
Cerianthus lloydii (Ceriantharia: Anthozoa: Cnidaria): New Status and New Perspectives †

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling and Identification
2.2. CO1 and 18S Sequence Acquisition
2.3. Phylogenetic Analyses
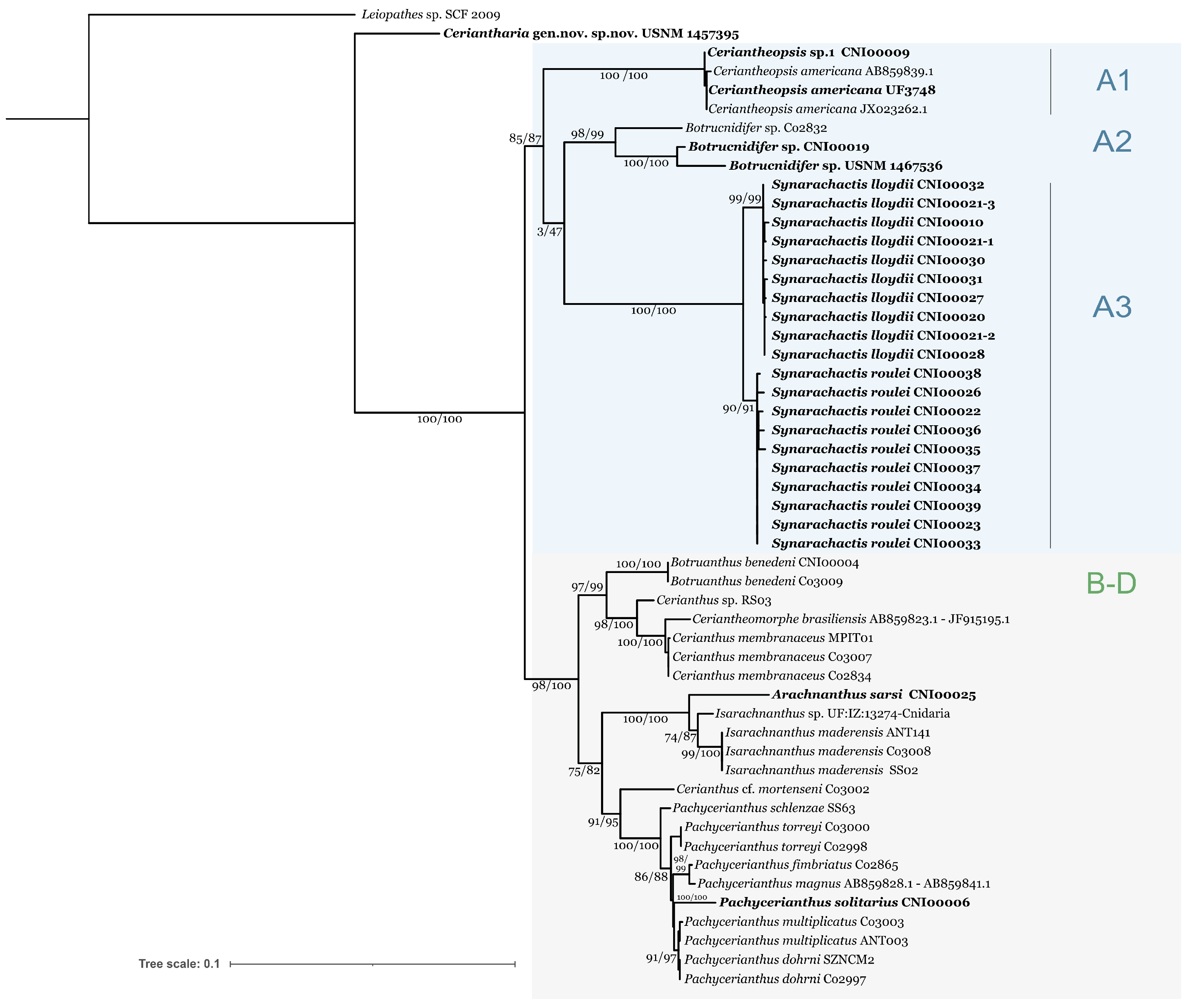
3. Results
3.1. Phylogenetic Analyses
3.2. Species in A3 Subclade
4. Discussion
4.1. Phylogeny of Ceriantharia
4.2. Taxonomic Implications
4.2.1. Synarachnactis Carlgren 1924 (Amended)
4.2.2. Synarachnactidae fam. nov. Molodtsova and Simakova, 2023
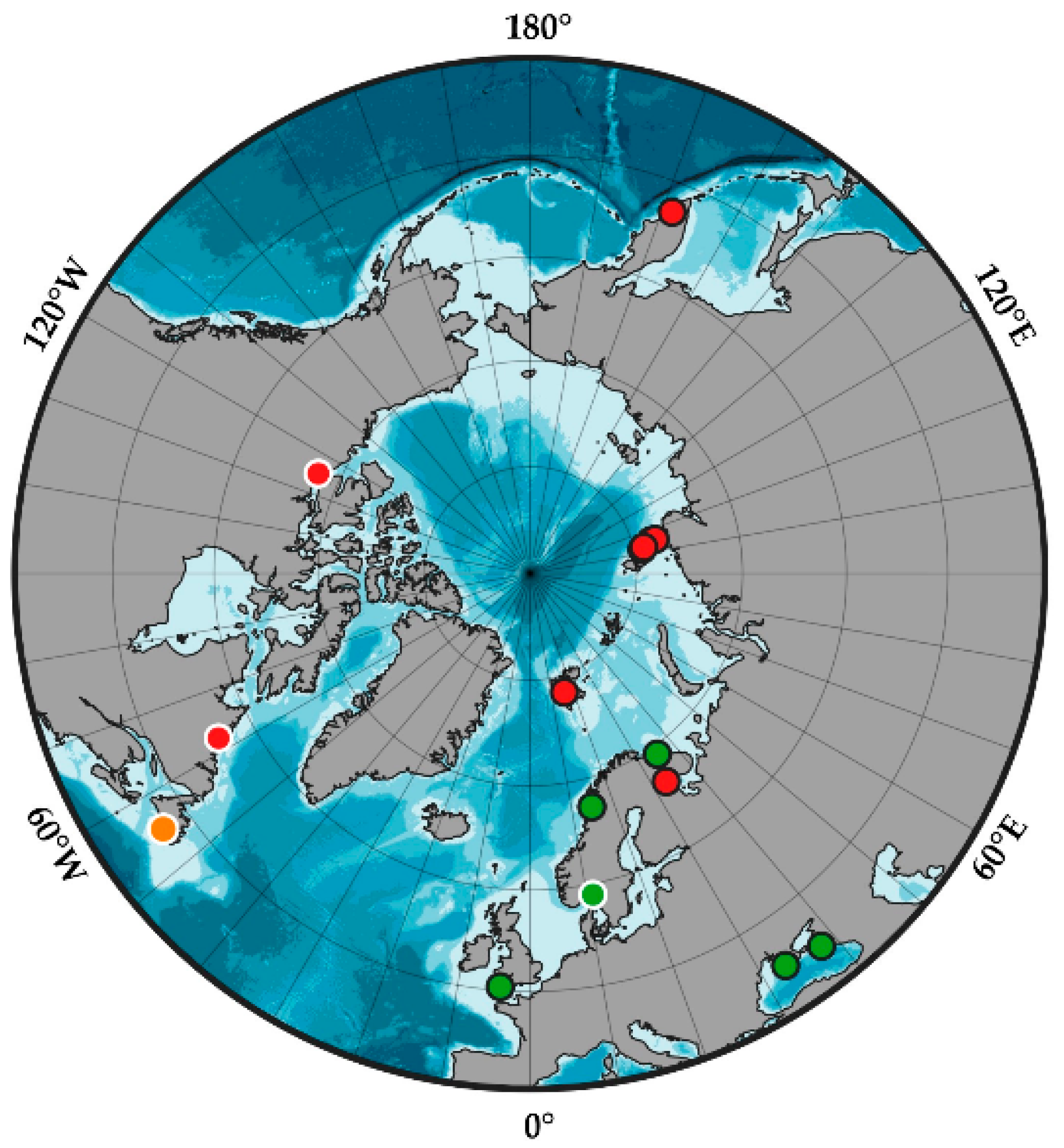
4.3. Geographic Distribution of the Genus Synarachnactis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daly, M.; Brugler, M.R.; Cartwright, P.; Collins, A.G.; Dawson, M.N.; Fautin, D.G.; France, S.C.; McFadden, C.S.; Romano, S.L. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa 2007, 1668, 127–182. [Google Scholar] [CrossRef]
- McFadden, C.S.; Quattrini, A.M.; Brugler, M.R.; Cowman, P.F.; Dueñas, L.F.; Kitahara, M.V.; Paz-García, D.A.; Reimer, J.D.; Rodríguez, E. Phylogenomics, origin, and diversification of Anthozoans (Phylum Cnidaria). Syst. Biol. 2021, 70, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Forero Mejia, A.C.; Molodtsova, T.; Östman, C.; Bavestrello, G.; Rouse, G.W. Molecular phylogeny of Ceriantharia (Cnidaria: Anthozoa) reveals non-monophyly of traditionally accepted families. Zool. J. Linn. Soc. 2020, 190, 397–416. [Google Scholar] [CrossRef]
- Stampar, S.N.; Reimer, J.D.; Maronna, M.M.; Lopes, C.S.; Ceriello, H.; Santos, T.B.; Acuña, F.H.; Morandini, A.C. Ceriantharia (Cnidaria) of the World: An annotated catalogue and key to species. ZooKeys 2020, 952, 1. [Google Scholar] [CrossRef] [PubMed]
- Stampar, S.N.; Beneti, J.S.; Acuña, F.H.; Morandini, A.C. Ultrastructure and tube formation in Ceriantharia (Cnidaria, Anthozoa). Zool. Anz. 2015, 254, 67–71. [Google Scholar] [CrossRef]
- Mariscal, R.N.; Conclin, E.J.; Bigger, C.H. The ptychocyst, a major new category of cnida used in tube construction by a cerianthid anemone. Biol. Bull. 1977, 152, 392–405. [Google Scholar] [CrossRef]
- Malakhov, V.V. Symmetry and the tentacular apparatus in Cnidaria. Russ. J. Mar. Biol. 2016, 42, 287–298. [Google Scholar] [CrossRef]
- Stampar, S.N.; Maronna, M.M.; Kitahara, M.V.; Reimer, J.D.; Morandini, A.C. Fast-evolving mitochondrial DNA in Ceriantharia: A reflection of hexacorallia paraphyly? PLoS ONE 2014, 9, e86612. [Google Scholar] [CrossRef]
- Molodtsova, T.N. Order Ceriantharia Perrier, 1883—Tube anemones. In Cnidaria. Ctenophora. Illustrated Keys to Free-Living Invertebrates of Eurasian Arctic Seas and Adjacent Deep Waters; Sirenko, B., Ed.; KMK: Moscow, Russia, 2012; Volume 3, pp. 182–187. [Google Scholar]
- Verrill, A., 3rd. The Actinaria of the Canadian Arctic Expeditions, with notes on interesting species from Hudson Bay and other Canadian localities. Rep. Can. Arct. Exped. 1913–18 1922, 8, 89–164. [Google Scholar]
- Molodtsova, T.; Malakhov, V. Cerianthus lloydii (Anthozoa, Ceriantharia) from volcanic ecosystem of Kraternaya Bay. I. Morphology and anatomy of adult polyps, geographical distribution. Zool. Zhurnal 1995, 74, 5–17. [Google Scholar]
- Molodtsova, T.N. Ceriantharia (Cnidaria: Anthozoa) from the Faroe Islands. Fródskaparrit 2004, 51, 292–297. [Google Scholar]
- Carlgren, O. Ceriantharia. Dan. Ingolf-Exp. 1912, 5, 1–72. [Google Scholar]
- Molodtsova, T.N. Deep-sea fauna of european seas: An annotated species check-list of benthic invertebrates living deeper than 2000 m in the seas bordering Europe. Ceriantharia. Zool. Bespozvon. 2014, 11, 99–100. [Google Scholar]
- Molodtsova, T.; Malakhov, V. Cerianthus lloydii (Anthozoa, Ceriantharia) from the volcanic ecosystem of Kraternaya bay. 2. Larval development. Zool. Zhurnal 1995, 74, 4–11. [Google Scholar]
- Molodtsova, T.N.; Griffiths, C.L.; Acuña, F.H. A new species of shallow-water cerianthid (Cnidaria: Anthozoa) from South Africa, with remarks on the genus Ceriantheopsis. Afr. Nat. Hist. 2011, 7, 1–8. [Google Scholar]
- Stampar, S.N.; Scarabino, F.; Pastorino, G.; Morandini, A.C. A new species of tube-dwelling anemone (Cnidaria, Anthozoa, Ceriantharia, Ceriantheopsis) from the warm temperate South-western Atlantic. J. Mar. Biol. Assoc. UK 2015, 96, 1475–1481. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Meyer, C.P. Molecular systematics of cowries (Gastropoda: Cypraeidae) and diversification patterns in the tropics. Biol. J. Linn. Soc. 2003, 79, 401–459. [Google Scholar] [CrossRef]
- Norén, M.; Jondelius, U. Phylogeny of the Prolecithophora (Platyhelminthes) inferred from 18S rDNA sequences. Cladistics 1999, 15, 103–112. [Google Scholar] [CrossRef]
- Jørgensen, A.; Faurby, S.; Hansen, J.G.; Møbjerg, N.; Kristensen, R.M. Molecular phylogeny of Arthrotardigrada (Tardigrada). Mol. Phylogenet. Evol. 2010, 54, 1006–1015. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2012, 41, D36–D42. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Zardoya, R.; Telford, M.J. TranslatorX: Multiple alignment of nucleotide sequences guided by amino acid translations. Nucleic Acids Res. 2010, 38, W7–W13. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Roule, L. Description des Antipathaires et Cérianthaires recueillis par SAS le Prince de Monaco dans l’Atlantique nord, 1886–1902. Rés. Camp. Sci. Prince Albert Ier. 1905, 30, 1–99. [Google Scholar]
- Molodtsova, T. World List of Ceriantharia. Ceriantharia. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=1361 (accessed on 21 July 2023).
- Quattrini, A.M.; Snyder, K.E.; Purow-Ruderman, R.; Seiblitz, I.G.; Hoang, J.; Floerke, N.; Ramos, N.I.; Wirshing, H.H.; Rodriguez, E.; McFadden, C.S. Mito-nuclear discordance within Anthozoa, with notes on unique properties of their mitochondrial genomes. Sci. Rep. 2023, 13, 7443. [Google Scholar] [CrossRef]
- Budaeva, N.; Rogacheva, A. Colonization of the Arctic Ocean by two cosmopolitan genera of marine invertebrates. Zool. Bespozvon. 2013, 10, 127–142. [Google Scholar] [CrossRef]
- Laakkonen, H.M.; Hardman, M.; Strelkov, P.; Väinölä, R. Cycles of trans-Arctic dispersal and vicariance, and diversification of the amphi-boreal marine fauna. J. Evol. Biol. 2021, 34, 73–96. [Google Scholar] [CrossRef]
- Agassiz, A.E.R. On Arachnactis brachiolata, a species of floating Actinia found at Nahant, Massachusetts. J. Boston Soc. Nat. Hist. 1863, 7, 525–531. [Google Scholar]
- Kolyuchkina, G.A.; Syomin, V.L.; Simakova, U.V.; Sergeeva, N.G.; Ananiev, R.A.; Dmitrevsky, N.N.; Lyubimov, I.V.; Zenina, M.A.; Podymov, O.I.; Basin, A.B. Benthic community structure near the margin of the oxic zone: A case study on the Black Sea. J. Mar. Syst. 2022, 227, 103691. [Google Scholar] [CrossRef]
- Kiseleva, M. Benthos of the Soft Bottoms of the Black Sea; Naukova Dumka: Kiev, Ukraine, 1981. [Google Scholar]
- Zaika, V.; Kiseleva, M.; Mikhailova, T.; Makkaveeva, E. Mnogoletnie izmeneniya zoobentosa Chernogo morya [Long-term Changes in the Zoobenthos of the Black Sea]; Naukova Dumka: Kiev, Ukraine, 1992. [Google Scholar]
- Chikina, M.V.; Kucheruk, N.V. Contemporary dynamics of coastal benthic communities of the north Caucasian coast of the Black Sea. In Proceedings of the International Workshop on the Black Sea Benthos, Istanbul, Turkey, 19–23 April 2004; pp. 155–160. [Google Scholar]
- Hubenov, Z. Species composition of the free living multicellular invertebrate animals (Metazoa: Invertebrata) from the Bulgarian sector of the Black Sea and the coastal brackish basins. Hist. Nat. Bulg. 2015, 21, 49–168. [Google Scholar]
- Luth, U. The benthos of the oxic/anoxic interface in the western Black Sea: Comparative macro-and meiofauna investigations on transects from the Ukrainian, Romanian and Turkish shelf. In Proceedings of the International Workshop on Black Sea Benthos, Istanbul, Turkey, 19–23 April 2004; pp. 2–69. [Google Scholar]
- Mazlumyan, S.; Boltachova, N. Long-term variations in macrobenthos diversity at the Istanbul Strait’s (Bosporus) outlet area of the Black Sea. Ecol. Montenegrina 2017, 14, 80–91. [Google Scholar] [CrossRef]
- Forbes, E. On two remarkable Marine Invertebrate inhabiting the Aegean Sea. Ann. Mag. Nat. 1842, 8, 48–54. [Google Scholar]
- Forbes, E.V. Retrospective Comments. Ann. Mag. Nat. 1843, 12, 40–42. [Google Scholar] [CrossRef]
- Haime, J. Memoire sur le cérianthe (Cerianthus membranaceus). Ann. Sci. Nat. 1854, 1, 341–389. [Google Scholar]
- Andres, A. Le attinie. Fauna und Flora des Golfes von Neapel: Und der angrenzenden Meeres-Abschnitte; W. Engelmann: Leipzig, Germany, 1884; Volume 9, pp. 1–459. [Google Scholar] [CrossRef]
- Murray, J. On the deposits of the Black Sea. Scott. Geogr. Mag. 1900, 16, 673–702. [Google Scholar] [CrossRef]
- Zernov, S. K voprosu ob izuchenii zhizni Chernogo morya (On the Issue of Studying the Life of the Black Sea). Zap. Imp. Akad. Nauk. Fiz.-Mat. Otd. 1913, 32, 1–299. [Google Scholar]
- Brückner, H.; Kelterbaum, D.; Marunchak, O.; Porotov, A.; Vött, A. The Holocene sea level story since 7500 BP–Lessons from the Eastern Mediterranean, the Black and the Azov Seas. Quat. Int. 2010, 225, 160–179. [Google Scholar] [CrossRef]
- Ivanova, E.V.; Marret, F.; Zenina, M.A.; Murdmaa, I.O.; Chepalyga, A.L.; Bradley, L.R.; Schornikov, E.I.; Levchenko, O.V.; Zyryanova, M.I. The Holocene Black Sea reconnection to the Mediterranean Sea: New insights from the northeastern Caucasian shelf. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 427, 41–61. [Google Scholar] [CrossRef]
- Kurt, O.; Öztürk, S. Observation of Marine Areas (Çandarlı and Gökova Bays) and their biodiversity. Sak. Univ. J. Sci. 2022, 26, 213–223. [Google Scholar] [CrossRef]
- Cinar, M.E.; Yokeş, M.B.; Açik, Ş.; Bakir, A.K. Checklist of Cnidaria and Ctenophora from the coasts of Turkey. Turk. Zool. Derg. 2014, 38, 677–697. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Direction | Sequence 5′–3′ | PCR Scheme | Reference |
|---|---|---|---|---|---|
| COI | LCO1490 | F | GGTCAACAAATCATAAAGATATTGG | at 95 °C for 15 s, annealing at 50 °C for 30 s, and extension at 72 °C for 45 s, for 5 cycles, at 95 °C for 15 s, annealing at 52 °C for 30 s, and extension at 72 °C for 45 s, for 30 cycles | [18] |
| HCO2198 | R | TAAACTTCAGGGTGACCAAAAAATCA | [18] | ||
| dgLCO1490 | F | GGTCAACAAATCATAAAGAYATYGG | [19] | ||
| dgHCO2198 | R | TAAACTTCAGGGTGACCAAARAAYCA | [19] | ||
| 18S | TimA | F | AMCTGGTTGATCCTGCCAG | at 95 °C for 15 s, annealing at 55 °C for 30 s, and extension at 72 °C for 60 s for 35 cycles | [20] |
| 1100R2 | R | CGGTATCTGATCGTCTTCGA | [20] | ||
| 18S–5F | F | GCGAAAGCATTTGCCAAGAA | [21] | ||
| 18S–9R | R | GATCCTTCCGCAGGTTCACCTAC | [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molodtsova, T.N.; Moskalenko, V.N.; Lipukhin, E.V.; Antokhina, T.I.; Ananeva, M.S.; Simakova, U.V. Cerianthus lloydii (Ceriantharia: Anthozoa: Cnidaria): New Status and New Perspectives. Biology 2023, 12, 1167. https://doi.org/10.3390/biology12091167
Molodtsova TN, Moskalenko VN, Lipukhin EV, Antokhina TI, Ananeva MS, Simakova UV. Cerianthus lloydii (Ceriantharia: Anthozoa: Cnidaria): New Status and New Perspectives. Biology. 2023; 12(9):1167. https://doi.org/10.3390/biology12091167
Chicago/Turabian StyleMolodtsova, Tina N., Viktoria N. Moskalenko, Elizabeth V. Lipukhin, Tatiana I. Antokhina, Marina S. Ananeva, and Ulyana V. Simakova. 2023. "Cerianthus lloydii (Ceriantharia: Anthozoa: Cnidaria): New Status and New Perspectives" Biology 12, no. 9: 1167. https://doi.org/10.3390/biology12091167
APA StyleMolodtsova, T. N., Moskalenko, V. N., Lipukhin, E. V., Antokhina, T. I., Ananeva, M. S., & Simakova, U. V. (2023). Cerianthus lloydii (Ceriantharia: Anthozoa: Cnidaria): New Status and New Perspectives. Biology, 12(9), 1167. https://doi.org/10.3390/biology12091167