wide comprehensive analysis of miRNAs and their target genes expressed in resistant and susceptible Capsicum annuum genotypes during Phytophthora capsici infection. Mol. Genet. Genom. 2023, 298, 273–292. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Jiang, Y.; Peng, J.; Guo, J.; Lin, M.; Jin, C.; Huang, J.; Tang, W.; Guan, D.; He, S. The transcriptional reprograming and functional identification of WRKY family members in pepper’s response to Phytophthora capsici infection. BMC Plant Biol. 2020, 20, 256. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Lin, M.L.; Qiu, M.; Kong, L.; Xu, Y.P.; Li, Y.N.; Wang, Y.; Ye, W.W.; Dong, S.M.; He, S.L.; et al. Chitin synthase is involved in vegetative growth, asexual reproduction and pathogenesis of Phytophthora capsici and Phytophthora sojae. Environ. Microbiol. 2019, 21, 4537–4547. [Google Scholar] [CrossRef] [PubMed]
- Fiancette, R.; Finlay, C.M.; Willis, C.; Bevington, S.L.; Soley, J.; Ng, S.T.H.; Baker, S.M.; Andrews, S.; Hepworth, M.R.; Withers, D.R. Reciprocal transcription factor networks govern tissue-resident ILC3 subset function and identity. Nat. Immunol. 2021, 22, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Wingett, S.W.; Andrews, S.; Fraser, P.; Morf, J. RNA proximity sequencing data and analysis pipeline from a human neuroblastoma nuclear transcriptome. Sci. Data 2020, 7, 35. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.Z.; Huang, J.J.; Ding, Y.; Wu, J.M.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L.P. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. Embnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Z.; Wang, Y.; Li, L.; Yang, X.Z. miRDeep-P2: Accurate and fast analysis of the microRNA transcriptome in plants. Bioinformatics 2019, 35, 2521–2522. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.B.; Zhuang, Z.H.; Zhao, P.X.C. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.J.; Ma, Y.K.; Chen, T.; Wang, M.; Wang, X.J. PsRobot: A web-based plant small RNA meta-analysis toolbox. Nucleic Acids Res. 2012, 40, W22–W28. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.S.; Cui, J.; Li, J.; Jiang, N.; Liu, P.; Meng, J. Effective enhancement of resistance to Phytophthora infestans by overexpression of miR172a and b in Solanum lycopersicum. Planta 2018, 247, 127–138. [Google Scholar] [CrossRef]
- de Vries, S.; Kukuk, A.; von Dahlen, J.K.; Schnake, A.; Kloesges, T.; Rose, L.E. Expression profiling across wild and cultivated tomatoes supports the relevance of early miR482/2118 suppression for Phytophthora resistance. Proc. R. Soc. B-Biol. Sci. 2018, 285, 20172560. [Google Scholar] [CrossRef]
- Cui, X.X.; Yan, Q.; Gan, S.P.; Xue, D.; Dou, D.L.; Guo, N.; Xing, H. Overexpression of gma-miR1510a/b suppresses the expression of a NB-LRR domain gene and reduces resistance to. Gene 2017, 621, 32–39. [Google Scholar] [CrossRef]
- Wong, J.; Gao, L.; Yang, Y.; Zhai, J.X.; Arikit, S.; Yu, Y.; Duan, S.Y.; Chan, V.; Xiong, Q.; Yan, J.; et al. Roles of small RNAs in soybean defense against Phytophthora sojae infection. Plant J. 2014, 79, 928–940. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Hong, Y.H.; Liu, Y.R.; Cui, J.; Luan, Y.S. Function identification of miR394 in tomato resistance to Phytophthora infestans. Plant Cell Rep. 2021, 40, 1831–1844. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Luan, Y.S.; Zhai, J.M. Sp-miR396a-5p acts as a stress-responsive genes regulator by conferring tolerance to abiotic stresses and susceptibility to Phytophthora nicotianae infection in transgenic tobacco. Plant Cell Rep. 2015, 34, 2013–2025. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.H.; Wang, N.Q.; Jalajakumari, M.; Blackman, L.; Shen, E.H.; Verma, S.; Wang, M.B.; Millar, A.A. miR159 represses a constitutive pathogen defense response in tobacco. Plant Physiol. 2020, 182, 2182–2198. [Google Scholar] [CrossRef]
- Natarajan, B.; Kalsi, H.S.; Godbole, P.; Malankar, N.; Thiagarayaselvam, A.; Siddappa, S.; Thulasiram, H.V.; Chakrabarti, S.K.; Banerjee, A.K. MiRNA160 is associated with local defense and systemic acquired resistance against Phytophthora infestans infection in potato. J. Exp. Bot. 2018, 69, 2023–2036. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Deng, S.S.; Xuan, H.D.; Fan, X.X.; Sun, R.D.; Zhao, J.M.; Wang, H.T.; Guo, N.; Xing, H. A novel TIR-NBS-LRR gene regulates immune response to Phytophthora root rot in soybean. Crop J. 2022, 10, 1644–1653. [Google Scholar] [CrossRef]
- Jiang, N.; Meng, J.; Cui, J.; Sun, G.X.; Luan, Y.S. Function identification of miR482b, a negative regulator during tomato resistance to Phytophthora infestans. Hortic. Res. 2018, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.Y.; Wei, Z.Y.; Song, P.W.; Zhou, L.Y.; Hu, H.Y.; Hu, P.; Li, C.W. MicroRNA expression profiles in response to Phytophthora infestans and Oidium neolycopersici and functional identification of sly-miR397 in tomato. Phytopathology 2023, 113, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.H.; Meng, J.; He, X.L.; Zhang, Y.Y.; Luan, Y.S. Overexpression of miR482c in tomato induces enhanced susceptibility to late blight. Cells 2019, 8, 822. [Google Scholar] [CrossRef]
- Li, Y.; Yu, T.; Wu, T.Q.; Wang, R.; Wang, H.M.; Du, H.; Xu, X.W.; Xie, D.S.; Xu, X.M. The dynamic transcriptome of pepper (Capsicum annuum) whole roots reveals an important role for the phenylpropanoid biosynthesis pathway in root resistance to Phytophthora capsici. Gene 2020, 728, 144288. [Google Scholar] [CrossRef]
- Xie, M.; Zhang, J.; Tschaplinski, T.J.; Tuskan, G.A.; Chen, J.G.; Muchero, W. Regulation of lignin biosynthesis and its role in growth-defense tradeoffs. Front. Plant Sci. 2018, 9, 1427. [Google Scholar] [CrossRef]
- Prasad, A.; Sharma, N.; Chirom, O.; Prasad, M. The sly-miR166-SlyHB module acts as a susceptibility factor during ToLCNDV infection. Theor. Appl. Genet. 2022, 135, 233–242. [Google Scholar] [CrossRef]
- Yang, Z.Y.; Hui, S.G.; Lv, Y.; Zhang, M.J.; Chen, D.; Tian, J.J.; Zhang, H.T.; Liu, H.B.; Cao, J.B.; Xie, W.Y.; et al. miR395-regulated sulfate metabolism exploits pathogen sensitivity to sulfate to boost immunity in rice. Mol. Plant 2022, 15, 671–688. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.F.; Bhutto, S.H.; He, X.R.; Yang, X.M.; Zhou, X.H.; Lin, X.Y.; Rajput, A.A.; Li, G.B.; Zhao, J.H.; et al. Blocking miR530 improves rice resistance, yield, and maturity. Front. Plant Sci. 2021, 12, 729560. [Google Scholar] [CrossRef]
- Bhar, A.; Roy, A. Emphasizing the role of long non-coding RNAs (lncRNA), circular RNA (circRNA), and micropeptides (miPs) in plant biotic stress tolerance. Plants 2023, 12, 3951. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wang, N.; Guo, J.; Zhou, X.; Bai, X.; Azeem, M.; Zhu, L.; Chen, L.; Chu, M.; Wang, H.; et al. Integrative Transcriptome Analysis of mRNA and miRNA in Pepper’s Response to Phytophthora capsici Infection. Biology 2024, 13, 186. https://doi.org/10.3390/biology13030186
Li Y, Wang N, Guo J, Zhou X, Bai X, Azeem M, Zhu L, Chen L, Chu M, Wang H, et al. Integrative Transcriptome Analysis of mRNA and miRNA in Pepper’s Response to Phytophthora capsici Infection. Biology. 2024; 13(3):186. https://doi.org/10.3390/biology13030186
Chicago/Turabian StyleLi, Yuan, Nan Wang, Jianwen Guo, Xianjun Zhou, Xueyi Bai, Muhammad Azeem, Liyun Zhu, Lin Chen, Moli Chu, Hui Wang, and et al. 2024. "Integrative Transcriptome Analysis of mRNA and miRNA in Pepper’s Response to Phytophthora capsici Infection" Biology 13, no. 3: 186. https://doi.org/10.3390/biology13030186
APA StyleLi, Y., Wang, N., Guo, J., Zhou, X., Bai, X., Azeem, M., Zhu, L., Chen, L., Chu, M., Wang, H., & Cheng, W. (2024). Integrative Transcriptome Analysis of mRNA and miRNA in Pepper’s Response to Phytophthora capsici Infection. Biology, 13(3), 186. https://doi.org/10.3390/biology13030186