
A Comparative Analysis of Milk Oligosaccharides via LC-MS: Globally Distributed Cattle Breeds and Native Northern Finncattle

 , , , ,
, , , , 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Milk Sample Collection
2.2. Oligosaccharide Extraction and Purification
2.3. Oligosaccharide Analysis by LC-QToF
2.4. Statistical Analysis
3. Results
3.1. Oligosaccharide Identification and Breed-Specific Library
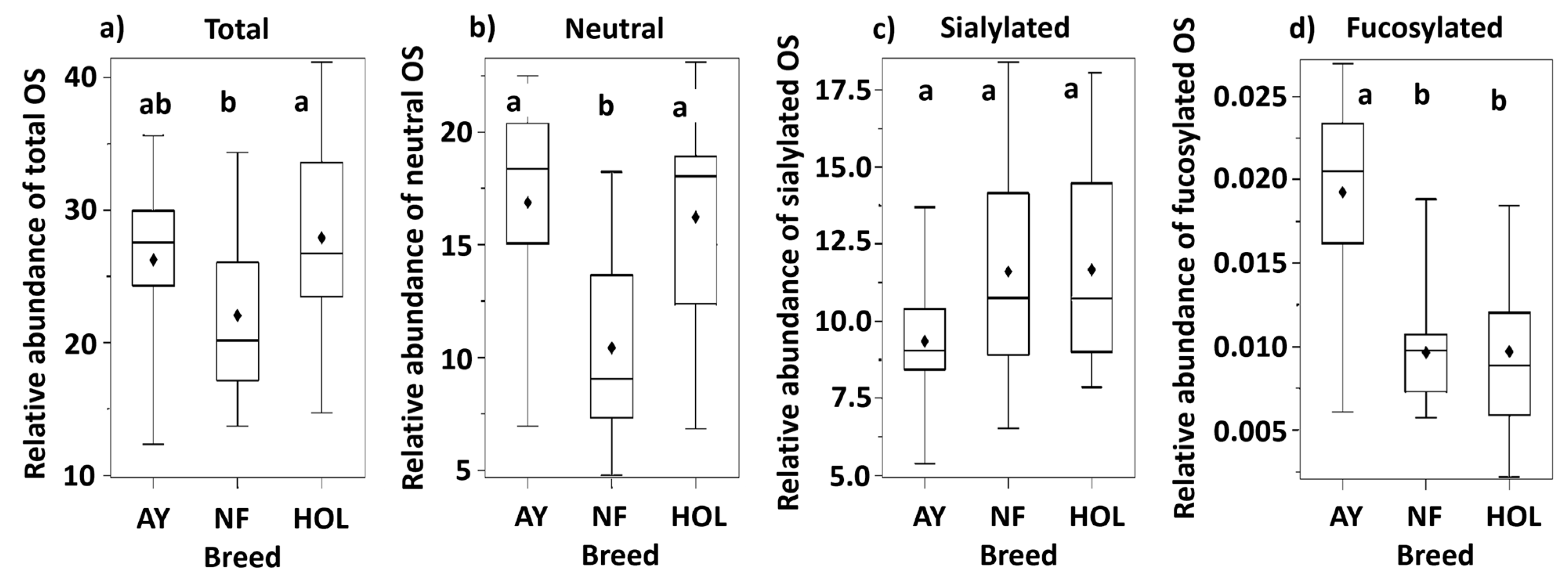
3.2. Relative Abundance
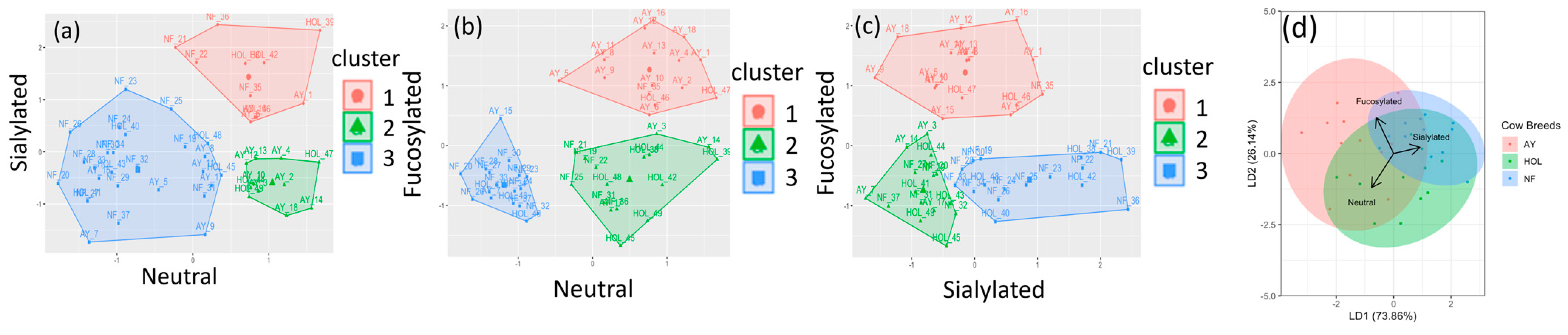
3.3. Clustering and Categorization Methods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roy, D.; Ye, A.; Moughan, P.J.; Singh, H. Composition, Structure, and Digestive Dynamics of Milk from Different Species—A Review. Front. Nutr. 2020, 7, 577759. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.C. Milk Nutritional Composition and Its Role in Human Health. Nutrition 2014, 30, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Ruhaak, L.R.; Lebrilla, C.B. Advances in Analysis of Human Milk Oligosaccharides. Adv. Nutr. 2012, 3, 406S–414S. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Davis, J.C.; Goonatilleke, E.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B. Absolute Quantitation of Human Milk Oligosaccharides Reveals Phenotypic Variations during Lactation. J. Nutr. 2017, 147, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Soyyılmaz, B.; Mikš, M.H.; Röhrig, C.H.; Matwiejuk, M.; Meszaros-Matwiejuk, A.; Vigsnæs, L.K. The Mean of Milk: A Review of Human Milk Oligosaccharide Concentrations throughout Lactation. Nutrients 2021, 13, 2737. [Google Scholar] [CrossRef]
- Bajic, D.; Wiens, F.; Wintergerst, E.; Deyaert, S.; Baudot, A.; Van Den Abbeele, P. HMOs Exert Marked Bifidogenic Effects on Children’s Gut Microbiota Ex Vivo, Due to Age-Related Bifidobacterium Species Composition. Nutrients 2023, 15, 1701. [Google Scholar] [CrossRef]
- Mills, D.A.; German, J.B.; Lebrilla, C.B.; Underwood, M.A. Translating Neonatal Microbiome Science into Commercial Innovation: Metabolism of Human Milk Oligosaccharides as a Basis for Probiotic Efficacy in Breast-Fed Infants. Gut Microbes 2023, 15, 2192458. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, Y.; Yang, Y.; Wu, S.; Zhao, J.; Li, Y.; Kang, X.; Li, Z.; Chen, J.; Shen, X.; et al. Bifidobacterium Infantis and 2′-Fucosyllactose Supplementation in Early Life May Have Potential Long-Term Benefits on Gut Microbiota, Intestinal Development, and Immune Function in Mice. J. Dairy Sci. 2023, 106, 7461–7476. [Google Scholar] [CrossRef]
- Rahman, T.; Sarwar, P.F.; Potter, C.; Comstock, S.S.; Klepac-Ceraj, V. Role of Human Milk Oligosaccharide Metabolizing Bacteria in the Development of Atopic Dermatitis/Eczema. Front. Pediatr. 2023, 11, 1090048. [Google Scholar] [CrossRef]
- Schalich, K.M.; Buendia, M.A.; Kaur, H.; Choksi, Y.A.; Washington, M.K.; Codreanu, G.S.; Sherrod, S.D.; McLean, J.A.; Peek, R.M.; Acra, S.A.; et al. A Human Milk Oligosaccharide Prevents Intestinal Inflammation in Adulthood via Modulating Gut Microbial Metabolism. mBio 2024, 15, e00298-24. [Google Scholar] [CrossRef]
- Manthey, C.F.; Autran, C.A.; Eckmann, L.; Bode, L. Human Milk Oligosaccharides Protect Against Enteropathogenic Escherichia Coli Attachment In Vitro and EPEC Colonization in Suckling Mice. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Morrow, A.L. Human milk glycans protect infants against enteric pathogens. Annu. Rev. Nutr. 2005, 25, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Chen, X. Human Milk Oligosaccharides (HMOS). In Advances in Carbohydrate Chemistry and Biochemistry; Elsevier: Amsterdam, The Netherlands, 2015; Volume 72, pp. 113–190. ISBN 978-0-12-802141-5. [Google Scholar]
- Moore, R.E.; Spicer, S.K.; Lu, J.; Chambers, S.A.; Noble, K.N.; Lochner, J.; Christofferson, R.C.; Vasco, K.A.; Manning, S.D.; Townsend, S.D.; et al. The Utility of Human Milk Oligosaccharides against Group B Streptococcus Infections of Reproductive Tissues and Cognate Adverse Pregnancy Outcomes. ACS Cent. Sci. 2023, 9, 1737–1749. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; McMath, A.L.; Donovan, S.M. Review on the Impact of Milk Oligosaccharides on the Brain and Neurocognitive Development in Early Life. Nutrients 2023, 15, 3743. [Google Scholar] [CrossRef]
- Rajhans, P.; Mainardi, F.; Austin, S.; Sprenger, N.; Deoni, S.; Hauser, J.; Schneider, N. The Role of Human Milk Oligosaccharides in Myelination, Socio-Emotional and Language Development: Observational Data from Breast-Fed Infants in the United States of America. Nutrients 2023, 15, 4624. [Google Scholar] [CrossRef]
- Tao, N.; DePeters, E.J.; Freeman, S.; German, J.B.; Grimm, R.; Lebrilla, C.B. Bovine Milk Glycome. J. Dairy Sci. 2008, 91, 3768–3778. [Google Scholar] [CrossRef]
- Robinson, R.C. Structures and Metabolic Properties of Bovine Milk Oligosaccharides and Their Potential in the Development of Novel Therapeutics. Front. Nutr. 2019, 6, 50. [Google Scholar] [CrossRef]
- Boudry, G.; Hamilton, M.K.; Chichlowski, M.; Wickramasinghe, S.; Barile, D.; Kalanetra, K.M.; Mills, D.A.; Raybould, H.E. Bovine Milk Oligosaccharides Decrease Gut Permeability and Improve Inflammation and Microbial Dysbiosis in Diet-Induced Obese Mice. J. Dairy Sci. 2017, 100, 2471–2481. [Google Scholar] [CrossRef]
- Zivkovic, A.M.; Barile, D. Bovine Milk as a Source of Functional Oligosaccharides for Improving Human Health. Adv. Nutr. 2011, 2, 284–289. [Google Scholar] [CrossRef]
- Quinn, E.M.; O’Callaghan, T.F.; Tobin, J.T.; Murphy, J.P.; Sugrue, K.; Slattery, H.; O’Donovan, M.; Hickey, R.M. Changes to the Oligosaccharide Profile of Bovine Milk at the Onset of Lactation. Dairy 2020, 1, 284–296. [Google Scholar] [CrossRef]
- Tao, N.; DePeters, E.J.; German, J.B.; Grimm, R.; Lebrilla, C.B. Variations in Bovine Milk Oligosaccharides during Early and Middle Lactation Stages Analyzed by High-Performance Liquid Chromatography-Chip/Mass Spectrometry. J. Dairy Sci. 2009, 92, 2991–3001. [Google Scholar] [CrossRef] [PubMed]
- Asakuma, S.; Ueda, Y.; Akiyama, F.; Uemura, Y.; Miyaji, M.; Nakamura, M.; Murai, M.; Urashima, T. Short Communication: Effect of Grazing on the Concentrations of Total Sialic Acid and Hexose in Bovine Milk. J. Dairy Sci. 2010, 93, 4850–4854. [Google Scholar] [CrossRef] [PubMed]
- Durham, S.D.; Lemay, D.G.; Wei, Z.; Kalscheur, K.F.; Finley, J.W.; Fukagawa, N.K.; Barile, D. Dietary Fiber to Starch Ratio Affects Bovine Milk Oligosaccharide Profiles. Curr. Dev. Nutr. 2022, 6, 6006004. [Google Scholar] [CrossRef] [PubMed]
- Sundekilde, U.K.; Barile, D.; Meyrand, M.; Poulsen, N.A.; Larsen, L.B.; Lebrilla, C.B.; German, J.B.; Bertram, H.C. Natural Variability in Bovine Milk Oligosaccharides from Danish Jersey and Holstein-Friesian Breeds. J. Agric. Food Chem. 2012, 60, 6188–6196. [Google Scholar] [CrossRef]
- Robinson, R.C.; Poulsen, N.A.; Colet, E.; Duchene, C.; Larsen, L.B.; Barile, D. Profiling of aminoxyTMT-Labeled Bovine Milk Oligosaccharides Reveals Substantial Variation in Oligosaccharide Abundance between Dairy Cattle Breeds. Sci Rep 2019, 9, 5465. [Google Scholar] [CrossRef]
- Sunds, A.V.; Bunyatratchata, A.; Robinson, R.; Glantz, M.; Paulsson, M.; Leskauskaite, D.; Pihlanto, A.; Inglingstad, R.; Devold, T.G.; Vegarud, G.E.; et al. Comparison of Bovine Milk Oligosaccharides in Native North European Cattle Breeds. Int. Dairy J. 2021, 114, 104917. [Google Scholar] [CrossRef]
- Pokharel, K.; Weldenegodguad, M.; Popov, R.; Honkatukia, M.; Huuki, H.; Lindeberg, H.; Peippo, J.; Reilas, T.; Zarovnyaev, S.; Kantanen, J. Whole Blood Transcriptome Analysis Reveals Footprints of Cattle Adaptation to Sub-arctic Conditions. Anim Genet 2019, 50, 217–227. [Google Scholar] [CrossRef]
- Soini, K.; Pouta, E.; Latvala, T.; Lilja, T. Agrobiodiversity Products in Alternative Food System: Case of Finnish Native Cattle Breeds. Sustainability 2019, 11, 3408. [Google Scholar] [CrossRef]
- Mazzullo, N.; Soppela, P. Reindeer, Cows and People: Sustainable Human–Animal Adaptations in Finnish Lapland. In Living and Working with Snow, Ice and Seasons in the Modern Arctic: Everyday Perspectives; Strauss-Mazzullo, H., Tennberg, M., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 59–83. ISBN 978-3-031-36445-7. [Google Scholar]
- Genes in Imprisonment—NordGen. Available online: https://www.nordgen.org/sv/nyheter/genes-in-imprisonment-1/ (accessed on 18 July 2024).
- Tapio, I.; Värv, S.; Bennewitz, J.; Maleviciute, J.; Fimland, E.; Grislis, Z.; Meuwissen, T.H.E.; Miceikiene, I.; Olsaker, I.; Viinalass, H.; et al. Prioritization for Conservation of Northern European Cattle Breeds Based on Analysis of Microsatellite Data. Conserv. Biol. 2006, 20, 1768–1779. [Google Scholar] [CrossRef]
- Soppela, P.; Tuomivaara, A. How Specialization of Products and Services Based on the Lapland Cattle Can Contribute to Northern Livelihoods. Available online: https://www.arcticcentre.org/FI/Lappari/Lapland-Cattle (accessed on 26 September 2024).
- Robinson, R.C.; Colet, E.; Tian, T.; Poulsen, N.A.; Barile, D. An Improved Method for the Purification of Milk Oligosaccharides by Graphitised Carbon-Solid Phase Extraction. Int. Dairy J. 2018, 80, 62–68. [Google Scholar] [CrossRef]
- Mehra, R.; Barile, D.; Marotta, M.; Lebrilla, C.B.; Chu, C.; German, J.B. Novel High-Molecular Weight Fucosylated Milk Oligosaccharides Identified in Dairy Streams. PLoS ONE 2014, 9, e96040. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-Q.; Treves, G.; Amicucci, M.; Guerrero, A.; Xu, G.; Gong, T.-Q.; Davis, J.; Park, D.; Galermo, A.; Wu, L.; et al. GlycoNote with Iterative Decoy Searching and Open-Search Component Analysis for High-Throughput and Reliable Glycan Spectral Interpretation. Anal. Chem. 2023, 95, 8223–8231. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.C.; Poulsen, N.A.; Barile, D. Multiplexed Bovine Milk Oligosaccharide Analysis with Aminoxy Tandem Mass Tags. PLoS ONE 2018, 13, e0196513. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://faba.fi/fi/lypsykarja/ayrshire (accessed on 26 September 2024).
- Soppela, P.; Tuomivaara, A. (Eds.) The Possibilities of Specialization Based on the Lapland Cattle Cow as Part of Northern Livelihoods: Final Report of the Lapland Cattle Livelihood Project (In Finnish, Abstract in English). Arctic Centre Reports. 2023, Volume 65, pp. 1–71. Available online: https://urn.fi/URN:ISBN:978-952-337-404-1 (accessed on 26 September 2024).
- Soppela, P. Looking to Lapland’s Past for a Sustainable Food Source. The Circle: WWF Magazine, 1 January 2018; 18–20. [Google Scholar]
- Liu, Z.; Wang, T.; Pryce, J.E.; MacLeod, I.M.; Hayes, B.J.; Chamberlain, A.J.; Jagt, C.V.; Reich, C.M.; Mason, B.A.; Rochfort, S.; et al. Fine-Mapping Sequence Mutations with a Major Effect on Oligosaccharide Content in Bovine Milk. Sci. Rep. 2019, 9, 2137. [Google Scholar] [CrossRef] [PubMed]
- Kantanen, J. Genetic Diversity and Population Structure of 20 North European Cattle Breeds. J. Hered. 2000, 91, 446–457. [Google Scholar] [CrossRef]
- Poulsen, N.A.; Robinson, R.C.; Barile, D.; Larsen, L.B.; Buitenhuis, B. A Genome-Wide Association Study Reveals Specific Transferases as Candidate Loci for Bovine Milk Oligosaccharides Synthesis. BMC Genom. 2019, 20, 404. [Google Scholar] [CrossRef]
- Jacobi, S.K.; Yatsunenko, T.; Li, D.; Dasgupta, S.; Yu, R.K.; Berg, B.M.; Chichlowski, M.; Odle, J. Dietary Isomers of Sialyllactose Increase Ganglioside Sialic Acid Concentrations in the Corpus Callosum and Cerebellum and Modulate the Colonic Microbiota of Formula-Fed Piglets. J. Nutr. 2016, 146, 200–208. [Google Scholar] [CrossRef]
- Oliveros, E.; Vázquez, E.; Barranco, A.; Ramírez, M.; Gruart, A.; Delgado-García, J.; Buck, R.; Rueda, R.; Martín, M. Sialic Acid and Sialylated Oligosaccharide Supplementation during Lactation Improves Learning and Memory in Rats. Nutrients 2018, 10, 1519. [Google Scholar] [CrossRef]
- Galuska, C.E.; Rudloff, S.; Kuntz, S.; Borsch, C.; Reutzel, M.; Eckert, G.; Galuska, S.P.; Kunz, C. Metabolic Fate and Organ Distribution of 13C-3′-Sialyllactose and 13C-N-Acetylneuraminic Acid in Wild-Type Mice—No Evidence for Direct Incorporation into the Brain. J. Funct. Foods 2020, 75, 104268. [Google Scholar] [CrossRef]
- Charbonneau, M.R.; O’Donnell, D.; Blanton, L.V.; Totten, S.M.; Davis, J.C.C.; Barratt, M.J.; Cheng, J.; Guruge, J.; Talcott, M.; Bain, J.R.; et al. Sialylated Milk Oligosaccharides Promote Microbiota-Dependent Growth in Models of Infant Undernutrition. Cell 2016, 164, 859–871. [Google Scholar] [CrossRef]
- Jahan, M.; Francis, N.; Wynn, P.; Wang, B. The Potential for Sialic Acid and Sialylated Glycoconjugates as Feed Additives to Enhance Pig Health and Production. Animals 2021, 11, 2318. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, M.; Jahan, M.; Ghorashi, S.A.; Wang, B. Current Perspective of Sialylated Milk Oligosaccharides in Mammalian Milk: Implications for Brain and Gut Health of Newborns. Foods 2021, 10, 473. [Google Scholar] [CrossRef] [PubMed]
- Fischer-Tlustos, A.J.; Hertogs, K.; Van Niekerk, J.K.; Nagorske, M.; Haines, D.M.; Steele, M.A. Oligosaccharide Concentrations in Colostrum, Transition Milk, and Mature Milk of Primi- and Multiparous Holstein Cows during the First Week of Lactation. J. Dairy Sci. 2020, 103, 3683–3695. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Kleinschmidt, T. Synthesis of Galactooligosaccharides in Milk and Whey: A Review: Synthesis of Galactooligosaccharides…. Compr. Rev. Food Sci. Food Saf. 2018, 17, 678–697. [Google Scholar] [CrossRef]
- Gray, T.E.; Narayana, K.; Garner, A.M.; Bakker, S.A.; Yoo, R.K.H.; Fischer-Tlustos, A.J.; Steele, M.A.; Zandberg, W.F. Analysis of the Biosynthetic Flux in Bovine Milk Oligosaccharides Reveals Competition between Sulfated and Sialylated Species and the Existence of Glucuronic Acid-Containing Analogues. Food Chem. 2021, 361, 130143. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OS ID | Composition | Monoisotopic Mass | Northern Finncattle | Ayrshire | Holstein |
|---|---|---|---|---|---|
| Number of Isomers | |||||
| 1 | 3_0_0_0_0 | 504.1690 | 5 | 5 | 5 |
| 2 | 2_1_0_0_0 | 545.1956 | 4 | 4 | 4 |
| 3 | 2_0_0_1_0 | 633.2116 | 2 | 2 | 2 |
| 4 | 4_0_0_0_0 | 666.2219 | 4 | 4 | 4 |
| 5 | 1_1_0_1_0 | 674.2382 | 1 | 1 | 1 |
| 6 | 3_1_0_0_0 | 707.2484 | 4 | 4 | 4 |
| 7 | 2_2_0_0_0 | 748.2750 | 3 | 3 | 3 |
| 8 | 3_0_0_1_0 | 795.2645 | 2 | 2 | 2 |
| 9 | 5_0_0_0_0 | 828.2747 | 3 | 3 | 3 |
| 10 | 2_1_0_1_0 | 836.2910 | 1 | 1 | 1 |
| 11 | 4_1_0_0_0 | 869.3012 | 3 | 2 | 2 |
| 12 | 3_2_0_0_0 | 910.3278 | 2 | 2 | 2 |
| 13 | 2_0_0_2_0 | 924.3070 | 1 | 1 | 1 |
| 14 | 6_0_0_0_0 | 990.3275 | 3 | 3 | 3 |
| 15 | 3_1_0_1_0 | 998.3438 | 1 | 1 | 1 |
| 16 | 4_2_0_0_0 | 1072.3806 | 2 | 2 | 2 |
| 17 | 3_0_0_2_0 | 1086.3599 | 1 | 1 | 1 |
| 18 | 3_3_0_0_0 | 1113.4072 | 1 | 1 | 1 |
| 19 | 2_1_0_2_0 | 1127.3864 | 1 | 1 | 1 |
| 20 | 4_1_0_1_0 | 1160.3967 | 1 | 1 | 1 |
| 21 | 4_2_0_1_0 | 1363.4760 | 1 | 1 | 1 |
| 22 | 5_4_0_0_0 | 1640.5922 | 1 | 1 | 1 |
| 23 | 5_4_1_0_0 | 1786.6501 | 1 | 1 | 1 |
| 24 | 3_6_1_0_0 | 1868.7032 | 1 | 1 | 1 |
| 25 | 5_4_1_1_0 | 2077.7455 | 1 | 1 | 1 |
| Number of isomers | 50 | 49 | 49 | ||
| AY 1 | 95% CI 2 | NF1 | 95% CI 2 | HOL 1 | 95% CI 2 | p-Value | |
|---|---|---|---|---|---|---|---|
| Neutral oligosaccharides | |||||||
| 3_0_0_0_0 | 2.9 a | 2.6, 3.1 | 2.4 b | 2.2, 2.6 | 2.8 ab | 2.5, 3.1 | 0.019 |
| 2_1_0_0_0 | 9.0 a | 7.2, 10.9 | 4.4 b | 2.6, 6.2 | 9.5 a | 7.2, 11.7 | <0.001 |
| 4_0_0_0_0 | 3.5 | 2.8, 4.3 | 2.4 | 1.7, 3.1 | 2.8 | 1.9, 3.7 | 0.080 |
| 3_1_0_0_0 | 0.44 a | 0.39, 0.49 | 0.35 b | 0.30, 0.40 | 0.36 ab | 0.30, 0.43 | 0.028 |
| 2_2_0_0_0 | 0.087 | 0.073, 0.101 | 0.079 | 0.066, 0.093 | 0.100 | 0.083, 0.117 | 0.173 |
| 5_0_0_0_0 3 | 0.024 | 0.018, 0.027 | 0.020 | 0.015, 0.022 | 0.021 | 0.015, 0.025 | 0.312 |
| 4_1_0_0_0 3 | 0.36 ab | 0.21, 0.37 | 0.34 a | 0.24, 0.41 | 0.21 b | 0.13, 0.26 | 0.047 |
| 3_2_0_0_0 3 | 0.054 a | 0.033, 0.058 | 0.048 a | 0.031, 0.054 | 0.020 b | 0.012, 0.024 | <0.001 |
| 6_0_0_0_0 | 0.35 a | 0.30, 0.39 | 0.25 b | 0.20, 0.29 | 0.34 ab | 0.28, 0.39 | 0.008 |
| 4_2_0_0_0 | 0.050 | 0.040, 0.060 | 0.037 | 0.028, 0.047 | 0.034 | 0.022, 0.046 | 0.094 |
| 3_3_0_0_0 3 | 0.032 | 0.023, 0.043 | 0.025 | 0.019, 0.034 | 0.022 | 0.016, 0.032 | 0.322 |
| 5_4_0_0_0 | 0.008 | 0.006, 0.010 | 0.009 | 0.007, 0.012 | 0.409 | ||
| Neutral fucosylated oligosaccharides | |||||||
| 5_4_1_0_0 | 0.012 | 0.010, 0.014 | 0.010 | 0.008, 0.011 | 0.010 | 0.007, 0.012 | 0.183 |
| 3_6_1_0_0 | 0.007 | 0.006, 0.009 | |||||
| Fucosylated and sialylated oligosaccharides | |||||||
| 5_4_1_1_0 | 0.005 | 0.005, 0.006 | |||||
| Sialylated oligosaccharides | |||||||
| 2_0_0_1_0 (α-2,6 isomer) | 1.6 | 1.2, 2.0 | 2.0 | 1.6, 2.4 | 2.0 | 1.5, 2.5 | 0.195 |
| 2_0_0_1_0 (α-2,3 isomer) | 4.3 | 3.8, 4.9 | 4.8 | 4.3, 5.3 | 4.5 | 3.8, 5.2 | 0.477 |
| 1_1_0_1_0 3 | 0.074 | 0.045, 0.087 | 0.112 | 0.057, 0.108 | 0.108 | 0.06, 0.134 | 0.348 |
| 3_0_0_1_0 | 0.64 | 0.56, 0.72 | 0.52 | 0.44, 0.60 | 0.58 | 0.48, 0.68 | 0.109 |
| 2_1_0_1_0 3 | 0.032 b | 0.023, 0.039 | 0.014 c | 0.009, 0.016 | 0.092 a | 0.045, 0.089 | <0.001 |
| 2_0_0_2_0 | 2.2 b | 1.8, 2.5 | 3.6 a | 3.3, 4.0 | 3.5 a | 3.0, 3.9 | <0.001 |
| 3_1_0_1_0 | 0.021 | 0.016, 0.026 | 0.025 | 0.019, 0.031 | 0.238 | ||
| 3_0_0_2_0 3 | 0.425 b | 0.236, 0.428 | 0.396 ab | 0.266, 0.473 | 0.656 a | 0.405, 0.837 | 0.035 |
| 4_1_0_1_0 | 0.073 | 0.053, 0.092 | 0.081 | 0.062, 0.100 | 0.047 | 0.023, 0.071 | 0.080 |
| 4_2_0_1_0 3 | 0.015 | 0.012, 0.021 | 0.022 | 0.017, 0.030 | 0.017 | 0.012, 0,024 | 0.165 |
| 2_1_0_2_0 | 0.13 | 0.08, 0.18 | |||||
| Total oligosaccharides | 26.2 ab | 23.2, 29.3 | 22.0 b | 19.1, 25.0 | 27.9 a | 24.2, 31.6 | 0.035 |
| Neutral oligosaccharides | 16.9 a | 14.8, 19.0 | 10.4 b | 8.4, 12.5 | 16.2 a | 13.6, 18.8 | <0.001 |
| Sialylated oligosaccharides 3 | 9.1 | 8.0, 10.3 | 11.2 | 9.9, 12.7 | 11.2 | 9.6, 13.1 | 0.042 |
| Fucosylated oligosaccharides | 0.019 a | 0.017, 0.022 | 0.010 b | 0.007, 0.012 | 0.010 b | 0.007, 0.012 | <0.001 |
| Neutral/Total oligosaccharides | 63.6 a | 59.8, 67.4 | 46.7 b | 43.0, 50.4 | 57.3 a | 52.6, 61.9 | <0.001 |
| Sialylated/Total oligosaccharides | 36.3 b | 32.5, 40.1 | 53.3 a | 49.6, 57.0 | 42.7 b | 38.0, 47.3 | <0.001 |
| Fucosylated/Total oligosaccharides | 0.074 a | 0.066, 0.082 | 0.046 b | 0.038, 0.054 | 0.035 b | 0.025, 0.045 | <0.001 |
| Neutral vs. Sialylated | AY | NF | HOL | Neutral vs. Fucosylated | AY | NF | HOL | Sialylated vs. Fucosylated | AY | NF | HOL |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cluster | n = 18 | n = 19 | n = 12 | Cluster | n = 18 | n = 19 | n = 12 | Cluster | n = 18 | n = 19 | n = 12 |
| 1 | 0.17 | 0.21 | 0.33 | 1 | 0.72 | 0.05 | 0.17 | 1 | 0.78 | 0.05 | 0.17 |
| 2 | 0.44 | 0 | 0.25 | 2 | 0.17 | 0.32 | 0.58 | 2 | 0.22 | 0.32 | 0.42 |
| 3 | 0.39 | 0.79 | 0.42 | 3 | 0.11 | 0.63 | 0.25 | 3 | 0 | 0.63 | 0.42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Huang, Y.-P.; Rogers, M.; Leskinen, H.; Soppela, P.; Tuomivaara, A.; Hyvönen, J.; Barile, D. A Comparative Analysis of Milk Oligosaccharides via LC-MS: Globally Distributed Cattle Breeds and Native Northern Finncattle. Biology 2024, 13, 855. https://doi.org/10.3390/biology13110855
Wang Y, Huang Y-P, Rogers M, Leskinen H, Soppela P, Tuomivaara A, Hyvönen J, Barile D. A Comparative Analysis of Milk Oligosaccharides via LC-MS: Globally Distributed Cattle Breeds and Native Northern Finncattle. Biology. 2024; 13(11):855. https://doi.org/10.3390/biology13110855
Chicago/Turabian StyleWang, Yu, Yu-Ping Huang, Mana Rogers, Heidi Leskinen, Päivi Soppela, Anne Tuomivaara, Juha Hyvönen, and Daniela Barile. 2024. "A Comparative Analysis of Milk Oligosaccharides via LC-MS: Globally Distributed Cattle Breeds and Native Northern Finncattle" Biology 13, no. 11: 855. https://doi.org/10.3390/biology13110855
APA StyleWang, Y., Huang, Y.-P., Rogers, M., Leskinen, H., Soppela, P., Tuomivaara, A., Hyvönen, J., & Barile, D. (2024). A Comparative Analysis of Milk Oligosaccharides via LC-MS: Globally Distributed Cattle Breeds and Native Northern Finncattle. Biology, 13(11), 855. https://doi.org/10.3390/biology13110855