
Light-Exposed Metabolic Responses of Cordyceps militaris through Transcriptome-Integrated Genome-Scale Modeling


 , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Retrofitting GSMM of C. militaris for Growth under Light Conditions
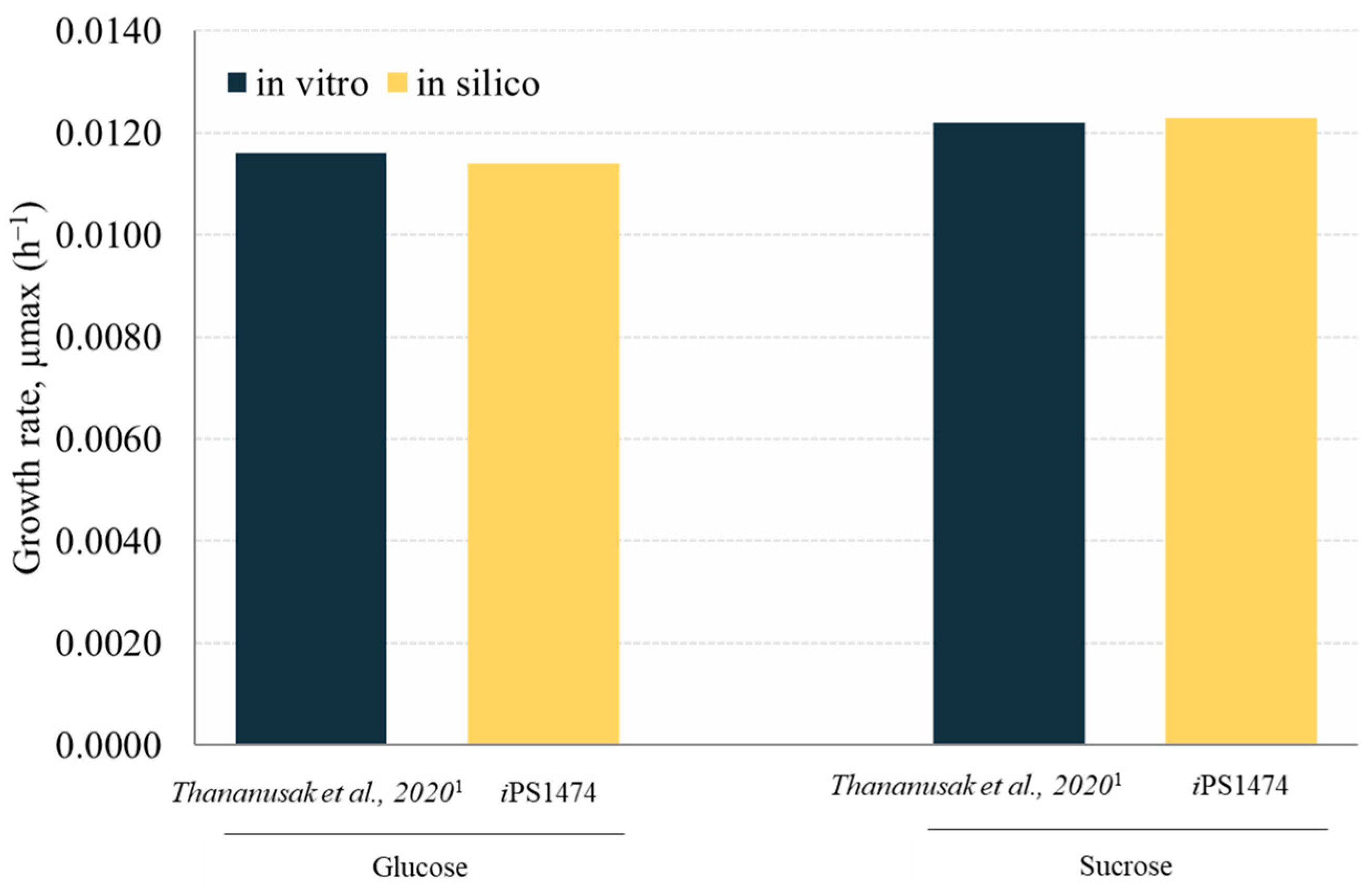
2.2. Validation of the Retrofitted GSMM of C. militaris
2.3. Transcriptome-Integrated Constraint-Based Metabolic Model of C. militaris Using GIMME
Subject to: Sij · vi = 0,
lbi < vi < ubi,
ci = {xthreshold − xi where xthreshold > xi, 0 otherwise},
2.4. Analysis of Active Flux Distribution and Metabolic Reaction under Light Exposure
2.5. Using the Retrofitted GSMM for Identifying Reporter Metabolites and Key Sub-Networks
3. Results and Discussion
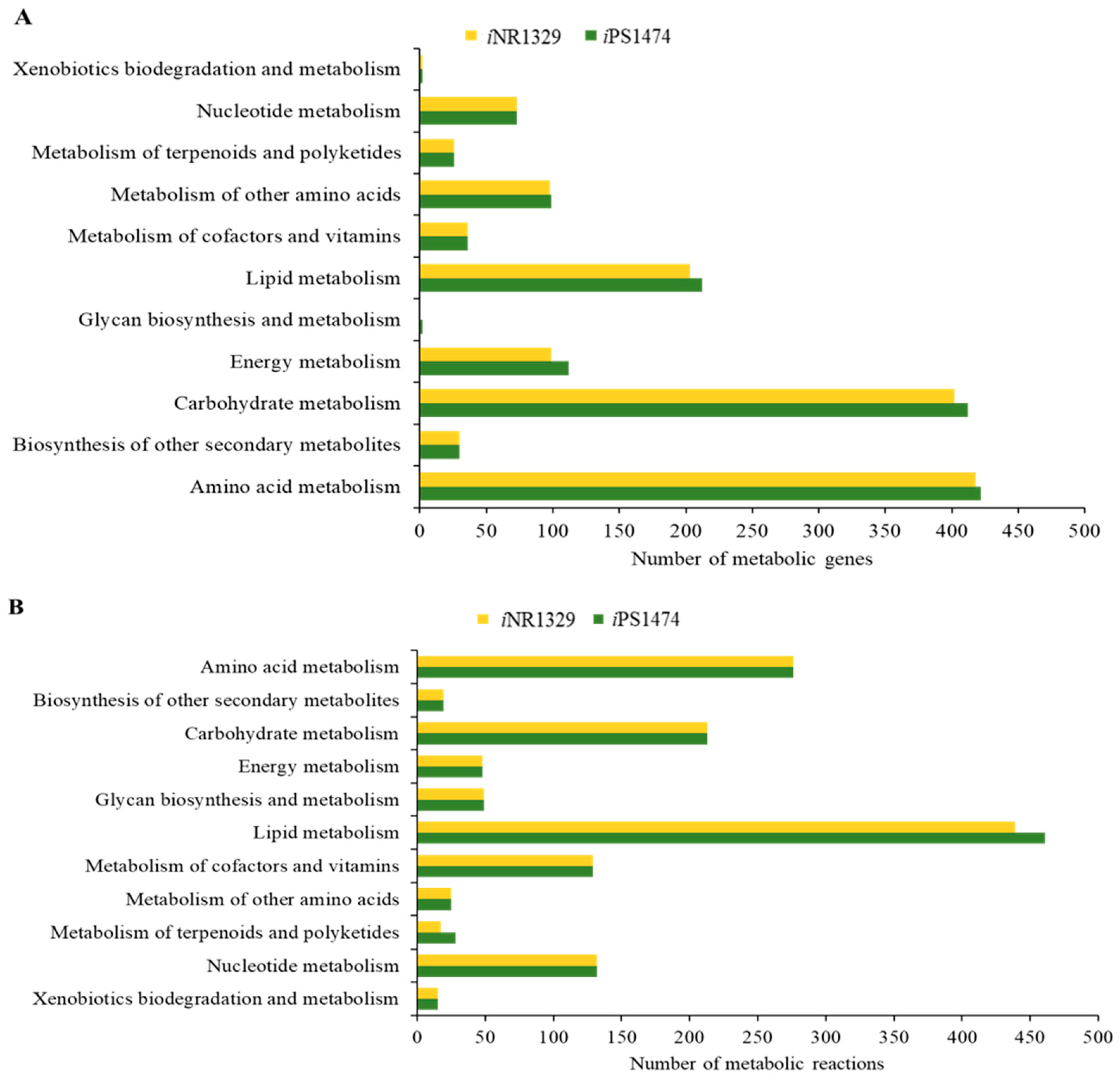
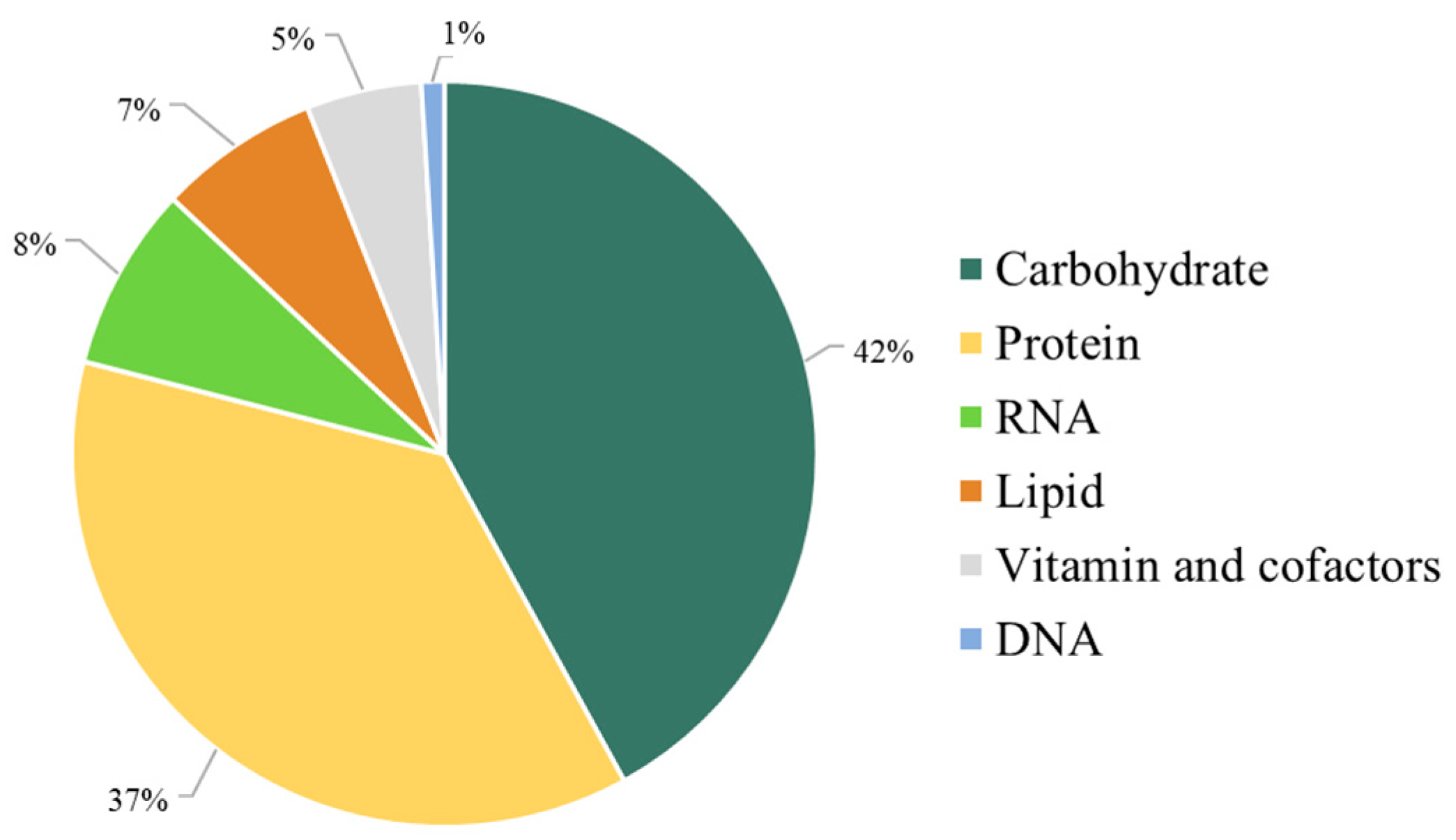
3.1. The Retrofitted iPS1474-GSMM of C. militaris in Comparison with Earlier Model Characteristics
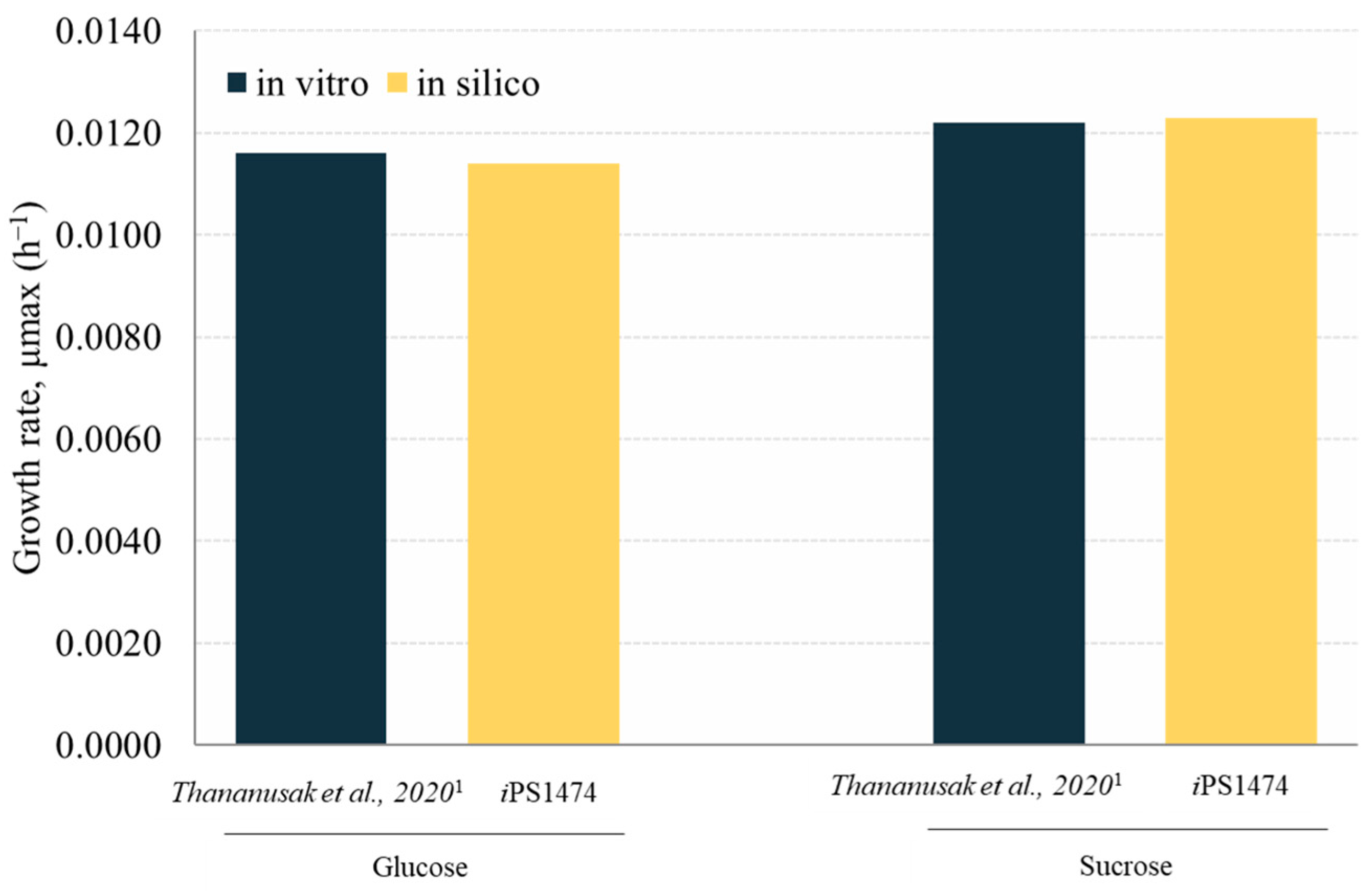
3.2. Validation and Performance of iPS1474-GSMM Model
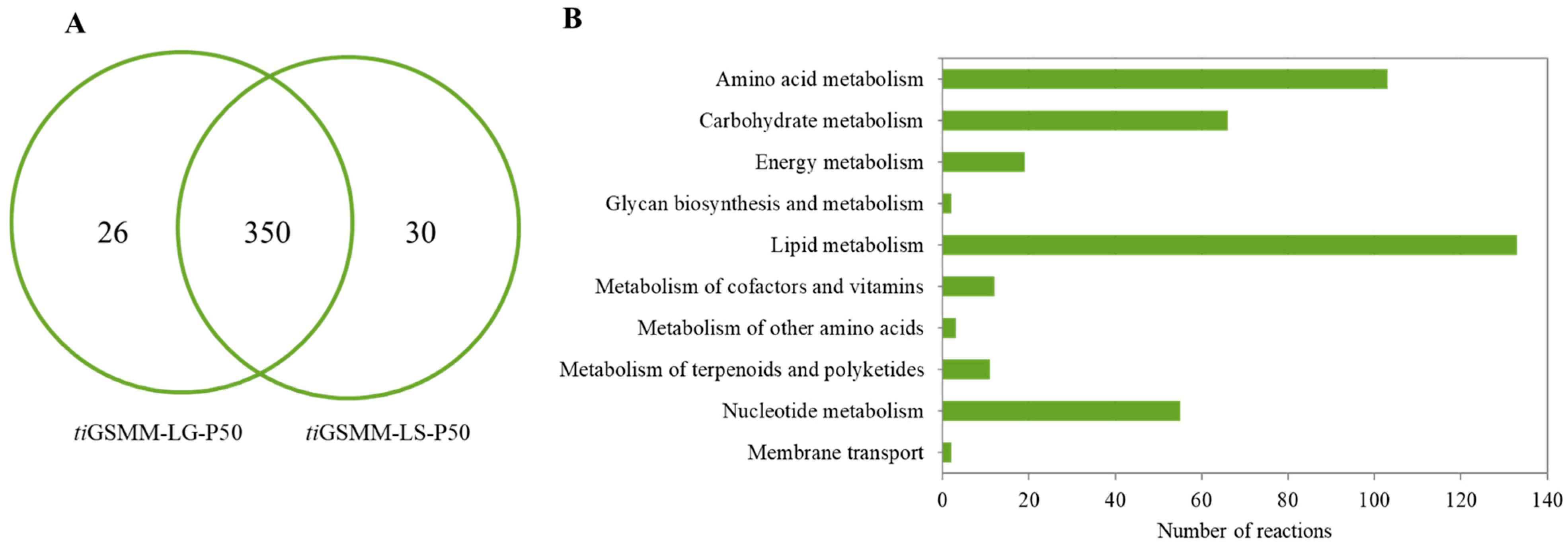
3.3. Transcriptome-Integrated GSMM (iPS1474-tiGSMM) Model of C. militaris under Light Conditions
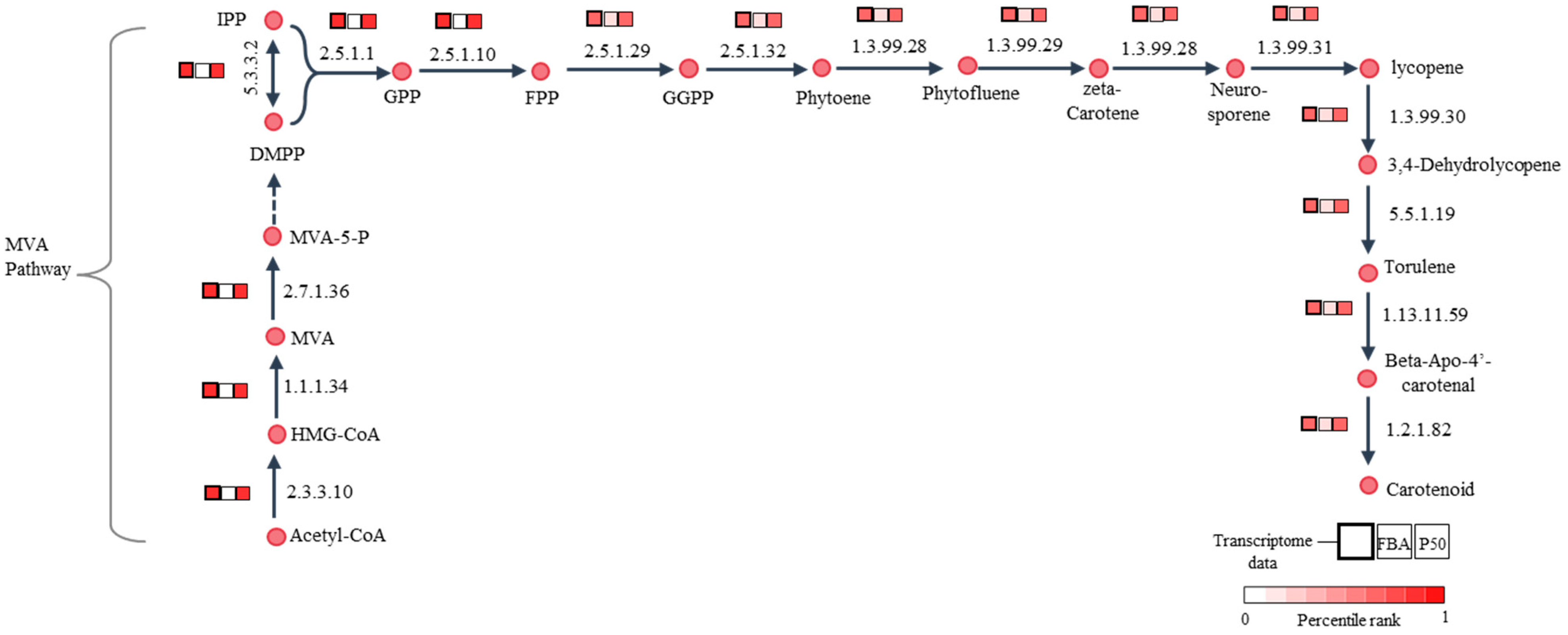
3.4. Active Light-Exposed Metabolic Reactions Predicted by iPS1474-tiGSMM-P50
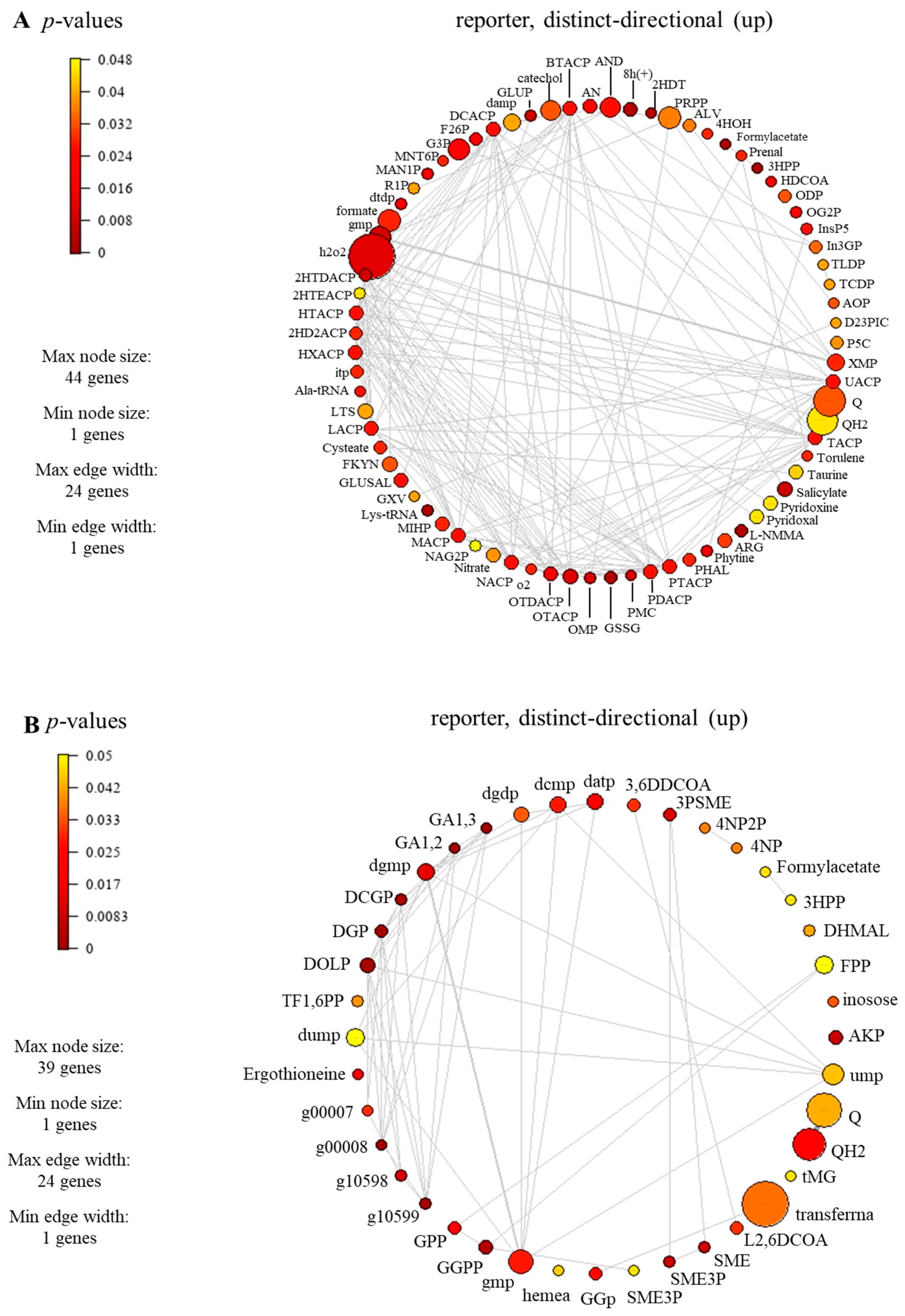
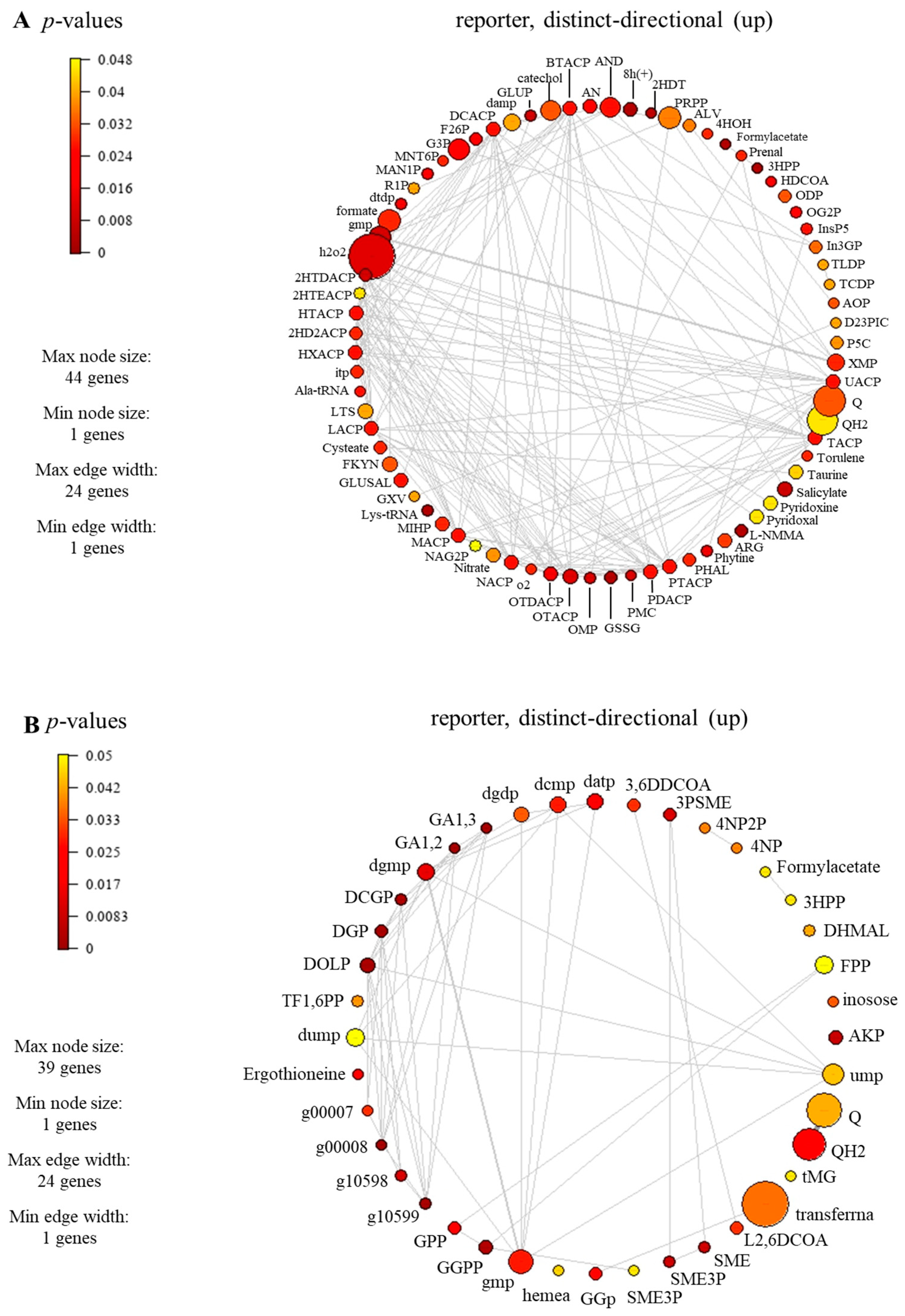
3.5. Identified Reporter Metabolites of C. militaris
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cui, J.D. Biotechnological production and applications of Cordyceps militaris, a valued traditional Chinese medicine. Crit. Rev. Biotechnol. 2015, 35, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.; Barseghyan, G.; Asatiani, M.; Wasser, S. Chemical composition and medicinal value of fruiting bodies and submerged cultured mycelia of caterpillar medicinal fungus Cordyceps militaris CBS-132098 (Ascomycetes). Int. J. Med. Mushrooms 2015, 17, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Jędrejko, K.J.; Lazur, J.; Muszyńska, B. Cordyceps militaris: An overview of its chemical constituents in relation to biological activity. Foods 2021, 10, 2634. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Lei, C.; Zheng, X.; Ai, X.; Wang, Y.; Wang, Q. Light wavelengths regulate growth and active components of Cordyceps militaris fruit bodies. J. Food Biochem. 2013, 37, 578–584. [Google Scholar] [CrossRef]
- Dong, J.; Wang, S.; Ai, X.; Yao, L.; Sun, Z.; Lei, C.; Wang, Y.; Wang, Q. Composition and characterization of cordyxanthins from Cordyceps militaris fruit bodies. J. Funct. Foods 2013, 5, 1450–1455. [Google Scholar] [CrossRef]
- Dong, Y.; Jing, T.; Meng, Q.; Liu, C.; Hu, S.; Ma, Y.; Liu, Y.; Lu, J.; Cheng, Y.; Wang, D.; et al. Studies on the antidiabetic activities of Cordyceps militaris extract in diet-streptozotocin-induced diabetic sprague-dawley rats. BioMed Res. Int. 2014, 2014, 160980. [Google Scholar] [CrossRef]
- Liu, C.; Song, J.; Teng, M.; Zheng, X.; Li, X.; Tian, Y.; Pan, M.; Li, Y.; Lee, R.; Wang, D. Antidiabetic and antinephritic activities of aqueous extract of Cordyceps militaris fruit body in diet-streptozotocin-induced diabetic sprague dawley rats. Oxidative Med. Cell. Longev. 2016, 2016, 9685257. [Google Scholar] [CrossRef]
- Rupa, E.J.; Li, J.F.; Arif, M.H.; Yaxi, H.; Puja, A.M.; Chan, A.J.; Hoang, V.A.; Kaliraj, L.; Yang, D.C.; Kang, S.C. Cordyceps militaris fungus extracts-mediated nanoemulsion for improvement antioxidant, antimicrobial, and anti-inflammatory activities. Molecules 2020, 25, 5733. [Google Scholar] [CrossRef]
- Tuli, H.S.; Sharma, A.; Sandhu, S.; Kashyap, D. Cordycepin: A bioactive metabolite with therapeutic potential. Life Sci. 2013, 93, 863–869. [Google Scholar] [CrossRef]
- Yu, S.-H.; Dubey, N.; Li, W.-S.; Liu, M.-C.; Chiang, H.S.; Leu, S.-J.; Shieh, Y.-H.; Tsai, F.-C.; Deng, W.P. Cordyceps militaris treatment preserves renal function in type 2 diabetic nephropathy mice. PLoS ONE 2016, 11, e0166342. [Google Scholar] [CrossRef]
- Jiaojiao, Z.; Fen, W.; Kuanbo, L.; Qing, L.; Ying, Y.; Caihong, D. Heat and light stresses affect metabolite production in the fruit body of the medicinal mushroom Cordyceps militaris. Appl. Microbiol. Biotechnol. 2018, 102, 4523–4533. [Google Scholar] [CrossRef]
- Dong, J.Z.; Lei, C.; Ai, X.R.; Wang, Y. Selenium enrichment on Cordyceps militaris link and analysis on its main active components. Appl. Biochem. Biotechnol. 2012, 166, 1215–1224. [Google Scholar] [CrossRef] [PubMed]
- Thananusak, R.; Laoteng, K.; Raethong, N.; Zhang, Y.; Vongsangnak, W. Metabolic responses of carotenoid and cordycepin biosynthetic pathways in Cordyceps militaris under light-programming exposure through genome-wide transcriptional analysis. Biology 2020, 9, 242. [Google Scholar] [CrossRef]
- Raethong, N.; Wang, H.; Nielsen, J.; Vongsangnak, W. Optimizing cultivation of Cordyceps militaris for fast growth and cordycepin overproduction using rational design of synthetic media. Comput. Struct. Biotechnol. J. 2020, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cheawchanlertfa, P.; Chitcharoen, S.; Raethong, N.; Liu, Q.; Chumnanpuen, P.; Soommat, P.; Song, Y.; Koffas, M.; Laoteng, K.; Vongsangnak, W. Enhancing Genome-Scale Model by Integrative Exometabolome and Transcriptome: Unveiling carbon assimilation towards sphingolipid biosynthetic capability of Cordyceps militaris. J. Fungi 2022, 8, 887. [Google Scholar] [CrossRef]
- Becker, S.A.; Palsson, B.O. Context-specific metabolic networks are consistent with experiments. PLoS Comput. Biol. 2008, 4, e1000082. [Google Scholar] [CrossRef]
- Zur, H.; Ruppin, E.; Shlomi, T. iMAT: An integrative metabolic analysis tool. Bioinformatics 2010, 26, 3140–3142. [Google Scholar] [CrossRef]
- Robaina Estévez, S.; Nikoloski, Z. Context-specific metabolic model extraction based on regularized least squares optimization. PLoS ONE 2015, 10, e0131875. [Google Scholar] [CrossRef] [PubMed]
- Casini, I.; McCubbin, T.; Esquivel-Elizondo, S.; Luque, G.G.; Evseeva, D.; Fink, C.; Beblawy, S.; Youngblut, N.D.; Aristilde, L.; Huson, D.H.; et al. An integrated systems biology approach reveals differences in formate metabolism in the genus Methanothermobacter. iScience 2023, 26, 108016. [Google Scholar] [CrossRef]
- Siriwach, R.; Matsuda, F.; Yano, K.; Hirai, M.Y. Drought stress responses in context-specific genome-scale metabolic models of Arabidopsis thaliana. Metabolites 2020, 10, 159. [Google Scholar] [CrossRef]
- Kamsen, R.; Kalapanulak, S.; Chiewchankaset, P.; Saithong, T. Transcriptome integrated metabolic modeling of carbon assimilation underlying storage root development in cassava. Sci. Rep. 2021, 11, 8758. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef]
- Caspi, R.; Billington, R.; Fulcher, C.A.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Latendresse, M.; Midford, P.E.; Ong, Q.; Ong, W.K.; et al. The MetaCyc database of metabolic pathways and enzymes. Nucleic Acids Res. 2018, 46, D633–D639. [Google Scholar] [CrossRef]
- Wang, H.; Marcišauskas, S.; Sánchez, B.J.; Domenzain, I.; Hermansson, D.; Agren, R.; Nielsen, J.; Kerkhoven, E.J. RAVEN 2.0: A versatile toolbox for metabolic network reconstruction and a case study on Streptomyces coelicolor. PLoS Comput. Biol. 2018, 14, e1006541. [Google Scholar] [CrossRef]
- Heirendt, L.; Arreckx, S.; Pfau, T.; Mendoza, S.N.; Richelle, A.; Heinken, A.; Haraldsdóttir, H.S.; Wachowiak, J.; Keating, S.M.; Vlasov, V.; et al. Creation and analysis of biochemical constraint-based models using the COBRA Toolbox v.3.0. Nat. Protoc. 2019, 14, 639–702. [Google Scholar] [CrossRef]
- Väremo, L.; Nielsen, J.; Nookaew, I. Enriching the gene set analysis of genome-wide data by incorporating directionality of gene expression and combining statistical hypotheses and methods. Nucleic Acids Res. 2013, 41, 4378–4391. [Google Scholar] [CrossRef]
- Park, J.P.; Kim, S.W.; Hwang, H.J.; Yun, J.W. Optimization of submerged culture conditions for the mycelial growth and exo-biopolymer production by Cordyceps militaris. Lett. Appl. Microbiol. 2001, 33, 76–81. [Google Scholar] [CrossRef]
- Suparmin, A.; Kato, T.; Dohra, H.; Park, E.Y. Insight into cordycepin biosynthesis of Cordyceps militaris: Comparison between a liquid surface culture and a submerged culture through transcriptomic analysis. PLoS ONE 2017, 12, e0187052. [Google Scholar] [CrossRef] [PubMed]
- Puckett, S.; Trujillo, C.; Wang, Z.; Eoh, H.; Ioerger, T.R.; Krieger, I.; Sacchettini, J.; Schnappinger, D.; Rhee, K.Y.; Ehrt, S. Glyoxylate detoxification is an essential function of malate synthase required for carbon assimilation in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2017, 114, E2225–E2232. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Yoon, D.-H.; Shrestha, B.; Choi, H.-K.; Sung, G.H. Metabolomic profiling reveals enrichment of cordycepin in senescence process of Cordyceps militaris fruit bodies. J. Microbiol. 2019, 57, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Guo, M.; Yang, H.; Guo, S.; Dong, C. The blue-light receptor CmWC-1 mediates fruit body development and secondary metabolism in Cordyceps militaris. Appl. Microbiol. Biotechnol. 2016, 100, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Wang, Z.; Lu, D.; Kang, H.; Li, G.; Guo, R.; Zhao, Y.; Han, R.; Ji, B.; Zeng, Y. Neutral lipid content in lipid droplets: Potential biomarker of cordycepin accumulation in cordycepin-producing fungi. Molecules 2019, 24, 3363. [Google Scholar] [CrossRef] [PubMed]
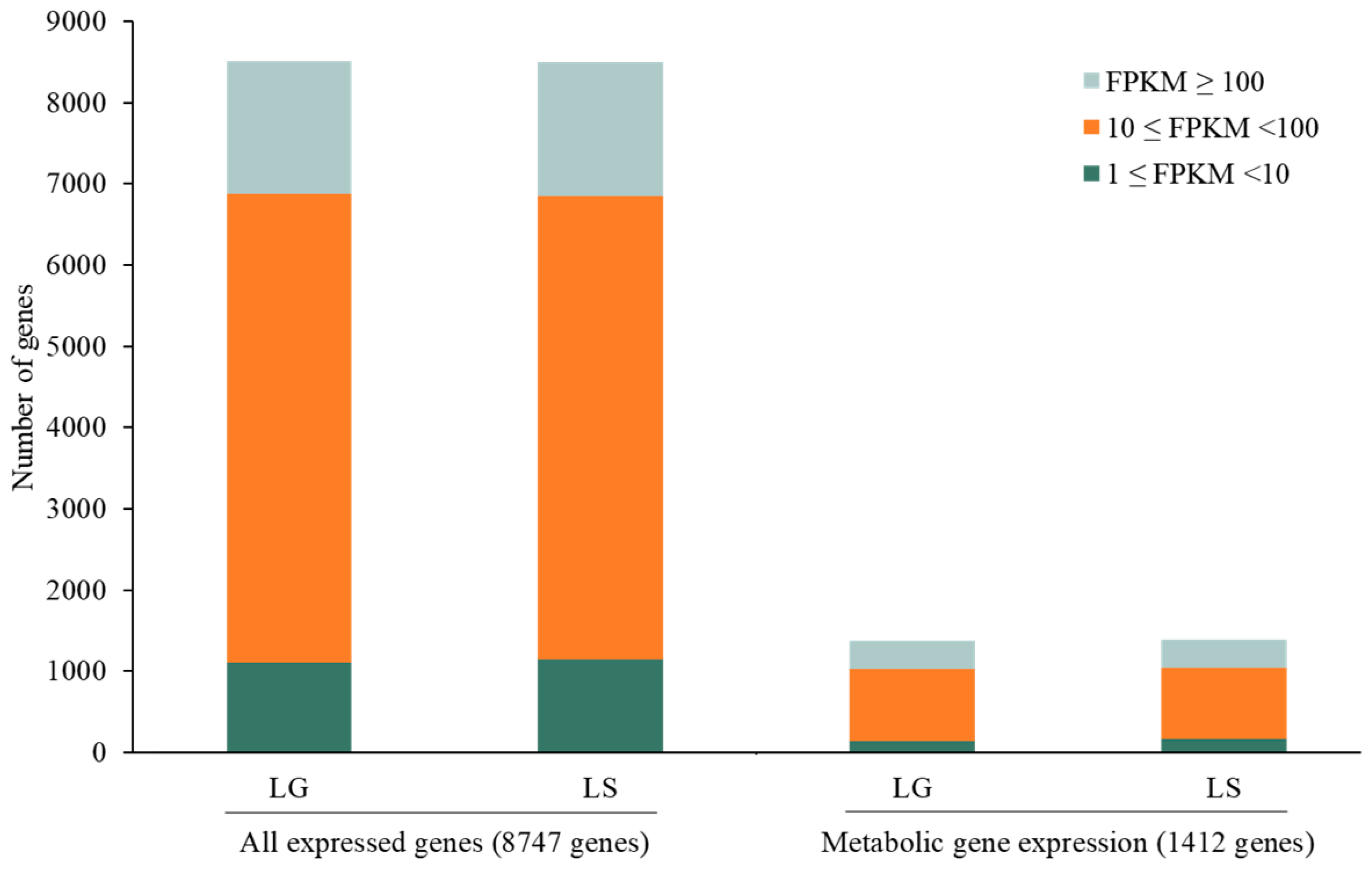

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Characteristics | iNR1329 1 | This Study iPS1474 |
|---|---|---|
| Total genes | 1329 | 1474 |
| Total metabolites | 1171 | 1699 |
| Total metabolic reactions | 1821 | 1916 |
| - Enzymatic reactions | 1391 | 1412 |
| - Non-enzymatic reactions | 430 | 504 |
| Spontaneous reactions | 21 | 21 |
| Transport reactions | 271 | 336 |
| Exchange reactions | 137 | 146 |
| Biomass reaction | 1 | 1 |
| Carotenoid biosynthesis | No | Yes |
| Pathway | Reporter Metabolites | Sucrose Up-Directional p-Value | Glucose Up-Directional p-Value |
|---|---|---|---|
| N-Glycan biosynthesis | Dolichyl phosphate | - | 0.001996 |
| Dolichyl-d-glucosyl phosphate | - | 0.001996 | |
| N-glycan (g10599) | - | 0.001996 | |
| D-glc-alpha-(1->2) | - | 0.001996 | |
| D-glc-alpha-(1->3) | - | 0.001996 | |
| N-glycan (g00008) | - | 0.001996 | |
| Dolichyl-beta-d-glucosyl phosphate | - | 0.003992 | |
| N-glycan (g10598) | - | 0.005988 | |
| Aminoacyl tRNA biosynthesis | L-lysyl-tRNA(lys) | 0.001996 | - |
| Cysteine and methionine metabolism | Protein n (omega)-methyl-l-arginine | 0.001996 | - |
| Oxidative phosphorylation | 8H (+) (energy metabolism) | 0.003992 | - |
| Phenylalanine, tyrosine and tryptophan biosynthesis | Shikimate3-phosphate | - | 0.005988 |
| Shikimate | - | 0.005988 | |
| 5-o-(1-carboxyvinyl)-3-phosphoshikimate | - | 0.007984 | |
| Carotenoid biosynthesis | Geranylgeranyl diphosphate (GGPP) | - | 0.005988 |
| Pyrimidine metabolism | 3-hydroxypropanoate | 0.005988 | - |
| 3-oxopropanoate | 0.005988 | - | |
| Lipid metabolism | 6,7-dihydropteridine | 0.005988 | - |
| Oxidized glutathione | 0.005988 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soommat, P.; Raethong, N.; Ruengsang, R.; Thananusak, R.; Laomettachit, T.; Laoteng, K.; Saithong, T.; Vongsangnak, W. Light-Exposed Metabolic Responses of Cordyceps militaris through Transcriptome-Integrated Genome-Scale Modeling. Biology 2024, 13, 139. https://doi.org/10.3390/biology13030139
Soommat P, Raethong N, Ruengsang R, Thananusak R, Laomettachit T, Laoteng K, Saithong T, Vongsangnak W. Light-Exposed Metabolic Responses of Cordyceps militaris through Transcriptome-Integrated Genome-Scale Modeling. Biology. 2024; 13(3):139. https://doi.org/10.3390/biology13030139
Chicago/Turabian StyleSoommat, Panyawarin, Nachon Raethong, Ratchaprapa Ruengsang, Roypim Thananusak, Teeraphan Laomettachit, Kobkul Laoteng, Treenut Saithong, and Wanwipa Vongsangnak. 2024. "Light-Exposed Metabolic Responses of Cordyceps militaris through Transcriptome-Integrated Genome-Scale Modeling" Biology 13, no. 3: 139. https://doi.org/10.3390/biology13030139
APA StyleSoommat, P., Raethong, N., Ruengsang, R., Thananusak, R., Laomettachit, T., Laoteng, K., Saithong, T., & Vongsangnak, W. (2024). Light-Exposed Metabolic Responses of Cordyceps militaris through Transcriptome-Integrated Genome-Scale Modeling. Biology, 13(3), 139. https://doi.org/10.3390/biology13030139