
The Impact of HPP-Assisted Biocontrol Approach on the Bacterial Communities’ Dynamics and Quality Parameters of a Fermented Meat Sausage Model

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms and Inoculum Preparation
2.2. Preparation of Alheira Paste
2.3. High Hydrostatic Pressure Treatments
2.4. Quality Parameters of Fermented Meat Sausage over Refrigerated Storage
2.4.1. Color Analysis
2.4.2. Texture Analysis—Compression/Extrusion Tests
2.4.3. Thiobarbituric Acid Reactive Substances (TBARS)
2.5. Bacterial Communities’ Dynamics of Fermented Meat Sausage Model
2.5.1. Total DNA Extraction Procedure
2.5.2. 16S rRNA Gene Amplicon Sequencing and Library Preparation
2.6. Statistical Analysis
3. Results and Discussion
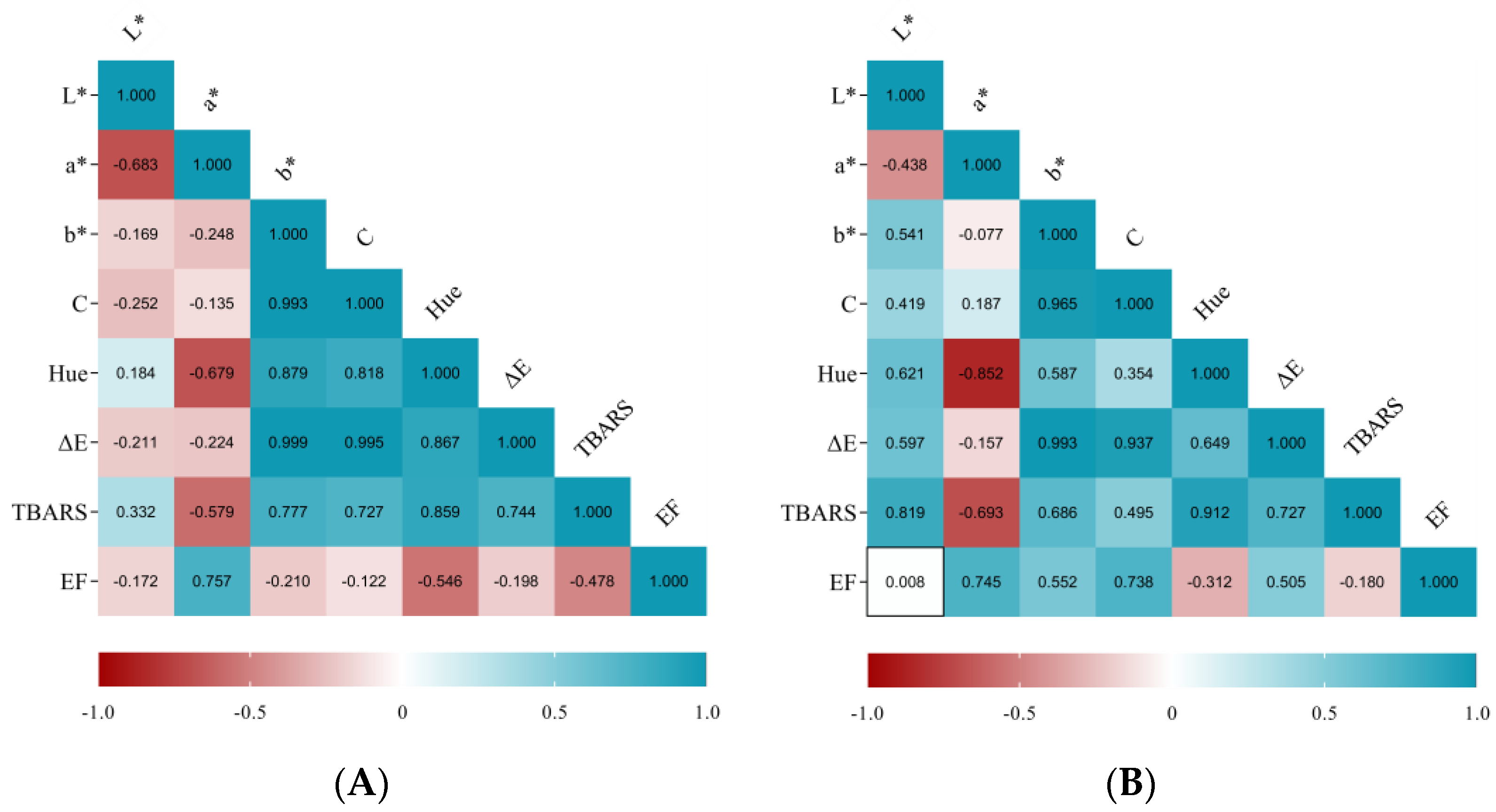
3.1. Instrumental Color Evaluation of Non- and HPP-Phage-LAB Treated Sausages
3.2. Minimal Processing Impact on Alheira Texture: Compression/Extrusion Tests
3.3. Lipid Peroxidation Evaluation through Thiobarbituric Acid Reactive Substances (TBARS) Assays
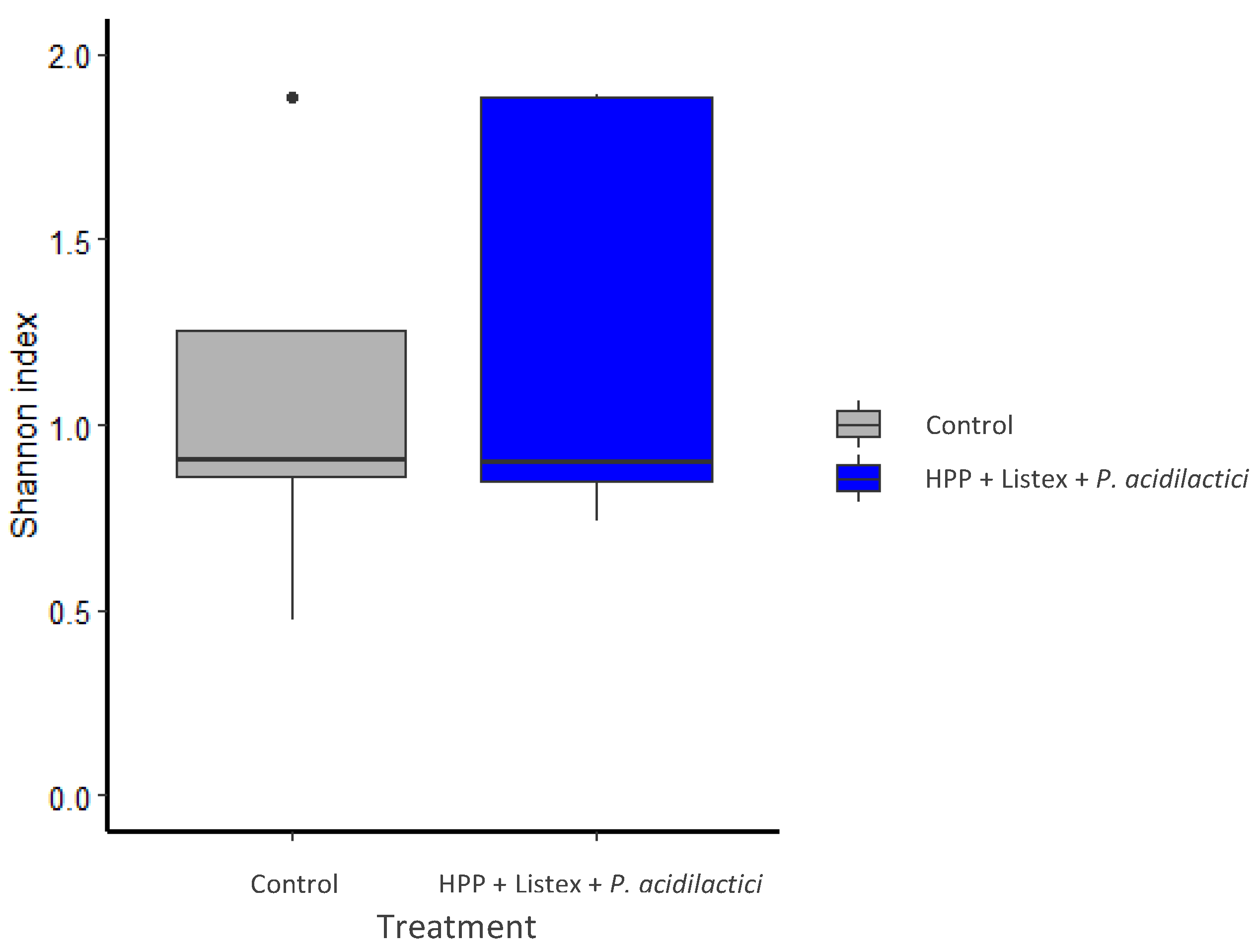
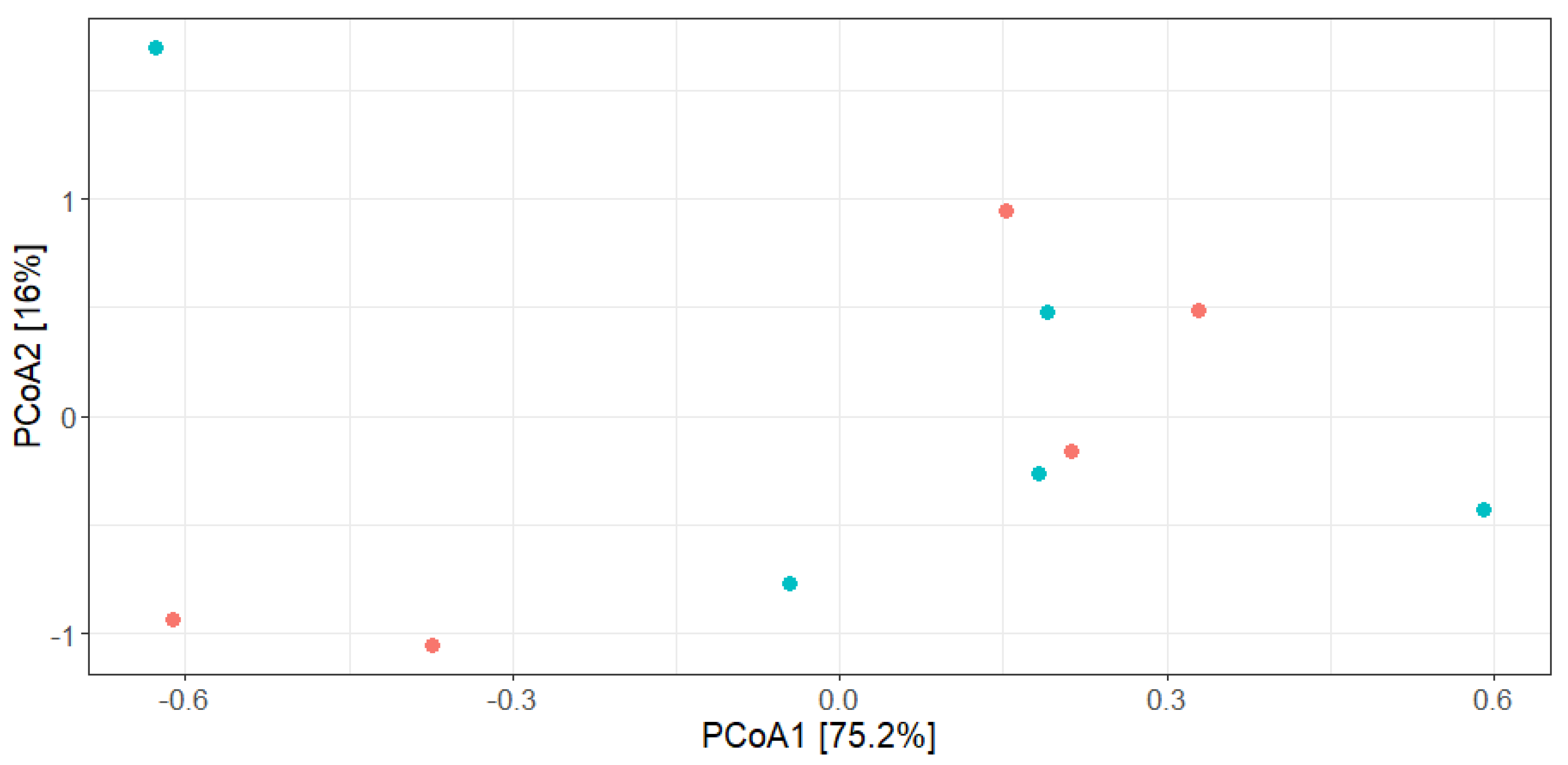
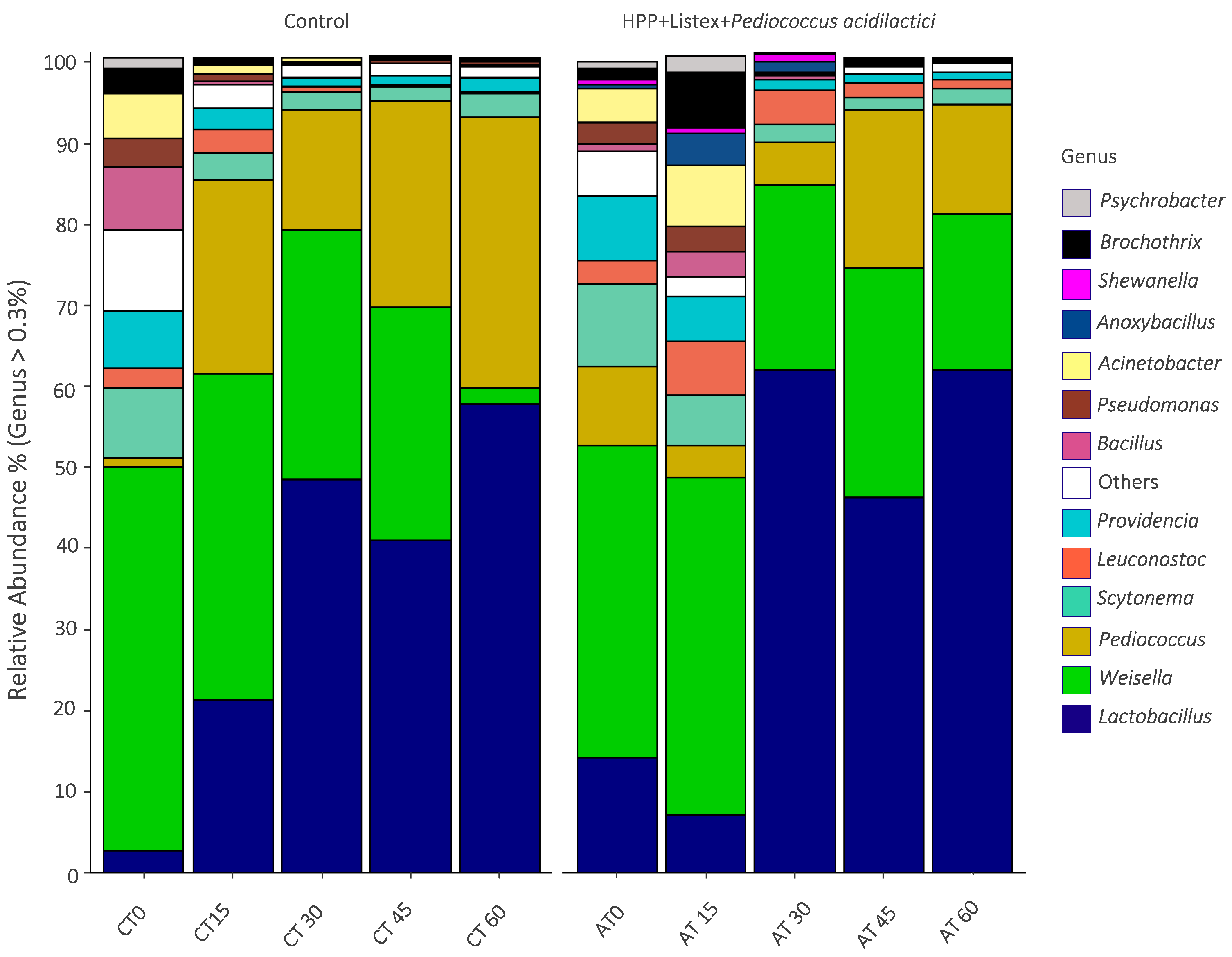
3.4. Impact of the Minimal Processing on Alheira Microbiota Dynamics Assessed by 16S rRNA Diversity Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fernández-Ferrín, P.; Calvo-Turrientes, A.; Bande, B.; Artaraz-Miñón, M.; Galán-Ladero, M.M. The valuation and purchase of food products that combine local, regional and traditional features: The influence of consumer ethnocentrism. Food Qual. Prefer. 2018, 64, 138–147. [Google Scholar] [CrossRef]
- Pieniak, Z.; Verbeke, W.; Vanhonacker, F.; Guerrero, L.; Hersleth, M. Association between traditional food consumption and motives for food choice in six European countries. Appetite 2009, 53, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Bardone, E.; Spalvēna, A. European Union food quality schemes and the transformation of traditional foods into European products in Latvia and Estonia. Appetite 2019, 135, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Kapelari, S.; Alexopoulos, G.; Moussouri, T.; Sagmeister, K.J.; Stampfer, F. Food heritage makes a difference: The importance of cultural knowledge for improving education for sustainable food choices. Sustainability 2020, 12, 1509. [Google Scholar] [CrossRef]
- Guerrero, L.; Guàrdia, M.D.; Xicola, J.; Verbeke, W.; Vanhonacker, F.; Zakowska-Biemans, S.; Sajdakowska, M.; Sulmont-Rossé, C.; Issanchou, S.; Contel, M.; et al. Consumer-driven definition of traditional food products and innovation in traditional foods. A qualitative cross-cultural study. Appetite 2009, 52, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Ben Slima, S.; Ktari, N.; Triki, M.; Trabelsi, I.; Abdeslam, A.; Moussa, H.; Makni, I.; Herrero, A.M.; Jiménez-Colmenero, F.; Ruiz-Capillas, C.; et al. Effects of probiotic strains, Lactobacillus plantarum TN8 and Pediococcus acidilactici, on microbiological and physico-chemical characteristics of beef sausages. LWT-Food. Sci. Technol. 2018, 92, 195–203. [Google Scholar] [CrossRef]
- Blázquez, I.O.; Burgos, M.J.G.; Pérez-Pulido, R.; Gálvez, A. Treatment With High-Hydrostatic Pressure, Activated Film Packaging With Thymol Plus Enterocin AS-48, and Its Combination Modify the Bacterial Communities of Refrigerated Sea Bream (Sparus aurata) Fillets. Front. Microbiol. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Baños, A.; García-López, J.D.; Núñez, C.; Martínez-Bueno, M.; Maqueda, M.; Valdivia, E. Biocontrol of Listeria monocytogenes in fish by enterocin AS-48 and Listeria lytic bacteriophage P100. LWT-Food Sci. Technol. 2016, 66, 672–677. [Google Scholar] [CrossRef]
- Rendueles, C.; Escobedo, S.; Rodríguez, A.; Martínez, B. Bacteriocin-phage interaction (BaPI): Phage predation of Lactococcus in the presence of bacteriocins. MicrobiologyOpen 2022, 11, e1308. [Google Scholar] [CrossRef] [PubMed]
- Aganovic, K.; Hertel, C.; Vogel, R.F.; Johne, R.; Schlüter, O.; Schwarzenbolz, U.; Jäger, H.; Holzhauser, T.; Bergmair, J.; Roth, A.; et al. Aspects of high hydrostatic pressure food processing: Perspectives on technology and food safety. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3225–3266. [Google Scholar] [CrossRef]
- Komora, N.; Maciel, C.; Pinto, C.A.; Ferreira, V.; Brandão, T.R.; Saraiva, J.M.; Castro, S.M.; Teixeira, P. Non-thermal approach to Listeria monocytogenes inactivation in milk: The combined effect of high pressure, pediocin PA-1 and bacteriophage P100. Food Microbiol. 2020, 86, 103315. [Google Scholar] [CrossRef]
- Ferreira, V.; Barbosa, J.; Silva, J.; Felício, M.T.; Mena, C.; Hogg, T.; Gibbs, P.; Teixeira, P. Characterisation of alheiras, traditional sausages produced in the North of Portugal, with respect to their microbiological safety. Food Control 2007, 18, 436–440. [Google Scholar] [CrossRef]
- Fusco, V.; Besten HM den Logrieco, A.F.; Rodriguez, F.P.; Skandamis, P.N.; Stessl, B.; Teixeira, P. Food safety aspects on ethnic foods: Toxicological and microbial risks. Curr. Opin. Food Sci. 2015, 6, 24–32. [Google Scholar] [CrossRef]
- Albano Helena Todorov, S.D.; van Reenen, C.A.; Hogg, T.; Dicks, L.M.T.; Teixeira, P. Characterization of two bacteriocins produced by Pediococcus acidilactici isolated from “Alheira”, a fermented sausage traditionally produced in Portugal. Int. J. Food Microbiol. 2007, 116, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Castro, S.M.; Kolomeytseva, M.; Casquete, R.; Silva, J.; Saraiva, J.A.; Teixeira, P. Effect of high pressure on growth and bacteriocin production of Pediococcus acidilactici HA-6111-2. High Press. Res. 2015, 35, 405–418. [Google Scholar] [CrossRef]
- Komora, N.; Maciel, C.; Amaral, R.; Fernandes, R.; Marília Castro, S.; Saraiva, J.A.; Teixeira, P. Innovative hurdle system towards Listeria monocytogenes inactivation in a fermented meat sausage model—High pressure processing assisted by bacteriophage P100 and bacteriocinogenic Pediococcus acidilactici. Food Res. Int. 2021, 148, 110628. [Google Scholar] [CrossRef] [PubMed]
- Komora, N.; Bruschi, C.; Ferreira, V.; Maciel, C.; Brandão, T.R.S.; Fernandes, R.; Saraiva, J.A.; Castro, S.M.; Teixeira, P. The protective effect of food matrices on Listeria lytic bacteriophage P100 application towards high pressure processing. Food Microbiol. 2018, 76, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Cruxen, C.E.d.S.; Braun, C.L.K.; Fagundes, M.B.; Gularte, M.A.; Wagner, R.; Padilha da Silva, W.; Fiorentini, Â.M. Development of fermented sausage produced with mutton and native starter cultures. LWT-Food. Sci. Technol. 2018, 95, 23–31. [Google Scholar] [CrossRef]
- Tomasevic, I.; Tomovic, V.; Milovanovic, B.; Lorenzo, J.; Đorđević, V.; Karabasil, N.; Djekic, I. Comparison of a computer vision system vs. traditional colorimeter for color evaluation of meat products with various physical properties. Meat Sci. 2019, 48, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Arslan, B.; Soyer, A. Effects of chitosan as a surface fungus inhibitor on microbiological, physicochemical, oxidative and sensory characteristics of dry fermented sausages. Meat Sci. 2018, 145, 107–113. [Google Scholar] [CrossRef]
- Chaillou, S.; Chaulot-Talmon, A.; Caekebeke, H.; Cardinal, M.; Christieans, S.; Denis, C.; Hélène Desmonts, M.; Dousset, X.; Feurer, C.; Hamon, E.; et al. Origin and ecological selection of core and food-specific bacterial communities associated with meat and seafood spoilage. ISME J. 2015, 9, 1105–1118. [Google Scholar] [CrossRef]
- Quijada, N.M.; De Filippis, F.; Sanz, J.J.; García-Fernández, M.d.C.; Rodríguez-Lázaro, D.; Ercolini, D.; Hernández, M. Different Lactobacillus populations dominate in “Chorizo de León” manufacturing performed in different production plants. Food Microbiol. 2018, 70, 94–102. [Google Scholar] [CrossRef]
- Ferrocino, I.; Bellio, A.; Giordano, M.; Macori, G.; Romano, A.; Rantsiou, K. Shotgun Metagenomics and Volatilome Profile of the Microbiota of Fermented Sausages. Appl. Environ. Microbiol. 2018, 84, e02120–17. [Google Scholar] [CrossRef]
- Macieira, A.; Albano, H.; Pinto, M.; Linheiro, R.; Barbosa, J.; Teixeira, P. Evaluation of a bacteriocinogenic Lactobacillus plantarum strain on the microbiological characteristics of “Alheira de Vitela”. AIMS Agric. Food 2019, 4, 223–236. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis (Use R!), 2nd ed.; Springer Nature: New York, NY, USA, 2016; pp. 1–260. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Cava, R.; García-Parra, J.; Ladero, L. Effect of high hydrostatic pressure processing and storage temperature on food safety, microbial counts, colour and oxidative changes of a traditional dry-cured sausage. LWT-Food. Sci. Technol. 2020, 128, 109462. [Google Scholar] [CrossRef]
- Rubio, B.; Martínez, B.; García-Cachán, M.D.; Rovira, J.; Jaime, I. The effects of high pressure treatment and of storage periods on the quality of vacuum-packed “salchichón” made of raw material enriched in monounsaturated and polyunsaturated fatty acids. Innov. Food Sci. Emerg. Technol. 2007, 8, 180–187. [Google Scholar] [CrossRef]
- Cava Ramón Ladero, L.; González, S.; Carrasco, A.; Ramírez, M.R. Effect of pressure and holding time on colour, protein and lipid oxidation of sliced dry-cured Iberian ham and loin during refrigerated storage. Innov. Food Sci. Emerg. Technol. 2009, 10, 76–81. [Google Scholar] [CrossRef]
- Van Ba, H.; Seo, H.-W.; Kim, J.-H.; Cho, S.-H.; Kim, Y.-S.; Ham, J.-S.; Park, B.-Y.; Kim, H.-W.; Kim, T.-B.; Seong, P.-N. The effects of starter culture types on the technological quality, lipid oxidation and biogenic amines in fermented sausages. LWT-Food. Sci. Technol. 2016, 74, 191–198. [Google Scholar] [CrossRef]
- Rigdon, M.; Thippareddi, H.; McKee, R.W.; Thomas, C.L.; Stelzleni, A.M. Texture of fermented summer sausage with differing pH, endpoint temperature, and high pressure processing times. Meat Muscle Biol. 2020, 4, 1–11. [Google Scholar] [CrossRef]
- Casquete, R.; Benito, M.J.; Martín, A.; Ruiz-Moyano, S.; Pérez-Nevado, F.; Córdoba, M.G. Comparison of the effects of a commercial and an autochthonous Pediococcus acidilactici and Staphylococcus vitulus starter culture on the sensory and safety properties of a traditional Iberian dry-fermented sausage “salchichón”. Int. J. Food Sci. Technol. 2012, 47, 1011–1019. [Google Scholar] [CrossRef]
- Wang, X.; Ren, H.; Wang, W.; Xie, Z.J. Effects of a Starter Culture on Histamine Reduction, Nitrite Depletion and Oxidative Stability of Fermented Sausages. J. Food Saf. 2016, 36, 195–202. [Google Scholar] [CrossRef]
- Ripoll, G.; Joy, M.; Muñoz, F. Use of dietary vitamin E and selenium (Se) to increase the shelf life of modified atmosphere packaged light lamb meat. Meat Sci. 2011, 87, 88–93. [Google Scholar] [CrossRef] [PubMed]
- GB-T 9959.2-2008; Fresh and Frozen Pork Lean, Cuts. National Standards of People’s Republic of China: Beijing, China, 2008.
- Campo, M.M.; Nute, G.R.; Hughes, S.I.; Enser, M.; Wood, J.D.; Richardson, R.I. Flavour perception of oxidation in beef. Meat Sci. 2006, 72, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Wenjiao, F.; Yongkui, Z.; Yunchuan, C.; Junxiu, S.; Yuwen, Y. TBARS predictive models of pork sausages stored at different temperatures. Meat Sci. 2014, 96, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Guyon, C.; Meynier, A.; De Lamballerie, M. Protein and lipid oxidation in meat: A review with emphasis on high-pressure treatments. Trends Food Sci. Technol. 2016, 50, 131–143. [Google Scholar] [CrossRef]
- Liu, G.; Wang, Y.; Gui, M.; Zheng, H.; Dai, R.; Li, P. Combined effect of high hydrostatic pressure and enterocin LM-2 on the refrigerated shelf life of ready-to-eat sliced vacuum-packed cooked ham. Food Control 2012, 24, 64–71. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Martín, A.; Benito, M.J.; Hernández, A.; Casquete, R.; de Guia Córdoba, M. Application of Lactobacillus fermentum HL57 and Pediococcus acidilactici SP979 as potential probiotics in the manufacture of traditional Iberian dry-fermented sausages. Food Microbiol. 2011, 28, 839–847. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Ye, H.; Wang, Z.; Wu, X.; Han, Y.; Xu, B. Changes in the microbial communities in vacuum-packaged smoked bacon during storage. Food Microbiol. 2018, 77, 26–37. [Google Scholar] [CrossRef]
- Zhang, Y.; Qin, Y.; Wang, Y.; Huang, Y.; Li, P.; Li, P. Lactobacillus plantarum LPL-1, a bacteriocin producing strain, changed the bacterial community composition and improved the safety of low-salt fermented sausages. LWT-Food. Sci. Technol. 2020, 128, 109385. [Google Scholar] [CrossRef]
- Albano, H.; Henriques, I.; Correia, A.; Hogg, T.; Teixeira, P. Characterization of microbial population of ‘Alheira’ (a traditional Portuguese fermented sausage) by PCR-DGGE and traditional cultural microbiological methods. J. Appl. Microbiol. 2008, 105, 2187–2194. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Huang, J.; Wu, X.; Xu, Y.; Fan, J.; Wu, C.; Jin, Y. Characterizing the metabolites and the microbial communities of the soy sauce mash affected by temperature and hydrostatic pressure. Food Res. Int. 2019, 123, 801–808. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, P.; Wu, J.; Tao, D.; Wu, R. Effects of Leuconostoc mesenteroides on physicochemical and microbial succession characterization of soybean paste, Da-jiang. LWT-Food. Sci. Technol. 2019, 115, 108028. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Time (Days) | Color Parameters | TBARS (mg kg−1) | Extrusion Force (N g−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Lightness (L*) | Redness (a*) | Yellowness (b*) | Hue Angle (h°) | Chroma (Cab) | ΔE | ||||
| 0 | C | 59.60 ± 0.84 | 13.15 ± 0.94 | 28.49 ± 2.99 | 65.17 ± 0.82 | 31.38 ± 3.10 | 0 | 0.89 ± 0.09 | 0.65 ± 0.02 |
| HPP+Listex+LAB | 58.38 ± 0.82 | 13.24 ± 0.54 | 29.85 ± 2.53 | 66.03 ± 0.95 | 32.66 ± 2.53 | 0 | 0.87 ± 0.07 | 0.61 ± 0.02 | |
| 15 | C | 57.82 ± 0.73 | 13.82 ± 0.89 | 32.72 ± 2.13 | 67.09 ± 0.65 | 35.52 ± 2.27 | 4.68 ± 2.29 | 0.96 ± 0.10 | 0.66 ± 0.09 |
| HPP+Listex+LAB | 58.39 ± 1.13 | 14.43 ± 0.67 | 33.51 ± 2.02 | 66.69 ± 0.43 | 36.48 ± 2.11 | 3.93 ± 2.19 | 0.95 ± 0.05 | 0.68 ± 0.01 | |
| 30 | C | 58.53 ± 0.32 | 12.57 ± 1.10 | 32.22 ± 0.75 | 68.71 ± 1.26 | 34.59 ± 1.08 | 4.05 ± 0.58 | 1.11 ± 0.06 | 0.62 ± 0.04 |
| HPP+Listex+LAB | 58.23 ± 0.41 | 11.85 ± 0.29 | 32.10 ± 0.57 | 69.74 ± 0.53 | 34.22 ± 0.56 | 2.69 ± 0.49 | 1.09 ± 0.04 | 0.61 ± 0.02 | |
| 45 | C | 59.91 ± 0.65 | 12.22 ± 1.13 | 33.22 ± 2.89 | 69.81 ± 0.22 | 35.39 ± 3.10 | 5.04 ± 2.63 | 1.17 ± 0.03 | 0.64 ± 0.01 |
| HPP+Listex+LAB | 59.56 ± 1.21 | 12.28 ± 1.08 | 33.15 ± 2.95 | 69.68 ± 0.02 | 35.35 ± 3.14 | 4.09 ± 2.44 | 1.23 ± 0.05 | 0.64 ± 0.04 | |
| 60 | C | 59.43 ± 1.24 | 12.82 ± 1.16 | 33.78 ± 1.47 | 69.21 ± 1.94 | 36.15 ± 1.43 | 5.49 ± 1.43 | 1.34 ± 0.02 | 0.64 ± 0.05 |
| HPP+Listex+LAB | 59.41 ± 1.26 | 12.01 ± 0.98 | 33.82 ± 1.43 | 70.47 ± 0.70 | 35.89 ± 1.68 | 4.47 ± 1.44 | 1.41 ± 0.03 | 0.62 ± 0.04 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komora, N.; Maciel, C.; Isidro, J.; Pinto, C.A.; Fortunato, G.; Saraiva, J.M.A.; Teixeira, P. The Impact of HPP-Assisted Biocontrol Approach on the Bacterial Communities’ Dynamics and Quality Parameters of a Fermented Meat Sausage Model. Biology 2023, 12, 1212. https://doi.org/10.3390/biology12091212
Komora N, Maciel C, Isidro J, Pinto CA, Fortunato G, Saraiva JMA, Teixeira P. The Impact of HPP-Assisted Biocontrol Approach on the Bacterial Communities’ Dynamics and Quality Parameters of a Fermented Meat Sausage Model. Biology. 2023; 12(9):1212. https://doi.org/10.3390/biology12091212
Chicago/Turabian StyleKomora, Norton, Cláudia Maciel, Joana Isidro, Carlos A. Pinto, Gianuario Fortunato, Jorge M. A. Saraiva, and Paula Teixeira. 2023. "The Impact of HPP-Assisted Biocontrol Approach on the Bacterial Communities’ Dynamics and Quality Parameters of a Fermented Meat Sausage Model" Biology 12, no. 9: 1212. https://doi.org/10.3390/biology12091212
APA StyleKomora, N., Maciel, C., Isidro, J., Pinto, C. A., Fortunato, G., Saraiva, J. M. A., & Teixeira, P. (2023). The Impact of HPP-Assisted Biocontrol Approach on the Bacterial Communities’ Dynamics and Quality Parameters of a Fermented Meat Sausage Model. Biology, 12(9), 1212. https://doi.org/10.3390/biology12091212