Simple Summary
Rhizobia are soil proteobacteria able to establish nitrogen-fixing symbiosis with host legumes. This symbiotic interaction, which is highly important from ecological and agronomical points of view since it allows growth of legumes in soils poor in nitrogen, requires a complex interchange of molecular signals between both symbionts. The production of rhizobial molecular signals is elicited by flavonoids, which are phenolic compounds exuded by legume roots. Recent work has shown that osmotic stress can also promote the formation of symbiotic signals in some rhizobia. In this work, we show that this is also the case for Sinorhizobium fredii HH103, a rhizobial strain able to establish symbiosis with hundreds of legumes, including the very important crop, soybean. Non-ionic osmotic stress, which can be encountered by the bacterium in the rhizosphere or inside the legume host, affected the expression of hundreds of bacterial genes and, consequently, influenced diverse bacterial traits, including the production of symbiotic signals and certain characteristics that may be relevant for successful interaction with the host: motility, production of the phytohormone indole acetic acid, and production of molecules involved in bacterial cell-to-cell communication. Thus, our work provides new evidence of how stress can promote rhizobia-legume symbiosis.
Abstract
(1) Background: Some rhizobia, such as Rhizobium tropici CIAT 899, activate nodulation genes when grown under osmotic stress. This work aims to determine whether this phenomenon also takes place in Sinorhizobium fredii HH103. (2) Methods: HH103 was grown with and without 400 mM mannitol. β-galactosidase assays, nodulation factor extraction, purification and identification by mass spectrometry, transcriptomics by RNA sequencing, motility assays, analysis of acyl-homoserine lactones, and indole acetic acid quantification were performed. (3) Results: Non-ionic osmotic stress induced the production of nodulation factors. Forty-two different factors were detected, compared to 14 found in the absence of mannitol. Transcriptomics indicated that hundreds of genes were either activated or repressed upon non-ionic osmotic stress. The presence of 400 mM mannitol induced the production of indole acetic acid and acyl homoserine lactones, abolished swimming, and promoted surface motility. (4) Conclusions: In this work, we show that non-ionic stress in S. fredii HH103, caused by growth in the presence of 400 mM mannitol, provokes notable changes not only in gene expression but also in various bacterial traits, including the production of nodulation factors and other symbiotic signals.
1. Introduction
Rhizobia-legume symbiosis involves interactions mediated by molecular dialogue between prokaryotic and eukaryotic cells. In this signal exchange, flavonoids secreted by the roots of leguminous plants are key molecules. These phenolic compounds are believed to interact with the bacterial protein NodD, which belongs to the LysR family of bacterial transcriptional regulators, triggering the transcription of so-called nodulation (nod) genes by interacting with conserved promoter sequences, called nod boxes [1,2,3]. Some nod genes are responsible for the biosynthesis and secretion of another fundamental signal for the establishment of the symbiosis, nodulation factors (Nod factors, NFs). NFs, also called lipochitooligosaccharides or LCOs, are N-acetyl-glucosamine (GlcNAc) oligomers (3–6 residues) exhibiting different chemical decorations [1,4]. NFs are recognised by plant receptors of the LYSM type, initiating the whole reaction cascade that culminates in the formation of nitrogen-fixing nodules in the legume roots [2,3].
Molecular recognition between NFs and plant receptors is a key factor not only for the establishment of symbiosis but also for the specificity of the interaction between rhizobia and legumes. However, other bacterial molecular signals also play important roles in the symbiotic process [1,5,6]. Different surface polysaccharides function either as signal molecules required for the progression of symbiosis or as protective molecules that face the plant defence system. The rhizobial surface polysaccharides showing more symbiotic relevance include exopolysaccharides (EPS), lipopolysaccharides (LPS), K-antigen capsular polysaccharides (KPS) and cyclic glucans (CG) [1,7]. In some rhizobia, such as Sinorhizobium fredii and different species of the Bradyrhizobium genus, flavonoids and NodD also induce the expression of TtsI, the transcription of which is driven by a nod box. TtsI is the transcriptional activator of genes (rhc, nop) related to the assembly of a symbiotic type III secretion system (T3SS), which is responsible for the delivery of effector proteins (nodulation outer proteins, Nops) into host cells [1]. These effector proteins, called T3Es, can have positive, negative, or neutral effects on symbiosis depending on the rhizobial-legume pair [8]. In addition, the expression of some other rhizobial genes may be driven by nod boxes and thus activated by Nod and flavonoids. This is the case for genes involved in the production of the phytohormone indole acetic acid (IAA) in both Sinorhizobium fredii HH103 and Rhizobium tropici CIAT 899 [9]. Finally, flavonoids also manipulate quorum-sensing systems, activating N-acyl homoserine lactone (AHL) synthesis genes in some rhizobia, such as S. fredii SMH12, Rhizobium etli ISP42, or Rhizobium sullae IS123 [10].
During their free-living stage in the soil, rhizobia are exposed to multiple physical stresses, such as high temperatures, acidity, or elevated osmolarity. Interestingly, in R. tropici CIAT 899, in addition to flavonoids, some other molecules or conditions can activate the biosynthesis of NFs. Thus, this strain, which nodulates the common bean, can produce NFs not only in the presence of flavonoids but also under osmotic stress, either ionic (300 mM NaCl) or non-ionic (400 mM mannitol) [9,11]. In R. tropici CIAT 899, in addition to nod genes, non-ionic osmotic stress also activates the expression of nitrogen fixation genes and represses genes involved in motility or polysaccharide production [12]. Different studies indicate that R. tropici CIAT 899 has two different circuits for nod gene activation and Nod factor biosynthesis. The first is the classical flavonoid-mediated activation involving NodD1, whereas the second relies on osmotic stress activation and requires both NodD2 and OnfD, an AraC-type transcriptional regulator [13,14,15,16].
Sinorhizobium fredii HH103 is a fast-growing rhizobial strain that nodulates dozens of legume genera, including Glycine max (soybean). This strain was isolated from a soil sample from the Honghu county of Central China and was first described in 1985 [17]. S. fredii strain HH103 can nodulate American and Asiatic soybean cultivars and is the best studied fast-growing soybean-nodulating strain to date [18]. Legumes nodulated by S. fredii HH103 can form determinate nodules, such as Glycine max and Vigna unguiculata, or indeterminate nodules, such as Glycyrrhiza uralensis and Sophora tomentosa. The most abundant Nod factors produced by HH103 consist of a mix of tri-, tetra-, and pentasaccharides carrying a saturated or unsaturated C16 or C18 fatty acyl residue in the non-reducing end of the molecule and a fucose or methylfucose modification at the C6 position of the reducing GlcNAc residue [19]. S. fredii HH103 symbiotic genes are regulated in a complex manner. In the presence of effective flavonoids such as genistein, NodD1 acts as the master activator of approximately 100 genes, including those related to Nod factor production and T3SS functioning [9,20,21]. However, other regulatory proteins participate in the modulation of symbiotic gene expression, such as the global repressor protein NolR, LysR family regulators NodD2 and SyrM, and the zinc-finger transcriptional regulator MucR1, all of which have different positive, neutral, or negative effects on expression depending on the gene [6,22,23,24,25].
The influence of additional S. fredii HH103 symbiotic signals during the nodulation process has also been investigated. The production of surface polysaccharides in S. fredii HH103 has been extensively described. In fact, the chemical structures of EPS, LPS, KPS, and CG are known, many of the genes involved in their production have been characterised, and the effect of the absence of any of these polysaccharides in symbiosis with soybean (and some other host legumes) has been studied [1,26,27]. A similar landscape emerges from reports dealing with T3SS and QS in this rhizobium, since both systems have been deeply described and the role of both Nops and AHL molecules during the symbiotic process has been analysed in detail [1,28,29,30,31,32,33,34].
In this work, we studied how non-ionic stress caused by the presence of a high concentration of mannitol affected S. fredii HH103 gene expression and whether this growth condition affected Nod factors production and different physiological traits, such as motility and production of surface polysaccharides IAA or AHLs.
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
S. fredii HH103 RifR and its nodD1::Ω (= SVQ318) [35] and nodD2::Ω (= SVQ515) [34] were grown at 28 °C on tryptone yeast (TY) medium [36]. For the β-galactosidase assay, AHL production, and RNA-seq analysis, yeast extract mannitol (YM) medium [37] was used. B− minimal medium [38] was used for NF-related experiments. Minimal medium (MM) [39] was used for the surface motility and IAA assays. All of these media, except for TY, contained 10 g L−1 mannitol (55 mM). Swimming motility was studied in Bromfield medium (BM) (0.04% tryptone, 0.01% yeast extract, and 0.01% CaCl2 2H2O) [40]. When required, media were supplemented with mannitol or the flavonoid genistein at a final concentration of 400 mM or 3.7 µM, respectively.
Agrobacterium tumefaciens NT1 (pZLR4), carrying a traG::lacZ fusion and traR for producing blue colour in the presence of X-Gal (5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside) and appropriate AHLs, was grown at 28 °C in YM supplemented with 30 µg mL−1 gentamicin [41].
The growth curves of S. fredii HH103 in different media were obtained using a Sinergy HT microplate reader (Agilent, Santa Clara, CA, USA) for 72 h at 28 °C with continuous orbital shaking. Measurements were undertaken every 4 h.
2.2. β-Galactosidase Assay
The β-galactosidase assay was carried out as previously described [20,42]. This assay was carried out with S. fredii HH103 (pMP240) grown in YM medium with or without 400 mM mannitol or 3.7 µM genistein. Plasmid pMP240 belongs to the IncP incompatibility group and harbours a TcR gene and transcriptional fusion occurs between the R. leguminosarum bv. viciae nodA promoter and a lacZ gene lacking its own promoter [43]. Bacterial cultures at OD660 = 0.8–1.0 were diluted (approximately 100-fold) before culturing in YM and YM supplemented with 400 mM mannitol to ensure that the cultures had an OD660 in the range of 0.15–0.30 when β-galactosidase activity was measured (approximately 16 h later).
2.3. Nod Factor Purification, Biological Activity, and Structural Determination by UHPLC-HRMS/MS
Nod factors (NFs) were purified from 1 L of culture for each sample following the previously described procedure [11]. The purified NFs were resuspended in 50 mL of acetonitrile 20% and 1 μL per mL of plant nutrient solution was added for the biological activity assays. G. max cv. Williams seeds were surface-sterilised and mounted in test tubes on a curled wire with the roots in 30 mL of Fåhraeus medium [44]. Roots were protected from light and plants were grown for 10 days. The growth chamber conditions were 16 h at 26 °C in light and 8 h at 18 °C in the dark, with 70% humidity. To determine the presence of nodule primordia, roots were cleared with sodium hypochlorite and stained with methylene blue using the previously described method [45]. Each experiment was repeated three times with six plants for each treatment.
To determine the chemical structure of the NFs, samples were analysed using an Ultra-High Performance Liquid Chromatography (UHPLC) system consisting of a quaternary UHPLC Dionex Ultimate 3000 SD (Thermo Fisher Scientific, Waltham, MA, USA) connected to a quadrupole-orbitrap Qexactive hybrid high-resolution mass spectrometer (HRMS) (Thermo Fisher Scientific, Waltham, MA, USA) with a HESI ionisation probe, as previously described [12]. The lists of NFs produced in each condition are provided in Supplementary Dataset S1.
2.4. RNA Extraction and Sequencing
S. fredii HH103 was grown in 50 mL Falcon tubes containing 7 mL of YM medium to ensure good aeration and supplemented with 400 mM mannitol when necessary. Bacteria were incubated with an orbital shaker (180 rpm) for 72 h at 28 °C, until reaching an OD600 of 0.8–1.0. Total RNA was isolated using a High Pure RNA Isolation Kit (Roche, Basel, Switzerland) according to the manufacturer’s protocol. Verification of the quantity and quality of the total RNA samples was carried out using a Nanodrop 1000 spectrophotometer (Thermo Scientific, Waltham, MA, USA) and a Qubit 2.0 Fluorometer (Invitrogen, Waltham, MA, USA). Two independent total RNA extractions were obtained for each condition with or without osmotic stress.
Ribosomal RNA was depleted using a MICROB Express Bacterial mRNA Purification kit (Ambion, Waltham, MA, USA) following the manufacturer’s protocol. The integrity and quality of the ribosomal RNA-depleted RNA were checked using an Agilent Bioanalyzer 2100 (Agilent Technologies, Santa Clara, CA, USA). RNA sequencing was carried out by Sistemas Genómicos ((Valencia, Spain) using the Illumina Next Generation Sequencing (NGS) platform with 100 bp paired-end reads using the Illumina HiSeq 2000 sequencing instrument (Illumina, San Diego, CA, USA). Ribosomal RNA-depleted samples were used to generate whole transcriptome libraries following the manufacturer’s recommendations for sequencing on this NGS platform. Amplified cDNA quality was analysed using the Bioanalyzer 2100 DNA 1000 kit (Agilent Technologies, Santa Clara, CA, USA) and quantified using a Qubit 2.0 Fluorometer (Invitrogen, USA). The RNA-Seq data discussed in this publication were deposited in the Sequence Read Archive of NCBI (BioProject database) under BioProject IDs PRJNA313151 [46] and PRJNA911807.
A total of four RNA-seq libraries corresponding to S. fredii HH103 under control and 400 mM mannitol conditions were generated (two independent biological experiments for each condition). Quality control of each run, sample normalisation, and statistical procedures were performed as previously described [9,46]. Differentially expressed genes for each strain and condition were obtained by comparing against gene expression levels of the wild-type strain grown under control conditions (Supplementary Datasets S2 and S3). The dataset was validated by qRT-PCR as previously described [9,46]. Briefly, the (DNA free) RNA extracted from 24-h old cultures was reverse transcribed to cDNA using the QuantiTec Reverse Transcription Kit (Qiagen, Hilden, Germany). Quantitative PCR was performed using a LightCycler 480 (Roche) with the following conditions: 95 °C, 10 min; 95 °C, 30 s; 50 °C, 30 s; 72 °C, 20 s; for 40 cycles, followed by melting from 60 °C to 95 °C to verify the specificity of the reaction. The 16S RNA gene was used as an internal control to normalize gene expression. The fold-change in gene expression was obtained using the ΔΔCt method [47]. Selected genes and primers are listed in Table 1.

Table 1.
Selected genes and primer sequences for qRT-PCR assays.
2.5. RNA-seq Data Analysis
First, quality control of the raw data was carried out by FASTQ. Next, the initial whole transcriptome paired-end reads obtained from sequencing were mapped against the latest version of the NCBI HH103 genome: NC_016812 (Chromosome), NC_16813 (plasmid a1), NC_16814 (plasmid c), NC_16815 (plasmid e), NC_16836 (plasmid b), NC_16846 (plasmid d, part 1), NC_16847 (plasmid d, part 2), NC_16848 (plasmid d, part 3), NC_16849 (plasmid d, part4), and NC_16850 (plasmid d, part 5), using the mapping algorithm Bowtie2 v2.3.1 [48]. Low-quality reads were eliminated using Samtools [49,50] and Picard Tools (http://broadinstitute.github.io/picard/), leaving only high-quality reads. The genetic quantification was calculated using the htseq count 0.6.1p1 method [51]. Gene differential expression and quantification were obtained using DESeq2 [52]. Concordance between samples under the same conditions was determined by correlation and Euclidean distance using the statistic software R. To determine differential expression among groups of samples, Phyton and R software and the DESeq2 algorithm were used. Differentially expressed genes (DEGs) in this study were established as those genes with a fold-change lower or higher than −3.0 or 3.0, respectively, with a p value adjusted to 0.05.
2.6. Quantification of Indole Acetic Acid (IAA) Production
Quantification of IAA from S. fredii HH103 was carried out using the Salkowski colourimetric assay, as described by del Cerro et al. [13]. Briefly, 5 mL of TY medium with tryptophan (0.4 g L−1) (supplemented when required with 400 mM mannitol) was inoculated with S. fredii HH103 and incubated for 96 h at 28 °C with an orbital shaker (180 rpm). Of these cultures, 1 mL of each sample was centrifuged to remove cells. The cell-free culture supernatant was assayed for IAA production. Every experiment was performed three times with eight replicates each time.
2.7. Motility Assay
Swimming was examined on plates prepared with BM containing 0.3% Bacto agar and surface motility was analysed on semisolid MM plates containing 0.4% agarose. When necessary, plates were supplemented with mannitol, ethanol, or the inducer flavonoid genistein, at final concentrations of 400 mM, 0.1% v/v, and 3.7 µM, respectively. Preparation of plates, inoculation with S. fredii HH103, and calculation of the migration zone were carried out as previously described [53].
2.8. PAGE Analysis of LPS and KPS
LPS extraction from bacterial cultures grown in solid TY medium in the presence or absence of 400 mM mannitol, separation on SDS-PAGE gels, and silver staining were performed as previously described [54]. For visualisation by PAGE, KPS was extracted from bacterial cultures grown in solid TY medium in the presence or absence of 400 mM mannitol, according to Hidalgo et al. [54]. Samples were analysed by PAGE as described by Parada et al. [55], but absolute ethanol was added to the running buffer and running and stacking gels at a final concentration of 10% (v/v) in all cases. The acrylamide concentration of the running gel was 18% (w/v) and the acrylamide:N,N-methylenebisacrylamide ratio was 29:1. Gels were fixed using Alcian Blue (0.5% in 2% acetic acid) and silver-stained. At least two independent cultures of each strain were used for the extraction of LPS and KPS.
2.9. AHL Well Diffusion Assay
A. tumefaciens NT1 (pZRL4) was used for the detection of AHLs (acyl chains ranging from C4 to C18) from supernatants of the parental strain grown to the late stationary phase in a well diffusion assay in Petri dishes, as described by McClean et al. and Cha et al. [56,57].
2.10. HPLC and Mass Spectrometry Analysis of AHL
Rhizobia were grown in 500 mL of YM medium for six days at 28 °C with shaking. Cultures were centrifuged and supernatants were extracted with dichloromethane and the organic layer was evaporated. The extracts were dissolved in 1 mL of methanol:water (1:1 v/v) containing 0.1% (v/v) formic acid, microfiltered (0.2 µm), and 20 mL was injected into an HPLC system equipped with a Tracer Hypersyl ODS column: 250 × 4.6 mm, 5 µm particle size (Teknokroma, Sant Cugat del Vallès, Spain). Methods using high pressure liquid chromatography and tandem mass spectrometry (HPLC-MS/MS) were carryout out as previously described [33]. Mass spectrometric conditions were optimised by infusing solutions of standards dissolved in methanol (100 mg mL−1) at a flow between 10–100 mL min−1: C4-HSL, C6-HSL, 3-oxo-C6-HSL, C8-HSL, 3-oxo-C8-HSL, C10-HSL, 3-oxo-C10-HSL, C12-HSL, 3-oxo-C12-HSL, 3-OH-C12-HSL, C14-HSL, 3-oxo-C14-HSL, and 3-OH-C14-HSL. The probe capillary voltage was optimised at 5500 V. The desolvation temperature was set to 50 °C. The pressures of curtain, nebulising, and turbo spray gases were set to 35, 20, and 0 (arbitrary units), respectively. Nitrogen was used for CID (collision-induced dissociation). Ions were scanned from m/z 150 to 500 at a scan rate of 4000 Th s−1.
3. Results
3.1. Expression of S. fredii HH103 Nod Genes Is Induced by the Presence of 400 mM Mannitol
In previous works, we showed that non-ionic osmotic stress, 400 mM mannitol, induces nod gene expression and Nod factor production in R. tropici CIAT 899 [12]. In the present work, we wanted to investigate whether this also occurs in another rhizobial strain, S. fredii HH103. For this purpose, we analysed lacZ expression driven by the promoter of the nodA gene of Rhizobium leguminosarum bv. viciae in S. fredii HH103 harbouring the pMP240 plasmid, when grown in the presence of different mannitol concentrations: from 55 mM (the concentration of mannitol present in YM medium) to 1 M. The expression of pnodA::lacZΔp (measured as β-galactosidase activity) was significantly higher than that found in YM medium in the range of 100 to 800 mM mannitol, reaching a maximum of 2-fold at 300–500 mM mannitol (Supplementary Figure S1). These results indicated that a high concentration of mannitol positively influenced nod gene expression, not only in R. tropici CIAT 899 [12] but also in S. fredii HH103. However, this effect was more limited than the presence of nod gene-inducing flavonoids such as genistein, which caused a 12-fold increase in pnodA::lacZΔp expression (Figure 1).
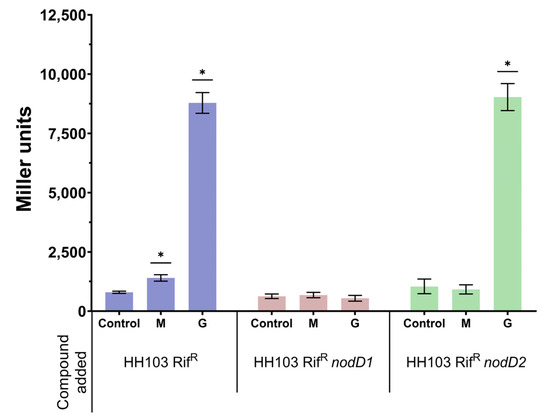
Figure 1.
β-galactosidase activity of S. fredii HH103 and its nodD1 and nodD2 mutant derivatives carrying plasmid pMP240 upon growth in YM (Control) or YM supplemented with 400 mM mannitol (M) or 3.7 µM genistein (G). Asterisks (*) indicate a significant difference from the corresponding control sample using the non-parametric test of Mann–Whitney, α = 5%.
Since S. fredii HH103 carries two different copies of nodD (nodD1 and nodD2), we investigated if either gene mediated the observed increase in nod gene expression in the presence of 400 mM mannitol. For this purpose, we studied the expression of pnodA::lacZΔp in the presence of 400 mM mannitol or 3.7 µM genistein in the wild-type and in nodD1 and nodD2 S. fredii HH103 mutant backgrounds (Figure 1). As expected, genistein enhanced β-galactosidase activity in S. fredii HH103 nodD2 but not in S. fredii HH103 nodD1 since NodD1 is responsible for the flavonoid-induced expression of nod genes in this bacterium [20]. Interestingly, the presence of 400 mM mannitol did not enhance pnodA::lacZΔp expression in either the nodD1 or the nodD2 S. fredii HH103 mutants. This result suggested that both NodD1 and NodD2 may play a role in the induction of nod gene expression caused by the presence of non-ionic osmotic stress.
3.2. S. fredii HH103 Produces Biologically Active Nod Factors in the Presence of 400 mM Mannitol
As mentioned above, the enhanced expression of R. tropici CIAT 899 nod genes provoked by the presence of 400 mM mannitol resulted in an increase in the production of NFs [12]. In addition, these NFs were shown to be biologically active since they promoted the formation of nodule primordia in the roots of the host legume Phaseolus vulgaris.
The fact that the presence of 400 mM mannitol also increased nod gene expression in S. fredii HH103 prompted us to investigate whether Nod factor production was also affected by this growth condition. For this purpose, we carried out ultra-high performance liquid chromatography (UHPLC) followed by data-dependent high-resolution mass spectrometry (UHPLC–HRMS/MS) of S. fredii HH103 cultures grown in YM medium in the absence or presence of non-ionic osmotic stress (400 mM mannitol). Cultures of S. fredii HH103 grown in YM medium supplemented with 3.7 µM genistein were also included in this study as a control for NF production. As shown in Figure 2A, when S. fredii HH103 was grown in YM medium, 14 different NFs were detected, whereas the presence of genistein or high mannitol concentrations led to an increase in the number of Nod factors produced (124 and 42, respectively).
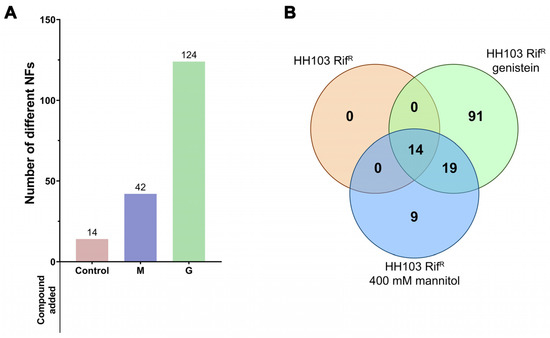
Figure 2.
(A) Number of NFs produced by S. fredii HH103 grown in B− medium in the absence (Control) or presence of mannitol (M) or genistein (G). (B) Venn diagram of the sets of Nod factors produced by S. fredii HH103 grown in B− medium in the absence (orange circle) or presence of mannitol (blue circle) or genistein (green circle).
The complete list of Nod factors produced under each condition is provided in Supplementary Dataset S1. All the NFs produced in YM (14 NFs) were also found among those produced in the presence of either genistein or high mannitol concentrations (Figure 2B). In addition, 19 other NFs were also found in both inducing conditions, whereas 9 and 91 different NFs were exclusively found in the presence of either 400 mM mannitol or 3.7 µM genistein, respectively. Thus, a core of 33 different NFs was produced under both nod gene-inducing conditions. Among them, NFs having 3 to 5 GlcNAc residues and harbouring predominantly saturated or monounsaturated C16 or C18 acyl groups as well as fucose or methyl fucose decorations could be found (Supplementary Dataset S1).
The putative biological activity of the NFs produced by S. fredii HH103 upon growth in the presence of 400 mM mannitol was studied by adding NF extracts to germinated soybean seedlings. Thirteen days after inoculation, the roots were stained with methylene blue and examined to detect the possible presence of nodule primordia (Figure 3). In the case of non-induced S. fredii HH103 cultures (B− medium, 55 mM mannitol), the purified NF extract slightly induced the formation of primordia (2.8 ± 0.9 primordia/root), a phenomenon that was previously described for R. tropici CIAT 899 [12,15] and might be because a small number of NFs could be detected in such extracts. However, when the purified extracts of S. fredii HH103 cultures grown in 400 mM mannitol were added to soybean seedlings, a significant increase (approximately 3-fold) in the formation of nodule primordia could be detected (7.6 ± 1.5 primordia/root). In fact, the number of nodule primordia induced under the latter condition was very similar to that obtained when NFs purified from genistein-induced S. fredii HH103 cultures were added (7.9 ± 1.0 primordia/root) to soybean roots. These results clearly indicated that the NFs produced upon growth in B− supplemented with a final concentration of 400 mM mannitol were, like those produced upon induction with genistein, biologically active.
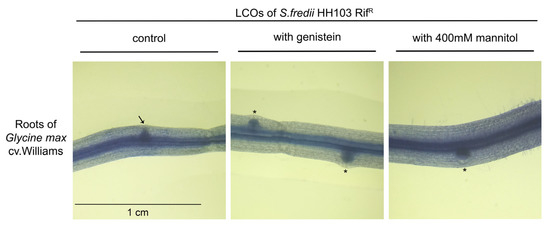
Figure 3.
Biological activity assay of the NFs purified from S. fredii HH103 cultures grown in B− medium in the absence (control) or presence of 3.7 µM genistein or 400 mM mannitol applied to soybean roots. Nodule primordia (circular shape) are denoted by asterisks, whereas an arrow marks an emerging lateral root (spiky shape).
3.3. A High Concentration of Mannitol Affects the Expression of Hundreds of S. fredii HH103 Genes
In a previous work, it was determined that non-ionic stress caused by the presence of 400 mM mannitol altered the expression of a high number of genes and consequently affected different biological processes in R. tropici CIAT 899 [12]. To check whether this also takes place in S. fredii HH103, we performed RNA-seq analysis of S. fredii HH103 cultures grown in YM medium (55 mM mannitol) and YM supplemented with a final concentration of 400 mM mannitol. The number of differentially expressed genes (DEGs) with fold-changes ≥3 with mannitol compared to the control was 743, including 350 induced DEGs and 393 repressed DEGs (Supplementary Datasets S2 and S3). Since the genome of S. fredii HH103 is estimated to have 6960 genes [58], the presence of mannitol affected the expression of more than the 10% genome of this strain. Remarkably, the percentage of induced or repressed genes varied depending on the replicon (Table 2). For example, regarding the symbiotic plasmid (pSfHH103d), 77 genes were induced whereas only 6 were repressed. On the other hand, the number of repressed genes (292) was considerably higher than that of induced genes (119) in the chromosome. Regarding the largest plasmid (pSfHH103e), the number of induced genes was double the number of repressed genes (148 vs. 72). Among the different small plasmids present in S. fredii HH103, it is noteworthy that more than 20% of the genes present in plasmid pSfHH103a2 (8) were repressed whereas none of them were induced in the presence of 400 mM mannitol. Supplementary Datasets S2 and S3 contain detailed lists of all the S. fredii HH103 DEGs sorted by their fold-change or genome location, respectively. In both cases, the annotation of each DEG is provided.
Table 2.
Distribution of S. fredii HH103 DEGs in the presence of 400 mM mannitol among the different replicons in the genome.
Transcription of 11 of the found DEGs was quantified by qRT-PCR to validate the dataset (Table 3). In all cases, except for nodA, the same type of change of expression (either induction or repression) upon growth with 400 mM mannitol was detected.
Table 3.
RNA-seq data validation using qRT-PCR. The expression of 11 S. fredii genes was measured both in the presence or absence of 400 mM mannitol. For qRT-PCR, fold-change values were calculated using the ΔΔCt method and normalised to the reference 16S rRNA gene.
3.4. The Presence of a High Concentration of Mannitol Induces the Expression of 350 S. fredii HH103 Genes
Among the 350 induced genes in the presence of 400 mM mannitol, 91 genes (26.0%) corresponded to hypothetical proteins of unknown function. 38 genes were involved in ABC-type transport and another seven genes were involved in other types of transport of different metabolites. According to their predicted functions, these transport systems would be responsible for the uptake of sugars, such as glucose or fructose, dicarboxylic acids, nitrate, sulphate, glycine betaine, proline, or spermidine (Supplementary Datasets S2 and S3). Similarly, three genes coding for products with BON domains, which are found in proteins that protect cells from osmotic stress, were overexpressed in 400 mM mannitol. In addition, two genes coding for universal stress proteins, psfHH103d_567 (+4.2) and psfHH103d_586 (+4.1), one encoding a DnaJ-like protein (psfHH103d_507, +3.6), and two genes coding for GroEL chaperones, SFHH103_00877 (+4.1) and psfHH103d_587 (+3.1), were also induced.
Regarding genes related to symbiosis, eight nif or fix genes involved in the biological fixation of atmospheric nitrogen were induced, including nifS, nifN, fixP, fixB or fixJ (Supplementary Dataset S2). Other nif and fix important genes showed levels of induction lower than 3-fold, such as nifHDK, which codes for the different polypeptides constituting the nitrogenase enzyme, the levels of expression of which increased approximately 2-fold. In S. fredii HH103, two clusters of nod genes account for the production of NFs: the nodABCIJnolO’noeI (psfHH103d_126 to psfHH103d_132) operon and the nolK-noeL-nodZ-noeK and noeJ genes (psfHH103d_379 to psfHH103d_383) [35,58,59]. Surprisingly, none of these nod genes were scored as DEGs in the presence of 400 mM mannitol, although some of them displayed transcriptional activation higher than 2-fold (Supplementary Dataset S2). Instead, two copies of the nodU gene exhibited increased expression: SFHH103_05865 (+4.1), located on pSfHH103e, and the pseudogene 454 (+3.9), harboured by the symbiotic plasmid pSfHH103d. Different genes involved in the formation of the T3SS apparatus (rhcJ, nopX, and rhcN, among others), as well as genes coding for effectors secreted by this apparatus such as nodD and gunA, were also induced by the presence of 400 mM mannitol. In addition, different genes involved in the production of surface polysaccharides, such as rkpI and rkpJ (K-antigen capsular polysaccharide or KPS) [18,54] and exoI, exoN, or exoF genes (exopolysaccharide, EPS) [60], were induced under this condition. Finally, other important symbiotic genes, such as those responsible for the biosynthesis of indole acetic acid (IAA), psfHH103d_257 to psfHH103d_259, which are located on the pSym and are under the transcriptional control of a nod box [58], were strongly upregulated upon the high concentration of mannitol (Supplementary Dataset S2).
Other genes showing increased expression in the presence of 400 mM mannitol included: (i) sinI, responsible for the synthesis of long-chain acyl-homoserine lactones (AHLs) [33]; (ii) different genes involved in the synthesis of cytochromes and the enzymes NADH dehydrogenase and nitrate reductase; (iii) 12 genes coding for different types of transcriptional regulators, including members of the HTH, GntR, and XRE families and the negative regulator TraM.
3.5. The Presence of a High Concentration of Mannitol Represses the Expression of 393 S. fredii HH103 Genes
Among the 393 genes differentially repressed in the presence of 400 mM mannitol, 97 of them (24.7%) coded for hypothetical proteins of unknown function Similar to the induced genes, a high number of genes (18) involved in ABC-type transport processes (iron, zinc, cyclic glucans, methionine, phosphonate, sugars) were repressed. Other processes that could be affected by the repressed genes included cell division and chromosome or plasmid segregation (eight genes), bacterial cell wall biosynthesis (ten), or chemotaxis (eight).
Concerning genes related to stress responses, four different copies of the tandem genes coding for GroES/GroEL chaperones were strongly repressed (between −8.3 and −20.1-fold), in contrast to the two previously mentioned copies of GroEL, one located on the pSym and one on the chromosome, that were induced 3.1–4.0-fold. Two of the repressed copies were located on the chromosome (SFHH103_00476-SFHH103_00475, SFHH103_03405-SFHH103_03404), one on the plasmid c (SFHH103_04126-SFHH103_04125), and the other on plasmid a2 (psfHH103a2_17-psfHH103a2_16). Additionally, the chromosomal copy of dnaJ (SFHH103_00574) was strongly repressed (9.4-fold), in contrast to the copy harboured by the pSym, which was induced 3.6-fold. Five multidrug efflux pump genes, which are related to the exclusion of toxic substances from the bacterial cell, were also repressed.
Regarding genes involved in surface appendages, two genes required for flagellum biosynthesis, fliP and flaC, as well as several genes involved in type IV pili biosynthesis, including pilQ or cpaA, were repressed in the presence of 400 mM mannitol. Although four genes responsible for EPS biosynthesis (exoN, exoO, exoI, and exoF3) were induced in the presence of mannitol, exoN, exoB, a possible EPS transporter, and exoR, which codes for a negative regulator of EPS biosynthesis, were repressed. Another repressed gene was grlR, the product of which is predicted to act as a negative regulator of T3SS, a system that, as mentioned before, is activated in the presence of non-ionic osmotic stress. In contrast, four out of seven genes (rhcV-2, rhcU-2, rhcT-2, and rhcC2-2) involved in the production of a second T3SS apparatus (T3SS-2) were strongly repressed at the high concentration of mannitol (Supplementary Dataset S2).
In addition, a high number of genes coding for transcriptional regulators (at least 18) were repressed by the presence of 400 mM mannitol, including members of the LuxR, LysR, LacI, and GntR families, among others.
3.6. The Presence of a High Concentration of Mannitol Affects different Physiological Traits in S. fredii HH103 Genes
The transcriptomics analysis of the effect of non-ionic stress on S. fredii HH103 mentioned above showed differential expression of genes involved in different biological processes beyond NF production, T3SS formation, or nitrogen fixation, including those related to motility, surface polysaccharide biosynthesis, and production of IAA and AHLs (Supplementary Dataset S2). Therefore, we decided to investigate whether these processes were affected by the presence of 400 mM mannitol. Before studying these bacterial traits, we examined whether the growth of S. fredii HH103 might be altered by the presence of 400 mM mannitol in the different culture media that would be employed in those analyses (see Material and Methods). As shown in Supplementary Figure S2, this non-ionic osmotic stress negatively affected S. fredii HH103 growth in TY and YM media and MM, but had a positive effect in BM.
3.6.1. Motility
In a recent study, we demonstrated that S. fredii HH103 exhibited surface motility in the presence of genistein in a NodD1- and TtsI (master activator of the T3SS)-dependent manner. However, the presence of this nod gene-inducing flavonoid did not influence the swimming capacity of this strain [53]. Since the expression of some S. fredii HH103 flagellar genes was repressed by the presence of 400 mM mannitol, we decided to investigate whether this stress condition might affect S. fredii HH103 motility. For this purpose, we carried out swimming and surface motility assays in the presence or absence of 400 mM mannitol. As shown in Figure 4, the swimming ability of S. fredii HH103 was fully abolished in the presence of a high concentration of mannitol. In contrast, the presence of 400 mM mannitol promoted S. fredii HH103 surface motility, although to a lower extent than genistein (Figure 5).
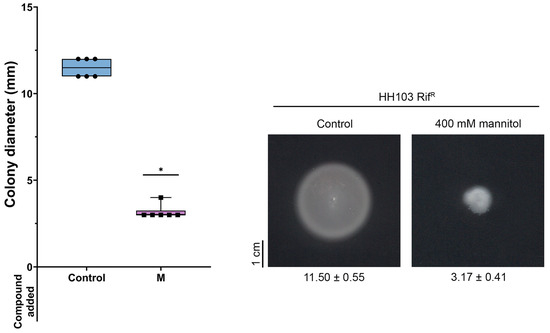
Figure 4.
Swimming ability of S. fredii HH103 in Bromfield medium in the absence (Control) or presence of 400 mM mannitol (M). All data points are shown in box and whisker plots from at least three biological replicates performed with two technical replicates. Values under images represent the average and standard deviation of migration (given in millimetres and determined as described in the text). Asterisks (*) indicate a significant difference from the corresponding control sample using the non-parametric test of Mann–Whitney, α = 1%.
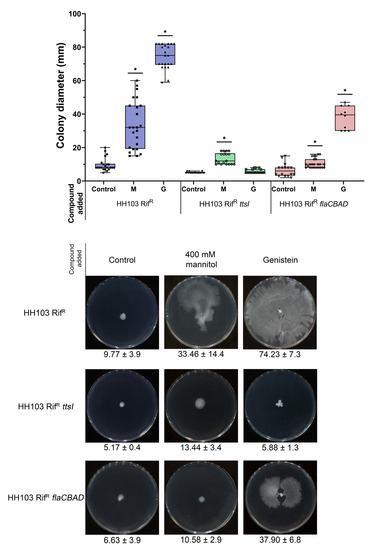
Figure 5.
Surface motility of S. fredii HH103 and its ttsI and flaCBAD mutant derivatives in MM in the absence (Control) or presence of 400 mM mannitol (M) or 3.7 µM genistein (G). All data points are shown in box and whisker plots from at least three biological replicates performed with two technical replicates. Values under images represent the average and standard deviation of migration (given in millimetres and determined as described in the text). For each strain, asterisks (*) indicate a significant difference from the corresponding control sample using the non-parametric test of Mann–Whitney, α = 1%.
As described earlier, genistein-induced surface motility of S. fredii HH103 is dependent on the presence of a functional T3SS [53]. To check whether this was also the case for surface motility induced by a high concentration of mannitol, this trait was studied in an HH103 ttsI::Ω, a mutant derivative of S. fredii HH103 that lacked the positive regulator of this system, TtsI [34]. The growth of this mutant slightly extended from the centre of the plate (Figure 5). However, this could have been a consequence of high EPS production due to the high amount of carbon source available rather than surface motility, which suggested that T3SS was also essential for surface motility when induced by non-ionic osmotic stress in S. fredii HH103.
Genistein-induced surface motility of S. fredii HH103 is mediated by a combination of flagellum-dependent and -independent mechanisms [53]. Thus, we also investigated whether the presence of flagella was required for S. fredii HH103 surface motility upon growth in the presence of 400 mM mannitol. The growth of HH103 ΔflaCBAD, a derivative of S. fredii HH103 that lacked flagella [26], slightly extended from the centre of the plate, which, again, could have been the result of increased EPS production rather than surface motility (Figure 5). This result suggested that the surface motility exhibited by HH103 upon non-ionic osmotic stress was mediated by a flagellum-dependent mechanism.
3.6.2. Surface Polysaccharides Biosynthesis
The fact that the expression of various genes involved in surface polysaccharide biosynthesis was affected by non-ionic osmotic stress led us to investigate whether the production of some of those polysaccharides might be altered by the growth condition. Regarding EPS, as commented above, several exo genes were affected, some of which were induced, and others were repressed. When grown in YM medium, no obvious changes in the appearance of mucoid colonies could be scored (data not shown), but a clear increase in mucoidy was observed in the presence of 400 mM mannitol when grown in TY medium (Figure 6).

Figure 6.
Mucoidy of S. fredii HH103 grown in TY medium in the absence or presence of 400 mM mannitol.
On the other hand, two genes involved in KPS production (rkpIJ) were induced and several genes putatively involved in LPS synthesis (cpsA, SFHH103_02601, SFHH103_05300, SFHH103_05856) were either induced or repressed when S. fredii HH103 was grown in the presence of 400 mM mannitol (Supplementary Dataset S2). However, the electrophoretic profiles of these two polysaccharides were apparently not affected by non-ionic osmotic stress (Supplementary Figure S3).
3.6.3. Production of IAA
In S. fredii HH103, the psfHH103d_257-psfHH103d_259 genes, which are under the transcriptional control of a nod box, are involved in IAA synthesis [46,58]. Since these genes were also induced by non-ionic osmotic stress, we analysed the production of IAA by S. fredii HH103 when grown in the presence or absence of a high concentration of mannitol. As expected, the presence of 400 mM mannitol significantly enhanced (2-fold) the production of IAA (45.9 ± 5.5 mg/L) compared to that under control conditions (22.9 ± 1.4 mg/L) (non-parametric test of Mann–Whitney, α = 1%).
3.6.4. Production of AHLs
S. fredii HH103 presents two known genes (traI and sinI) coding for AHL synthases. TraI and SinI are responsible for the production of short-(C4 to C8) and long-(C10 to C16) acyl chain AHLs, respectively, but at very low, non-physiological concentrations [33,58]. According to the RNA-seq analysis, the sinI gene was highly upregulated in the presence of 400 mM mannitol (Supplementary Dataset S2). Thus, S. fredii HH103 supernatants from cultures grown with or without a high concentration of mannitol were inspected for AHL production in well diffusion assays using the biosensor A. tumefaciens NT1 (pZRL4). Interestingly, the presence of AHLs could only be detected under the non-ionic osmotic stress condition (Figure 7A).
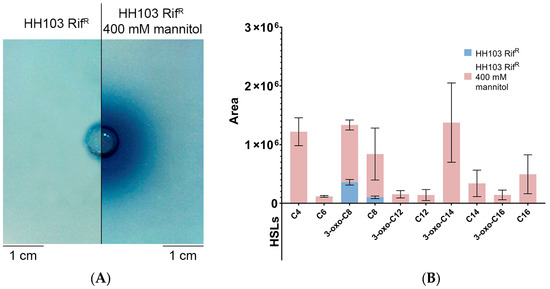
Figure 7.
(A) Well diffusion assay, using the biosensor A. tumefaciens NT1 (pZRL4), for detection of AHLs produced by S. fredii HH103 grown in TY medium in the absence (left) or presence (right) of 400 mM mannitol. (B) Specific AHLs produced by S. fredii HH103 grown in TY medium in the absence (blue) or presence (pink) of 400 mM mannitol, as determined by HPLC-MS/MS.
In a previous work, qualitative mass spectrometry analysis using HPLC-MS/MS unequivocally identified C8-HSL and 3-oxo-C8-HSL, produced by TraI, and C12-HSL, C14-HSL, and 3-oxo-C14-HSL, synthesised by SinI, in supernatants of S. fredii HH103 cultures, despite long-chain AHLs being present as trace compounds [33]. Here, we performed quantitative mass spectrometry analysis, establishing correlations between signal areas of any particular AHL and its relative abundance in supernatants from cultures with or without high concentration of mannitol (Supplementary Dataset S4). Figure 7B shows the areas of AHL signals present in supernatants from S. fredii HH103 cultures grown in the presence or absence of 400 mM mannitol (only AHLs detected in four biological replicates with average area values ≥ 105 are represented). Following this approach, quantitative variations for individual AHLs were estimated. We found that short-chain AHLs (3-oxo-C8-HSL and C8-HSL) were present at high concentrations in both control and mannitol supernatants. Conversely, the set of long-chain AHLs (3-oxo-C12-HSL, C12-HSL, 3-oxo-C14-HSL, C14-HSL, 3-oxo-C16-HSL, and C16-HSL) was only detected when S. fredii HH103 was grown with a high concentration of mannitol. Altogether, these results suggested that non-ionic osmotic stress conditions induced the production of long-chain AHLs by sinI transcriptional activation in S. fredii HH103.
4. Discussion
Rhizobia are soil bacteria that can shift between two different styles of life: free-living saprophytic bacteria in the soil and endosymbionts hosted inside specialised organs, called nodules, formed by the roots of legumes in response to the presence of compatible rhizobia [61]. Because of this, rhizobia have to survive in different environments and cope with different kind of stresses, including high temperatures, acidity, or basicity, as well as elevated osmolarity due to high concentrations of different solutes either in the soil or inside the host plant [62].
Non-ionic osmotic stress created by the presence of 400 mM mannitol had a substantial impact on both the bacterial growth and gene expression of S. fredii HH103. On the one hand, as expected, this condition negatively affected S. fredii HH103 growth in TY and YM media and MM, but, interestingly, it had a positive effect in BM, most likely because of the low osmolarity of this medium (it is basically a 10-fold diluted version of TY). On the other hand, growth upon 400 mM mannitol affected the expression of more than 10% of the annotated genes of S. fredii HH103, and the numbers of induced and repressed genes were similar (350 and 393 respectively). Remarkably, the percentages of induced and repressed genes varied greatly depending on the replicon (Table 2). In the case of the symbiotic plasmid, these percentages were 11.5% and 0.9% respectively, in contrast to the 0.0% and 21.1% found for plasmid pSfHH103a2. In the case of pSfHH103e, another plasmid important for symbiosis [46,58], these values were 7.4% and 3.6%, respectively. Thus, interestingly, the two plasmids more relevant for symbiosis were likely the replicons showing the highest percentages of induced genes upon non-ionic osmotic stress. The expression of a high number of genes was affected by this condition; thus, it is not possible to comment on all of them. However, it is worth considering the presence of numerous genes involved in ABC transport systems, some induced, others repressed, which indicates that the presence of 400 mM mannitol might alter the transport of solutes across the bacterial envelope. Another group of genes for which transcription was also clearly altered was that coding for chaperonins, especially GroEL which is required for the appropriate folding of hundreds of proteins, approximately 10% to 15% of the cellular proteins [63,64]. Rhizobial species harbour several copies of groEL (i.e., there are 5 copies in S. meliloti and 3 in Mesorhizobium huakuii) [65,66]. Interestingly, all four of the groEL copies present in S. fredii HH103 that were co-expressed with the coding gene of the co-chaperone GroES were repressed by non-ionic osmotic stress, whereas two additional copies of groEL that were not associated with groES were induced in that condition. This result suggested that some chaperonins might be more relevant, and thus differentially expressed, for adaptation to non-ionic stress conditions. Clearly, more research is necessary to shed light on this issue.
Recent studies have demonstrated that different osmotic stresses, either ionic (NaCl) or non-ionic (mannitol, dulcitol), activate the expression of R. tropici CIAT 899 symbiotic genes, including those involved in NFs (nod genes) and those related to nitrogen fixation (nif and fix genes) [9,12]. In the case of osmotic stress caused by a high concentration of mannitol, we observed strong repression of several exo and kps genes, which are involved in the synthesis of exopolysaccharide (EPS) and capsular polysaccharide (KPS) respectively, and induction of genes related to the production of cyclin glucans (CG) in R. tropici CIAT 899 [12]. On the other hand, in this bacterium, ionic osmotic stress provoked by high concentrations of NaCl also induced the expression of genes related to the production of IAA [9].
In this work. we showed that non-ionic osmotic stress caused by the presence of a high concentration of mannitol had an effect on different physiological traits of S. fredii HH103, including the production of different symbiotic molecular signals. Upon this growth condition, this rhizobial strain exhibited enhanced expression of nod genes (as scored using the reporter plasmid pMP240 in the β-galactosidase assay and by qPCR) and produced significant amounts of biologically active NFs, although showing a lower diversity that those measured upon induction with flavonoids. However, our transcriptomics results did not detect induced expression of the main nod genes (nodABC) upon non-ionic osmotic stress. Most likely, this may be due to the fact that the RNA-seq experiments were carried out using S. fredii HH103 cultures grown in the presence of mannitol for 72 h, whereas both the β-galactosidase assay and qPCR experiments were performed using 16–24-h-old cultures. In fact, preliminary analyses of new RNA-seq experiments following 16 h incubation in the presence of 400 mM mannitol has revealed a slight induction (2–2.5-fold) of S. fredii HH103 nodABC genes (data not shown). Taking into account all of these data, we hypothesize that the presence of 400 mM mannitol might cause a transient induction of S. fredii nodABC genes. In any case, the data provided in this work showed that non-ionic osmotic stress positively influenced the production of NFs by S. fredii HH103.
We also observed that non-ionic osmotic stress caused by growth in the presence of a high concentration of mannitol may affect the production of other S. fredii HH103 symbiotic signals, since this condition influenced the expression of different genes related to symbiotic T3SS and the production of different surface polysaccharides (EPS, LPS, KPS), as determined by RNA-seq. On the one hand, several genes involved in T3SS were induced. Since the expression of the ttsI gene, which codes for the transcriptional activator of this secretion system [35], was very slightly affected (+1.5-fold), one possibility is that the enhanced expression of some T3SS-related genes might be due to the observed repression of grlR (−4.9-fold), the product of which is predicted to act as a negative regulator of T3SS. Unfortunately, we could not study whether the secretion of T3Es was enhanced since the presence of 400 mM mannitol negatively affected the extraction of extracellular proteins. Regarding KPS and LPS, the rkpIJ genes involved in KPS export [54,67] were strongly induced (+35.2- and 5.6-fold, respectively), whereas several genes putatively involved in LPS production were either induced or repressed. Although the electrophoretic profiles of these polysaccharides were not altered upon growth in the presence of 400 mM mannitol, this methodology was not quantitative and thus the possibility of increased/decreased production of LPS and KPS cannot be discounted. Concerning EPS, we observed the altered expression of several genes, some of them induced, others repressed. Interestingly, exoR, of which the S. meliloti orthologue has been identified as a repressor of EPS production [68,69], was a repressed gene. This fact might be the cause of the observed enhanced production of EPS by S. fredii HH103 when grown in TY medium supplemented with 400 mM mannitol. Furthermore, non-ionic osmotic stress and the presence of flavonoids have opposing effects on EPS production since the production of EPS in S. fredii HH103 is repressed by these phenolic compounds in a NodD1- and SyrM-dependent mechanism [70,71]. Finally, it is worth mentioning that the ndvA gene, which codes for the protein responsible for CG export from the cytoplasm to the periplasmic space [72,73], was repressed (−5.6-fold) upon growth in the presence of 400 mM mannitol. This fact suggests that S. fredii HH103 might accumulate intracellular CGs in response to that stress, but elucidating this possibility would require further investigation.
There are additional S. fredii HH103 traits that were influenced by non-ionic stress and might be relevant for symbiosis. Among them is the above-mentioned altered pattern of expression of different S. fredii HH103 groEL copies. In fact, studies carried out in S. meliloti and M. huakuii showed that specific copies of groEL were essential for symbiosis with their host plants [65,66]. On the other hand, the presence of a high concentration of mannitol clearly affected S. fredii HH103 motility, abolishing swimming (most probably because of the increase in viscosity in the culture medium), but promoting surface propagation by a flagellum- and T3SS-dependent mechanism. Interestingly, recent work has shown that the presence of nod gene-inducing flavonoids also enhanced S. fredii HH103 surface motility, although it did not affect swimming capacity [53]. Flavonoid-induced surface motility also requires the presence of a functional T3SS, but is mediated by both flagellum-dependent and -independent mechanisms. To elucidate whether both surface motility-inducing conditions (presence of nod-gene-inducing flavonoids or a high mannitol concentration) share the same flagellum-dependent mechanism would require further investigation. Although rhizobial motility appears not to be essential for symbiosis, it might enhance nodulation efficiency and competitiveness [74,75]. Another interesting trait induced in S. fredii HH103 by the presence of a high mannitol concentration was the production of IAA, a phytohormone that has been described as an enhancer of nodule development and symbiotic performance of rhizobia [76]. In fact, many plant growth-promoting rhizobacteria enhance legume–rhizobial symbioses through the production of IAA. Curiously, genistein also enhances the production of IAA in S. fredii HH103 through the induction of the same genes (under the transcriptional control of a nod box) as mannitol [46,58]. In R. tropici CIAT 899, the expression of genes related to the production of IAA is also driven by a nod box and induced by flavonoids and ionic osmotic stress, but not in the presence of a high concentration of mannitol [9,12]. In any case, there is the possibility that at least some rhizobia have evolved the ability to produce IAA as a way of improving symbiotic interactions with their host plants. Finally, and very interestingly, the non-ionic stress caused by the presence of 400 mM mannitol provoked a clear increase in the amount and diversity of AHLs produced by S. fredii HH103. Most probably, this enhancement was due to the induced expression of the AHL-synthase coding gene sinI. In fact, we show in this work that this stress condition acted as a potent elicitor of the production of long chain AHLs in S. fredii HH103, which, otherwise, are produced at very low concentrations (non-physiological) when this bacterium is grown under normal culture conditions [33]. Quorum-sensing systems regulate cooperative behaviours in bacteria, and, in the case of rhizobia, many studies show its relevance for symbiosis [77]. In any case, further studies are required to elucidate the relevance of this increased AHL production in the different phenotypes of S. fredii HH103 upon growth in the presence of 400 mM mannitol.
In order to distinguish between osmotic and putative effects on growth upon a too high concentration of a readily metabolisable substrate, we plan to extend our studies of non-ionic osmotic stress conditions caused by the presence of non-metabolisable compounds. In any case, similar results have been previously observed with other rhizobia, such as R. tropici CIAT 899, by using metabolisable (mannitol) or non-metabolisable (dulcitol) substrates [12].
5. Conclusions
In summary, this work demonstrates that non-ionic stress caused by growth in the presence of a high concentration of mannitol provoked notable changes in S. fredii HH103, not only in gene expression but also in various bacterial traits, including the production of NFs, other symbiotic signals, as well as other characteristics that might be relevant for symbiosis with legumes. Our work provides new evidence that, as it has been recently reviewed [62], rhizobial responses to stress might play key roles in symbiotic signalling.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/biology12020148/s1, Figure S1. β-galactosidase activity of S. fredii HH103 carrying plasmid pMP240 upon growth in YM supplemented with different mannitol concentration (from 55 mM to 1 M). Figure S2. Growth curves of S. fredii HH103 in different media supplemented with or without 400 mM mannitol. Figure S3. LPS (A) and KPS (B) electrophoretic profiles of S. fredii HH103 grown in TY medium in the absence or presence of 400 mM mannitol. Supplementary Dataset S1. Complete list of Nod factors produced by Sinorhizobium fredii HH103 in YM medium in the absence (HH−) or presence of 3.7 µM genistein (HH+) or 400 mM mannitol (HH mannitol). Supplementary Dataset S2. S. fredii HH103 DEGs upon growth in 400 mM mannitol ordered by fold-change. Supplementary Dataset S3. List of S. fredii HH103 induced genes in the presence of 400 mM mannitol, ordered by replicon. Supplementary Dataset S4. List of the different AHLs detected as produced by S. fredii HH103 grown in the absence (269) or presence (269_Manitol) of 400 mM mannitol.
Author Contributions
Conceptualization, F.J.O. and J.-M.V.; investigation, F.F.-R., P.A.-G., I.M.-B., M.Á.R.-C. and S.A.-J.; data analyses, F.P.-M. and S.A.-J.; writing-review and editing, S.A.-J., F.P.-M., F.J.O. and J.-M.V.; funding acquisition, J.-M.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the projects PID2019-107634RB-I00 from the Spanish “Ministerio de Ciencia, Innovación y Universidades” and by Grant US-1250546 funded by FEDER/Universidad de Sevilla.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The list of Nod Factors produced by S. fredii HH103 under the different conditions studied in this work are available as Supplementary Dataset S1. Fold-changes of all S. fredii HH103 genes in the presence of 400 mM mannitol regarding growth in YM medium are available as Supplementary Datasets S2 and S3. The list of the different AHLs detected as produced by S. fredii HH103 grown in the presence or absence of 400 mM mannitol is available as Supplementary Dataset 4. The RNA-Seq data discussed in this publication can be found in the Sequence Read Archive of NCBI (BioProject database) under BioProject IDs PRJNA911807 and PRJNA313151.
Acknowledgments
We thank the Biology and Mass Spectrometry Research Services of the “Centro de Investigación, Tecnología e Innovación” (CITIUS) of the University of Seville. We thank Catherine Jacott and Pablo del Cerro, both members of the Department of Microbiology of the University of Seville, for English language reviewing of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- López-Baena, F.J.; Ruiz-Sainz, J.E.; Rodríguez-Carvajal, M.A.; Vinardell, J.M. Bacterial molecular signals in the Sinorhizobium fredii-soybean symbiosis. Int. J. Mol. Sci. 2016, 17, 755. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.; Dickstein, R.; Udvardi, M.K. Celebrating 20 years of genetic discoveries in legume nodulation and symbiotic nitrogen fixation. Plant Cell. 2020, 32, 15–41. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lan, L.; Jin, Y.; Yu, N.; Wang, D.; Wang, E. Mechanisms underlying legume-rhizobium symbioses. J. Integr. Plant Biol. 2022, 64, 244–267. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, F.; Wall, L.; Fabra, A. Starting points in plant-bacteria nitrogen-fixing symbioses: Intercellular invasion of the roots. J. Exp. Bot. 2017, 68, 1905–1918. [Google Scholar] [CrossRef]
- Janczarek, M.; Rachwał, K.; Marzec, A.; Grzadziel, J.; Palusińska-Szysz, M. Signal molecules and cell-surface components involved in early stages of the legume-rhizobium interactions. Appl. Soil Ecol. 2015, 85, 94–113. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Fuentes-Romero, F.; Ruiz-Sainz, J.E.; Janzarek, M.; Vinardell, J.M. Rhizobial exopolysaccharides: Genetic regulation of their synthesis and relevance in symbiosis with legumes. Int. J. Mol.Sci. 2021, 22, 6233. [Google Scholar] [CrossRef]
- Fraysse, N.; Couderc, F.; Poinsot, V. Surface polysaccharide involvement in establishing the rhizobium-legume symbiosis. Eur. J. Biochem. 2003, 270, 1365–1380. [Google Scholar] [CrossRef]
- Jiménez-Guerrero, I.; Medina, C.; Vinardell, J.M.; Ollero, F.J.; López-Baena, F.J. The rhizobial type 3 secretion system: The Dr. Jekyll and Mr. Hyde in the rhizobium-legume symbiosis. Int. J. Mol. Sci. 2022, 23, 11089. [Google Scholar] [CrossRef]
- Pérez-Montaño, F.; del Cerro, P.; Jiménez-Guerrero, I.; López-Baena, G.J.; Cubo, M.T.; Hungria, M.; Megías, M.; Ollero, F.J. RNA-seq analysis of the Rhizobium tropici CIAT 899 transcriptome shows similarities in the activation patterns of symbiotic genes in the presence of apigenin and salt. BMC Genom. 2016, 17, 198. [Google Scholar] [CrossRef]
- Pérez-Montaño, F.; Guasch-Vidal, B.; González-Barroso, S.; López-Baena, F.J.; Cubo, T.; Ollero, F.J.; Gil-Serrano, A.M.; Rodríguez-Carvajal, M.A.; Bellogín, R.A.; Espuny, M.R. Nodulation-gene-inducing flavonoids increase overall production of autoinducers and expression of N-acyl homoserine lactone synthesis genes in rhizobia. Res. Microbiol. 2011, 162, 715–723. [Google Scholar] [CrossRef]
- Del Cerro, P.; Rolla-Santos, A.A.P.; Valderrama-Fernández, R.; Gil-Serrano, A.M.; Bellogín, R.A.; Gomes, D.F.; Pérez-Montaño, F.; Megías, M.; Hungria, M.; Ollero, F.J. NrcR, a new transcriptional regulator of Rhizobium tropici CIAT 899 involved in the legume root-nodule symbiosis. PLoS ONE 2016, 11, e0154029. [Google Scholar] [CrossRef]
- del Cerro, P.; Megías, M.; López-Baena, F.J.; Gil-Serrano, M.; Pérez-Montaño, F.; Ollero, F.J. Osmotic stress activates nif and fix genes and induces the Rhizobium tropici CIAT 899 Nod factor production via NodD2 by up-regulation of the nodA2 operon and the nodA3 gene. PLoS ONE 2019, 14, e0213298. [Google Scholar] [CrossRef]
- del Cerro, P.; Rolla-Santos, A.A.P.; Gomes, D.F.; Marks, B.; Pérez-Montaño, F.; Rodríguez-Carvajal, M.A.; Gil-Serrano, A.; Megías, M.; Ollero, F.J.; Hungría, M. Regulatory nodD1 and nodD2 genes of Rhizobium tropici strain CIAT 899 and their role in early steps of molecular signaling and host nodulation. BMC Genom. 2015, 16, 251. [Google Scholar] [CrossRef]
- del Cerro, P.; Rolla-Santos, A.A.; Gomes, D.F.; Marks, B.B.; Espuny, M.R.; Rodríguez-Carvajal, M.A.; Soria-Díaz, M.E.; Nakatani, A.S.; Hungría, M.; Ollero, F.J.; et al. Opening the “black box” of nodD3, nodD4 and nodD5 genes of Rhizobium tropici strain CIAT 899. BMC Genom. 2015, 16, 864. [Google Scholar] [CrossRef]
- Del Cerro, P.; Pérez-Montaño, F.; Gil-Serrano, A.; López-Baena, F.J.; Megías, M.; Hungria, M.; Ollero, F.J. The Rhizobium tropici CIAT 899 NodD2 protein regulates the production of Nod factors under salt stress in a flavonoid-independent manner. Sci. Rep. 2017, 7, 46712. [Google Scholar] [CrossRef]
- del Cerro, P.; Ayala-García, P.; Buzón, P.; Castells-Graells, R.; López-Baena, F.J.; Ollero, F.J.; Pérez-Montaño, F. OnfD, an AraC-type transcriptional regulator encoded by Rhizobium tropici CIAT 899 and involved in Nod factor synthesis and symbiosis. Appl. Environ. Microbiol. 2020, 86, e01297-20. [Google Scholar] [CrossRef]
- Dowdle, S.F.; Bohlool, B.B. Predominance of fast-growing Rhizobium japonicum in a soybean field in the People’s Republic of China. Appl. Environ. Microbiol. 1985, 50, 1171–1176. [Google Scholar] [CrossRef]
- Margaret, I.; Becker, A.; Blom, J.; Bonilla, I.; Groesmann, A.; Göttfert, M.; Lloret, J.; Mittard-Runte, V.; Rückert, C.; Ruiz-Sainz, J.E.; et al. Symbiotic properties and first analyses of the genomic sequence of the fast growing model strain Sinorhizobium fredii HH103 nodulating soybean. J. Biotechnol. 2011, 155, 11–19. [Google Scholar] [CrossRef]
- Gil-Serrano, A.M.; Franco-Rodríguez, G.; Tejero-Mateo, P.; Thomas-Oates, J.; Spaink, H.P.; Ruiz-Sainz, J.E.; Megías, M.; Lamrabet, Y. Structural determination of the lipo-chitinoligosaccharide nodulation signals produced by Rhizobium fredii HH103. Carbohydr. Res. 1997, 303, 435–443. [Google Scholar] [CrossRef]
- Vinardell, J.M.; López-Baena, F.J.; Hidalgo, A.; Ollero, F.J.; Bellogín, R.; Espuny, M.R.; Temprano, F.; Romero, F.; Krishnan, H.B.; Pueppke, S.G.; et al. The effect of FITA mutations on the symbiotic properties of Sinorhizobium fredii varies in a chromosomal-background-dependent manner. Arch. Microbiol. 2004, 181, 144–154. [Google Scholar] [CrossRef]
- Fuentes-Romero, F.; Navarro-Gómez, P.; Ayala-García, P.; Moyano-Bravo, I.; López-Baena, F.J.; Pérez-Montaño, F.; Ollero, F.J.; Acosta-Jurado, S.; Vinardell, J.M. The nodD1 gene of Sinorhizobium fredii HH103 restores nodulation capacity on bean in a Rhizobium tropici CIAT 899 nodD1/nodD2 mutant, but the secondary symbiotic regulators nolR, nodD2 or syrM prevent HH103 to nodulate with this legume. Microorganisms 2022, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.; Pueppke, S.G.; Vinardell, J.M.; Ruiz-Sainz, J.E.; Krishnan, H.B. Expression of nodD1 and nodD2 in Sinorhizobium fredii, a nitrogen-fixing symbiont of soybean and other legumes. Mol. Plant-Microbe Interact. 1998, 11, 375–382. [Google Scholar] [CrossRef]
- Vinardell, J.M.; Ollero, F.J.; Hidalgo, A.; López-Baena, F.J.; Medina, C.; Ivanov-Vangelov, K.; Parada, M.; Madinabeitia, N.; Espuny, M.R.; Bellogín, R.A.; et al. NolR regulates diverse symbiotic signals of Sinorhizobium fredii HH103. Mol. Plant-Microbe Interact. 2004, 17, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Jurado, S.; Alías-Villegas, C.; Navarro-Gómez, P.; Zehner, S.; Murdoch, P.D.; Rodríguez-Carvajal, M.A.; Soto, M.J.; Ollero, F.J.; Ruiz-Sainz, J.E.; Göttfert, M.; et al. The Sinorhizobium fredii HH103 MucR1 global regulator is connected with the nod regulon and is required for efficient symbiosis with Lotus burttii and Glycine max cv. Williams. Mol. Plant-Microbe Interact. 2016, 29, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Jurado, S.; Rodríguez-Navarro, D.N.; Kawaharada, Y.; Rodríguez-Carvajal, M.A.; Gil-Serrano, A.; Soria-Díaz, M.E.; Pérez-Montaño, F.; Fernández-Perea, J.; Niu, Y.; Alías-Villegas, C.; et al. Sinorhizobium fredii HH103 nolR and nodD2 mutants gain capacity for infection thread invasion of Lotus japonicus Gifu and Lotus burttii. Environ. Microbiol. 2019, 21, 1718–1739. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Rodríguez-Navarro, D.N.; Kawaharada, Y.; Perea, J.F.; Gil-Serrano, A.; Jin, H.; An, Q.; Rodríguez-Carvajal, M.A.; Andersen, S.U.; Sandal, N.; et al. Sinorhizobium fredii HH103 invades Lotus burttii by crack entry in a Nod factor-and surface polysaccharide-dependent manner. Mol. Plant-Microbe Interact. 2016, 29, 925–937. [Google Scholar] [CrossRef]
- Di Lorenzo, F.; Speciale, I.; Silipo, A.; Alías-Villegas, C.; Acosta-Jurado, S.; Rodríguez-Carvajal, M.A.; Dardanelli, M.S.; Palmigiano, A.; Garozzo, D.; Ruiz-Sainz, J.E.; et al. Structure of the unusual Sinorhizobium fredii HH103 lipopolysaccharide and its role in symbiosis. J. Biol. Chem. 2020, 295, 10969–10987. [Google Scholar] [CrossRef]
- Jiménez-Guerrero, I.; Pérez-Montaño, F.; Monreal, J.A.; Preston, G.M.; Fones, H.; Vioque, B.; Ollero, F.J.; López-Baena, F.J. The Sinorhizobium (Ensifer) fredii HH103 type 3 secretion system suppresses early defense responses to effectively nodulate soybean. Mol. Plant-Microbe Interact. 2015, 28, 790–7909. [Google Scholar] [CrossRef]
- Jiménez-Guerrero, I.; Pérez-Montaño, F.; Medina, C.; Ollero, F.J.; López-Baena, F.J. NopC is a Rhizobium-specific type 3 secretion system effector secreted by Sinorhizobium (Ensifer) fredii HH103. PLoS ONE 2015, 10, e0142866. [Google Scholar] [CrossRef]
- Jiménez-Guerrero, I.; Pérez-Montaño, F.; Medina, C.; Ollero, F.J.; López-Baena, F.J. The Sinorhizobium (Ensifer) fredii HH103 nodulation outer protein NopI Is a determinant for efficient nodulation of soybean and cowpea plants. Appl. Environ. Microbiol. 2017, 83, e02770-16. [Google Scholar] [CrossRef]
- Jiménez-Guerrero, I.; Pérez-Montaño, F.; Zdyb, A.; Beutler, M.; Werner, G.; Göttfert, M.; Ollero, F.J.; Vinardell, J.M.; López-Baena, F.J. GunA of Sinorhizobium (Ensifer) fredii HH103 is a T3SS-secreted cellulase that differentially affects symbiosis with cowpea and soybean. Plant Soil 2019, 435, 15–26. [Google Scholar] [CrossRef]
- Jiménez-Guerrero, I.; Acosta-Jurado, S.; Medina, C.; Ollero, F.J.; Alías-Villegas, C.; Vinardell, J.M.; Pérez-Montaño, F.; López-Baena, F.J. The Sinorhizobium fredii HH103 type III secretion system effector NopC blocks nodulation with Lotus japonicus Gifu. J. Exp. Bot. 2020, 71, 6043–6056. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Alías-Villegas, C.; Almozara, A.; Espuny, M.R.; Vinardell, J.M.; Pérez-Montaño, F. Deciphering the symbiotic significance of quorum sensing systems of Sinorhizobium fredii HH103. Microorganisms 2020, 8, 68. [Google Scholar] [CrossRef]
- Lamrabet, Y.; Bellogín, R.A.; Cubo, T.; Espuny, R.; Gil, A.; Krishnan, H.B.; Megias, M.; Ollero, F.J.; Pueppke, S.G.; Ruiz-Sainz, J.E.; et al. Mutation in GDP-fucose synthesis genes of Sinorhizobium fredii alters Nod factors and significantly decreases competitiveness to nodulate soybeans. Mol. Plant-Microbe Interact. 1999, 12, 207–217. [Google Scholar] [CrossRef]
- López-Baena, F.J.; Vinardell, J.M.; Pérez-Montaño, F.; Crespo-Rivas, J.C.; Bellogín, R.A.; Espuny, M.R.; Ollero, F.J. Regulation and symbiotic significance of nodulation outer proteins secretion in Sinorhizobium fredii HH103. Microbiol. Read. 2008, 154, 1825–1836. [Google Scholar] [CrossRef]
- Beringer, J.E. R factor transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 1974, 84, 188–198. [Google Scholar] [CrossRef]
- Nutman, P.S. The Modified Fåhraeus Slide Technique. In A Manual for the Practical Study of Root Nodule Bacteria. Handbook, Number 15, Appendix III; Vincent, J.M., Ed.; Blackwell Scientific: Oxford, UK; Edinburgh, UK, 1970; pp. 144–145. [Google Scholar]
- Spaink, H.P.; Aarts, A.; Stacey, G.; Bloemberg, G.V.; Lugtenberg, B.J.J.; Kennedy, E.P. Detection and separation of Rhizobium and Bradyrhizobium Nod metabolites using thin-layer chromatography. Mol. Plant-Microbe Interact. 1992, 5, 72–80. [Google Scholar] [CrossRef]
- Robertsen, B.K.; Aiman, P.; Darwill, A.G.; Mcneil, M.; Albersheim, P. The structure of acidic extracellular polysaccharides secreted by Rhizobium leguminosarum and Rhizobium trifolii. Plant Physiol. 1981, 67, 389–400. [Google Scholar] [CrossRef]
- Sourjik, V.; Schmitt, R. Different roles of CheY1 and CheY2 in the chemotaxis of Rhizobium meliloti. Mol. Microbiol. 1996, 22, 427–436. [Google Scholar] [CrossRef]
- Farrand, S.K.; Qin, Y.; Oger, P. Quorum-sensing system of Agrobacterium plasmids: Analysis and utility. Methods Enzymol. 2002, 358, 452–484. [Google Scholar] [CrossRef]
- Miller, J.H. Experiment in Molecular Genetics; Cold Spring Harbor: New York, NY, USA, 1972. [Google Scholar]
- de Maagd, R.A.; Wijffelman, C.A.; Pees, E.; Lugtenberg, B.J.J. Detection and subcellular localization of two Sym plasmid dependent proteins of Rhizobium leguminosarum biovar viciae. J. Bacteriol. 1998, 170, 4424–4427. [Google Scholar] [CrossRef] [PubMed]
- López-Lara, I.M.; van Der Drift, K.M.; van Brussel, A.A.; Haverkamp, J.; Lugtenberg, B.J.; Thomas-Oates, J.E.; Spaink, H.P. Induction of nodule primordia on Phaseolus and Acacia by lipo-chitin oligosaccharide nodulation signals from broad-host-range Rhizobium strain GRH2. Plant Mol. Biol. 1995, 29, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Truchet, G.; Camut, S.; de Billy, F.; Odorico, R.; Vasse, J. The Rhizobium-legume symbiosis. Two methods to discriminate between nodules and other root derived structures. Protoplasma 1989, 149, 82–89. [Google Scholar] [CrossRef]
- Pérez-Montaño, F.; Jiménez-Guerrero, I.; Acosta-Jurado, S.; Navarro-Gómez, P.; Ollero, F.J.; Ruiz-Sainz, J.E.; López-Baena, F.J.; Vinardell, J.M. A transcriptomic analysis of the effect of genistein on Sinorhizobium fredii HH103 reveals novel rhizobial genes putatively involved in symbiosis. Sci. Rep. 2016, 6, 31592. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map forma The Sequence alignment/map (SAM) format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 987–2993. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-a Phyton framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 3166–3169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Alías-Villegas, C.; Fuentes-Romero, F.; Cuéllar, V.; Navarro-Gómez, P.; Soto, M.J.; Vinardell, J.-M.; Acosta-Jurado, S. Surface Motility regulation of Sinorhizobium fredii HH103 by plant flavonoids and the NodD1, TtsI, NolR, and MucR1 symbiotic bacterial regulators. Int. J. Mol. Sci. 2022, 23, 7698. [Google Scholar] [CrossRef]
- Hidalgo, Á.; Margaret, I.; Crespo-Rivas, J.C.; Parada, M.; Murdoch, P.D.S.; López, A.; Buendía-Clavería, A.M.; Moreno, J.; Albareda, M.; Gil-Serrano, A.M.; et al. The rkpU gene of Sinorhizobium fredii HH103 is required for bacterial K-antigen polysaccharide production and for efficient nodulation with soybean but not with cowpea. Microbiol. Read. 2010, 156, 3398–3411. [Google Scholar] [CrossRef]
- Parada, M.; Vinardell, J.M.; Ollero, F.J.; Hidalgo, A.; Gutiérrez, R.; Buendía-Clavería, A.M.; Lei, W.; Margaret, I.; López-Baena, F.J.; Gil-Serrano, A.M.; et al. Sinorhizobium fredii HH103 mutants affected in capsular polysaccharide (KPS) are impaired for nodulation with soybean and Cajanus cajan. Mol. Plant-Microbe Interact. 2006, 19, 43–52. [Google Scholar] [CrossRef]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef]
- Cha, C.; Gao, P.; Chen, Y.C.; Shaw, P.D.; Farrand, S.K. Production of acyl-homoserine lactone quorum-sensing signals by gram-negative plant associated bacteria. Mol. Plant-Microbe Interact. 1998, 11, 1119–1129. [Google Scholar] [CrossRef]
- Vinardell, J.M.; Acosta-Jurado, S.; Göttfert, M.; Zehner, S.; Becker, A.; Baena-Ropero, I.; Blom, J.; Crespo-Rivas, J.C.; Goesmann, A.; Jaenicke, S.; et al. The Sinorhizobium fredii HH103 genome: A comparative analysis with S. fredii strains differing in their symbiotic behaviour with soybean. Mol. Plant-Microbe Interact. 2015, 28, 811–824. [Google Scholar] [CrossRef]
- Madinabeitia, N.; Bellogín, R.A.; Buendía-Clavería, A.M.; Camacho, M.; Cubo, T.; Espuny, M.R.; Gil-Serrano, A.M.; Lyra, M.C.; Moussaid, A.; Ollero, F.J.; et al. Sinorhizobium fredii HH103 has a truncated nolO gene due to a -1 frameshift mutation that is conserved among other geographically distant S. fredii strains. Mol. Plant-Microbe Interact. 2002, 15, 150–159. [Google Scholar] [CrossRef]
- Rodríguez-Navarro, D.N.; Rodríguez-Carvajal, M.A.; Acosta-Jurado, S.; Soto, M.J.; Margaret, I.; Crespo-Rivas, J.C.; Sanjuan, J.; Temprano, F.; Gil-Serrano, A.; Ruiz-Sainz, J.E.; et al. Structure and biological roles of Sinorhizobium fredii HH103 exopolysaccharide. PLoS ONE 2014, 9, e115391. [Google Scholar] [CrossRef]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef]
- Hawkins, J.P.; Oresnik, I.J. The rhizobium-legume symbiosis: Co-opting successful stress management. Front. Plant Sci. 2022, 12, 796045. [Google Scholar] [CrossRef]
- Kumar, C.M.; Mande, S.C.; Mahajan, G. Multiple chaperonins in bacteria-novel functions and non-canonical behaviors. Cell Stress Chaperones 2015, 20, 555–574. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.; Achom, M.; Wilkinson, H.; Lagunas, B.; Gifford, M.L. Symbiotic outcome modified by the diversification from 7 to over 700 nodule-specific cysteine-rich peptides. Genes 2020, 11, 348. [Google Scholar] [CrossRef] [PubMed]
- Bittner, A.N.; Foltz, A.; Oke, V. Only one of five groEL genes is required for viability and successful symbiosis in Sinorhizobium meliloti. J. Bacteriol. 2007, 189, 1884–1889. [Google Scholar] [CrossRef]
- Wei, F.; Liu, Y.; Zhou, D.; Zhao, W.; Chen, Z.; Chen, D.; Li, Y.; Zhang, X.X. Transcriptomic identification of a unique set of nodule-specific cysteine-rich peptides expressed in the nitrogen-fixing root nodule of Astragalus sinicus. Mol. Plant-Microbe Interact. 2022, 35, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Margaret-Oliver, I.; Lei, W.; Parada, M.; Rodríguez-Carvajal, M.A.; Crespo-Rivas, J.C.; Hidalgo, A.; Gil-Serrano, A.; Moreno, J.; Rodríguez-Navarro, D.N.; Buendía-Clavería, A.; et al. Sinorhizobium fredii HH103 does not strictly require KPS and/or EPS to nodulate Glycyrrhiza uralensis, an indeterminate nodule-forming legume. Arch. Microbiol. 2012, 192, 87–102. [Google Scholar] [CrossRef]
- Glazebrook, J.; Reed, J.W.; Reuber, T.L.; Walker, G.C. Genetic analyses of Rhizobium meliloti exopolysaccharides. Int. J. Biol. Macromol. 1990, 12, 67–70. [Google Scholar] [CrossRef]
- Barnett, M.J.; Long, S.R. The Sinorhizobium meliloti SyrM regulon: Effects on global gene expression are mediated by syrA and nodD3. J. Bacteriol. 2015, 197, 1792–17806. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Navarro-Gómez, P.; Murdoch, P.; Del, S.; Crespo-Rivas, J.C.; Jie, S.; Cuesta-Berrio, L.; Ruiz-Sainz, J.E.; Rodríguez-Carvajal, M.A.; Vinardell, J.M. Exopolysaccharide production by Sinorhizobium fredii HH103 is repressed by genistein in a NodD1-dependent manner. PLoS ONE 2016, 11, e0160499. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Alias-Villegas, C.; Navarro-Gómez, P.; Almozara, A.; Rodríguez-Carvajal, M.A.; Medina, C.; Vinardell, J.M. Sinorhizobium fredii HH103 syrM inactivation affects the expression of a large number of genes, impairs nodulation with soybean and extends the host-range to Lotus japonicus. Environ. Microbiol. 2020, 22, 1104–1124. [Google Scholar] [CrossRef]
- Breedveld, M.W.; Miller, K.J. Cyclic beta-glucans of members of the family Rhizobiaceae. Microbiol. Rev. 1994, 58, 145–161. [Google Scholar] [CrossRef]
- Crespo-Rivas, J.C.; Margaret, I.; Hidalgo, A.; Buendía-Clavería, A.M.; Ollero, F.J.; López-Baena, F.J.; del Socorro Murdoch, P.; Rodríguez-Carvajal, M.A.; Soria-Díaz, M.E.; Reguera, M.; et al. Sinorhizobium fredii HH103 cgs mutants are unable to nodulate determinate- and indeterminate nodule-forming legumes and overproduce an altered EPS. Mol. Plant-Microbe Interact. 2009, 22, 575–588. [Google Scholar] [CrossRef]
- Wheatley, R.M.; Ford, B.L.; Li, L.; Aroney, S.T.N.; Knights, H.E.; Ledermann, R.; East, A.K.; Ramachandran, V.K.; Poole, P.S. Lifestyle adaptations of Rhizobium from rhizosphere to symbiosis. Proc. Natl. Acad. Sci. USA 2020, 117, 23823–23834. [Google Scholar] [CrossRef]
- Aroney, S.T.N.; Poole, P.S.; Sánchez-Cañizares, C. Rhizobial chemotaxis and motility systems at work in the soil. Front. Plant Sci. 2021, 12, 725338. [Google Scholar] [CrossRef]
- Alemneh, A.A.; Zhou, Y.; Ryder, M.H.; Denton, M.D. Mechanisms in plant growth-promoting rhizobacteria that enhance legume-rhizobial symbioses. J. Appl. Microbiol. 2020, 129, 1133–1156. [Google Scholar] [CrossRef]
- Calatrava-Morales, N.; McIntosh, M.; Soto, M.J. Regulation mediated by N-acyl homoserine lactone quorum sensing signals in the Rhizobium-legumes symbiosis. Genes 2018, 9, 263. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).