
Assessing the Impact of Anthropic Pressures on Aquatic Macroinvertebrates: A Functional Trait Approach in the Irtysh River Watershed

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
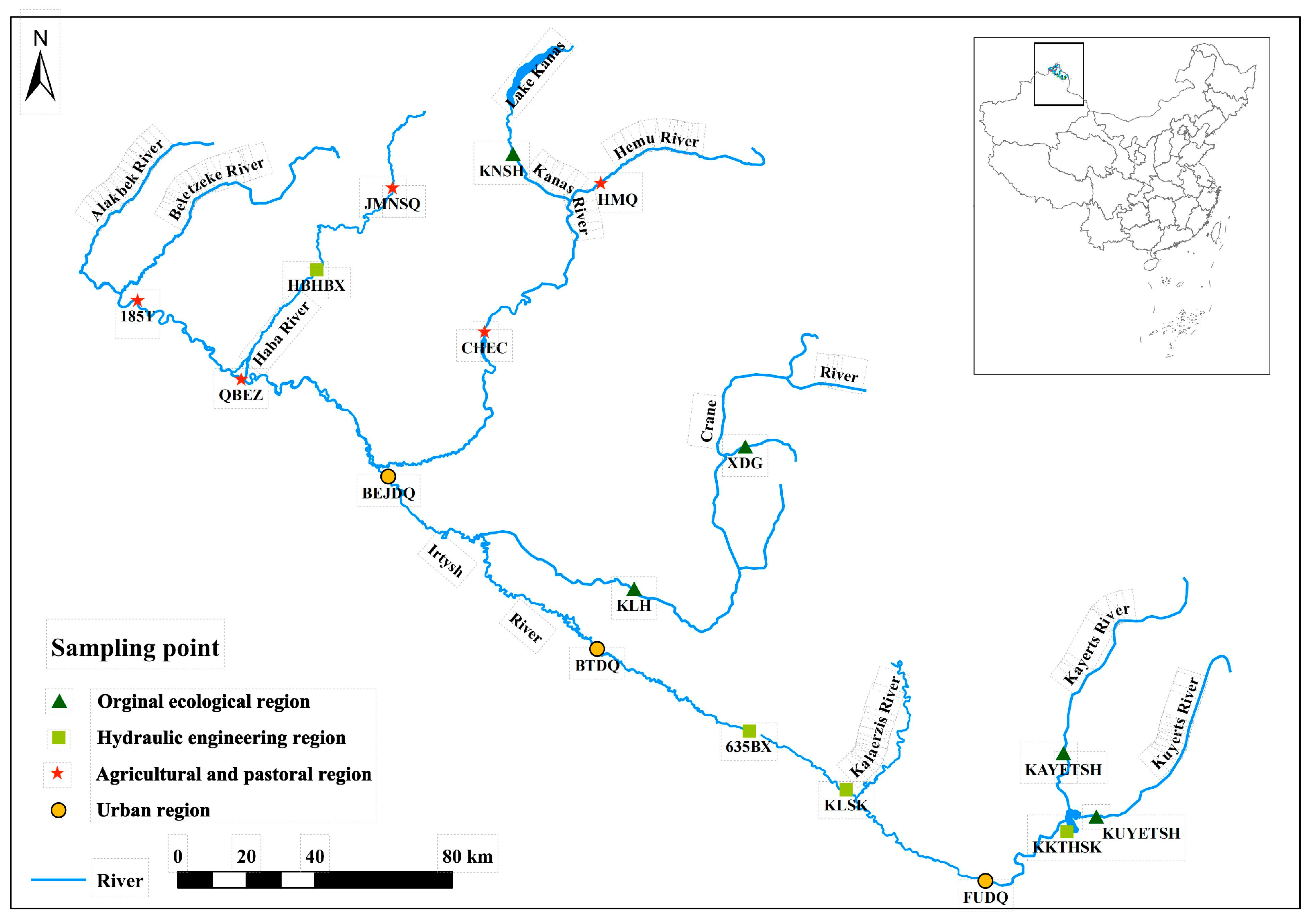
2.1. Field Sampling and Data Acquisition
2.2. Data Analysis
2.2.1. Physico-Chemical, Biological and Trait Analysis
2.2.2. Functional Diversity and Community-Weighted Means
3. Results
3.1. Environmental Factors and Family Distribution
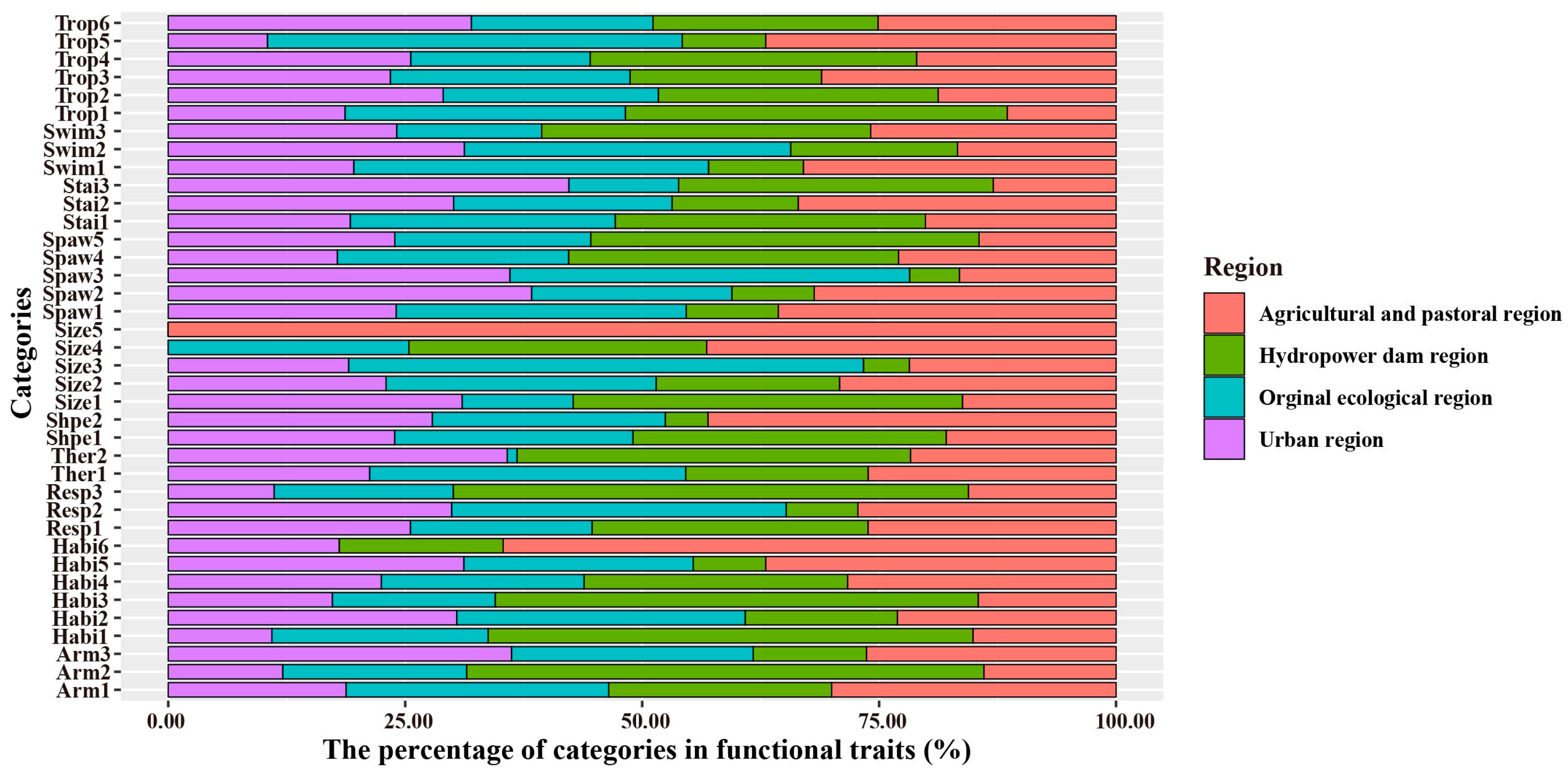
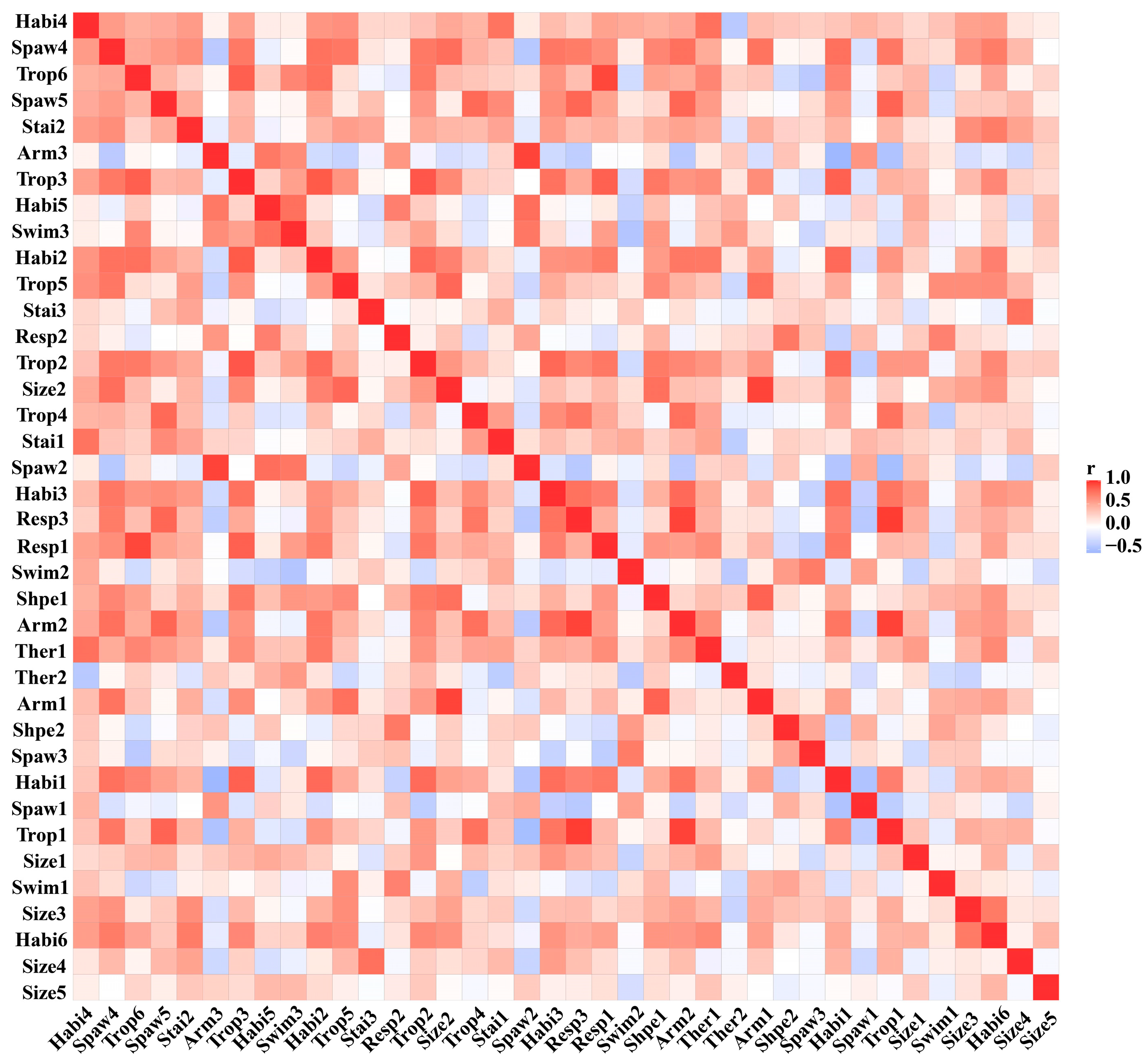
3.2. Biological Traits
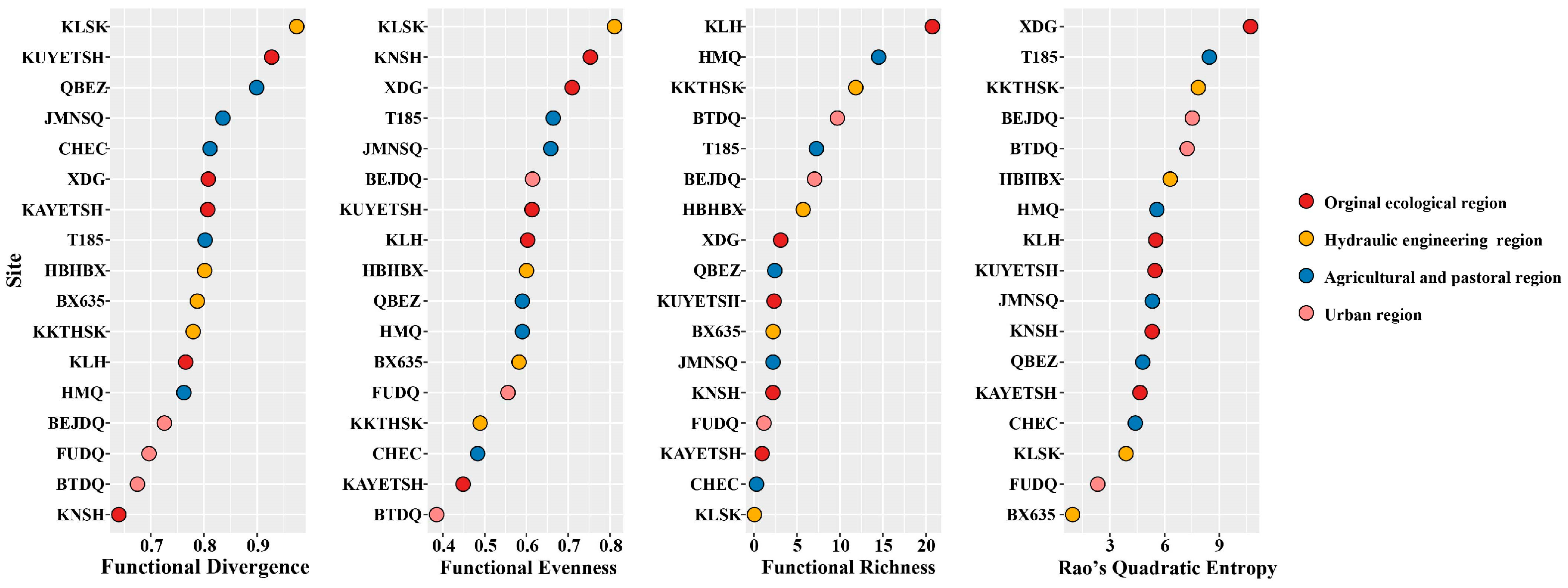
3.3. Functional Diversity and Community Weighted Means
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Section |
|---|---|
| SP1 | Lymnaeidae |
| SP2 | Elmidae |
| SP3 | Ephemerellidae |
| SP4 | Siphlonuridae |
| SP5 | Tipulidae |
| SP6 | Gammaridae |
| SP7 | Orthocladiinae |
| SP8 | Leptoceridae |
| SP9 | Chironominae |
| SP10 | Lumbriculidae |
| SP11 | Leptophlebiidae |
| SP12 | Baetidae |
| SP13 | Haplotaxidae |
| SP14 | Ephemeridae |
| SP15 | Psychomyiidae |
| SP16 | Heptageniidae |
| SP17 | Planorbidae |
| SP18 | Hemiclepsis |
| SP19 | Nemouridae |
| SP20 | Taeniopterygidae |
| SP21 | Philopotamidae |
| SP22 | Brachycentridae |
| SP23 | Polycentropodidae |
| SP24 | Psychodidae |
| SP25 | Aphelocheiridae |
| SP26 | Muscidae |
| SP27 | Potamanthidae |
| SP28 | Capniidae |
| SP29 | Corixidae |
| SP30 | Ameletidae |
| SP31 | Perlidae |
| SP32 | Dytiscidae |
| SP33 | Chloroperlidae |
| SP34 | Cetatqogoridae |
| SP35 | Libellulidae |
| SP36 | Simuliidae |
| SP37 | Caropterygldae |
| SP38 | Glossosomatidae |
| SP39 | Glossiphoniidae |
| SP40 | Erpobdellidae |
| SP41 | Hirudidae |
| SP42 | Perlodidae |
| SP43 | Corduliidae |
| SP44 | Hydropsychidae |
| SP45 | Rhyacophilidae |
| SP46 | Polichopudidae |
| SP47 | Tanypodinae |
| SP48 | Limoniidae |
| SP49 | Arachnida |
References
- García–Girón, J.; Tolonen, K.T.; Soininen, J.; Snåre, H.; Pajunen, V.; Heino, J. Antropogenic landuse inpacts on the size structure of macroinvertebrate assemblages are jointly modulated by local conditions and spatial processes. Environ. Res. 2022, 204 Pt B, 112055. [Google Scholar] [CrossRef]
- Feng, Q.; Endo, K.N.; Cheng, G.D. Towards sustainable development of the environmentally degraded arid rivers of China—A case study from Tarim River. Environ. Geol. 2001, 41, 229–238. [Google Scholar] [CrossRef]
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef]
- Bian, L.; Wang, J.; Liu, J.; Han, B. Spatiotemporal changes of soil salinization in the Yellow River Delta of China from 2015 to 2019. Sustainability 2021, 13, 822. [Google Scholar] [CrossRef]
- Yan, Z.; Zhou, Z.; Liu, J.; Wang, H.; Li, D. Water use characteristics and impact factors in the Yellow River Basin, China. Water Int. 2021, 28, 148–168. [Google Scholar] [CrossRef]
- Shuai, F.M.; Li, X.H.; Liu, Q.F.; Li, Y.F.; Yang, J.P.; Li, J.; Chen, F.C. Spatial patterns of fish diversity and distribution in the Pearl River. Sheng Tai Xue Bao 2017, 37, 3182–3192. [Google Scholar] [CrossRef]
- Ali, R.; Kuriqi, A.; Abubaker, S.; Kisi, O. Hydrologic alteration at the upper and middle part of the Yangtze River, China: Towards sustainable water resource management under increasing water exploitation. Sustainability 2019, 11, 5176. [Google Scholar] [CrossRef]
- Gaglio, M.; Aschonitis, V.G.; Gissi, E.; Castaldelli, G.; Fano, E.A. Land use change effects on ecosystem services of river deltas and coastal wetlands: Case study in Volano–Mesola–Goro in Poriver delta (Italy). Wetlands Ecol. Manag. 2017, 25, 67–86. [Google Scholar] [CrossRef]
- Gál, B.; Szivák, I.; Heino, J.; Schmera, D. The effrcts of urbanization on freshwater macroinvertebrates knowledge gaps and future research directions. Ecol. Indic. 2019, 104, 357–364. [Google Scholar] [CrossRef]
- Huang, Y.; Zhou, B.; Li, N.; Li, Y.; Han, R.; Qi, J.; Lu, X.; Li, S.; Feng, C.; Liang, S. Spatial-temporal analysis of selected industrial aquatic heavy metal pollution in China. J. Clean. Prod. 2019, 238, 117944. [Google Scholar] [CrossRef]
- Yang, S.; Hao, H.; Liu, B.; Wang, Y.; Yang, Y.; Liang, R.; Li, K. Influence of socioeconomic development on river water quality: A case study of two river basins in China. Environ. Sci. Pollut. Res. Int. 2021, 28, 53857–53871. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, M.; Su, F.; Zhang, X.; Gao, S.; Xu, M. Changes in land cover and ecological stress in Borneo based on remote sensing and an ecological footprint method. Landsc. Ecol. Eng. 2020, 16, 319–333. [Google Scholar] [CrossRef]
- Jiang, Z.F.; Huo, T.B. Survey and Study on the Aquatic Species Resources in Irtysh River, Tarim River and Ulungur Lake in Xinjiang Urgur Autonomous Region; China Environmental Science Press: Beijing, China, 2013; pp. 24–127. ISBN 978-7-5111-1475-4. [Google Scholar]
- Wang, D.; Zhang, S.; Wang, G.; Gu, J.; Wang, H.; Chen, X. Ecohydrological variation and multi-objective ecological water demand of the Irtysh River Basin. Water 2022, 14, 2876. [Google Scholar] [CrossRef]
- Ullrich, S.M.; Ilyushchenko, M.A.; Kamberov, I.M.; Tanton, T.W. Mercury contamination in the vicinity of a derelict chlor-alkali plant. Part I: Sediment and water contamination of Lake Balkyldak and the River Irtysh. Sci. Total Environ. 2007, 381, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Huang, Q.; Liu, D.; Deng, M.; Zhang, H.; Pan, B.; Zhang, H. Long-term and mid-term ecological operation of cascade hydropower plants considering ecological water demands in arid region. J. Clean. Prod. 2021, 279, 1235999. [Google Scholar] [CrossRef]
- Dittrich, A.; Buerkert, A.; Brinkmann, K. Assessment of land use and land cover changes during the last 50 years in oases and surrounding rangelands of Xinjiang, NW China. J. Agric. Rural. Dev. Trop. 2010, 111, 129–142. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, Y.; Wang, M.X.; Wang, F.; Tang, Z.; He, H. Spatial and temporal distribution characteristics of drought and its relationship with meteorological factors in Xinjiang in last 17 years. Sheng Tai Xue Bao 2020, 40, 1077–1088. [Google Scholar] [CrossRef]
- Gusev, E.M.; Nasonova, O.N.; Shurkhno, E.A.; Dzhogan, L.Y. Scenario forecasting of changes in water balance components in the Ob–Irtysh Basin in the context of possible climate change. Water Resour. 2019, 46, 647–658. [Google Scholar] [CrossRef]
- Urbanič, G.; Mihaljević, Z.; Petkovska, V.; Urbanič, M.P. Disentangling the effects of multiple stressors on large rivers using benthic invertebrates-a study of southeastern European large rivers with implications for management. Int. J. Appl. Mech. 2020, 12, 621. [Google Scholar] [CrossRef]
- Desrosiers, M.; Usseglio-Polatera, P.; Archaimbault, V.; Larras, F.; Méthot, G.; Pinel-Alloul, B. Assessing anthropic pressure in the St. Lawrence River using traits of benthic macroinvertebrates. Sci. Total Environ. 2019, 649, 233–246. [Google Scholar] [CrossRef]
- Ji, F.; Han, D.; Yan, L.; Yan, S.; Zha, J.; Shen, J. Assessment of benthic invertebrate diversity and river ecological status along an urbanized gradient using environmental DNA metabarcoding and a traditional survey method. Sci. Total Environ. 2022, 806 Pt 2, 150587. [Google Scholar] [CrossRef]
- Vitecek, S.; Johnson, R.K.; Poikane, S. Assessing the ecological status of European rivers and lakes using benthic invertebrate communities: A practical catalogue of metrics and methods. Water 2021, 13, 346. [Google Scholar] [CrossRef]
- Vaate, A.B.D.; Pavluk, T.I. Practicability of the index of trophic completeness for running waters. Hydrobiologia 2004, 519, 49–60. [Google Scholar] [CrossRef]
- Thomas, J.F.; Hall, T.J. A comparison of three methods of evaluating aquatic community impairment in streams. J. Freshw. Ecol. 2006, 21, 53–63. [Google Scholar] [CrossRef]
- Bady, P.; Doledec, S.; Fesl, C.; Gayraud, S.; Bacchi, M.; Scholl, F. Use of invertebrate traits for the biomonitoring of European large rivers: The effects of sampling effort on genus richness and functional diversity. Freshw. Biol. 2005, 50, 159–173. [Google Scholar] [CrossRef]
- Leah-Bêche, A.; Statzner, B. Richness gradients of stream invertebrates across the USA: Taxonomy and trait-based approaches. Biodivers Conserv. 2009, 18, 3909–3930. [Google Scholar] [CrossRef]
- Magliozzi, C.; Usseglio-Polatera, P.; Meyer, A.; Grabowski, R.C. Functional traits of hyporheic and benthic invertebrates reveal importance of wood-driven geomorphological processes in rivers. Funct. Ecol. 2019, 33, 1758–1770. [Google Scholar] [CrossRef]
- Bendary, R.E.; Ibrahim, S.M.; Goher, M.E.; Elsaied, H.E.; El Shabrawy, G.M.; El Mordy, M.A.; Khalil, M.T. Taxonomic and functional structure of macrobenthic invertebrate communities and their response to environmental variables along the subbranches of the Nile River (rayahs), Egypt. Environ. Sci. Pollut. Res. Int. 2023, 30, 28803–28817. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.J.; Bray, J.; Nichols, S.J.; Shenton, M.; Kaserzon, S.; Mac Nally, R.; Kefford, B.J. Sensitivity and specificity of macroinvertebrate responses to gradients of multiple agricultural stressors. Environ. Pollut. 2021, 291, 118092. [Google Scholar] [CrossRef]
- Duan, X.H.; Wang, Z.Y.; Xu, M.Z. Benthic Macroinvertebrate and Application in the Assessment of Streamecology; Tsinghua University Press: Beijing, China, 2010; pp. 113–167. ISBN 978-7-302-21860-9. [Google Scholar]
- Fekadu, M.B.; Agembe, S.; Kiptum, C.K.; Mingist, M. Impacts of anthropic activities on the benthic macroinvertebrate assemblages during the wet season in Kipsinende River, Kenya. Turk. J. Fish Aquat. Sci. 2022, 22, 1–10. [Google Scholar] [CrossRef]
- Sudarso, J.; Suryono, T.S.T.; Yoga, G.P.; Imroatusshoolikhah, I.; Ibrahim, A.; Sari, L.S.L.; Badjoeri, M.B.M.; Samir, O.S.O. Effect of anthropic activity on benthic macroinvertebrate functional feeding groups in small streams of west Sumatra, Indonesia. Sains Malays. 2022, 51, 3551–3566. [Google Scholar] [CrossRef]
- The State environmental protection administration the water and wastewater monitoring analysis method editorial board. In Water and Wastewater Monitoring and Analysis Methods, 4th ed.; China Environmental Science Press: Beijing, China, 2002; pp. 56–79. ISBN 978-7-8016-3400-9.
- Tachet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. The Invertebresd’eau Douce Systématique, Biologie, Écologie; China Water Conservancy and Hydropower Press: Beijing, China, 2015; pp. 17–158. ISBN 978-7-5170-3003-4. [Google Scholar]
- Pallottini, M.; Cappelletti, D.; Fabrizi, A.; Gaino, E.; Goretti, E.; Selvaggi, R.; Céréghino, R. Macroinvertebrate functional trait responses to chemical pollution in agricultural-industrial landscapes. River Res. Appl. 2017, 33, 505–513. [Google Scholar] [CrossRef]
- Mokany, K.; Ash, J.; Roxburgh, S. Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland. J. Ecol. 2008, 96, 884–893. [Google Scholar] [CrossRef]
- Somerfield, P.J.; Clarke, K.R.; Warwick, R.M.; Dulvy, N.K. Average functional distinctness as a measure of the composition of assemblages. ICES J. Mar. Sci. 2008, 65, 1462–1468. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.R. Diversity and dissimilarity coefficients: A unified approach. Theor. Popul. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K.; Studer, M.; Roudier, P.; Gonzalez, J.; Kozlowski, K.; Schubert, E.; et al. Finding Groups in Data: Cluster Analysis Extensions Rousseeuw etal. The Comprehensive R Archive Network Home Page. Available online: https://svn.r-project.org/R-packages/trunk/cluster/ (accessed on 27 August 2023).
- Dray, S.; Pettorelli, N.; Chessel, D. Matching data sets from two different spatial samplings. J. Veg. Sci. 2002, 13, 867–874. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. Measuring Functional Diversity (FD) from Multiple Traits, and Other Tools for Functional Ecology. The Comprehensive R Archive Network Home Page. Available online: https://cran.r-project.org/web/packages/FD/ (accessed on 27 August 2023).
- Chung, N.; Bae, M.-J.; Li, F.; Kwon, Y.-S.; Kwon, T.-S.; Kim, J.-S.; Park, Y.-S. Habitat characteristics and trophic structure of benthic macroinvertebrates in a forested headwater stream. J. Asia-Pac. Entomol. 2012, 15, 495–505. [Google Scholar] [CrossRef]
- Ow, L.F.; Whitehead, D.; Walcroft, A.S.; Turnbull, M.H. Seasonal variation in foliar carbon exchange in Pinus radiata and Populus deltoides: Respiration acclimates fully to changes in temperature but photosynthesis does not. Global Chang. Biol. 2010, 16, 288–302. [Google Scholar] [CrossRef]
- Bolam, S.; Garcia, C.; Eggleton, J.; Kenny, A.; Buhl-Mortensen, L.; Gonzalez-Mirelis, G.; van Kooten, T.; Dinesen, G.; Hansen, J.; Hiddink, J.; et al. Differences in biological traits composition of benthic assemblages between unimpacted habitats. Mar. Environ. Res. 2017, 126, 1–13. [Google Scholar] [CrossRef]
- Paz, L.; Rodriguez, M.; Gullo, B.; Capítulo, A.R. Impacts of urban and industrial pollution on functional traits of benthic macroinvertebrates: Are some traits advantageous for survival? Sci. Total Environ. 2022, 807 Pt 2, 150650. [Google Scholar] [CrossRef]
- Weigel, B.; Blenckner, T.; Bonsdorff, E. Maintained functional diversity in benthic communities in spite of diverging functional identities. Oikos 2016, 125, 1421–1433. [Google Scholar] [CrossRef]
- Lizarralde, Z.I.; Pittaluga, S.; Peronism, M. Changes of benthic macrofaunal composition on a tidal flat of Rio Gallegos estuary, argentina. Thalassas 2018, 34, 131–138. [Google Scholar] [CrossRef]
- Jiang, S.; Ren, L.; Yong, B.; Singh, V.P.; Yang, X.; Yuan, F. Quantifying the effects of climate variability and human activities on runoff from the Laohahe Basin in northern China using three different methods. Hydrol. Processes 2011, 25, 2492–2505. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, M.; Ren, L.; Xu, C.; Yuan, F.; Liu, Y.; Lu, Y.; Shen, H. A framework for quantifying the impacts of climate change and human activities on hydrological drought in a semiarid basin of northern China. Hydrol. Processes 2019, 33, 1075–1088. [Google Scholar] [CrossRef]
- Llanos, E.N.; Jaubet, M.L.; Rodolfo, E. The intertidal benthic community of SW Atlantic as an environmental indicator of 50 years of human-mediated changes. Mar. Biol. Res. 2019, 15, 458–475. [Google Scholar] [CrossRef]
- Gadawski, P.; Riss, H.W.; Płóciennik, M.; Meyer, E.I. City channel chironomids-benthic diversity in urban conditions. River Res. Appl. 2016, 32, 1978–1988. [Google Scholar] [CrossRef]
- Liu, X.; Ren, L.; Yuan, F.; Singh, V.P.; Fang, X.; Yu, Z.; Zhang, W. Quantifying the effect of land use and land cover changes on green water and blue water in northern part of China. Hydrol. Earth Syst. Sci. 2009, 13, 735–747. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, J.; He, R.; Jiang, N.; Jing, X. Runoff reduction due to environmental changes in the Sanchuanhe River Basin. Int. J. Sediment Res. 2008, 23, 174–180. [Google Scholar] [CrossRef]
- Li, C.W.; Kang, L. A new modified tennant method with spatial-temporal variability. Water Resour. Manag. 2014, 28, 4911–4926. [Google Scholar] [CrossRef]
- Mezgebu, A.; Lakew, A.; Lemma, B. Water quality assessment using benthic macroinvertebrates as bioindicators in streams and rivers around Sebeta, Ethiopia. Afr. J. Aquat. Sci. 2019, 44, 361–367. [Google Scholar] [CrossRef]
- Habersack, H.; Haspel, D.; Kondolf, M. Large rivers in the anthropocene: Insights and tools for understanding climatic, land use, and reservoir influences. Water Resour. Res. 2014, 50, 3641–3646. [Google Scholar] [CrossRef]
- Oliveira, J.; Sousa-Pinto, I.; Weber, G.; Bertocci, I. Urban vs. extra-urban environments: Scales of variation of intertidal benthic assemblages in north portugal. Mar. Environ. Res. 2014, 97, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Drury, B.; Rosi-Marshall, E.; Kelly, J.J. Wastewater treatment effluent reduces the abundance and diversity of benthic bacterial communities in urban and suburban rivers. Appl. Environ. Microbiol. 2013, 79, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.P.; Gayeski, L.M.; Tundis, J.G. Benthic macroinvertebrate community in the Sinos River drainage basin, Rio Grande do Sul, Brazil. Braz. J. Biol. 2016, 76, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Ren, Z.; Zhang, M.; Liu, X.; Peng, W. Sediment heavy metals and benthic diversities in Hun-Tai River, northeast of China. Environ. Sci. Pollut. Res. Int. 2017, 24, 10662–10673. [Google Scholar] [CrossRef] [PubMed]
- Tlig-Zouari, S.; Mami, T.; Maamouri, F. Structure of benthic macroinvertebrates and dynamics in the northern lagoon of Tunis. J. Mar. Biol. Assoc. UK 2009, 89, 1305–1318. [Google Scholar] [CrossRef]
- Smakhtin, V.U.; Shilpakar, R.L.; Hughes, D.A. Hydrology-based assessment of environmental flows: An example from Nepal. Hydrol. Sci. J. 2006, 51, 207–222. [Google Scholar] [CrossRef]
- Gao, Y.; Vogel, R.M.; Kroll, C.N.; Poff, N.L.; Olden, J.D. Development of representative indicators of hydrologic alteration. J. Hydrol. 2009, 374, 136–147. [Google Scholar] [CrossRef]
- Haghighi, T.A.; Klove, B. Development of a general river regime index (RRI) for intra-annual flow variation based on the unit river concept and flow variation end-points. J. Hydrol. 2013, 503, 169–177. [Google Scholar] [CrossRef]
- Utz, M.R.; Hilderbrand, R. Interregional variation in urbanization-induced geomorphic change and macroinvertebrate habitat colonization in headwater streams. J. N. Am. Benthol. Soc. 2011, 30, 25–37. [Google Scholar] [CrossRef]
- Thorne, R.; Williams, P. The response of benthic macroinvertebrate to pollution in developing countries: A multi-metric system of bioassessment. Freshw. Biol. 2003, 37, 671–686. [Google Scholar] [CrossRef]
- Yong, B.; Ren, L.; Hong, Y.; Gourley, J.J.; Chen, X.; Dong, J.; Wang, W.; Shen, Y.; Hardy, J. Spatial temporal changes of water resources in a Typical Semiarid Basin of north China over the past 50 years and assessment of possible natural and socioeconomic causes. J. Hydrometeorol. 2013, 14, 1009–1034. [Google Scholar] [CrossRef]
- Sellami, H.; Benabdallah, S.; La Jeunesse, I.; Vanclooster, M. Quantifying hydrological responses of small mediterranean catchments under climate change projections. Sci. Total Environ. 2016, 543 Pt B, 924–936. [Google Scholar] [CrossRef]
- Arman, N.Z.; Salmiati, S.; Said, M.I.M.; Aris, A. Development of macroinvertebrate based multimetric index and establishment of biocriteria for river health assessment in malaysia science direct. Ecol. Indic. 2019, 104, 449–458. [Google Scholar] [CrossRef]
- Zhao, C.; Yang, Y.; Yang, S.; Xiang, H.; Zhang, Y.; Wang, Z.; Chen, X.; Mitrovic, S. Predicting future river health in a minimally influenced mountainous area under climate change. Sci. Total Environ. 2019, 656, 1373–1385. [Google Scholar] [CrossRef]
- Yu, M.; Wood, P.; van de Giesen, N.; Liu, X.; Li, Q.; Wang, G.; Zhang, J. Enhanced potential ecological risk induced by a large scale water diversion project. Stoch. Environ. Res. Risk A 2020, 34, 2125–2138. [Google Scholar] [CrossRef]


| Trait | Category | Code | Abb |
|---|---|---|---|
| Trophic habit | Parasites | 1 | Trop1 |
| Predator | 2 | Trop2 | |
| Collector-gatherer | 3 | Trop3 | |
| Collector-filterer | 4 | Trop4 | |
| Scraper | 5 | Trop5 | |
| Shredder | 6 | Trop6 | |
| Habit | Burrower | 1 | Habi1 |
| Clinger | 2 | Habi2 | |
| Climber | 3 | Habi3 | |
| Sprawler | 4 | Habi4 | |
| Swimmer | 5 | Habi5 | |
| Divers | 6 | Habi6 | |
| Stain resistance value | Pollution-sensitive organisms < 3 | 1 | Stai1 |
| Semi-tolerant to pollution 3–7 | 2 | Stai2 | |
| Pollution-tolerant organisms > 7 | 3 | Stai3 | |
| Maximum size (mm) | 5–10 mm | 1 | Size1 |
| 10–20 mm | 2 | Size2 | |
| 20–40 mm | 3 | Size3 | |
| 40–80 mm | 4 | Size4 | |
| >80 mm | 5 | Size5 | |
| Reproduction | Isolated eggs, free | 1 | Spaw1 |
| Isolated eggs, cemented | 2 | Spaw2 | |
| Spawning | 3 | Spaw3 | |
| Aquatic spawning | 4 | Spaw4 | |
| Monogamy/Asexual reproduction | 5 | Spaw5 | |
| Respiration technique | Tegument | 1 | Resp1 |
| Gill | 2 | Resp2 | |
| Air (spiracles, tracheae, plastrons) | 3 | Resp3 | |
| Swimming ability | Weak | 1 | Swim1 |
| Poor | 2 | Swim2 | |
| strong | 3 | Swim3 | |
| Armoring | None | 1 | Arm1 |
| Poor | 2 | Arm2 | |
| Good | 3 | Arm3 | |
| Shape | Streamlined | 1 | Shpe1 |
| Not streamlined | 2 | Shpe2 | |
| Thermal preference | Stenothermy | 1 | Ther1 |
| Eurytherm | 2 | Ther2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Zi, F.; Wang, X.; Zeng, H.; Huo, B.; Wang, C.; Ge, J.; Chen, S.; Wang, B. Assessing the Impact of Anthropic Pressures on Aquatic Macroinvertebrates: A Functional Trait Approach in the Irtysh River Watershed. Biology 2023, 12, 1315. https://doi.org/10.3390/biology12101315
Liu F, Zi F, Wang X, Zeng H, Huo B, Wang C, Ge J, Chen S, Wang B. Assessing the Impact of Anthropic Pressures on Aquatic Macroinvertebrates: A Functional Trait Approach in the Irtysh River Watershed. Biology. 2023; 12(10):1315. https://doi.org/10.3390/biology12101315
Chicago/Turabian StyleLiu, Fei, Fangze Zi, Xinyue Wang, Honghui Zeng, Bin Huo, Chengxin Wang, Jianmin Ge, Shengao Chen, and Baoqiang Wang. 2023. "Assessing the Impact of Anthropic Pressures on Aquatic Macroinvertebrates: A Functional Trait Approach in the Irtysh River Watershed" Biology 12, no. 10: 1315. https://doi.org/10.3390/biology12101315
APA StyleLiu, F., Zi, F., Wang, X., Zeng, H., Huo, B., Wang, C., Ge, J., Chen, S., & Wang, B. (2023). Assessing the Impact of Anthropic Pressures on Aquatic Macroinvertebrates: A Functional Trait Approach in the Irtysh River Watershed. Biology, 12(10), 1315. https://doi.org/10.3390/biology12101315