

Blue Light and Temperature Actigraphy Measures Predicting Metabolic Health Are Linked to Melatonin Receptor Polymorphism

 ,
,  , , ,
, , , 
 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
We dedicate this paper to the memory of our dear colleague, Dr. Konstantin Danilenko, who helped initiate this project but sadly passed away on 18 January 2023.
2. Materials and Methods
2.1. Subjects and Data Collection
2.2. Actigraphy
2.3. Biochemical Assessment
2.4. Genotyping
2.5. Data and Statistical Analyses
3. Results
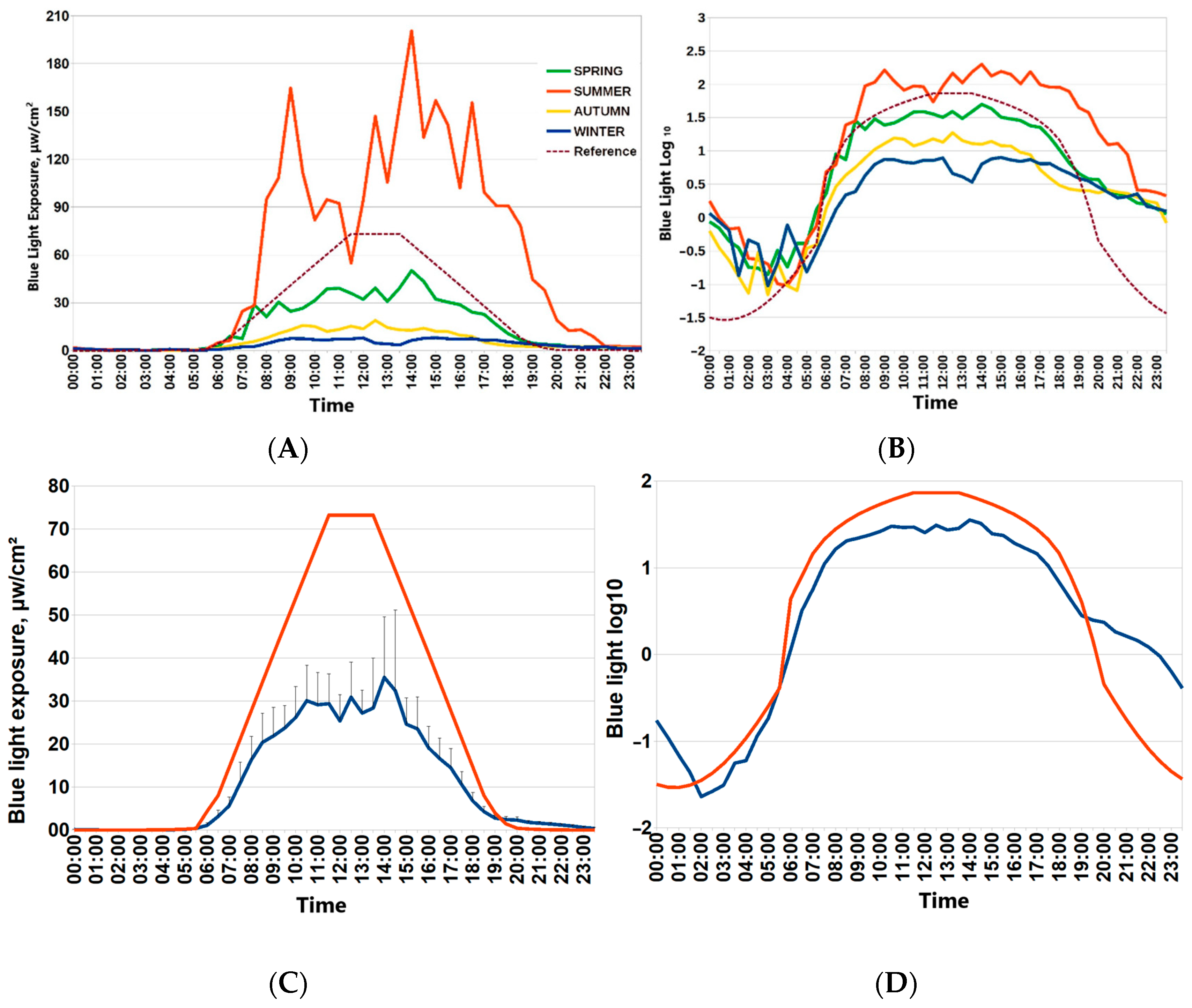
3.1. Seasonal Features of Light Exposure in the Arctic Spring as a Season with the Most Favorable Circadian Light Environment
3.2. General Characteristics of Participants of the Spring Equinox Session
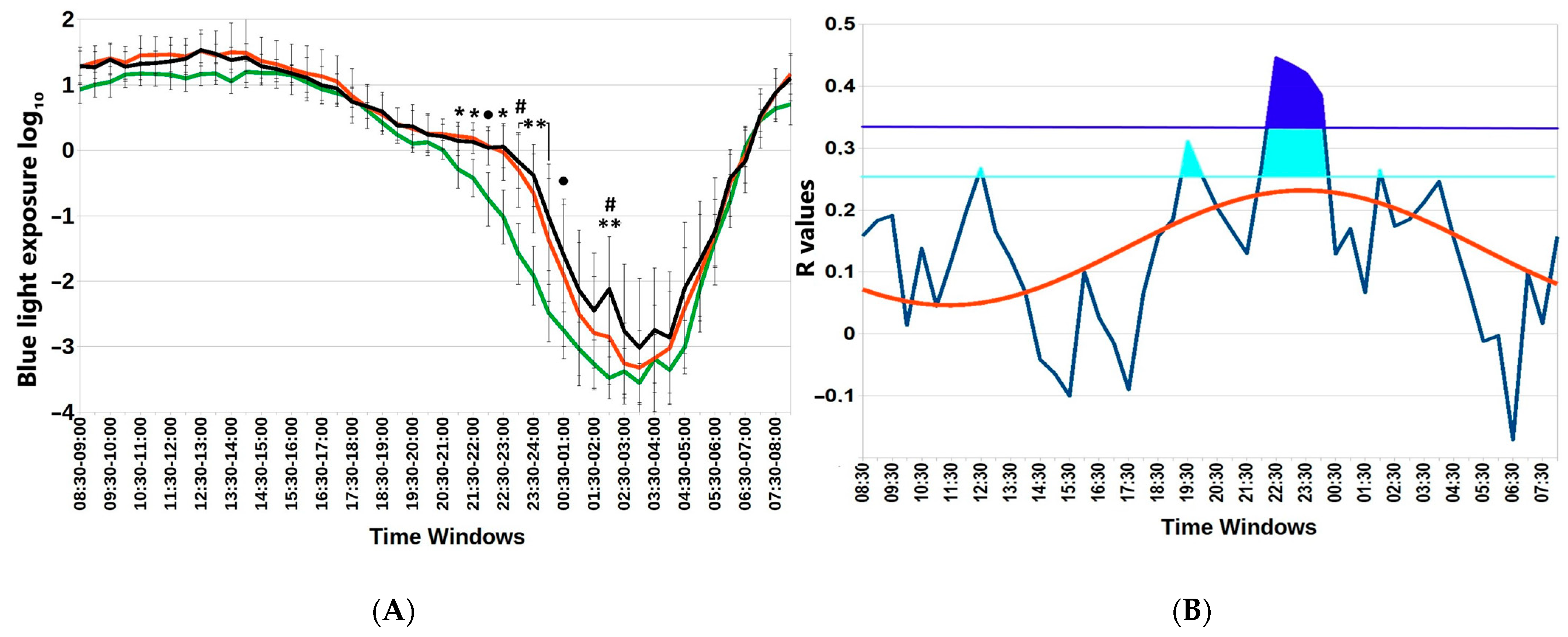
3.3. Blue Light Nocturnal Excess and Lower Wrist Temperature Predict Body Mass Index
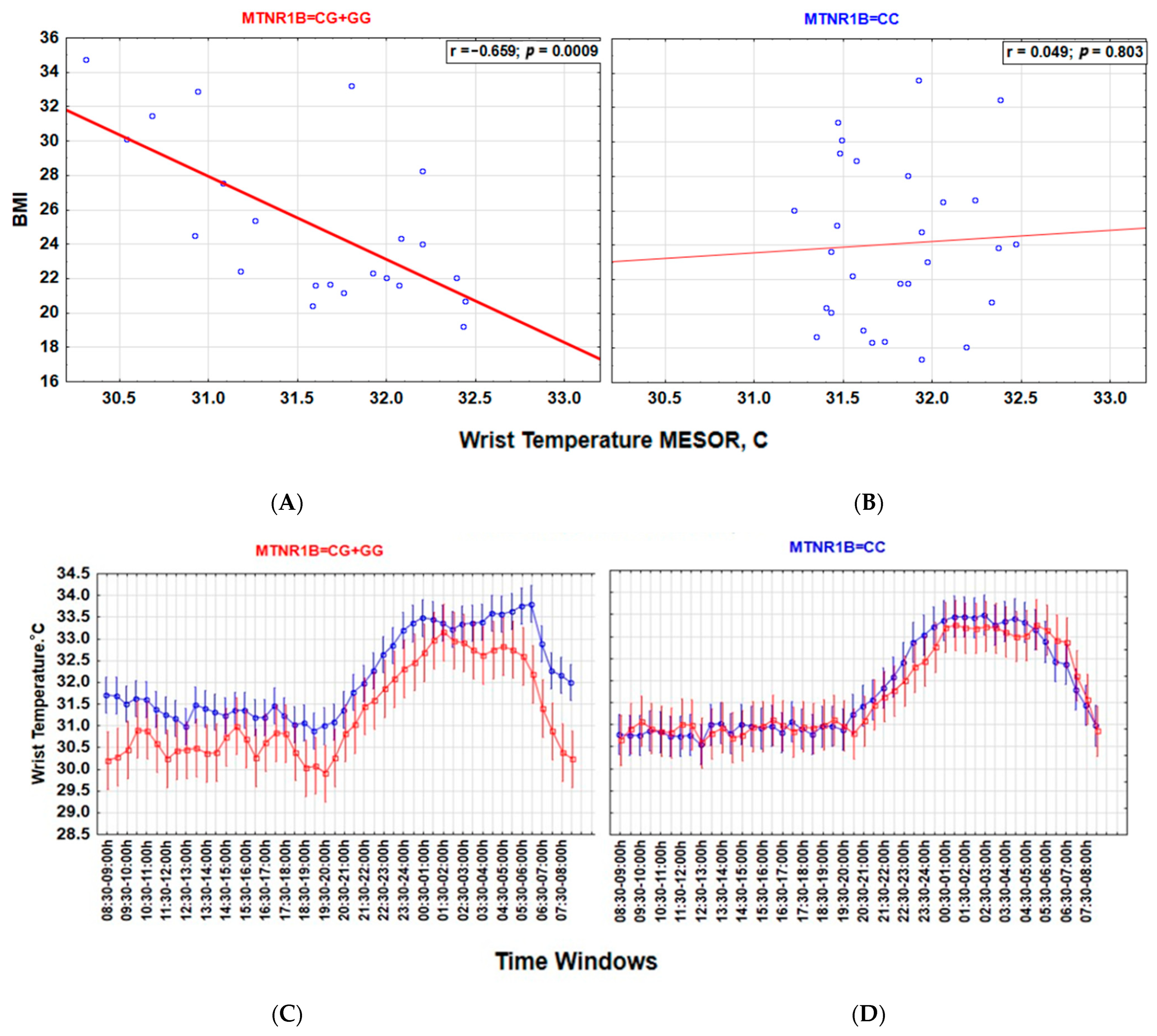
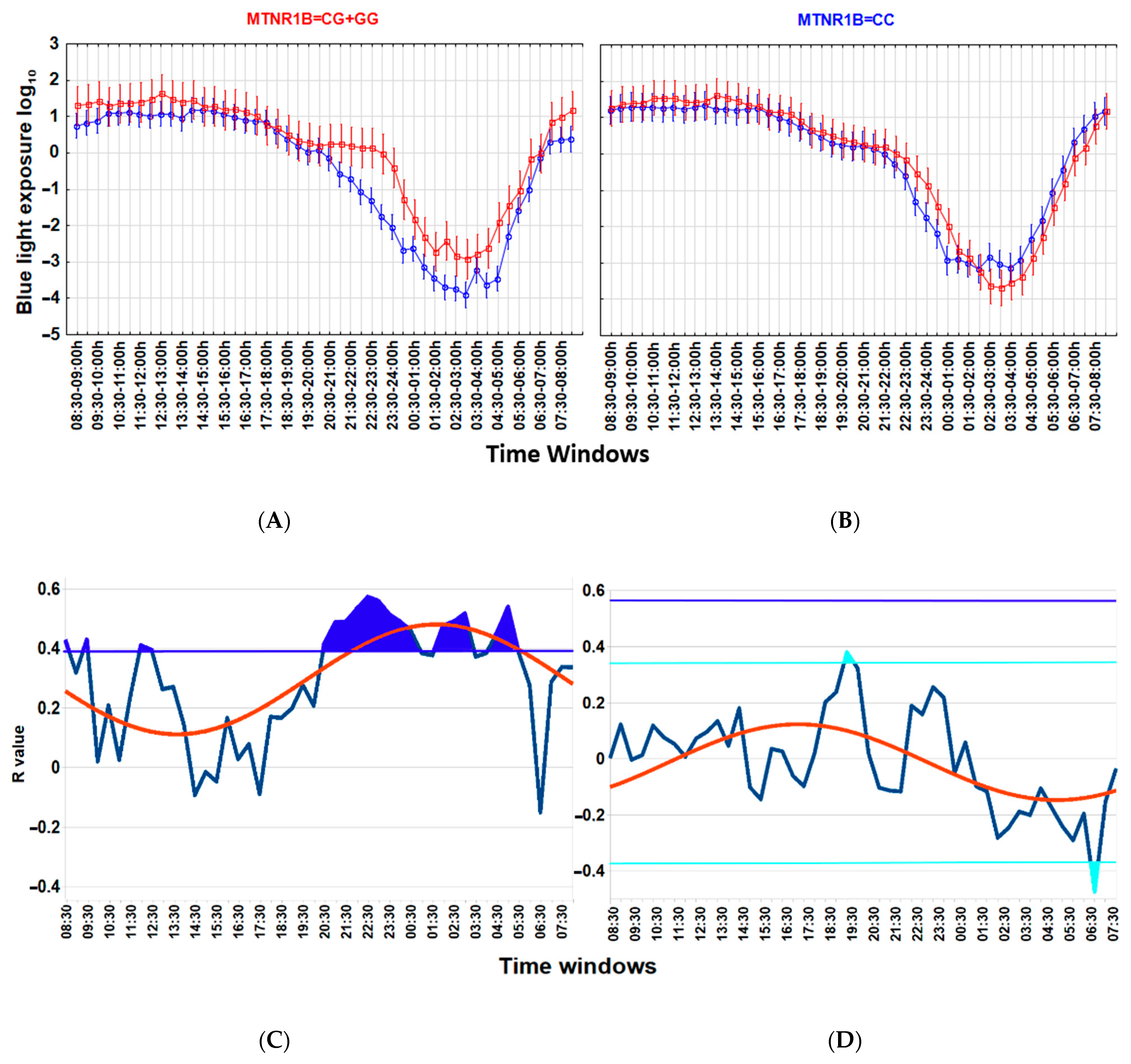
3.4. MTNR1B Polymorphism Accounts for the Interaction between Light, Wrist Temperature and Metabolic Health
3.5. Associations of Leptin and Cortisol with Actimetry-Derived Indices
3.6. Age-, Sex- and Indigeneity-Related Aspects of Actigraphy-Based Indices
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- LeGates, T.A.; Fernandez, D.C.; Hattar, S. Light as a central modulator of circadian rhythms, sleep and affect. Nat. Rev. Neurosci. 2014, 15, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Blume, C.; Garbazza, C.; Spitschan, M. Effects of light on human circadian rhythms, sleep and mood. Somnologie 2019, 23, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Wirz-Justice, A.; Skene, D.J.; Münch, M. The relevance of daylight for humans. Biochem. Pharmacol. 2021, 191, 114304. [Google Scholar] [CrossRef]
- Roenneberg, T.; Kantermann, T.; Juda, M.; Vetter, C.; Allebrandt, K.V. Light and the human circadian clock. Handb. Exp. Pharmacol. 2013, 217, 311–331. [Google Scholar]
- Husse, J.; Eichele, G.; Oster, H. Synchronization of the mammalian circadian timing system: Light can control peripheral clocks independently of the SCN clock: Alternate routes of entrainment optimize the alignment of the body’s circadian clock network with external time. BioEssays 2015, 37, 1119–1128. [Google Scholar] [CrossRef]
- Bano-Otalora, B.; Martial, F.; Harding, C.; Bechtold, D.A.; Allen, A.E.; Brown, T.M.; Belle, M.D.; Lucas, R.J. Bright daytime light enhances circadian amplitude in a diurnal mammal. Proc. Natl. Acad. Sci. USA 2021, 118, e2100094118. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.M.; Brainard, G.C.; Cajochen, C.; Czeisler, C.A.; Hanifin, J.P.; Lockley, S.W.; Lucas, R.J.; Münch, M.; O’Hagan, J.B.; Peirson, S.N.; et al. Recommendations for daytime, evening, and nighttime indoor light exposure to best support physiology, sleep, and wakefulness in healthy adults. PLoS Biol. 2022, 20, e3001571. [Google Scholar] [CrossRef]
- Vollmer, C.; Michel, U.; Randler, C. Outdoor light at night (LAN) is correlated with eveningness in adolescents. Chronobiol. Int. 2012, 29, 502–508. [Google Scholar] [CrossRef]
- Wright, K.P., Jr.; McHill, A.W.; Birks, B.R.; Griffin, B.R.; Rusterholz, T.; Chinoy, E.D. Entrainment of the human circadian clock to the natural light-dark cycle. Curr. Biol. 2013, 23, 1554–1558. [Google Scholar] [CrossRef]
- Eto, T.; Kitamura, S.; Nishimura, K.; Takeoka, K.; Nishimura, Y.; Lee, S.I.; Ohashi, M.; Shikano, A.; Noi, S.; Higuchi, S. Circadian phase advances in children during camping life according to the natural light-dark cycle. J. Physiol. Anthropol. 2022, 41, 42. [Google Scholar] [CrossRef]
- Gapon, L.I.; Shurkevich, N.P.; Vetoshkin, A.S.; Gubin, D.G. The rhythms of arterial pressure and heart rate in individuals with arterial hypertension under the conditions of Far North. Klin. Med. 2006, 84, 39–44. [Google Scholar]
- Arendt, J. Biological rhythms during residence in Polar Regions. Chronobiol. Int. 2012, 29, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Gubin, D.; Cornelissen, G.; Weinert, D.; Vetoshkin, A.S.; Gapon, L.I.; Shurkevich, N.P.; Poshinov, F.V.; Belozerova, N.V.; Danilova, L.A. Circadian disruption and Vascular Variability Disorders (VVD)—Mechanisms linking aging, disease state and Arctic shift-work: Applications for chronotherapy. World Heart J. 2013, 5, 285–306. [Google Scholar]
- Paul, M.A.; Love, R.J.; Hawton, A.; Brett, K.; McCreary, D.R.; Arendt, J. Sleep deficits in the High Arctic summer in relation to light exposure and behaviour: Use of melatonin as a countermeasure. Sleep. Med. 2015, 16, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Danilenko, K.V.; Kobelev, E.; Semenova, E.A.; Aftanas, L.I. Summer-winter difference in 24-h melatonin rhythms in subjects on a 5-workdays schedule in Siberia without daylight saving time transitions. Physiol. Behav. 2019, 212, 112686. [Google Scholar] [CrossRef] [PubMed]
- Unnarsdóttir, A.B.; Hauksdóttir, A.; Aspelund, T.; Gunnarsdóttir, V.; Tómasson, G.; Jakobsdóttir, J.; Anna Valdimarsdóttir, U.; Thordardottir, E.B. Sleep disturbances among women in a Subarctic region: A nationwide study. Sleep 2022, 45, zsac100. [Google Scholar] [CrossRef] [PubMed]
- Sahar, S.; Sassone-Corsi, P. Regulation of metabolism: The circadian clock dictates the time. Trends Endocrinol. Metab. 2012, 23, 1–8. [Google Scholar] [CrossRef]
- Gnocchi, D.; Pedrelli, M.; Hurt-Camejo, E.; Parini, P. Lipids around the Clock: Focus on Circadian Rhythms and Lipid Metabolism. Biology 2015, 4, 104–132. [Google Scholar] [CrossRef]
- Jha Kumar, P.; Challet, E.; Kalsbeek, A. Circadian rhythms in glucose and lipid metabolism in nocturnal and diurnal mammals. Mol. Cell Endocrinol. 2015, 418 Pt 1, 74–88. [Google Scholar] [CrossRef]
- Gooley, J.J. Circadian regulation of lipid metabolism. Proc. Nutr. Soc. 2016, 75, 440–450. [Google Scholar] [CrossRef]
- Plano, S.A.; Casiraghi, L.P.; García Moro, P.; Paladino, N.; Golombek, D.A.; Chiesa, J.J. Circadian and Metabolic Effects of Light: Implications in Weight Homeostasis and Health. Front. Neurol. 2017, 8, 558. [Google Scholar] [CrossRef] [PubMed]
- Poggiogalle, E.; Jamshed, H.; Peterson, C.M. Circadian regulation of glucose, lipid, and energy metabolism in humans. Metabolism 2018, 84, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, G. Circadian disruption and metabolic disease risk. In Functional Foods and Nutraceuticals in Metabolic and Non-Communicable Diseases; Singh, R., Watanabe, S., Isaza, A., Eds.; Academic Press: New York, NY, USA, 2022; pp. 513–524. [Google Scholar]
- Okuliarova, M.; Rumanova, V.S.; Stebelova, K.; Zeman, M. Dim Light at Night Disturbs Molecular Pathways of Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6919. [Google Scholar] [CrossRef] [PubMed]
- Kent, B.A.; Rahman, S.A.; St. Hilaire, M.A.; Grant, L.K.; Rüger, M.; Czeisler, C.A.; Lockley, S.W. Circadian lipid and hepatic protein rhythms shift with a phase response curve different than melatonin. Nat. Commun. 2022, 13, 681. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, A.; Courville, A.B.; Chen, K.Y. The Complex Effects of Light on Metabolism in Humans. Nutrients 2023, 15, 1391. [Google Scholar] [CrossRef] [PubMed]
- Danilenko, K.V.; Stefani, O.; Voronin, K.A.; Mezhakova, M.S.; Petrov, I.M.; Borisenkov, M.F.; Markov, A.A.; Gubin, D.G. Wearable Light-and-Motion Dataloggers for Sleep/Wake Research: A Review. Appl. Sci. 2022, 12, 11794. [Google Scholar] [CrossRef]
- Stampfli, J.; Schrader, B.; di Battista, C.; Häfliger, R.; Schälli, O.; Wichmann, G.; Zumbühl, C.; Blattner, P.; Cajochen, C.; Lazar, R.; et al. The Light-Dosimeter: A new device to help advance research on the non-visual responses to light. Light. Res. Technol. 2023, 55, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef]
- Mure, L.S. Intrinsically Photosensitive Retinal Ganglion Cells of the Human Retina. Front. Neurol. 2021, 12, 636330. [Google Scholar] [CrossRef]
- Garfield, V. The Association Between Body Mass Index (BMI) and Sleep Duration: Where Are We after nearly Two Decades of Epidemiological Research? Int. J. Environ. Res. Public Health 2019, 16, 4327. [Google Scholar] [CrossRef]
- Reid, K.J.; Santostasi, G.; Baron, K.G.; Wilson, J.; Kang, J.; Zee, P.C. Timing and intensity of light correlate with body weight in adults. PLoS ONE 2014, 9, e92251. [Google Scholar] [CrossRef] [PubMed]
- Constantino, D.B.; Xavier, N.B.; Levandovski, R.; Roenneberg, T.; Hidalgo, M.P.; Pilz, L.K. Relationship between Circadian Strain, Light Exposure, and Body Mass Index in Rural and Urban Quilombola Communities. Front. Physiol. 2022, 12, 773969. [Google Scholar] [CrossRef] [PubMed]
- Cespedes Feliciano, E.M.; Quante, M.; Weng, J.; Mitchell, J.A.; James, P.; Marinac, C.R.; Mariani, S.; Redline, S.; Kerr, J.; Godbole, S.; et al. Actigraphy-Derived Daily Rest-Activity Patterns and Body Mass Index in Community-Dwelling Adults. Sleep 2017, 40, zsx168. [Google Scholar] [CrossRef] [PubMed]
- Quante, M.; Cespedes Feliciano, E.M.; Rifas-Shiman, S.L.; Mariani, S.; Kaplan, E.R.; Rueschman, M.; Oken, E.; Taveras, E.M.; Redline, S. Association of Daily Rest-Activity Patterns with Adiposity and Cardiometabolic Risk Measures in Teens. J. Adolesc. Health 2019, 65, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Martinez-Lozano, N.; Tvarijonaviciute, A.; Rios, R.; Scheer, F.A.J.L.; Garaulet, M. Blunted rest-activity rhythms link to higher body mass index and inflammatory markers in children. Sleep 2021, 44, zsaa256. [Google Scholar] [CrossRef] [PubMed]
- Weinert, D.; Gubin, D. The Impact of Physical Activity on the Circadian System: Benefits for Health, Performance and Wellbeing. Appl. Sci. 2022, 12, 9220. [Google Scholar] [CrossRef]
- Obayashi, K.; Saeki, K.; Kurumatani, N. Ambient Light Exposure and Changes in Obesity Parameters: A Longitudinal Study of the HEIJO-KYO Cohort. J. Clin. Endocrinol. Metab. 2016, 101, 3539–3547. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, K.; Yamagami, Y.; Kurumatani, N.; Saeki, K. Bedroom lighting environment and incident diabetes mellitus: A longitudinal study of the HEIJO-KYO cohort. Sleep Med. 2020, 65, 1–3. [Google Scholar] [CrossRef]
- Kim, M.; Vu, T.H.; Maas, M.B.; Braun, R.I.; Wolf, M.S.; Roenneberg, T.; Daviglus, M.L.; Reid, K.J.; Zee, P.C. Light at night in older age is associated with obesity, diabetes, and hypertension. Sleep 2023, 46, zsac130. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, Z.J.; Liu, H.Y.; Cai, J.; Lu, Q.K.; Ji, L.D.; Xu, J. The melatonin receptor 1B gene links circadian rhythms and type 2 diabetes mellitus: An evolutionary story. Ann. Med. 2023, 55, 1262–1286. [Google Scholar] [CrossRef]
- Lane, J.M.; Chang, A.M.; Bjonnes, A.C.; Aeschbach, D.; Anderson, C.; Cade, B.E.; Cain, S.W.; Czeisler, C.A.; Gharib, S.A.; Gooley, J.J.; et al. Impact of Common Diabetes Risk Variant in MTNR1B on Sleep, Circadian, and Melatonin Physiology. Diabetes 2016, 65, 1741–1751. [Google Scholar] [CrossRef]
- Tan, X.; Ciuculete, D.M.; Schiöth, H.B.; Benedict, C. Associations between chronotype, MTNR1B genotype and risk of type 2 diabetes in UK Biobank. J. Intern. Med. 2020, 287, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Gubin, D.; Neroev, V.; Malishevskaya, T.; Cornelissen, G.; Astakhov, S.Y.; Kolomeichuk, S.; Yuzhakova, N.; Kabitskaya, Y.; Weinert, D. Melatonin mitigates disrupted circadian rhythms, lowers intraocular pressure, and improves retinal ganglion cells function in glaucoma. J. Pineal Res. 2021, 70, e12730. [Google Scholar] [CrossRef] [PubMed]
- Vejrazkova, D.; Vankova, M.; Vcelak, J.; Krejci, H.; Anderlova, K.; Tura, A.; Pacini, G.; Sumova, A.; Sladek, M.; Bendlova, B. The rs10830963 Polymorphism of the MTNR1B Gene: Association with Abnormal Glucose, Insulin and C-peptide Kinetics. Front. Endocrinol. 2022, 13, 868364. [Google Scholar] [CrossRef] [PubMed]
- Xia, A.Y.; Zhu, H.; Zhao, Z.J.; Liu, H.Y.; Wang, P.H.; Ji, L.D.; Xu, J. Molecular Mechanisms of the Melatonin Receptor Pathway Linking Circadian Rhythm to Type 2 Diabetes Mellitus. Nutrients 2023, 15, 1406. [Google Scholar] [CrossRef]
- Goni, L.; Sun, D.; Heianza, Y.; Wang, T.; Huang, T.; Martínez, J.A.; Shang, X.; Bray, G.A.; Smith, S.R.; Sacks, F.M.; et al. A circadian rhythm-related MTNR1B genetic variant modulates the effect of weight-loss diets on changes in adiposity and body composition: The POUNDS Lost trial. Eur. J. Nutr. 2019, 58, 1381–1389. [Google Scholar] [CrossRef]
- de Onis, M.; Onyango, A.W.; Borghi, E.; Siyam, A.; Nishida, C.; Siekmann, J. Development of a WHO growth reference for school-aged children and adolescents. Bull. World Health Organ. 2007, 85, 660–667. [Google Scholar] [CrossRef]
- Girardin, J.-L.; Kripke, D.F.; Mason, W.J.; Elliott, J.A.; Youngstedt, S.D. Sleep estimation from wrist movement quantified by different actigraphic modalities. J. Neurosci. Methods 2001, 105, 185–191. [Google Scholar]
- Blackwell, T.; Redline, S.; Ancoli-Israel, S.; Schneider, J.L.; Surovec, S.; Johnson, N.L.; Cauley, J.A.; Stone, K.L. Study of Osteoporotic Fractures Research Group. Comparison of sleep parameters from actigraphy and polysomnography in older women: The SOF study. Sleep 2008, 1, 283–291. [Google Scholar] [CrossRef]
- Price, L.L.; Lyachev, A.; Khazova, M. Optical performance characterization of light-logging actigraphy dosimeters. J. Opt. Soc. Am. A 2017, 34, 545–557. [Google Scholar] [CrossRef]
- Price, L.L.A.; Lyachev, A. Modification of a personal dosimetry device for logging melanopic irradiance. Light. Res. Technol. 2017, 49, 922–927. [Google Scholar] [CrossRef]
- St Hilaire, M.A.; Ámundadóttir, M.L.; Rahman, S.A.; Rajaratnam, S.M.W.; Rüger, M.; Brainard, G.C.; Czeisler, C.A.; Andersen, M.; Gooley, J.J.; Lockley, S.W. The spectral sensitivity of human circadian phase resetting and melatonin suppression to light changes dynamically with light duration. Proc. Natl. Acad. Sci. USA 2022, 119, e2205301119. [Google Scholar] [CrossRef] [PubMed]
- CIE S 026/E:2018; CIE System for Metrology of Optical Radiation for ipRGC-Influenced Responses to Light. CIE Central Bureau: Vienna, Austria, 2018.
- Cole, R.J.; Kripke, D.F.; Gruen, W.; Mullaney, D.J.; Gillin, J.C. Automatic sleep/wake identification from wrist activity. Sleep 1992, 15, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, G. Cosinor-based rhythmometry. Theor. Biol. Med. Model. 2014, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Diedenhofen, B.; Much, J. Cocor: A comprehensive solution for the statistical comparison of correlations. PLoS ONE 2015, 10, e0121945. [Google Scholar] [CrossRef] [PubMed]
- Cheung, I.N.; Zee, P.C.; Shalman, D.; Malkani, R.G.; Kang, J.; Reid, K.J. Morning and Evening Blue-Enriched Light Exposure Alters Metabolic Function in Normal Weight Adults. PLoS ONE 2016, 11, e0155601. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, J.F.; Wefers, J.; Doligkeit, D.; Schlangen, L.; Dautzenberg, B.; Rense, P.; van Moorsel, D.; Hoeks, J.; Moonen-Kornips, E.; Gordijn, M.C.M.; et al. The influence of bright and dim light on substrate metabolism, energy expenditure and thermoregulation in insulin-resistant individuals depends on time of day. Diabetologia 2022, 65, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Fleury, G.; Masís-Vargas, A.; Kalsbeek, A. Metabolic Implications of Exposure to Light at Night: Lessons from Animal and Human Studies. Obesity 2020, 28 (Suppl. S1), S18–S28. [Google Scholar] [CrossRef]
- Benedito-Silva, A.A.; Evans, S.; Viana Mendes, J.; Castro, J.; Gonçalves, B.D.S.B.; Ruiz, F.S.; Beijamini, F.; Evangelista, F.S.; Vallada, H.; Krieger, J.E.; et al. Association between light exposure and metabolic syndrome in a rural Brazilian town. PLoS ONE 2020, 15, e0238772. [Google Scholar] [CrossRef]
- Meléndez-Fernández, O.H.; Walton, J.C.; DeVries, A.C.; Nelson, R.J. The role of daylight exposure on body mass in male mice. Physiol. Behav. 2023, 266, 114186. [Google Scholar] [CrossRef]
- von Gall, C. The Effects of Light and the Circadian System on Rhythmic Brain Function. Int. J. Mol. Sci. 2022, 23, 2778. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Ryu, S.H.; Lee, B.R.; Kim, K.H.; Lee, E.; Choi, J. Effects of artificial light at night on human health: A literature review of observational and experimental studies applied to exposure assessment. Chronobiol. Int. 2015, 32, 1294–1310. [Google Scholar] [CrossRef] [PubMed]
- Touitou, Y.; Reinberg, A.; Touitou, D. Association between light at night, melatonin secretion, sleep deprivation, and the internal clock: Health impacts and mechanisms of circadian disruption. Life Sci. 2017, 173, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Bumgarner, J.R.; Nelson, R.J. Light at Night and Disrupted Circadian Rhythms Alter Physiology and Behavior. Integr. Comp. Biol. 2021, 61, 1160–1169. [Google Scholar] [CrossRef] [PubMed]
- Fonken, L.K.; Aubrecht, T.G.; Meléndez-Fernández, O.H.; Weil, Z.M.; Nelson, R.J. Dim light at night disrupts molecular circadian rhythms and increases body weight. J. Biol. Rhythm. 2013, 28, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Aubrecht, T.G.; Jenkins, R.; Nelson, R.J. Dim light at night increases body mass of female mice. Chronobiol. Int. 2015, 32, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Opperhuizen, A.L.; Stenvers, D.J.; Jansen, R.D.; Foppen, E.; Fliers, E.; Kalsbeek, A. Light at night acutely impairs glucose tolerance in a time-, intensity- and wavelength-dependent manner in rats. Diabetologia 2017, 60, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- McFadden, E.; Jones, M.E.; Schoemaker, M.J.; Ashworth, A.; Swerdlow, A.J. The relationship between obesity and exposure to light at night: Cross-sectional analyses of over 100,000 women in the Breakthrough Generations Study. Am. J. Epidemiol. 2014, 180, 245–250. [Google Scholar] [CrossRef]
- Nelson, R.J.; Chbeir, S. Dark matters: Effects of light at night on metabolism. Proc. Nutr. Soc. 2018, 77, 223–229. [Google Scholar] [CrossRef]
- Park, Y.M.; White, A.J.; Jackson, C.L.; Weinberg, C.R.; Sandler, D.P. Association of Exposure to Artificial Light at Night While Sleeping with Risk of Obesity in Women. JAMA Intern. Med. 2019, 179, 1061–1071. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, R.; Xin, Z.; Zhao, Z.; Li, M.; Wang, T.; Xu, M.; Lu, J.; Wang, S.; Li, H.; et al. Sex- and age-specific association between outdoor light at night and obesity in Chinese adults: A national cross-sectional study of 98,658 participants from 162 study sites. Front. Endocrinol. 2023, 14, 1119658. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.J.; Xie, Z.Y.; Gong, Y.C.; Wang, L.B.; Xie, Y.Y.; Lin, L.Z.; Zeng, X.W.; Yang, B.Y.; Zhang, W.; Liu, R.Q.; et al. The association between outdoor light at night exposure and adult obesity in Northeastern China. Int. J. Environ. Health Res. 2023, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Danilenko, K.V.; Mustafina, S.V.; Pechenkina, E.A. Bright light for weight loss: Results of a controlled crossover trial. Obes. Facts 2013, 6, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Pattinson, C.L.; Allan, A.C.; Staton, S.L.; Thorpe, K.J.; Smith, S.S. Environmental Light Exposure Is Associated with Increased Body Mass in Children. PLoS ONE 2016, 11, e0143578. [Google Scholar] [CrossRef] [PubMed]
- Melanson, E.L.; Ritchie, H.K.; Dear, T.B.; Catenacci, V.; Shea, K.; Connick, E.; Moehlman, T.M.; Stothard, E.R.; Higgins, J.; McHill, A.W.; et al. Daytime bright light exposure, metabolism, and individual differences in wake and sleep energy expenditure during circadian entrainment and misalignment. Neurobiol. Sleep Circadian Rhythm. 2018, 4, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Burns, A.C.; Phillips, A.J.K.; Rutter, M.K.; Saxena, R.; Cain, S.W.; Lane, J.M. Genome-wide gene by environment study of time spent in daylight and chronotype identifies emerging genetic architecture underlying light sensitivity. Sleep 2023, 46, zsac287. [Google Scholar] [CrossRef] [PubMed]
- Chellappa, S.L.; Steiner, R.; Oelhafen, P.; Cajochen, C. Sex differences in light sensitivity impact on brightness perception, vigilant attention and sleep in humans. Sci. Rep. 2017, 7, 14215. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Morris, C.J.; Caputo, R.; Wang, W.; Garaulet, M.; Scheer, F.A.J.L. Sex differences in the circadian misalignment effects on energy regulation. Proc. Natl. Acad. Sci. USA 2019, 116, 23806–23812. [Google Scholar] [CrossRef]
- Figueiro, M.G.; Overington, D. Self-luminous devices and melatonin suppression in adolescents. Light. Res. Technol. 2016, 48, 966–975. [Google Scholar] [CrossRef]
- Chellappa, S.L. Aging, light sensitivity and circadian health. Aging 2021, 13, 25604–25606. [Google Scholar] [CrossRef]
- Phillips, A.J.K.; Vidafar, P.; Burns, A.C.; McGlashan, E.M.; Anderson, C.; Rajaratnam, S.M.W.; Lockley, S.W.; Cain, S.W. High sensitivity and interindividual variability in the response of the human circadian system to evening light. Proc. Natl. Acad. Sci. USA 2019, 116, 12019–12024. [Google Scholar] [CrossRef] [PubMed]
- Vartanian, G.V.; Li, B.Y.; Chervenak, A.P.; Walch, O.J.; Pack, W.; Ala-Laurila, P.; Wong, K.Y. Melatonin Suppression by Light in Humans Is More Sensitive than Previously Reported. J. Biol. Rhythm. 2015, 30, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Khalsa, S.B.; Jewett, M.E.; Cajochen, C.; Czeisler, C.A. A phase response curve to single bright light pulses in human subjects. J. Physiol. 2003, 549 Pt 3, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Brooks, T.G.; Lahens, N.F.; Grant, G.R.; Sheline, Y.I.; FitzGerald, G.A.; Skarke, C. Diurnal rhythms of wrist temperature are associated with future disease risk in the UK Biobank. Nat. Commun. 2023, 14, 5172. [Google Scholar] [CrossRef] [PubMed]
- Refinetti, R. Circadian rhythmicity of body temperature and metabolism. Temperature 2020, 7, 321–362. [Google Scholar] [CrossRef] [PubMed]
- Kräuchi, K.; Wirz-Justice, A. Circadian clues to sleep onset mechanisms. Neuropsychopharmacology 2001, 25, S92–S96. [Google Scholar] [CrossRef] [PubMed]
- Kalsbeek, A.; la Fleur, S.; Fliers, E. Circadian control of glucose metabolism. Mol. Metab. 2014, 3, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Corbalán-Tutau, M.D.; Madrid, J.A.; Ordovás, J.M.; Smith, C.E.; Nicolás, F.; Garaulet, M. Differences in daily rhythms of wrist temperature between obese and normal-weight women: Associations with metabolic syndrome features. Chronobiol. Int. 2011, 28, 425–433. [Google Scholar] [CrossRef]
- Zerón-Rugerio, M.F.; Diez-Noguera, A.; Izquierdo-Pulido, M.; Cambras, T. Adiposity and body mass index of young women are associated with altered 24-hour profile of wrist temperature and sleep quality. Chronobiol. Int. 2020, 37, 1580–1590. [Google Scholar] [CrossRef]
- Hoffmann, M.E.; Rodriguez, S.M.; Zeiss, D.M.; Wachsberg, K.N.; Kushner, R.F.; Landsberg, L.; Linsenmeier, R.A. 24-h core temperature in obese and lean men and women. Obesity 2012, 20, 1585–1590. [Google Scholar] [CrossRef]
- Gubin, D.G.; Nelaeva, A.A.; Uzhakova, A.E.; Hasanova, Y.V.; Cornelissen, G.; Weinert, D. Disrupted circadian rhythms of body temperature, heart rate and fasting blood glucose in prediabetes and type 2 diabetes mellitus. Chronobiol. Int. 2017, 34, 1136–1148. [Google Scholar] [CrossRef] [PubMed]
- Heikens, M.J.; Gorbach, A.M.; Eden, H.S.; Savastano, D.M.; Chen, K.Y.; Skarulis, M.C.; Yanovski, J.A. Core body temperature in obesity. Am. J. Clin. Nutr. 2011, 93, 963–967. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, H.; Liu, N.; Cao, X.; Yang, Z.; Lu, B.; Hu, R.; Wang, X.; Wen, J. Melatonin exerts an inhibitory effect on insulin gene transcription via MTNR1B and the downstream Raf-1/ERK signaling pathway. Int. J. Mol. Med. 2018, 41, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Jia, W. Linking MTNR1B variants to diabetes: The role of circadian rhythms. Diabetes. 2016, 65, 1490–1492. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.J.; Cajochen, C.; Borbély, A.A. Effect of a single 3-hour exposure to bright light on core body temperature and sleep in humans. Neurosci. Lett. 1991, 121, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Cajochen, C.; Dijk, D.J.; Borbély, A.A. Dynamics of EEG slow-wave activity and core body temperature in human sleep after exposure to bright light. Sleep 1992, 15, 337–343. [Google Scholar] [PubMed]
- Gubin, D.G.; Weinert, D.; Rybina, S.V.; Danilova, L.A.; Solovieva, S.V.; Durov, A.M.; Prokopiev, N.Y.; Ushakov, P.A. Activity, sleep and ambient light have a different impact on circadian blood pressure, heart rate and body temperature rhythms. Chronobiol. Int. 2017, 34, 632–649. [Google Scholar] [CrossRef] [PubMed]
- Gubin, D.; Weinert, D.; Solovieva, S.V.; Durov, A.M.; Litvinova, N.S.; Danilova, L.A.; Prokopiev, N.Y.; Trushnikov, D.Y.; Kartashova, E.A. Melatonin attenuates light-at-night effects on systolic blood pressure and body temperature but does not affect diastolic blood pressure and heart rate circadian rhythms. Biol. Rhythm. Res. 2020, 51, 780–793. [Google Scholar] [CrossRef]
- Choi, Y.; Nakamura, Y.; Akazawa, N.; Park, I.; Kwak, H.B.; Tokuyama, K.; Maeda, S. Effects of nocturnal light exposure on circadian rhythm and energy metabolism in healthy adults: A randomized crossover trial. Chronobiol. Int. 2022, 39, 602–612. [Google Scholar] [CrossRef]
- Aschbacher, K.; Rodriguez-Fernandez, M.; van Wietmarschen, H.; Tomiyama, A.J.; Jain, S.; Epel, E.; Doyle, F.J., 3rd; van der Greef, J. The hypothalamic-pituitary-adrenal-leptin axis and metabolic health: A systems approach to resilience, robustness and control. Interface Focus 2014, 4, 20140020. [Google Scholar] [CrossRef]
- Malcher-Lopes, R.; Di, S.; Marcheselli, V.S.; Weng, F.J.; Stuart, C.T.; Bazan, N.G.; Tasker, J.G. Opposing crosstalk between leptin and glucocorticoids rapidly modulates synaptic excitation via endocannabinoid release. J. Neurosci. 2006, 26, 6643–6650. [Google Scholar] [CrossRef] [PubMed]
- Pralong, F.P.; Roduit, R.; Waeber, G.; Castillo, E.; Mosimann, F.; Thorens, B.; Gaillard, R.C. Leptin inhibits directly glucocorticoid secretion by normal human and rat adrenal gland. Endocrinology 1998, 139, 4264–4268. [Google Scholar] [CrossRef] [PubMed]
- Paul, R.F.; Hassan, M.; Nazar, H.S.; Gillani, S.; Afzal, N.; Qayyum, I. Effect of body mass index on serum leptin levels. J. Ayub. Med. Coll. Abbottabad. 2011, 23, 40–43. [Google Scholar] [PubMed]
- Cheng, J.; Luo, Y.; Li, Y.; Zhang, F.; Zhang, X.; Zhou, X.; Ji, L. Sex- and body mass index-specific reference intervals for serum leptin: A population based study in China. Nutr. Metab. 2022, 19, 54. [Google Scholar] [CrossRef] [PubMed]
- Mäntele, S.; Otway, D.T.; Middleton, B.; Bretschneider, S.; Wright, J.; Robertson, M.D.; Skene, D.J.; Johnston, J.D. Daily rhythms of plasma melatonin, but not plasma leptin or leptin mRNA, vary between lean, obese and type 2 diabetic men. PLoS ONE 2012, 7, e37123. [Google Scholar] [CrossRef] [PubMed]
- Figueiro, M.G.; Plitnick, B.; Rea, M.S. Light modulates leptin and ghrelin in sleep-restricted adults. Int. J. Endocrinol. 2012, 2012, 530726. [Google Scholar] [CrossRef] [PubMed]
- Driller, M.W.; Jacobson, G.; Uiga, L. Hunger hormone and sleep responses to the built-in blue-light filter on an electronic device: A pilot study. Sleep Sci. 2019, 12, 171–177. [Google Scholar] [CrossRef]
- Jung, C.M.; Khalsa, S.B.; Scheer, F.A.; Cajochen, C.; Lockley, S.W.; Czeisler, C.A.; Wright, K.P., Jr. Acute effects of bright light exposure on cortisol levels. J. Biol. Rhythm. 2010, 25, 208–216. [Google Scholar] [CrossRef]
- Leproult, R.; Colecchia, E.F.; L’Hermite-Balériaux, M.; Van Cauter, E. Transition from dim to bright light in the morning induces an immediate elevation of cortisol levels. J. Clin. Endocrinol. Metab. 2001, 86, 151–157. [Google Scholar] [CrossRef]
- Schöllhorn, I.; Deuring, G.; Stefani, O.; Strumberger, M.A.; Rosburg, T.; Lemoine, P.; Pross, A.; Wingert, B.; Mager, R.; Cajochen, C. Effects of nature-adapted lighting solutions (“Virtual Sky”) on subjective and objective correlates of sleepiness, well-being, visual and cognitive performance at the workplace. PLoS ONE 2023, 18, e0288690. [Google Scholar] [CrossRef]

{kind=link}
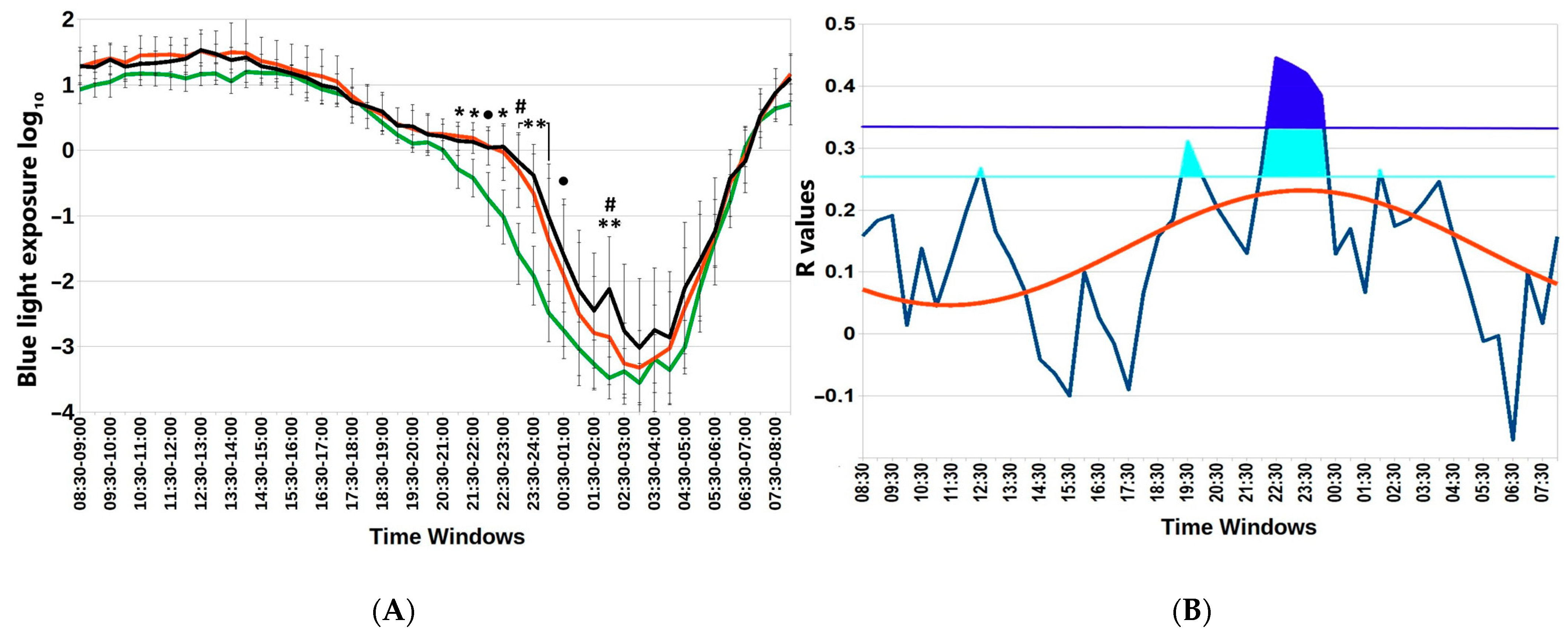{kind=link}
{kind=link}
{kind=link}
| Sex | Indigeneity | MTNR1B Genotype | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Females (n) | Males (n) | p-Value | Non-Natives (n) | Natives (n) | p-Value | GG + GC (n) | CC (n) | p-Value | |
| Age | 28.4 ± 12.6 (51) | 38.0 ± 14.0 (11) | 0.041 | 40.7 ± 11.0 (38) | 29.2 ± 15.9 (24) | 0.001 | 39.8 ± 12.6 (22) | 40.4 ± 11.4 (28) | 0.854 |
| Age, >20 y. | 37.1 ± 3.8 (41) | 43.7 ± 8.6 (7) | 0.054 | 43.0 ± 8.0 (35) | 42.0 ± 9.6 (13) | 0.709 | 43.6 ± 8.4 (19) | 42.5 ± 8.5 (26) | 0.675 |
| Age, 12–16 y. | 13.0 ± 1.4 (10) | 14.4 ± 1.4 (4) | 0.109 | 14.0 ± 2.6 (3) | 14.0 ± 1.2 (11) | 0.999 | 15.3 ± 1.5 (3) | 12.5 ± 0.7 (2) | 0.100 |
| BMI | 24.2 ± 4.6 (51) | 24.2 ± 4.9 (11) | 0.995 | 25.4 ± 4.6 (38) | 22.2 ± 4.6 (24) | 0.010 | 25.1 ± 4.7 (22) | 24.1 ± 4.7 (28) | 0.456 |
| BMI, >20 y. | 25.1 ± 5.0 (41) | 26.8 ± 3.4 (7) | 0.489 | 25.8 ± 4.5 (35) | 24.1 ± 5.4 (13) | 0.278 | 25.8 ± 4.6 (19) | 24.1 ± 5.4 (26) | 0.312 |
| BMI, 12–16 y. | 20.2 ± 1.7 (10) | 19.5 ± 1.5 (4) | 0.389 | 20.3 ± 1.5 (3) | 19.9 ± 1.7 (11) | 0.750 | 20.4 ± 1.2 (3) | 20.2 ± 2.2 (2) | 0.896 |
| Variable | BMI Groups | Regression with BMI | |||||
|---|---|---|---|---|---|---|---|
| 1. Normal (<25, n = 40) | 2. Overweight (25–30, n = 11) | 3. Obesity (30–35, n = 11) | K-W p-Value | r | p | MR + CF p | |
| Activity, PIM # | |||||||
| MESOR | 2606 ± 609 | 2226 ± 539 | 2708 ± 1075 | 0.257 | −0.079 | 0.541 | 0.494 |
| 24 h A | 2002 ± 638 | 1633 ± 447 | 1823 ± 853 | 0.182 | −0.234 | 0.067 | 0.727 |
| Phase | 14:47 ± 1:16 | 14:54 ± 1:22 | 14:31 ± 1:16 | 0.854 | −0.153 | 0.234 | 0.222 |
| M10 | 4270 ± 1070 | 3650 ± 816 | 4288 ± 1861 | 0.208 | −0.120 | 0.354 | 0.428 |
| M10 Onset | 9:08 ± 1:17 | 8:32 ± 1:25 | 8:58 ± 1:00 | 0.402 | 0.175 | 0.174 | 0.334 |
| L5 | 231 ± 208 | 124 ± 69 | 271 ± 242 | 0.068 | 0.077 | 0.554 | 0.855 |
| L5 Onset | 1:34 ± 1:11 | 1:29 ± 1:13 | 2:04 ± 1:23 | 0.406 | −0.123 | 0.342 | 0.843 |
| IV | 0.888 ± 0.167 | 0.847 ± 0.164 | 0.836 ± 0.180 | 0.484 | −0.147 | 0.255 | 0.178 |
| IS | 0.563 ± 0.113 | 0.523 ± 0.103 | 0.497 ± 0.067 | 0.068 | −0.186 | 0.147 | 0.894 |
| RA | 0.893 ± 0.065 | 0.903 ± 0.050 | 0.836 ± 0.071 | 0.056 | −0.345 | 0.006 * | 0.299 |
| CFI | 0.676 ± 0.058 | 0.663 ± 0.056 | 0.600 ± 0.071 | 0.546 | −0.172 | 0.182 | 0.994 |
| Wrist temperature, °C # | |||||||
| MESOR | 31.99 ± 0.50 | 31.63 ± 0.40 | 31.29 ± 0.64 | 0.002 1–3 | −0.491 | <0.0001 * | 0.008 * |
| 24 h A | 1.42 ± 0.53 | 1.29 ± 0.57 | 1.29 ± 0.59 | 0.694 | −0.072 | 0.576 | 0.161 |
| Phase | 2:58 ± 1:27 | 2:19 ± 1:43 | 2:22:1:50 | 0.397 | −0.322 | 0.011 * | 0.012 * |
| Sleep characteristics, hh:mm # | |||||||
| Bedtime | 22:35 ± 0:56 | 22:59 ± 1:15 | 22:49 ± 1:33 | 0.587 | −0.003 | 0.983 | 0.618 |
| Wake time | 6:51 ± 1:03 | 7:13 ± 1:38 | 7:07 ± 2:09 | 0.395 | −0.028 | 0.826 | 0.576 |
| Sleep phase | 2:43 ± 0:54 | 3:06 ± 1:15 | 2:54 ± 1:33 | 0.514 | −0.040 | 0.760 | 0.432 |
| Time in bed | 8:13 ± 0:50 | 8:13 ± 1:27 | 8:17 ± 2:06 | 0.663 | 0.013 | 0.921 | 0.931 |
| Total sleep | 7:19 ± 0:44 | 7:10 ± 1:18 | 7:22 ± 2:06 | 0.467 | 0.074 | 0.568 | 0.767 |
| Sleep latency, min | 02.05 ± 1.68 | 3.08 ± 2.92 | 3.18 ± 2.63 | 0.279 | −0.228 | 0.075 | 0.174 |
| Sleep efficiency, % | 88.67 ± 4.03 | 85.58 ± 6.24 | 88.15 ± 6.73 | 0.396 | −0.207 | 0.106 | 0.613 |
| WASO | 0:50 ± 0:21 | 0:58 ± 0:27 | 0:50 ± 0:30 | 0.619 | 0.147 | 0.254 | 0.721 |
| Blue light, μw/cm2 # | |||||||
| MESOR | 9.65 ± 7.00 | 15.23 ± 10.19 | 12.46 ± 5.75 | 0.074 | 0.158 | 0.221 | 0.146 |
| 24 h A | 13.89 ± 9.78 | 22.54 ± 15.81 | 18.11 ± 9.57 | 0.136 | 0.149 | 0.249 | 0.167 |
| Phase | 12:58 ± 0:53 | 12:34 ± 0:51 | 12:58 ± 0:29 | 0.699 | 0.099 | 0.442 | 0.420 |
| M10 | 19.81 ± 11.98 | 34.06 ± 23.28 | 26.44 ± 13.43 | 0.075 | 0.178 | 0.166 | 0.661 |
| M10 Onset | 7:57 ± 0:51 | 7:43 ± 0:39 | 7:57 ± 0:38 | 0.780 | 0.080 | 0.535 | 0.550 |
| L5 | 0.06 ± 0.14 | 0.07 ± 0.16 | 0.10 ± 0.11 | 0.126 | 0.086 | 0.506 | 0.006 * |
| L5 Onset | 1:15 ± 1:47 | 0:40 ± 0:43 | 2:26 ± 2:20 | 0.035 2–3 | −0.237 | 0.063 | 0.505 |
| L5 log10 | −2.42 ± 1.28 | −2.29 ± 1.25 | −1.55 ± 0.93 | 0.129 | 0.308 | 0.015 * | 0.185 |
| IV | 0.937 ± 0.352 | 0.924 ± 0.361 | 0.879 ± 0.297 | 0.933 | −0.041 | 0.752 | 0.168 |
| IS | 0.381 ± 0.133 | 0.444 ± 0.188 | 0.375 ± 0.105 | 0.455 | 0.103 | 0.428 | 0.110 |
| RA | 0.988 ± 0.023 | 0.985 ± 0.022 | 0.969 ± 0.027 | 0.022 1–3 | −0.300 | 0.018 * | 0.148 |
| DDIbl | 367 ± 106 | 277 ± 150 | 313 ± 106 | 0.052 | −0.185 | 0.150 | 0.943 |
| NEIbl | 1.49 ± 1.58 | 3.37 ± 3.38 | 3.39 ± 2.21 | 0.003 1–2/1–3 | 0.417 | 0.0008 * | 0.005 * |
| Variable | Leptin | Cortisol | ||||
|---|---|---|---|---|---|---|
| r | p | MR + CF p | r | p | MR + CF p | |
| Activity (power integrative mode) | ||||||
| MESOR | −0.288 | 0.024 | 0.132 | −0.174 | 0.184 | 0.374 |
| 24 h Amplitude | −0.289 | 0.024 * | 0.583 | −0.134 | 0.308 | 0.360 |
| Phase | −0.279 | 0.030 * | 0.210 | 0.186 | 0.155 | 0.071 |
| M10 | −0.308 | 0.016 | 0.208 | −0.205 | 0.116 | 0.177 |
| M10 Onset | −0.242 | 0.060 | 0.325 | −0.116 | 0.378 | 0.518 |
| L5 | 0.069 | 0.597 | 0.809 | −0.266 | 0.040 | 0.074 |
| L5 Onset | 0.079 | 0.545 | 0.807 | −0.033 | 0.802 | 0.969 |
| IV | −0.045 | 0.728 | 0.277 | −0.017 | 0.895 | 0.686 |
| IS | 0.123 | 0.344 | 0.819 | 0.012 | 0.927 | 0.685 |
| RA | −0.159 | 0.222 | 0.867 | 0.227 | 0.081 | 0.077 |
| CFI | 0.015 | 0.912 | 0.631 | −0.049 | 0.708 | 0.910 |
| Wrist temperature | ||||||
| MESOR | −0.130 | 0.318 | 0.299 | 0.106 | 0.420 | 0.269 |
| 24 h Amplitude | −0.159 | 0.220 | 0.235 | 0.114 | 0.387 | 0.617 |
| Phase | −0.302 | 0.014 * | 0.104 | 0.003 | 0.982 | 0.788 |
| Sleep characteristics | ||||||
| Bedtime | 0.214 | 0.098 | 0.132 | −0.161 | 0.220 | 0.166 |
| Wake time | −0.013 | 0.387 | 0.777 | 0.137 | 0.298 | 0.317 |
| Sleep phase | 0.179 | 0.167 | 0.353 | −0.168 | 0.201 | 0.353 |
| Time in bed | −0.078 | 0.550 | 0.258 | −0.034 | 0.795 | 0.657 |
| Total sleep | 0.036 | 0.785 | 0.571 | −0.027 | 0.840 | 0.678 |
| Sleep latency | −0.061 | 0.640 | 0.850 | 0.054 | 0.680 | 0.773 |
| Sleep efficiency, % | −0.179 | 0.167 | 0.336 | 0.070 | 0.595 | 0.618 |
| WASO | −0.339 | 0.008 * | 0.049 | 0.032 | 0.220 | 0.836 |
| Blue light | ||||||
| MESOR | −0.099 | 0.446 | 0.390 | −0.187 | 0.152 | 0.324 |
| 24 h Amplitude | −0.110 | 0.399 | 0.344 | −0.201 | 0.123 | 0.909 |
| Phase | −0.162 | 0.213 | 0.668 | −0.147 | 0.262 | 0.546 |
| M10 | −0.082 | 0.532 | 0.408 | −0.241 | 0.064 | 0.186 |
| M10 Onset | −0.130 | 0.318 | 0.776 | −0.317 | 0.014 | 0.016 |
| L5 | 0.017 | 0.899 | 0.085 | −0.058 | 0.659 | 0.425 |
| L5 Onset | 0.182 | 0.161 | 0.309 | −0.010 | 0.934 | 0.679 |
| L5 log10 | 0.178 | 0.169 | 0.412 | −0.010 | 0.934 | 0.995 |
| IV | 0.228 | 0.078 | 0.052 | 0.215 | 0.100 | 0.093 |
| IS | −0.037 | 0.428 | 0.052 | −0.092 | 0.486 | 0.571 |
| RA | −0.175 | 0.177 | 0.713 | −0.079 | 0.550 | 0.248 |
| DDIbl | 0.047 | 0.733 | 0.762 | 0.256 | 0.049 | 0.148 |
| NEIbl | 0.155 | 0.234 | 0.115 | −0.150 | 0.251 | 0.650 |
| Variable | Population | Age | Sex | |||
|---|---|---|---|---|---|---|
| Non-Natives (n = 38) | Natives (n = 24) | 12–16 (n = 14) | 20–59 (n = 48) | Males (n = 11) | Females (n = 51) | |
| Activity, PIM | ||||||
| MESOR | 2263 ± 534 | 3022 ± 706 *** | 2803 ± 687 | 2485 ± 703 | 2706 ± 823 | 2525 ± 685 |
| 24 h Amplitude | 1596 ± 401 | 2393 ± 695 ** | 2472 ± 748 | 1739 ± 532 * | 2232 ± 969 | 1834 ± 558 |
| Phase | 14:33 ± 1:24 | 15:05 ± 0:56 | 14:48 ± 0:49 | 14:45 ± 1:22 | 15:29 ± 1:04 | 14:36 ± 1.15 * |
| M10 | 3634 ± 812 | 5000 ± 1279 *** | 4842 ± 1288 | 3965 ± 1125 | 4606 ± 1582 | 4067 ± 1112 |
| M10 Onset | 8:43 ± 1:25 | 9:27 ± 0:49 | 9:15 ± 1:00 | 8:56 ± 1:20 | 9:32 ± 0:51 | 8:53 ± 1:19 |
| L5 | 217 ± 219 | 222 ± 171 | 125 ± 46 | 246 ± 219 * | 151 ± 79 | 234 ± 215 |
| L5 Onset | 1:42 ± 1:20 | 1:32 ± 1:02 | 0:54 ± 0:52 | 1:51 ± 1:14 * | 1:36 ± 1:14 | 1:39 ± 1:14 |
| IV | 0.894 ± 0.173 | 0.837 ± 0.156 | 0.881 ± 0.183 | 0.869 ± 0.165 | 0.820 ± 0.157 | 0.883 ± 0.169 |
| IS | 0.519 ± 0.098 | 0.584 ± 0.110 | 0.597 ± 0.121 | 0.529 ± 0.098 | 0.448 ± 0.122 | 0.565 ± 0.092 * |
| RA | 0.872 ± 0.073 | 0.905 ± 0.052 | 0.948 ± 0.019 | 0.867 ± 0.065 *** | 0.899 ± 0.057 | 0.882 ± 0.069 |
| CFI | 0.651 ± 0.057 | 0.695 ± 0.054 | 0.699 ± 0.056 | 0.660 ± 0.058 | 0.654 ± 0.065 | 0.671 ± 0.058 |
| Wrist temperature, °C | ||||||
| MESOR | 31.63 ± 0.53 | 32.07 ± 0.55 * | 32.26 ± 0.56 | 31.67 ± 0.51 | 31.55 ± 0.73 | 31.86 ± 0.53 |
| 24 h Amplitude | 1.49 ± 0.58 | 1.19 ± 0.44 * | 1.29 ± 0.52 | 1.40 ± 0.55 | 1.60 ± 0.51 | 1.33 ± 0.54 |
| Phase | 2:25 ± 1:39 | 3:16 ± 1:18 | 2:59:1:36 | 2:41:1:34 | 3:27 ± 1:02 | 2:36 ± 1:38 |
| Sleep characteristics, hh:mm | ||||||
| Bedtime | 22:42 ± 1:14 | 22:42 ± 0:54 | 22:28 ± 1:01 | 22:46 ± 1:12 | 22:58 ± 1:16 | 22:38 ± 1:05 |
| Wake time | 7:01 ± 1:31 | 6:52 ± 1:09 | 6:51 ± 2:09 | 6:59 ± 1:29 | 7:36 ± 2:06 | 6:49 ± 1:09 |
| Sleep phase | 2:51 ± 1:12 | 2:45 ± 0:54 | 2:39 ± 0:45 | 2:52 ± 1:10 | 3:17 ± 1:22 | 2:43 ± 0:57 |
| Time in bed | 8:17 ± 1:23 | 8:08 ± 0:58 | 8:20 ± 0:59 | 8:12 ± 1:18 | 8:38 ± 1:34 | 8:08 ± 1:09 |
| Total sleep | 7:18 ± 1:17 | 7:17 ± 0:58 | 7:34 ± 0:54 | 7:13 ± 1:14 | 7:50 ± 1:40 | 7:11 ± 1:01 |
| Sleep latency, min | 02.58 ± 2.35 | 02.21 ± 1.80 | 1.78 ± 1.58 | 2.63 ± 2.27 | 01.92 ± 2.36 | 02.55 ± 2.12 |
| Sleep efficiency, % | 88.67 ± 4.03 | 85.58 ± 6.24 | 88.15 ± 6.73 | 88.15 ± 6.73 | 88.59 ± 6.35 | 87.90 ± 4.79 |
| WASO | 0:54 ± 0:27 | 0:47 ± 0:18 | 0:42 ± 0:17 | 0:54 ± 0:25 | 0:45 ± 0:21 | 0:53 ± 0:24 |
| Blue light, μw/cm2 | ||||||
| MESOR | 13.00 ± 7.83 | 8.20 ± 6.36 | 8.10 ± 6.36 | 12.03 ± 7.83 | 16.98 ± 11.73 | 9.88 ± 5.90 ** |
| 24 h Amplitude | 18.61 ± 12.38 | 12.31 ± 8.30 | 12.18 ± 10.29 | 17.34 ± 11.46 | 25.21 ± 17.03 | 14.22 ± 8.77 ** |
| Phase | 12:46 ± 0:52 | 13:07 ± 0:55 | 13:27 ± 1:02 | 12:44 ± 0:48 * | 13:30 ± 1:18 | 12:48 ± 0:46 |
| M10 | 27.00 ± 16.91 | 18.00 ± 14.60 | 18.54 ± 14.60 | 24.96 ± 15.62 | 34.18 ± 21.55 | 21.21 ± 13.06 * |
| M10 Onset | 7:50 ± 0:46 | 8:01 ± 0:47 | 8:10 ± 0:51 | 7:49 ± 0:44 | 8:15 ± 0:55 | 7:50 ± 0:44 |
| L5 | 0.100 ± 0.169 | 0.024 ± 0.049 | 0.003 ± 0.006 | 0.091 ± 0.154 | 0.075 ± 0.158 | 0.070 ± 0.138 |
| L5 Onset | 1:34 ± 2:10 | 1:01 ± 1:01 | 0:29 ± 0:48 | 1:36 ± 1:57 | 1:13 ± 1:21 | 1:23 ± 1:55 |
| L5 log10 | −2.02 ± 1.28 | −2.60 ± 1.13 | −3.40 ± 0.88 | −1.90 ± 1.13 ** | −2.35 ± 1.43 | −2.22 ± 1.22 |
| IV | 0.915 ± 0.341 | 0.904 ± 0.345 | 1.030 ± 0.351 | 0.894 ± 0.334 | 1.077 ± 0.385 | 0.892 ± 0.324 |
| IS | 0.397 ± 0.153 | 0.382 ± 0.119 | 0.362 ± 0.134 | 0.400 ± 0.142 | 0.322 ± 0.181 | 0.406 ± 0.127 |
| RA | 0.980 ± 0.029 | 0.992 ± 0.014 | 0.999 ± 0.001 | 0.980 ± 0.026 * | 0.988 ± 0.022 | 0.984 ± 0.025 |
| DDIbl | 308 ± 88 | 394 ± 64 ** | 400 ± 96 | 324 ± 119 * | 282 ± 175 | 354 ± 100 |
| NEIbl | 2.81 ± 2.39 | 1.12 ± 1.50 *** | 0.89 ± 1.65 | 2.56 ± 2.29 ** | 2.55 ± 2.25 | 2.07 ± 2.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gubin, D.; Danilenko, K.; Stefani, O.; Kolomeichuk, S.; Markov, A.; Petrov, I.; Voronin, K.; Mezhakova, M.; Borisenkov, M.; Shigabaeva, A.; et al. Blue Light and Temperature Actigraphy Measures Predicting Metabolic Health Are Linked to Melatonin Receptor Polymorphism. Biology 2024, 13, 22. https://doi.org/10.3390/biology13010022
Gubin D, Danilenko K, Stefani O, Kolomeichuk S, Markov A, Petrov I, Voronin K, Mezhakova M, Borisenkov M, Shigabaeva A, et al. Blue Light and Temperature Actigraphy Measures Predicting Metabolic Health Are Linked to Melatonin Receptor Polymorphism. Biology. 2024; 13(1):22. https://doi.org/10.3390/biology13010022
Chicago/Turabian StyleGubin, Denis, Konstantin Danilenko, Oliver Stefani, Sergey Kolomeichuk, Alexander Markov, Ivan Petrov, Kirill Voronin, Marina Mezhakova, Mikhail Borisenkov, Aislu Shigabaeva, and et al. 2024. "Blue Light and Temperature Actigraphy Measures Predicting Metabolic Health Are Linked to Melatonin Receptor Polymorphism" Biology 13, no. 1: 22. https://doi.org/10.3390/biology13010022
APA StyleGubin, D., Danilenko, K., Stefani, O., Kolomeichuk, S., Markov, A., Petrov, I., Voronin, K., Mezhakova, M., Borisenkov, M., Shigabaeva, A., Yuzhakova, N., Lobkina, S., Weinert, D., & Cornelissen, G. (2024). Blue Light and Temperature Actigraphy Measures Predicting Metabolic Health Are Linked to Melatonin Receptor Polymorphism. Biology, 13(1), 22. https://doi.org/10.3390/biology13010022