

Spatial Pattern of Host Tree Size, Rather than of Host Tree Itself, Affects the Infection Likelihood of a Fungal Stem Disease

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
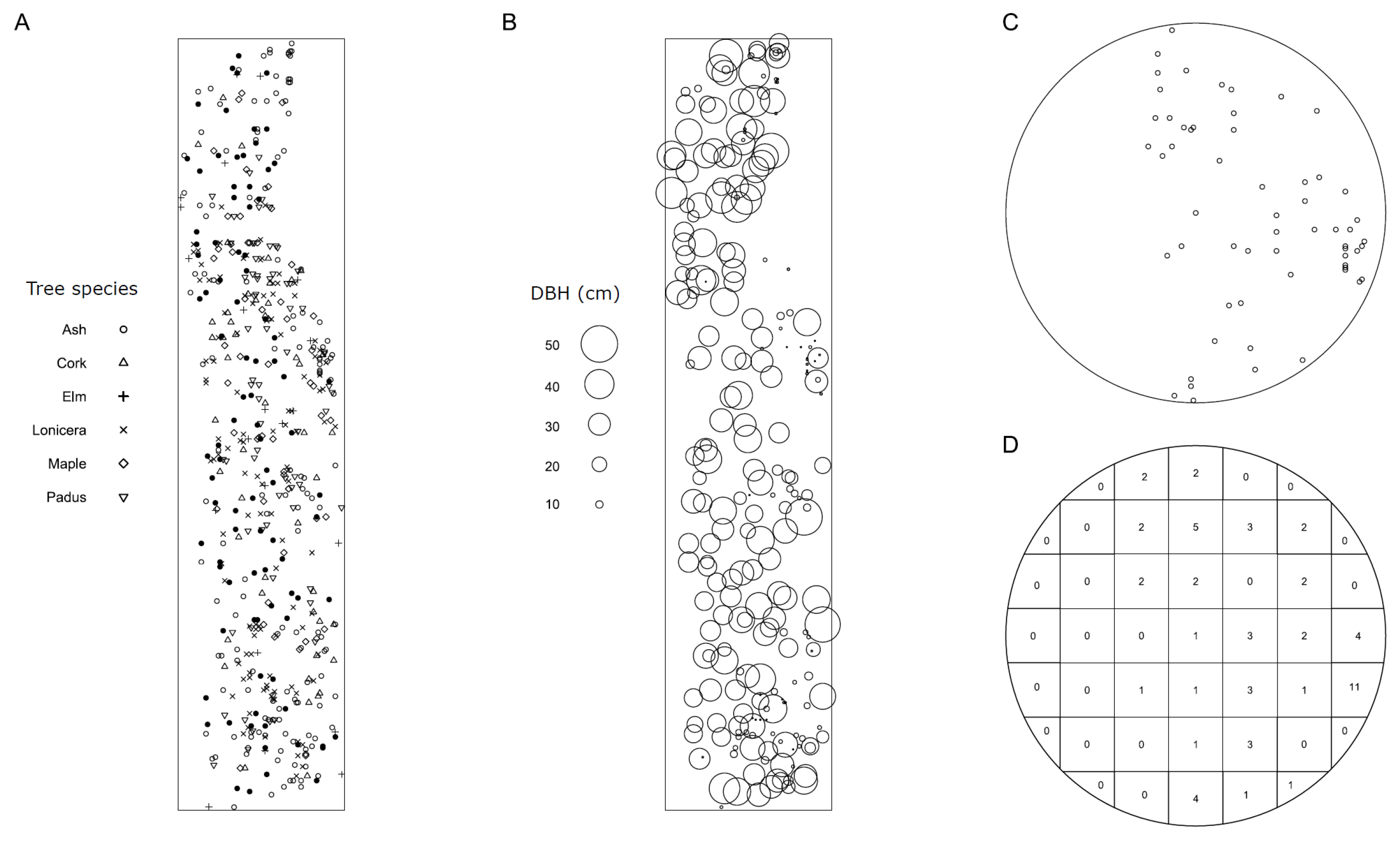
2.1. Study System
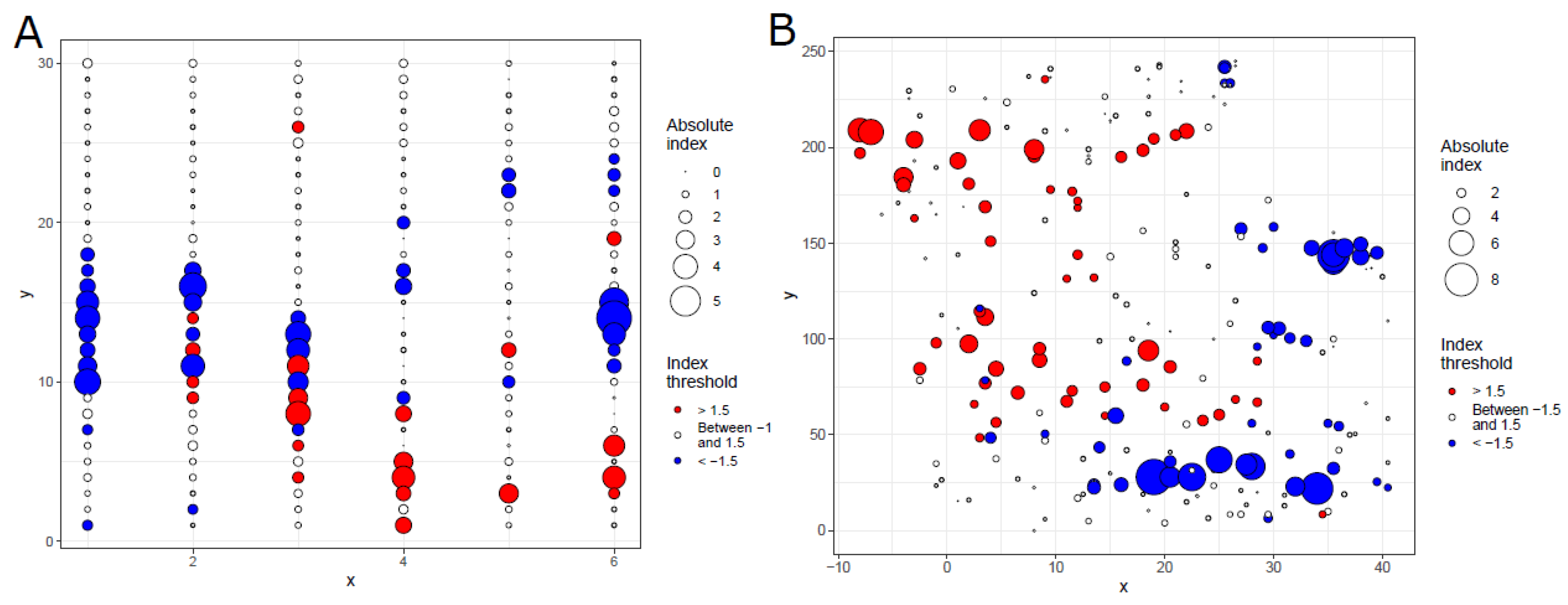
2.2. SADIE Analysis
2.3. Random Labeling and Trivariate Random Labeling
2.4. Spatial Point Pattern Test
2.5. Testing the Effect of DBH on Infection Patterns
2.6. Generalized Linear Models (GLMs) and Meta-Analysis
2.7. Statistics and Software
3. Results
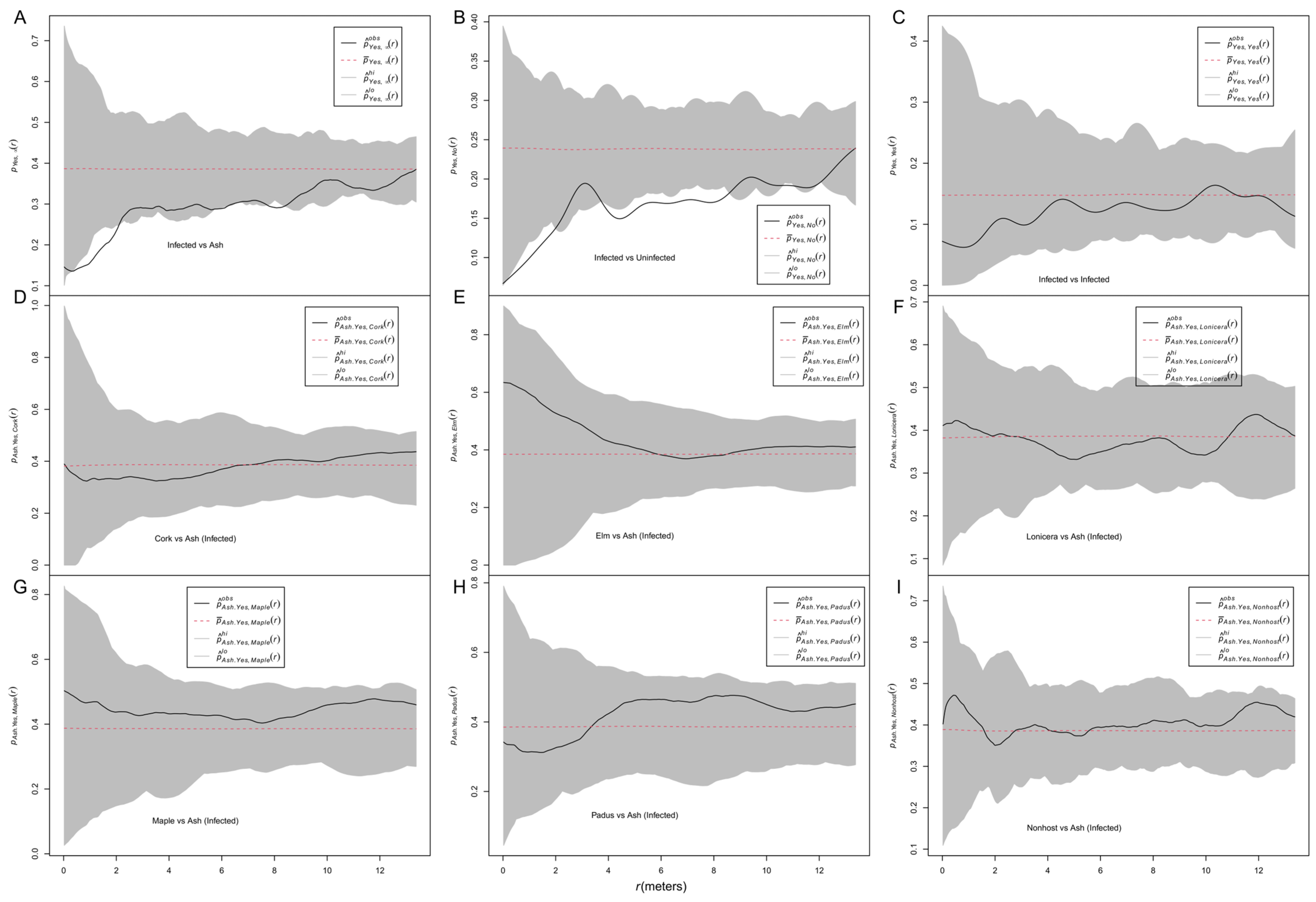
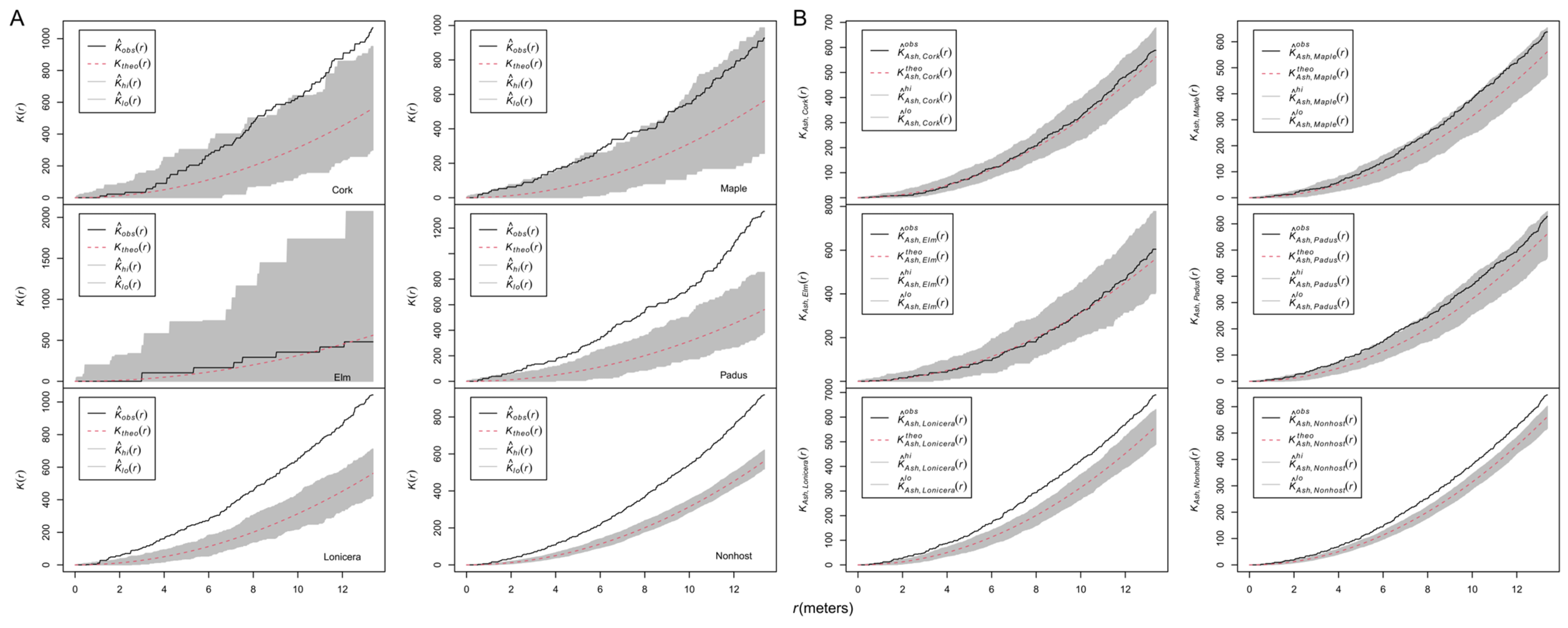
3.1. Correlation between Infection and Antecedent Spatial Patterns of Hosts/Nonhosts
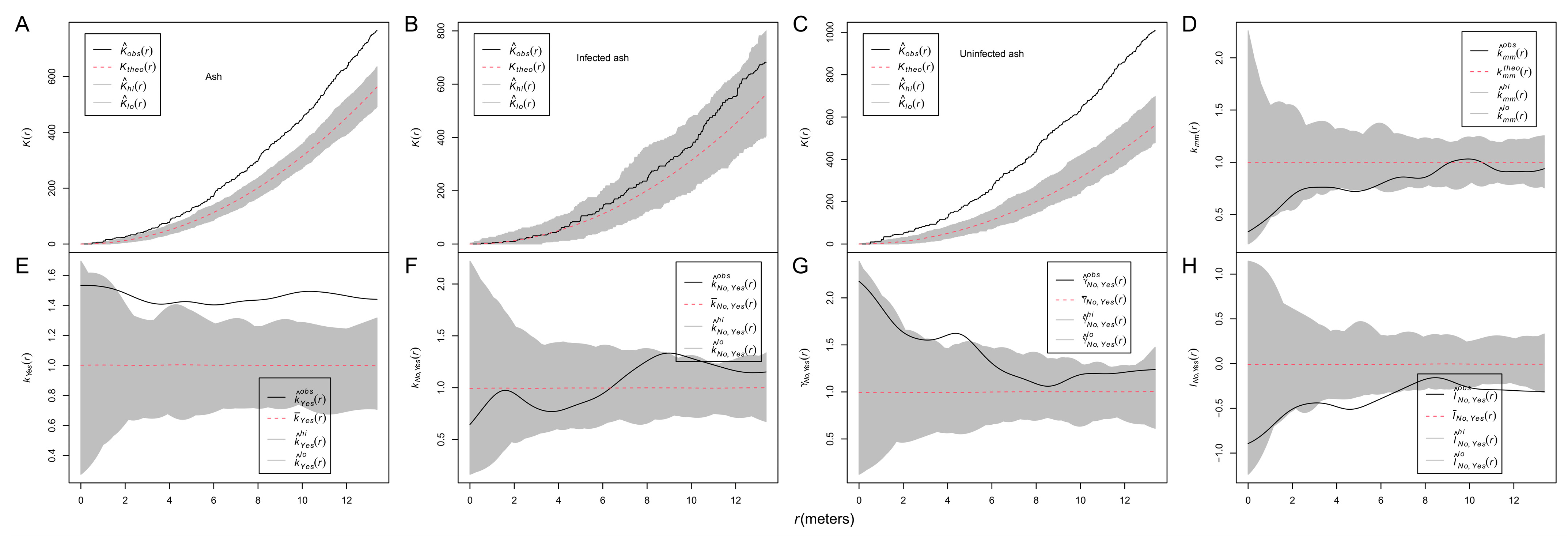
3.2. Spatial Patterns of Ash Trees and Their Sizes
3.3. Spatial Pattern of Nonhost Species
3.4. Spatial Pattern Effect of Ash Trees, Their DBHs, and Nonhost Trees on Infection Likelihood
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Real, L.A.; McElhany, P. Spatial pattern and process in plant-pathogen interactions. Ecology 1996, 77, 1011–1025. [Google Scholar] [CrossRef]
- Ndoungué Djeumekop, M.M.; Ngo Bieng, M.-A.; Ribeyre, F.; Bonnot, F.; Cilas, C.; Neema, C.; ten Hoopen, G.M. Spatial and temporal analysis of phytophthora megakarya epidemic in newly established cacao plantations. Plant Dis. 2021, 105, 1448–1460. [Google Scholar] [CrossRef] [PubMed]
- Ben-Hamo, M.; Ezra, D.; Krasnov, H.; Blank, L. Spatial and temporal dynamics of mal secco disease spread in lemon orchards in israel. Phytopathology 2020, 110, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Wallis, A.; Cox, K.D. Examining spatial distribution and spread of fire blight in apple orchards: Two case studies. Plant Health Progr. 2021, 22, 445–449. [Google Scholar] [CrossRef]
- Sasal, Y.; Amico, G.C.; Morales, J.M. Host spatial structure and disperser activity determine mistletoe infection patterns. Oikos 2021, 130, 440–452. [Google Scholar] [CrossRef]
- Gilligan, C.A. Statistical analysis of the spatial pattern of botrytis fabae on vicia faba: A methodological study. Trans. Br. Mycol. Soc. 1982, 79, 193–200. [Google Scholar] [CrossRef]
- Koricheva, J.; Vehviläinen, H.; Riihimäki, J.; Ruohomäki, K.; Kaitaniemi, P.; Ranta, H. Diversification of tree stands as a means to manage pests and diseases in boreal forests: Myth or reality? Can. J. For. Res. 2006, 36, 324–336. [Google Scholar] [CrossRef]
- Zhai, L.; Coyle, D.R.; Li, D.; Jonko, A. Fire, insect and disease-caused tree mortalities increased in forests of greater structural diversity during drought. J. Ecol. 2022, 110, 673–685. [Google Scholar] [CrossRef]
- Jules, E.S.; Carroll, A.L.; Garcia, A.M.; Steenbock, C.M.; Kauffman, M.J. Host heterogeneity influences the impact of a non-native disease invasion on populations of a foundation tree species. Ecosphere 2014, 5, art105. [Google Scholar] [CrossRef]
- Schwarze, F.; Fink, S. Reaction zone penetration and prolonged persistence of xylem rays in london plane wood degraded by the basidiomycete inonotus hispidus. Mycol. Res. 1997, 101, 1207–1214. [Google Scholar] [CrossRef]
- Schwarze, F.; Lonsdale, D.; Fink, S. Soft rot and multiple t-branching by the basidiomycete inonotus hispidus in ash and london plane. Mycol. Res. 1995, 99, 813–820. [Google Scholar] [CrossRef]
- Fischer, M.; García, V.G. An annotated checklist of european basidiomycetes related to white rot of grapevine (vitis vinifera). Phytopathol. Mediterr. 2015, 54, 281–298. [Google Scholar]
- Meadows, J.S.; Leininger, T.D.; Nebeker, T.E. Thinning to Improve Growth and Control the Canker Decay Fungus Inonotus Hispidus in a Red Oak-Sweetgum Stand in the Mississippi Delta; General Technical Report SRS-48; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2002; pp. 183–188.
- Dai, Y.-C.; Qin, G.-F.; Xiu, M.-Q. The forest pathogens of root and butt rot in northeast china. For. Res. 2000, 13, 15–22. [Google Scholar]
- Krimi, Z.; Mehdid, S. Identification of wood-decay fungi infecting various forest trees. EPPO Bull. 2001, 31, 114–115. [Google Scholar] [CrossRef]
- Khasanov, B.A. Fungal diseases of persian walnut in uzbekistan and their control. Ind. J. Pure App. Biosci. 2019, 7, 327–334. [Google Scholar] [CrossRef]
- Gavrikov, V.; Stoyan, D. The use of marked point processes in ecological and environmental forest studies. Environ. Ecol. Stat. 1995, 2, 331–344. [Google Scholar] [CrossRef]
- Perry, J.N. Spatial analysis by distance indices. J. Anim. Ecol. 1995, 64, 303–314. [Google Scholar] [CrossRef]
- Kennington, J.L.; Helgason, R.V. Algorithms for Network Programming; Wiley: New York, NY, USA, 1980. [Google Scholar]
- Greene, A.D.; Reay-Jones, F.P.F.; Kirk, K.R.; Peoples, B.K.; Greene, J.K. Spatial associations of key lepidopteran pests with defoliation, ndvi, and plant height in soybean. Environ. Entomol. 2021, 50, 1378–1392. [Google Scholar] [CrossRef] [PubMed]
- Raventós, J.; Wiegand, T.; de Luis, M. Evidence for the spatial segregation hypothesis: A test with nine-year survivorship data in a mediterranean shrubland. Ecology 2010, 917, 2110–2120. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Handbook of Spatial Point-Pattern Analysis in Ecology; Chapman and Hall/CRC: Boca Raton, FL, USA, 2014. [Google Scholar]
- Ripley, B.D. The second-order analysis of stationary point processes. J. Appl. Probab. 1976, 13, 255–266. [Google Scholar] [CrossRef]
- Goreaud, F.; Pélissier, R. Avoiding misinterpretation of biotic interactions with the intertype k12-function: Population independence vs. Random labelling hypotheses. J. Veg. Sci. 2003, 14, 681–692. [Google Scholar]
- Stoyan, D.; Stoyan, H. Fractals, Random Shapes and Point Fields: Methods of Geometrical Statistics; John Wiley and Sons: Chichester, UK, 1994. [Google Scholar]
- Wälder, O.; Stoyan, D. On variograms in point process statistics. Biometrical J. 1996, 38, 895–905. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Chacón-Labella, J.; de la Cruz, M.; Pescador, D.S.; Escudero, A. Individual species affect plant traits structure in their surroundings: Evidence of functional mechanisms of assembly. Oecologia 2016, 180, 975–987. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Mejia, L.; Kitching, I.J.; Jetz, W.; Beck, J. Putting insects on the map: Near-global variation in sphingid moth richness along spatial and environmental gradients. Ecography 2017, 40, 698–708. [Google Scholar] [CrossRef]
- Baddeley, R.A.; Rubak, E.; Turner, R. Spatial Point Patterns: Methodology and Applications with R; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Gigot, C. Epiphy: Analysis of Plant Disease Epidemics, R Package Version 0.3.4. 2018. Available online: https://CRAN.R-project.org/package=epiphy (accessed on 11 August 2024).
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- Vacher, C.; Vile, D.; Helion, E.; Piou, D.; Desprez-Loustau, M.-L. Distribution of parasitic fungal species richness: Influence of climate versus host species diversity. Divers. Distrib. 2008, 14, 786–798. [Google Scholar] [CrossRef]
- Nguyen, D.; Castagneyrol, B.; Bruelheide, H.; Bussotti, F.; Guyot, V.; Jactel, H.; Jaroszewicz, B.; Valladares, F.; Stenlid, J.; Boberg, J. Fungal disease incidence along tree diversity gradients depends on latitude in european forests. Ecol. Evol. 2016, 6, 2426–2438. [Google Scholar] [CrossRef]
- Franić, I.; Allan, E.; Prospero, S.; Adamson, K.; Attorre, F.; Auger-Rozenberg, M.-A.; Augustin, S.; Avtzis, D.; Baert, W.; Barta, M.; et al. Climate, host and geography shape insect and fungal communities of trees. Sci. Rep. 2023, 13, 11570. [Google Scholar] [CrossRef]
- Kuang, J.; Han, S.; Chen, Y.; Bates, C.T.; Wang, P.; Shu, W. Root-associated fungal community reflects host spatial co-occurrence patterns in a subtropical forest. ISME Commun. 2021, 1, 65. [Google Scholar] [CrossRef]
- Amico, G.C.; Rodriguez-Cabal, M.A.; Aizen, M.A. Geographic variation in fruit colour is associated with contrasting seed disperser assemblages in a south-andean mistletoe. Ecography 2011, 34, 318–326. [Google Scholar] [CrossRef]
- Koyani, R.D.; Sanghvi, G.V.; Bhatt, I.M.; Rajput, K.S. Pattern of delignification in ailanthus excelsa roxb. Wood by inonotus hispidus (bull.: Fr.) karst. Mycology 2010, 1, 204–211. [Google Scholar] [CrossRef]
- Saadani, M.; Hönig, L.; Bien, S.; Koehler, M.J.; Rutten, G.; Wubet, T.; Braun, U.; Bruelheide, H. Local tree diversity suppresses foliar fungal infestation and decreases morphological but not molecular richness in a young subtropical forest. J. Fungi 2021, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Pélissier, R.; Buendia, L.; Brousse, A.; Temple, C.; Ballini, E.; Fort, F.; Violle, C.; Morel, J.-B. Plant neighbour-modulated susceptibility to pathogens in intraspecific mixtures. J. Exp. Bot. 2021, 72, 6570–6580. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, M.J.; Jules, E.S. Heterogeneity shapes invasion: Host size and environment influence susceptibility to a nonnative pathogen. Ecol. Appl. 2006, 16, 166–175. [Google Scholar] [CrossRef]
- Xu, X.-M.; Madden, L.V. Use of sadie statistics to study spatial dynamics of plant disease epidemics. Plant Pathol. 2004, 53, 38–49. [Google Scholar] [CrossRef]
- Tredennick, A.T.; Hooker, G.; Ellner, S.P.; Adler, P.B. A practical guide to selecting models for exploration, inference, and prediction in ecology. Ecology 2021, 102, e03336. [Google Scholar] [CrossRef]
- Fagard, R.H.; Staessen, J.A.; Thijs, L. Advantages and disadvantages of the meta-analysis approach. J. Hypertens. 1996, 14, S9–S13. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Explanatory | Estimate | Std. Error | z Value | Pr(>|z|) | Radius of Plot (m) |
|---|---|---|---|---|---|---|
| global vi(tree) | (Intercept) | −6.73 × 10−15 | 4.71 × 10−1 | 0 | 1 | |
| vi.CatRandom | −5.94 × 10−1 | 4.99 × 10−1 | −1.189 | 0.234 | ||
| vi.CatPatch | −2.68 × 10−1 | 5.39 × 10−1 | −0.498 | 0.618 | ||
| global vi(DBH) | (Intercept) | −2.2824 | 0.4694 | −4.862 | 1.16 × 10−6 | |
| vi.CatRandom | 1.7756 | 0.5024 | 3.534 | 0.0004 | ||
| vi.CatPatch | 3.1474 | 0.556 | 5.661 | 1.51 × 10−8 | ||
| local vi(DBH) | vi.CatRandom | 2.4980 | 1.0487 | 2.3820 | 0.0172 | 10 |
| vi.CatRandom | 2.6935 | 1.0468 | 2.5729 | 0.0101 | 11 | |
| vi.CatRandom | 2.2678 | 0.7652 | 2.9637 | 0.0030 | 12 | |
| vi.CatRandom | 2.9618 | 1.0446 | 2.8353 | 0.0046 | 13 | |
| vi.CatPatch | 3.1781 | 1.2802 | 2.4825 | 0.0130 | 13 | |
| vi.CatRandom | 2.5421 | 1.0590 | 2.4005 | 0.0164 | 14 | |
| vi.CatPatch | 2.3026 | 1.0488 | 2.1954 | 0.0281 | 15 | |
| vi.CatRandom | 1.6458 | 0.8310 | 1.9804 | 0.0477 | 19 | |
| vi.CatRandom | 1.9363 | 0.7224 | 2.6806 | 0.0073 | 20 | |
| vi.CatPatch | 2.0794 | 0.8898 | 2.3371 | 0.0194 | 20 | |
| vi.CatRandom | 2.1893 | 0.8435 | 2.5956 | 0.0094 | 21 | |
| vi.CatPatch | 2.5903 | 1.0288 | 2.5179 | 0.0118 | 21 | |
| vi.CatRandom | 2.5745 | 0.8790 | 2.9290 | 0.0034 | 22 | |
| vi.CatPatch | 3.1135 | 1.1005 | 2.8292 | 0.0047 | 22 | |
| vi.CatRandom | 2.1972 | 0.8975 | 2.4481 | 0.0144 | 23 | |
| vi.CatPatch | 3.3322 | 1.3296 | 2.5062 | 0.0122 | 23 | |
| vi.CatRandom | 2.8214 | 1.1992 | 2.3527 | 0.0186 | 24 |
| Explanatory | K | I2 | Pooled Effect Size | Meta-Regression | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pooled Beta | Lower Bound | Upper Bound | Zval | Pval | Beta | se | Zval | Pval | R2 | |||
| vi.CatRandom | 20 | 0 | 1.6772 | 1.3151 | 2.0392 | 9.0804 | 1.08 × 10−19 | 0.0161 | 0.0366 | 0.4406 | 0.6595 | 0 |
| vi.CatPatch | 20 | 0 | 1.9380 | 1.3748 | 2.5013 | 6.7440 | 1.54 × 10−11 | 0.0784 | 0.0631 | 1.2415 | 0.2144 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Gao, X.; Jiang, Y.; Zhang, J.; Qi, F.-H.; Jing, T.-Z. Spatial Pattern of Host Tree Size, Rather than of Host Tree Itself, Affects the Infection Likelihood of a Fungal Stem Disease. Biology 2024, 13, 616. https://doi.org/10.3390/biology13080616
Shi Y, Gao X, Jiang Y, Zhang J, Qi F-H, Jing T-Z. Spatial Pattern of Host Tree Size, Rather than of Host Tree Itself, Affects the Infection Likelihood of a Fungal Stem Disease. Biology. 2024; 13(8):616. https://doi.org/10.3390/biology13080616
Chicago/Turabian StyleShi, Yanli, Xinbo Gao, Yunxiao Jiang, Junsheng Zhang, Feng-Hui Qi, and Tian-Zhong Jing. 2024. "Spatial Pattern of Host Tree Size, Rather than of Host Tree Itself, Affects the Infection Likelihood of a Fungal Stem Disease" Biology 13, no. 8: 616. https://doi.org/10.3390/biology13080616
APA StyleShi, Y., Gao, X., Jiang, Y., Zhang, J., Qi, F.-H., & Jing, T.-Z. (2024). Spatial Pattern of Host Tree Size, Rather than of Host Tree Itself, Affects the Infection Likelihood of a Fungal Stem Disease. Biology, 13(8), 616. https://doi.org/10.3390/biology13080616