
Biogeographic Analysis Suggests Two Types of Planktonic Prokaryote Communities in the Barents Sea

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
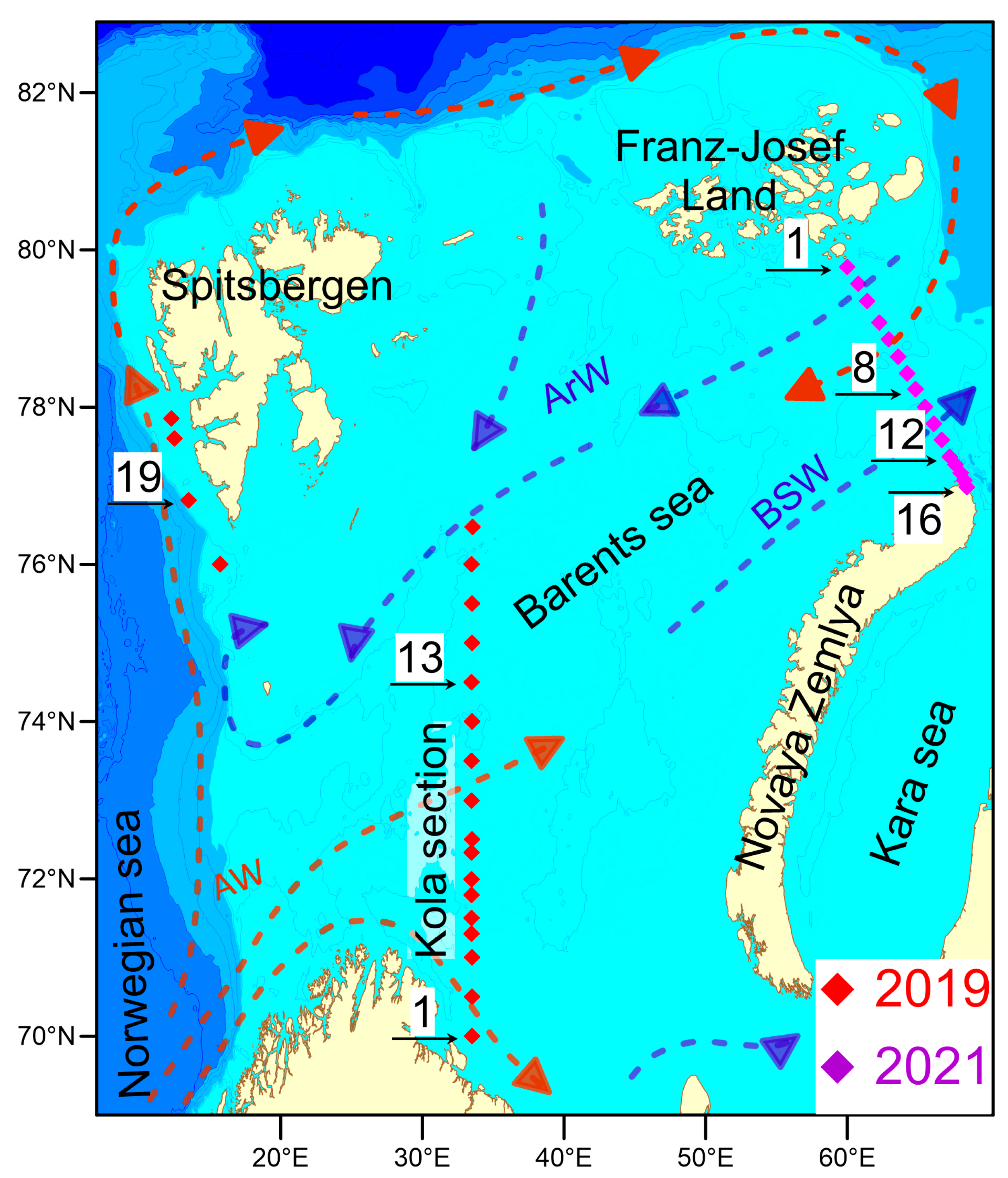
2. Materials and Methods
3. Results
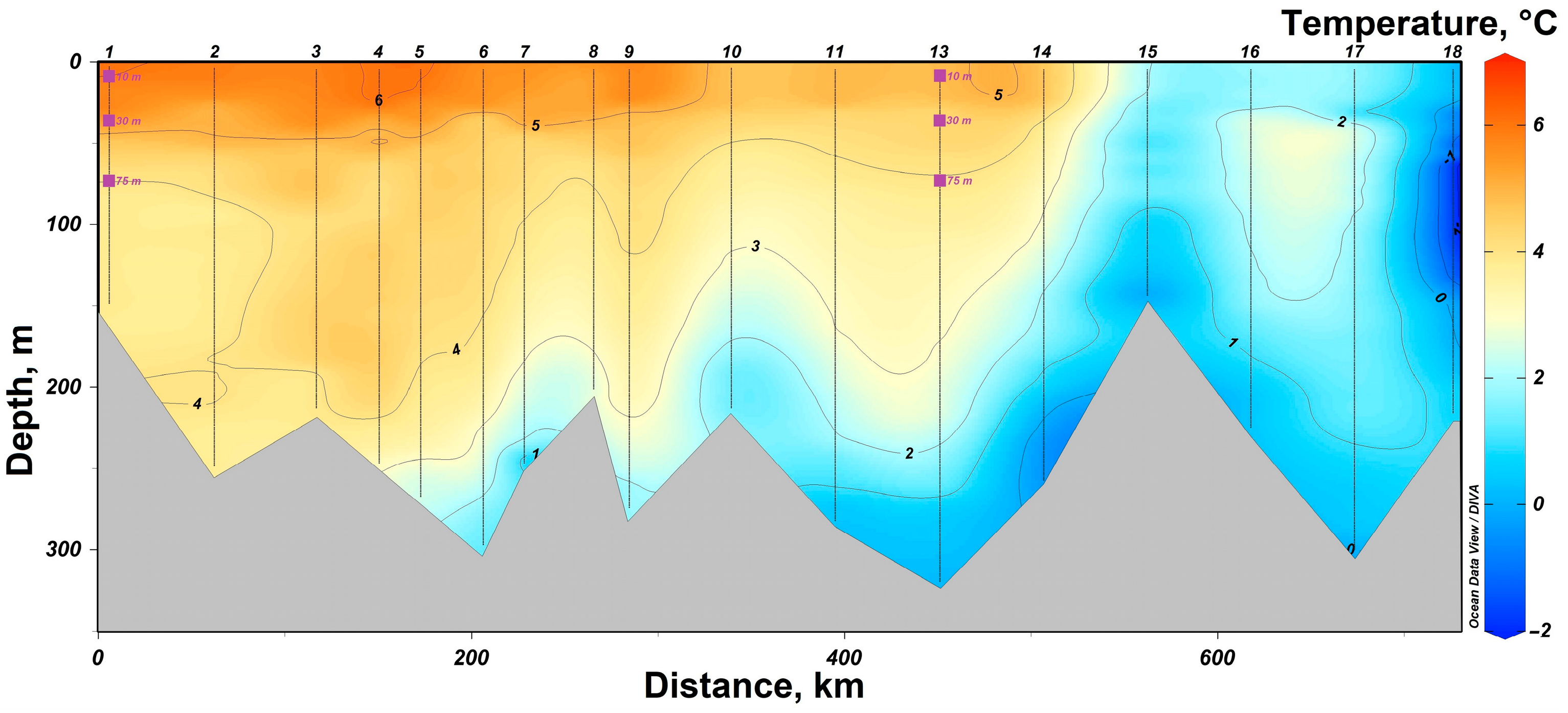
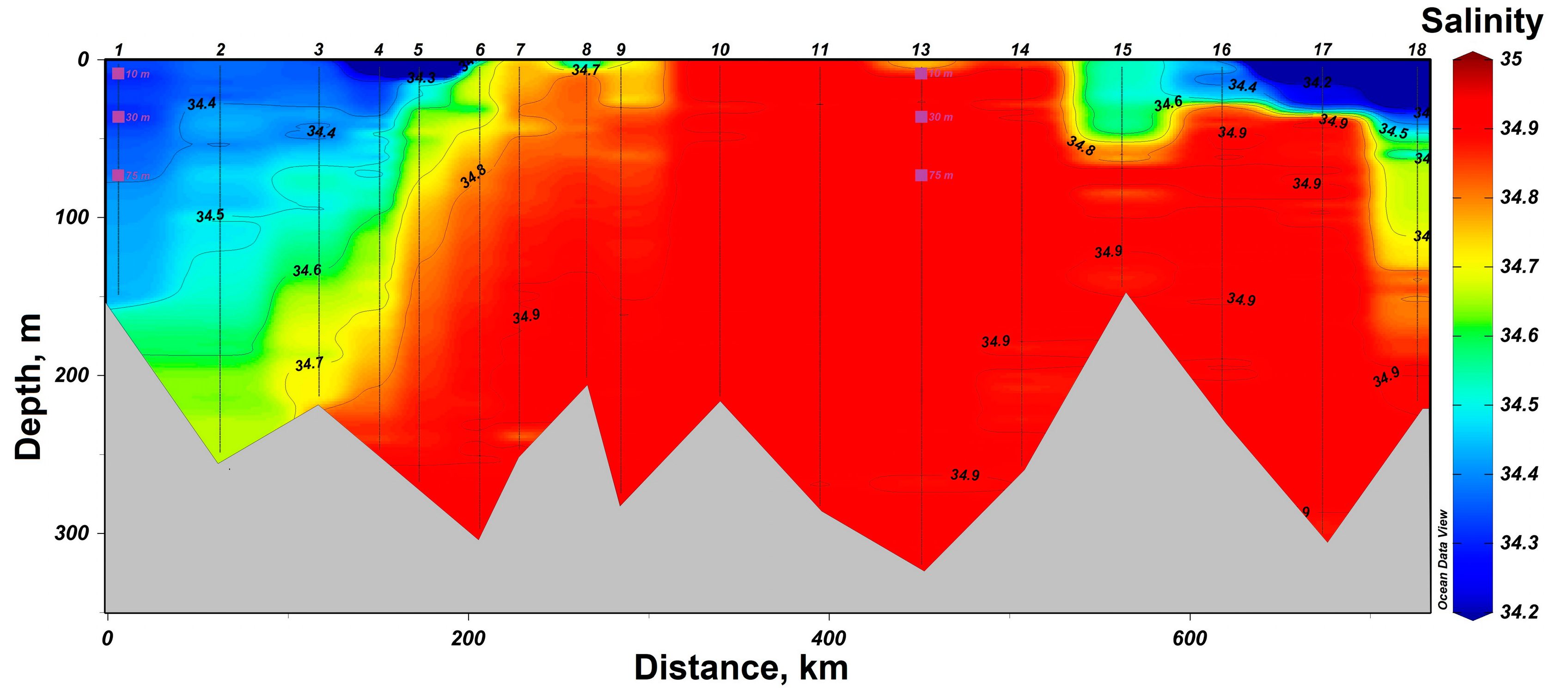
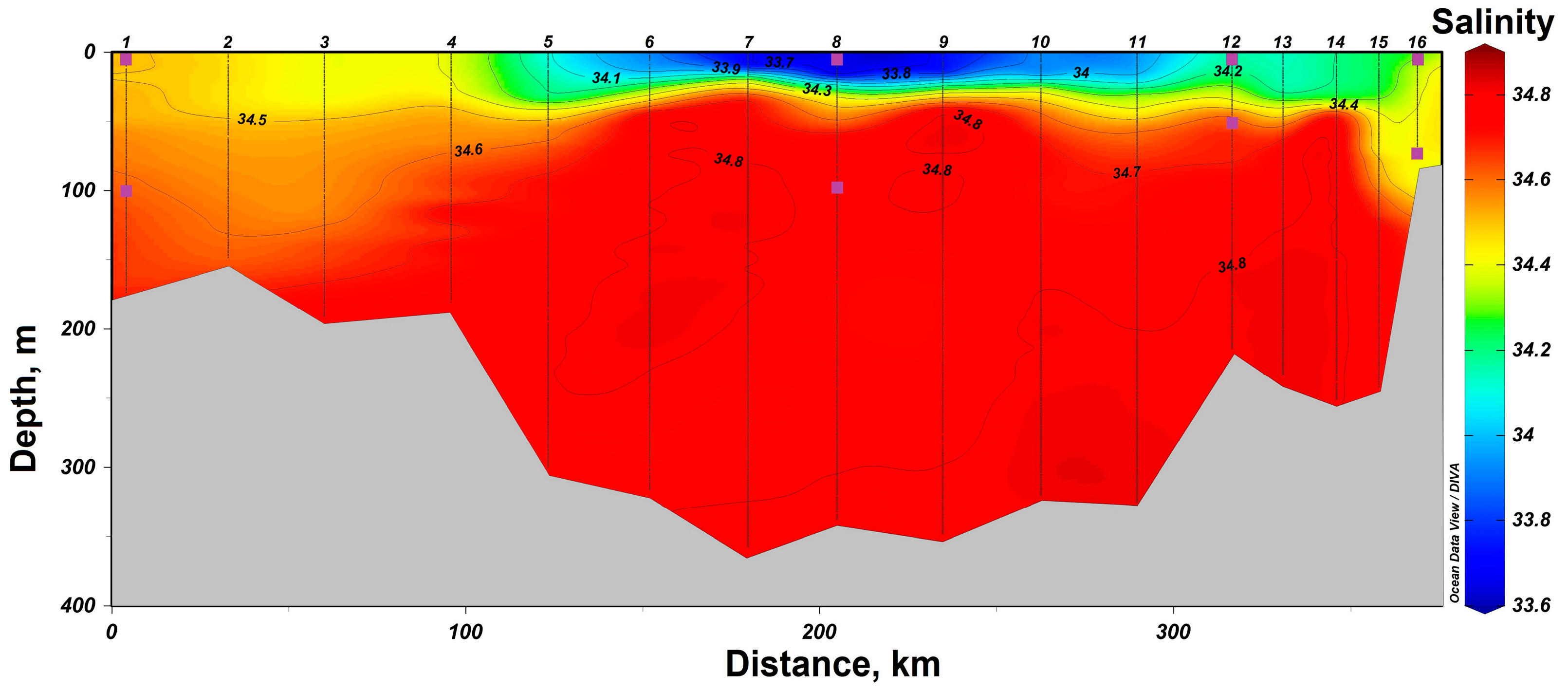
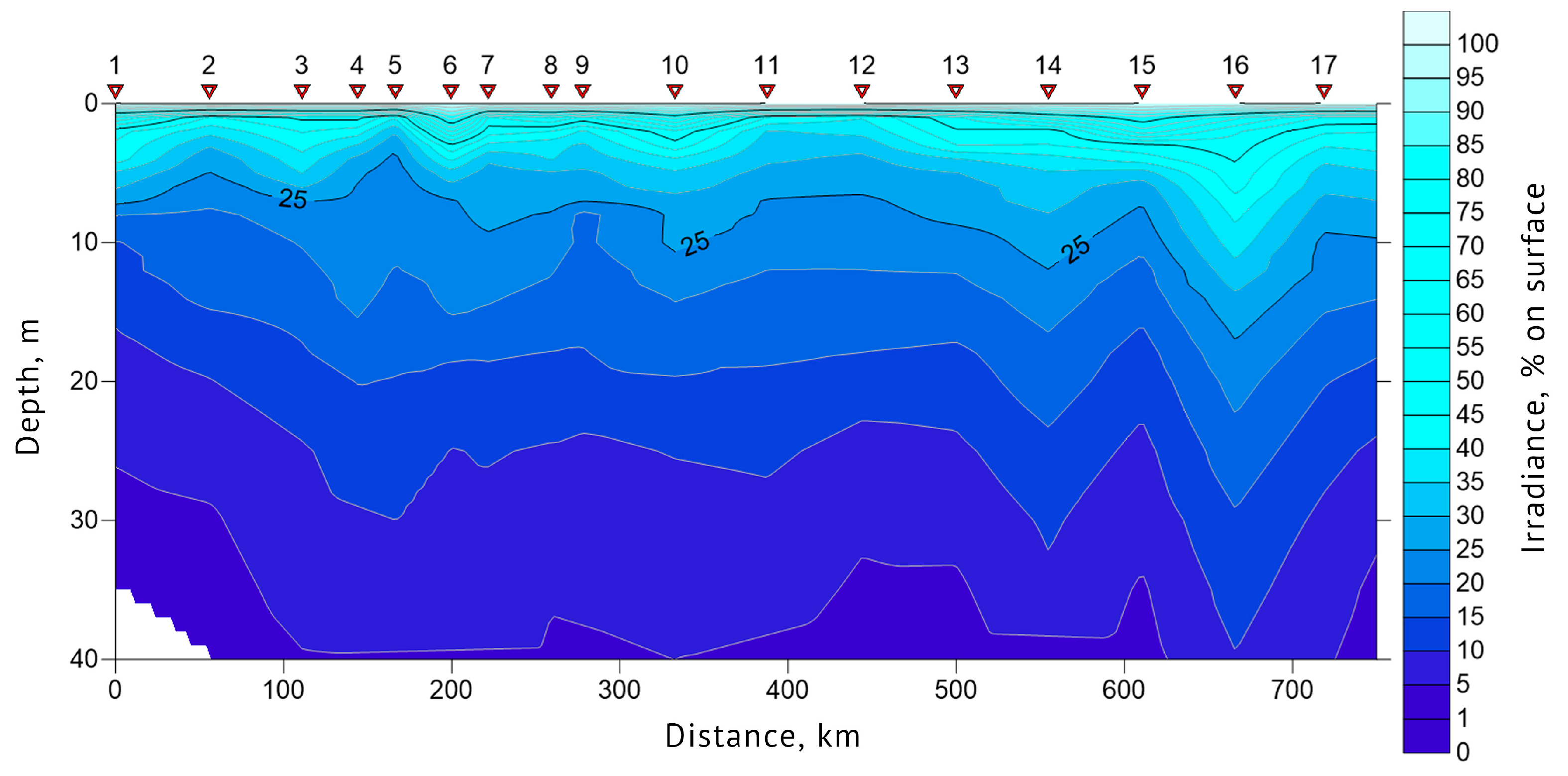
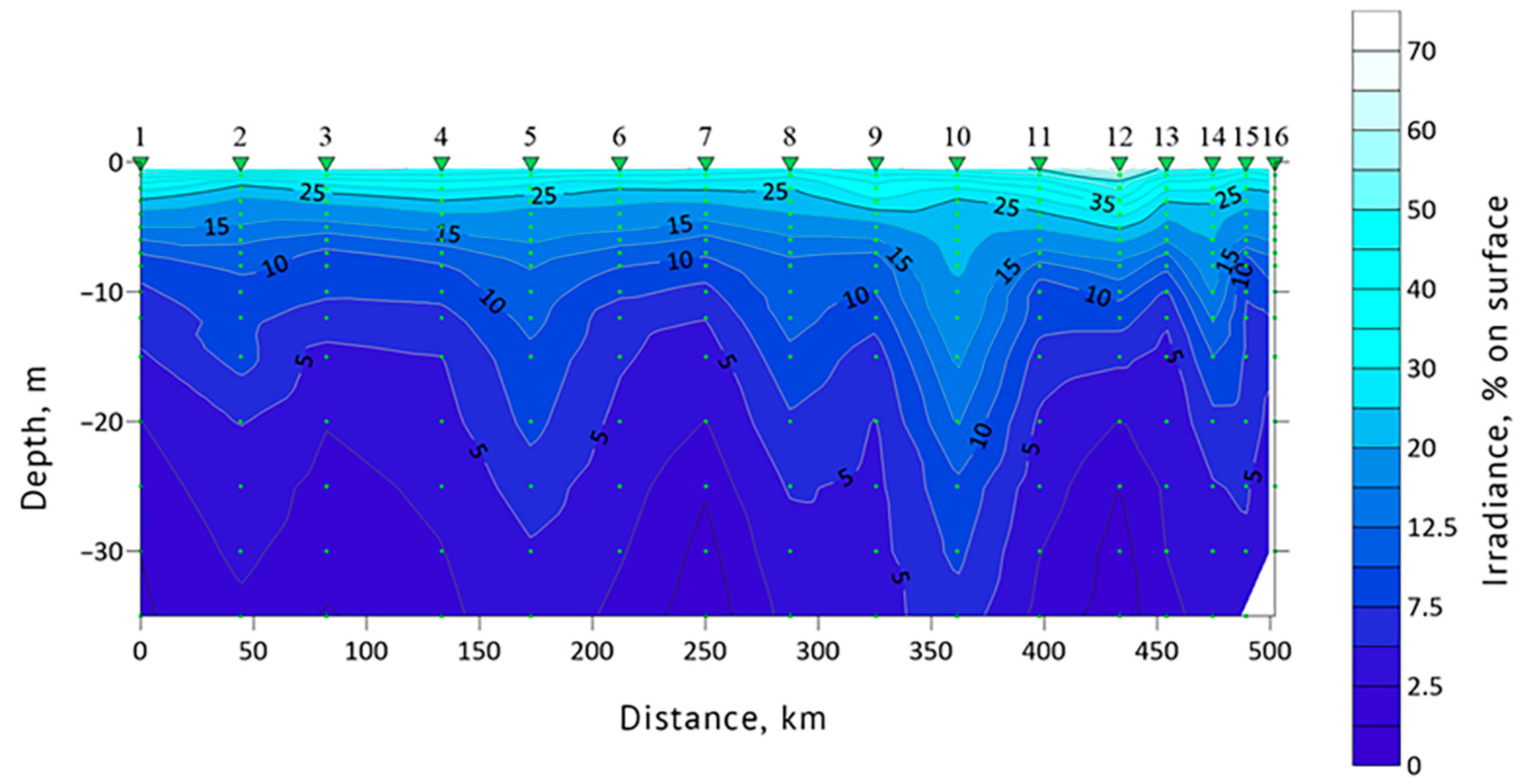
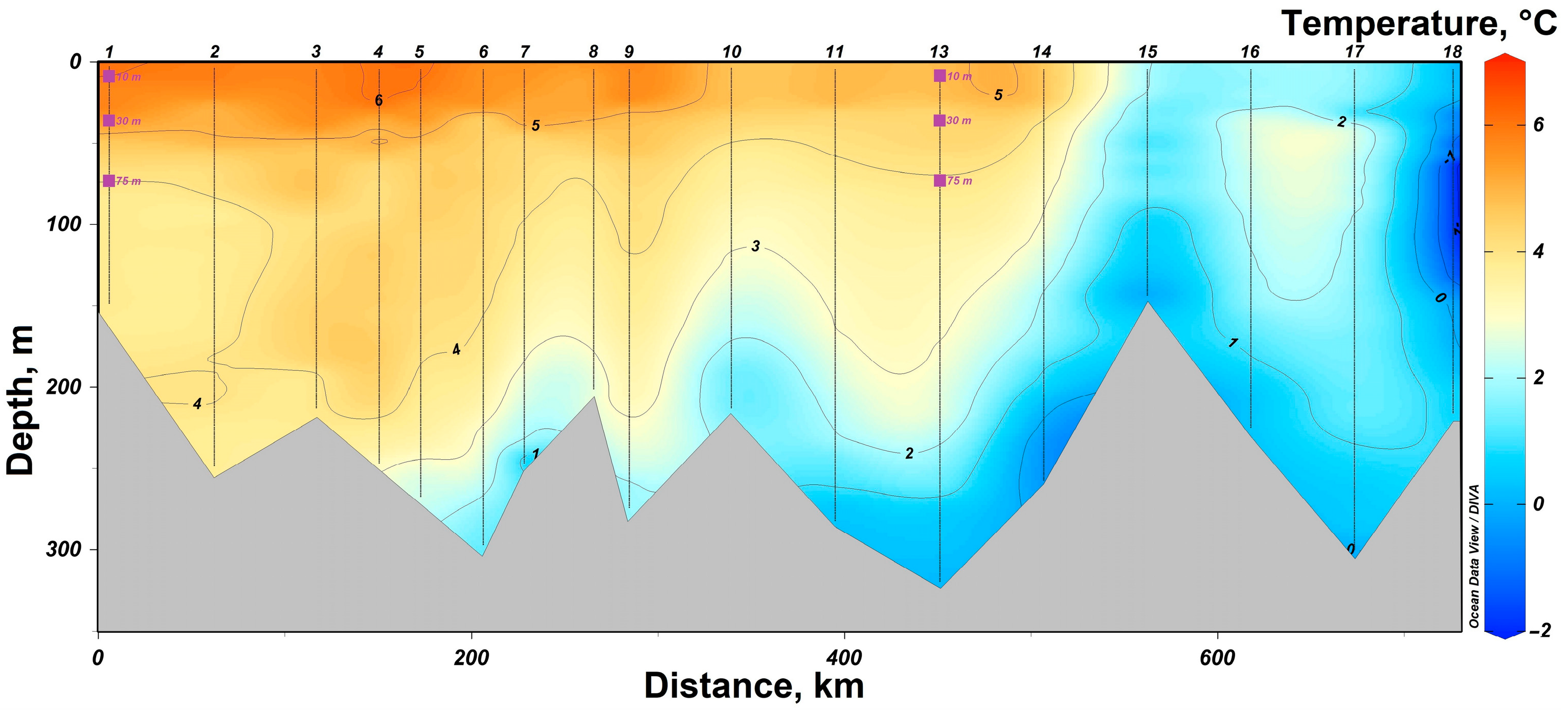
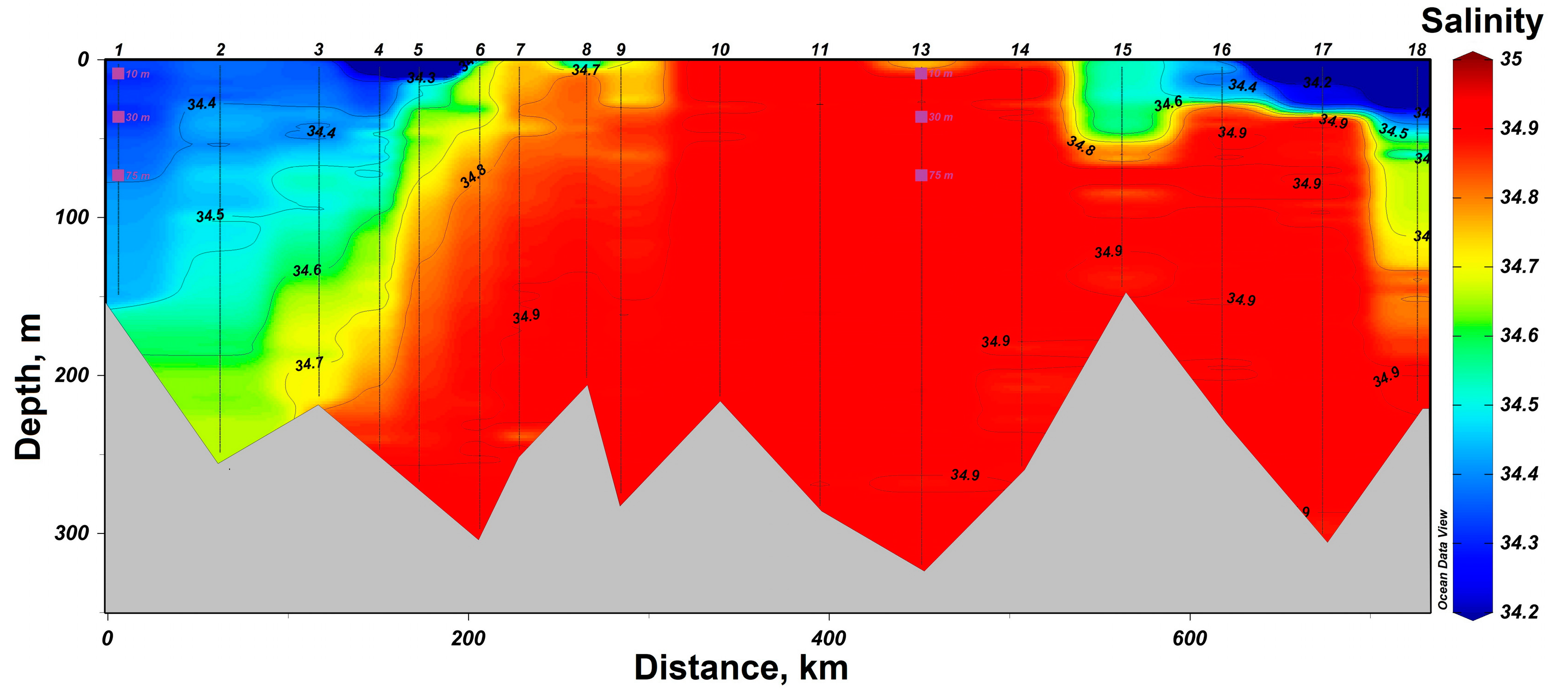
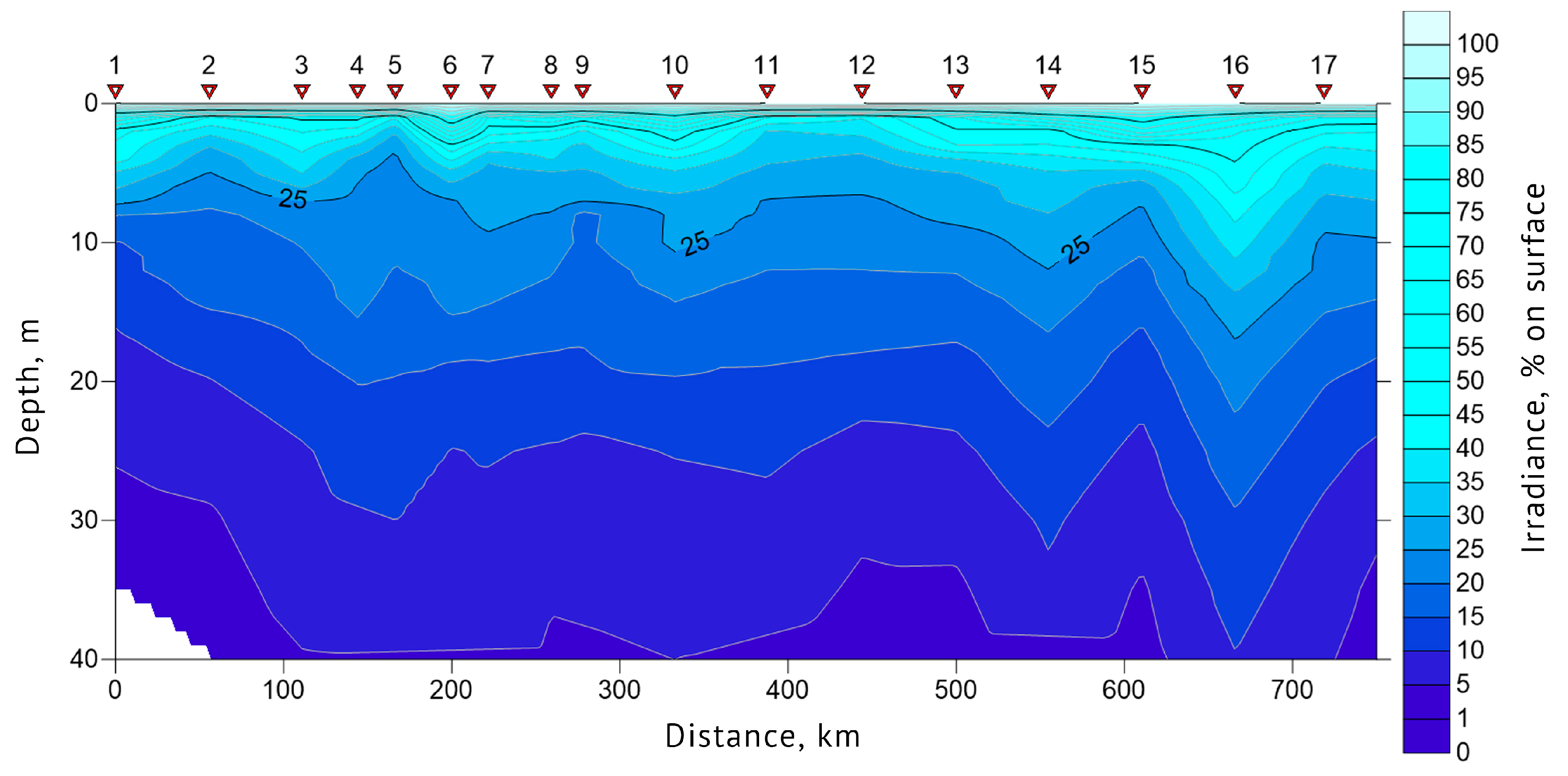
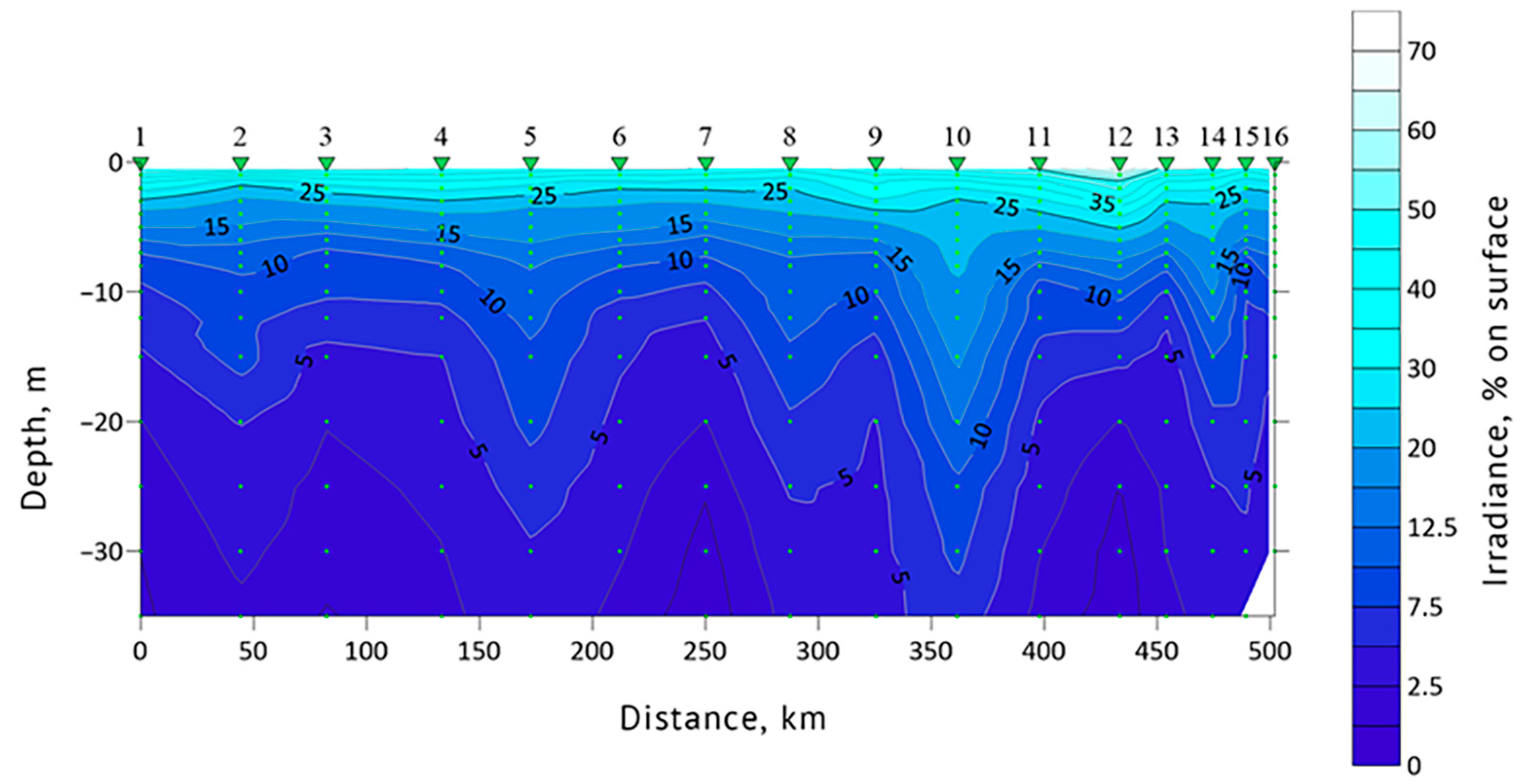
3.1. Determination of the Seawater Parameters
3.2. Analysis of Prokaryote Diversity Using 16S Metabarcoding
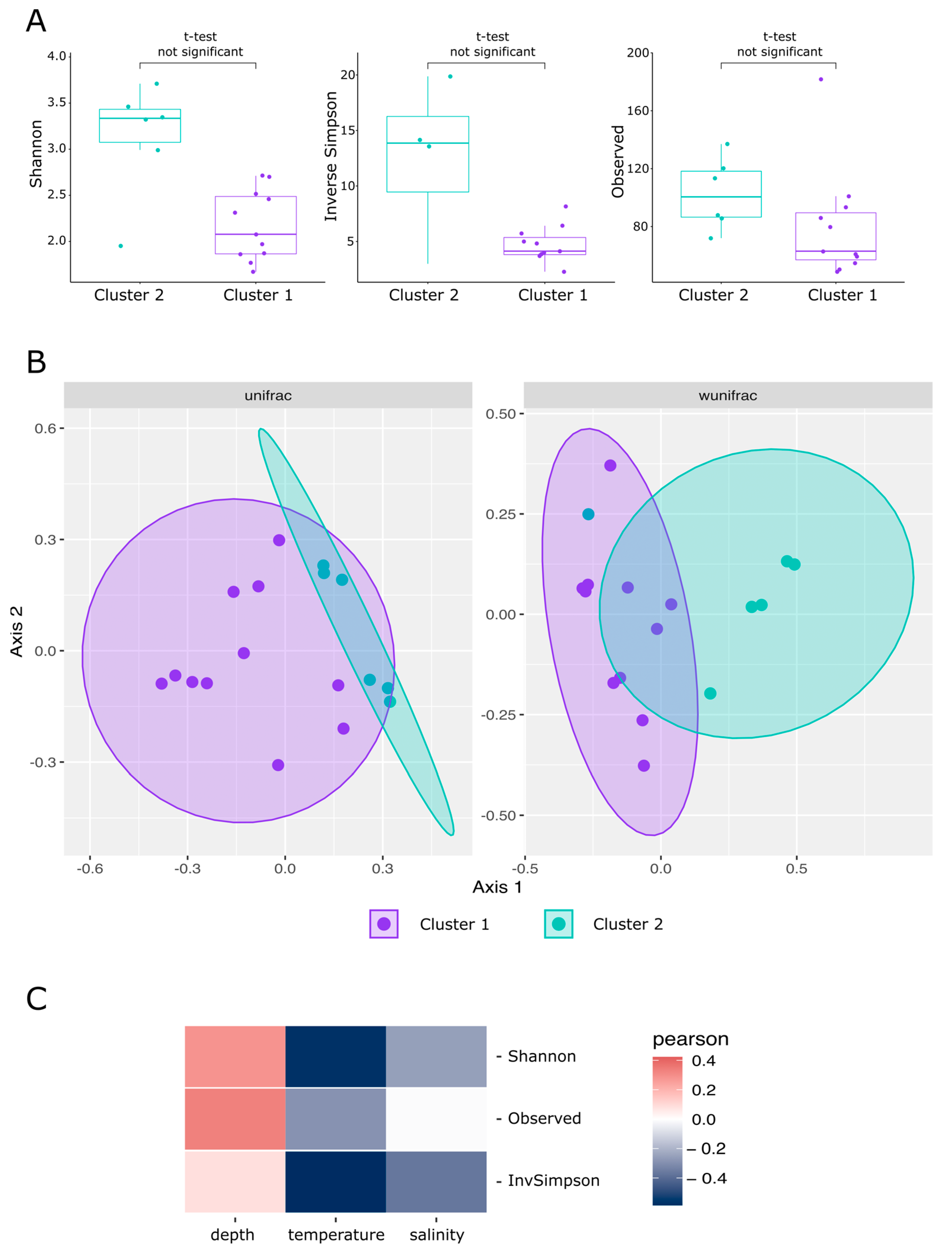
3.2.1. Hypothesis-Independent Clusterization of Samples by Community Type
3.2.2. Beta-Diversity
3.2.3. Alpha-Diversity
3.2.4. Prokaryote Community Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ingvaldsen, R.B.; Assmann, K.M.; Primicerio, R.; Fossheim, M.; Polyakov, I.V.; Dolgov, A.V. Physical Manifestations and Ecological Implications of Arctic Atlantification. Nat. Rev. Earth Environ. 2021, 2, 874–889. [Google Scholar] [CrossRef]
- Park, H.; Sherstiukov, A.B.; Fedorov, A.N.; Polyakov, I.V.; Walsh, J.E. An Observation-Based Assessment of the Influences of Air Temperature and Snow Depth on Soil Temperature in Russia. Environ. Res. Lett. 2014, 9, 064026. [Google Scholar] [CrossRef]
- Polyakov, I.V.; Pnyushkov, A.V.; Alkire, M.B.; Ashik, I.M.; Baumann, T.M.; Carmack, E.C.; Goszczko, I.; Guthrie, J.; Ivanov, V.V.; Kanzow, T.; et al. Greater Role for Atlantic Inflows on Sea-Ice Loss in the Eurasian Basin of the Arctic Ocean. Science 2017, 356, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, V.V.; Repina, I.A. Mid-Winter Anomaly of Sea Ice in the Western Nansen Basin in 2010s. IOP Conf. Ser. Earth Environ. Sci. 2019, 231, 012024. [Google Scholar] [CrossRef]
- Tesi, T.; Muschitiello, F.; Mollenhauer, G.; Miserocchi, S.; Langone, L.; Ceccarelli, C.; Panieri, G.; Chiggiato, J.; Nogarotto, A.; Hefter, J.; et al. Rapid Atlantification along the Fram Strait at the Beginning of the 20th Century. Sci. Adv. 2021, 7, eabj2946. [Google Scholar] [CrossRef]
- Rigor, I.G.; Wallace, J.M. Variations in the Age of Arctic Sea-Ice and Summer Sea-Ice Extent. Geophys. Res. Lett. 2004, 31, L09401. [Google Scholar] [CrossRef]
- Kohnemann, S.H.E.; Heinemann, G.; Bromwich, D.H.; Gutjahr, O. Extreme Warming in the Kara Sea and Barents Sea during the Winter Period 2000–16. J. Clim. 2017, 30, 8913–8927. [Google Scholar] [CrossRef]
- Dahlke, S.; Maturilli, M. Contribution of Atmospheric Advection to the Amplified Winter Warming in the Arctic North Atlantic Region. Adv. Meteorol. 2017, 2017, e4928620. [Google Scholar] [CrossRef]
- Lind, S.; Ingvaldsen, R.B.; Furevik, T. Arctic Warming Hotspot in the Northern Barents Sea Linked to Declining Sea-Ice Import. Nat. Clim. Chang. 2018, 8, 634–639. [Google Scholar] [CrossRef]
- Schlichtholz, P. Subsurface Ocean Flywheel of Coupled Climate Variability in the Barents Sea Hotspot of Global Warming. Sci. Rep. 2019, 9, 13692. [Google Scholar] [CrossRef]
- Ivanov, V.; Smirnov, A.; Alexeev, V.; Koldunov, N.V.; Repina, I.; Semenov, V. Contribution of Convection-Induced Heat Flux to Winter Ice Decay in the Western Nansen Basin. J. Geophys. Res. Ocean. 2018, 123, 6581–6597. [Google Scholar] [CrossRef]
- Onarheim, I.H.; Eldevik, T.; Smedsrud, L.H.; Stroeve, J.C. Seasonal and Regional Manifestation of Arctic Sea Ice Loss. J. Clim. 2018, 31, 4917–4932. [Google Scholar] [CrossRef]
- Carter-Gates, M.; Balestreri, C.; Thorpe, S.E.; Cottier, F.; Baylay, A.; Bibby, T.S.; Moore, C.M.; Schroeder, D.C. Implications of Increasing Atlantic Influence for Arctic Microbial Community Structure. Sci. Rep. 2020, 10, 19262. [Google Scholar] [CrossRef] [PubMed]
- Fadeev, E.; Rogge, A.; Ramondenc, S.; Nöthig, E.-M.; Wekerle, C.; Bienhold, C.; Salter, I.; Waite, A.M.; Hehemann, L.; Boetius, A.; et al. Sea Ice Presence Is Linked to Higher Carbon Export and Vertical Microbial Connectivity in the Eurasian Arctic Ocean. Commun. Biol. 2021, 4, 1255. [Google Scholar] [CrossRef]
- Lewis, K.M.; van Dijken, G.L.; Arrigo, K.R. Changes in Phytoplankton Concentration Now Drive Increased Arctic Ocean Primary Production. Science 2020, 369, 198–202. [Google Scholar] [CrossRef]
- Comeau, A.M.; Li, W.K.W.; Tremblay, J.-É.; Carmack, E.C.; Lovejoy, C. Arctic Ocean Microbial Community Structure before and after the 2007 Record Sea Ice Minimum. PLoS ONE 2011, 6, e27492. [Google Scholar] [CrossRef]
- Fossheim, M.; Primicerio, R.; Johannesen, E.; Ingvaldsen, R.B.; Aschan, M.M.; Dolgov, A.V. Recent Warming Leads to a Rapid Borealization of Fish Communities in the Arctic. Nat. Clim Chang. 2015, 5, 673–677. [Google Scholar] [CrossRef]
- Descamps, S.; Aars, J.; Fuglei, E.; Kovacs, K.M.; Lydersen, C.; Pavlova, O.; Pedersen, Å.Ø.; Ravolainen, V.; Strøm, H. Climate Change Impacts on Wildlife in a High Arctic Archipelago—Svalbard, Norway. Glob. Chang. Biol. 2017, 23, 490–502. [Google Scholar] [CrossRef]
- Namsaraev, Z.; Melnikova, A.; Komova, A.; Ivanov, V.; Rudenko, A.; Ivanov, E. Algal Bloom Occurrence and Effects in Russia. Water 2020, 12, 285. [Google Scholar] [CrossRef]
- Namsaraev, Z.; Bobrik, A.; Kozlova, A.; Krylova, A.; Rudenko, A.; Mitina, A.; Saburov, A.; Patrushev, M.; Karnachuk, O.; Toshchakov, S. Carbon Emission and Biodiversity of Arctic Soil Microbial Communities of the Novaya Zemlya and Franz Josef Land Archipelagos. Microorganisms 2023, 11, 482. [Google Scholar] [CrossRef]
- Melnikova, A.A.; Komova, A.V.; Namsaraev, Z.B. Trends and Driving Forces of Cyanobacterial Blooms in Russia in the 20th and Early 21st Centuries. Microbiology 2022, 91, 649–661. [Google Scholar] [CrossRef]
- Wietz, M.; Bienhold, C.; Metfies, K.; Torres-Valdés, S.; von Appen, W.-J.; Salter, I.; Boetius, A. The Polar Night Shift: Seasonal Dynamics and Drivers of Arctic Ocean Microbiomes Revealed by Autonomous Sampling. ISME Commun. 2021, 1, 76. [Google Scholar] [CrossRef]
- Priest, T.; von Appen, W.-J.; Oldenburg, E.; Popa, O.; Torres-Valdés, S.; Bienhold, C.; Metfies, K.; Boulton, W.; Mock, T.; Fuchs, B.M.; et al. Atlantic Water Influx and Sea-Ice Cover Drive Taxonomic and Functional Shifts in Arctic Marine Bacterial Communities. ISME J. 2023, 17, 1612–1625. [Google Scholar] [CrossRef] [PubMed]
- Galand, P.E.; Potvin, M.; Casamayor, E.O.; Lovejoy, C. Hydrography Shapes Bacterial Biogeography of the Deep Arctic Ocean. ISME J. 2010, 4, 564–576. [Google Scholar] [CrossRef]
- Bowman, J.S.; Rasmussen, S.; Blom, N.; Deming, J.W.; Rysgaard, S.; Sicheritz-Ponten, T. Microbial Community Structure of Arctic Multiyear Sea Ice and Surface Seawater by 454 Sequencing of the 16S RNA Gene. ISME J. 2012, 6, 11–20. [Google Scholar] [CrossRef]
- Ahme, A.; Von Jackowski, A.; McPherson, R.A.; Wolf, K.K.E.; Hoppmann, M.; Neuhaus, S.; John, U. Winners and Losers of Atlantification: The Degree of Ocean Warming Affects the Structure of Arctic Microbial Communities. Genes 2023, 14, 623. [Google Scholar] [CrossRef]
- Aalto, N.J.; Schweitzer, H.D.; Krsmanovic, S.; Campbell, K.; Bernstein, H.C. Diversity and Selection of Surface Marine Microbiomes in the Atlantic-Influenced Arctic. Front. Microbiol. 2022, 13, 892634. [Google Scholar] [CrossRef]
- Thiele, S.; Vader, A.; Thomson, S.; Saubrekka, K.; Petelenz, E.; Müller, O.; Bratbak, G.; Øvreås, L. Seasonality of the Bacterial and Archaeal Community Composition of the Northern Barents Sea. Front. Microbiol. 2023, 14, 1213718. [Google Scholar] [CrossRef]
- Thiele, S.; Vader, A.; Thomson, S.; Saubrekka, K.; Petelenz, E.; Armo, H.R.; Müller, O.; Olsen, L.; Bratbak, G.; Øvreås, L. The Summer Bacterial and Archaeal Community Composition of the Northern Barents Sea. Prog. Oceanogr. 2023, 215, 103054. [Google Scholar] [CrossRef]
- Thiele, S.; Storesund, J.E.; Fernández-Méndez, M.; Assmy, P.; Øvreås, L. A Winter-to-Summer Transition of Bacterial and Archaeal Communities in Arctic Sea Ice. Microorganisms 2022, 10, 1618. [Google Scholar] [CrossRef]
- Šupraha, L.; Klemm, K.; Gran-Stadniczeñko, S.; Hörstmann, C.; Vaulot, D.; Edvardsen, B.; John, U. Diversity and Biogeography of Planktonic Diatoms in Svalbard Fjords: The Role of Dispersal and Arctic Endemism in Phytoplankton Community Structuring. Elem. Sci. Anthr. 2022, 10, 00117. [Google Scholar] [CrossRef]
- Joo, H.M.; Kim, K.E.; Park, J.S.; Kim, H.-J.; Yang, E.J.; Lee, T.-K.; Cho, K.-H.; Jung, J.; Lee, Y.; Kang, S.-H.; et al. Ecological Responses of Core Phytoplankton by Latitudinal Differences in the Arctic Ocean in Late Summer Revealed by 18S rDNA Metabarcoding. Front. Mar. Sci. 2022, 9, 879911. [Google Scholar] [CrossRef]
- Kraemer, S.; Ramachandran, A.; Colatriano, D.; Lovejoy, C.; Walsh, D.A. Diversity and Biogeography of SAR11 Bacteria from the Arctic Ocean. ISME J. 2020, 14, 79–90. [Google Scholar] [CrossRef]
- Timmermans, M.-L.; Marshall, J. Understanding Arctic Ocean Circulation: A Review of Ocean Dynamics in a Changing Climate. J. Geophys. Res. Ocean. 2020, 125, e2018JC014378. [Google Scholar] [CrossRef]
- Quadfasel, D.; Rudels, B.; Selchow, S. The Central Bank Vortex in the Barents Sea: Water Mass Transformation and Circulation. ICES Mar. Sci. Symp. 1992, 195, 40–51. [Google Scholar]
- Ivanov, V.V.; Tuzov, F.K. Formation of Dense Water Dome over the Central Bank under Conditions of Reduced Ice Cover in the Barents Sea. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2021, 175, 103590. [Google Scholar] [CrossRef]
- Wilson, B.; Müller, O.; Nordmann, E.-L.; Seuthe, L.; Bratbak, G.; Øvreås, L. Changes in Marine Prokaryote Composition with Season and Depth Over an Arctic Polar Year. Front. Mar. Sci. 2017, 4, 95. [Google Scholar] [CrossRef]
- Boitsov, V.D.; Karsakov, A.L.; Trofimov, A.G. Atlantic Water Temperature and Climate in the Barents Sea, 2000–2009. ICES J. Mar. Sci. 2012, 69, 833–840. [Google Scholar] [CrossRef]
- Schlitzer, R. Interactive Analysis and Visualization of Geoscience Data with Ocean Data View. Comput. Geosci. 2002, 28, 1211–1218. [Google Scholar] [CrossRef]
- Lee, Z.; Weidemann, A.; Kindle, J.; Arnone, R.; Carder, K.L.; Davis, C. Euphotic Zone Depth: Its Derivation and Implication to Ocean-Color Remote Sensing. J. Geophys. Res. Ocean. 2007, 112, C03009. [Google Scholar] [CrossRef]
- Toshchakov, S.V.; Izotova, A.O.; Vinogradova, E.N.; Kachmazov, G.S.; Tuaeva, A.Y.; Abaev, V.T.; Evteeva, M.A.; Gunitseva, N.M.; Korzhenkov, A.A.; Elcheninov, A.G.; et al. Culture-Independent Survey of Thermophilic Microbial Communities of the North Caucasus. Biology 2021, 10, 1352. [Google Scholar] [CrossRef] [PubMed]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An Improved Dual-Indexing Approach for Multiplexed 16S rRNA Gene Sequencing on the Illumina MiSeq Platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Hugerth, L.W.; Wefer, H.A.; Lundin, S.; Jakobsson, H.E.; Lindberg, M.; Rodin, S.; Engstrand, L.; Andersson, A.F. DegePrime, a Program for Degenerate Primer Design for Broad-Taxonomic-Range PCR in Microbial Ecology Studies. Appl. Environ. Microbiol. 2014, 80, 5116–5123. [Google Scholar] [CrossRef] [PubMed]
- Merkel, A.Y.; Tarnovetskii, I.Y.; Podosokorskaya, O.A.; Toshchakov, S.V. Analysis of 16S rRNA Primer Systems for Profiling of Thermophilic Microbial Communities. Microbiology 2019, 88, 671–680. [Google Scholar] [CrossRef]
- Gohl, D.; MacLean, A.; Hauge, A.; Becker, A.; Walek, D.; Beckman, K.B. An Optimized Protocol for High-Throughput Amplicon-Based Microbiome Profiling. Protoc. Exch. 2016. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Renaud, G.; Stenzel, U.; Maricic, T.; Wiebe, V.; Kelso, J. deML: Robust Demultiplexing of Illumina Sequences Using a Likelihood-Based Approach. Bioinformatics 2015, 31, 770–772. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Bodenhofer, U.; Bonatesta, E.; Horejš-Kainrath, C.; Hochreiter, S. Msa: An R Package for Multiple Sequence Alignment. Bioinformatics 2015, 31, 3997–3999. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An Effective Distance Metric for Microbial Community Comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A. NbClust: An R Package for Determining the Relevant Number of Clusters in a Data Set. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar] [CrossRef]
- Rego, A.; Fernandez-Guerra, A.; Duarte, P.; Assmy, P.; Leão, P.N.; Magalhães, C. Secondary Metabolite Biosynthetic Diversity in Arctic Ocean Metagenomes. Microb. Genom. 2021, 7, 000731. [Google Scholar] [CrossRef] [PubMed]
- Malmstrom, R.R.; Straza, T.R.A.; Cottrell, M.T.; Kirchman, D.L. Diversity, Abundance, and Biomass Production of Bacterial Groups in the Western Arctic Ocean. Aquat. Microb. Ecol. 2007, 47, 45–55. [Google Scholar] [CrossRef]
- Haro-Moreno, J.M.; Rodriguez-Valera, F.; Rosselli, R.; Martinez-Hernandez, F.; Roda-Garcia, J.J.; Gomez, M.L.; Fornas, O.; Martinez-Garcia, M.; López-Pérez, M. Ecogenomics of the SAR11 Clade. Environ. Microbiol. 2020, 22, 1748–1763. [Google Scholar] [CrossRef] [PubMed]
- Rapp, J.Z.; Fernández-Méndez, M.; Bienhold, C.; Boetius, A. Effects of Ice-Algal Aggregate Export on the Connectivity of Bacterial Communities in the Central Arctic Ocean. Front. Microbiol. 2018, 9, 1035. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, A.G.G.; Tomasino, M.P.; Duarte, P.; Fernández-Méndez, M.; Assmy, P.; Ribeiro, H.; Surkont, J.; Leite, R.B.; Pereira-Leal, J.B.; Torgo, L.; et al. Diversity and Composition of Pelagic Prokaryotic and Protist Communities in a Thin Arctic Sea-Ice Regime. Microb. Ecol. 2019, 78, 388–408. [Google Scholar] [CrossRef]
- Bowman, J.P. The Genus Psychrobacter. In The Prokaryotes: A Handbook on the Biology of Bacteria Volume 6: Proteobacteria: Gamma Subclass; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 920–930. ISBN 978-0-387-30746-6. [Google Scholar]
- Moghadam, M.S.; Albersmeier, A.; Winkler, A.; Cimmino, L.; Rise, K.; Hohmann-Marriott, M.F.; Kalinowski, J.; Rückert, C.; Wentzel, A.; Lale, R. Isolation and Genome Sequencing of Four Arctic Marine Psychrobacter Strains Exhibiting Multicopper Oxidase Activity. BMC Genom. 2016, 17, 117. [Google Scholar] [CrossRef]
- Yinxin, Z.; Huirong, L.; Yong, Y.; Bo, C. Phylogenetic Relationship and Phenotypic Comparison of Psychrobacter Species Isolated from Polar Oceans. Prog. Nat. Sci. 2007, 17, 44–48. [Google Scholar] [CrossRef]
- Park, J.R.; Bae, J.-W.; Nam, Y.-D.; Chang, H.-W.; Kwon, H.-Y.; Quan, Z.-X.; Park, Y.-H. Sulfitobacter Litoralis Sp. Nov., a Marine Bacterium Isolated from the East Sea, Korea. Int. J. Syst. Evol. Microbiol. 2007, 57, 692–695. [Google Scholar] [CrossRef]
- Sorokin, D.Y. Sulfitobacter Pontiacus Gen. Nov., Sp. Nov.—A New Heterotrophic Bacterium from the Black Sea, Specialized on Sulfite Oxidation. Mikrobiologiya 1995, 64, 354–365. [Google Scholar]
- Chaffron, S.; Delage, E.; Budinich, M.; Vintache, D.; Henry, N.; Nef, C.; Ardyna, M.; Zayed, A.A.; Junger, P.C.; Galand, P.E.; et al. Environmental Vulnerability of the Global Ocean Epipelagic Plankton Community Interactome. Sci. Adv. 2021, 7, eabg1921. [Google Scholar] [CrossRef] [PubMed]
- Avcı, B.; Krüger, K.; Fuchs, B.M.; Teeling, H.; Amann, R.I. Polysaccharide Niche Partitioning of Distinct Polaribacter Clades during North Sea Spring Algal Blooms. ISME J. 2020, 14, 1369–1383. [Google Scholar] [CrossRef] [PubMed]
- González, J.M.; Fernández-Gómez, B.; Fernàndez-Guerra, A.; Gómez-Consarnau, L.; Sánchez, O.; Coll-Lladó, M.; del Campo, J.; Escudero, L.; Rodríguez-Martínez, R.; Alonso-Sáez, L.; et al. Genome Analysis of the Proteorhodopsin-Containing Marine Bacterium Polaribacter Sp. MED152 (Flavobacteria). Proc. Natl. Acad. Sci. USA 2008, 105, 8724–8729. [Google Scholar] [CrossRef]
- Celussi, M.; Balestra, C.; Fabbro, C.; Crevatin, E.; Cataletto, B.; Fonda Umani, S.; Del Negro, P. Organic-Matter Degradative Potential of Halomonas Glaciei Isolated from Frazil Ice in the Ross Sea (Antarctica). FEMS Microbiol. Ecol. 2008, 65, 504–512. [Google Scholar] [CrossRef]
- Williamson, A.; De Santi, C.; Altermark, B.; Karlsen, C.; Hjerde, E. Complete Genome Sequence of Halomonas Sp. R5-57. Stand. Genom. Sci. 2016, 11, 62. [Google Scholar] [CrossRef]
- Sinha, R.K.; Krishnan, K.P.; Hatha, A.A.M.; Rahiman, M.; Thresyamma, D.D.; Kerkar, S. Diversity of Retrievable Heterotrophic Bacteria in Kongsfjorden, an Arctic Fjord. Braz. J. Microbiol. 2017, 48, 51–61. [Google Scholar] [CrossRef]
- Gorrasi, S.; Pesciaroli, C.; Barghini, P.; Pasqualetti, M.; Fenice, M. Structure and Diversity of the Bacterial Community of an Arctic Estuarine System (Kandalaksha Bay) Subject to Intense Tidal Currents. J. Mar. Syst. 2019, 196, 77–85. [Google Scholar] [CrossRef]
- Médigue, C.; Krin, E.; Pascal, G.; Barbe, V.; Bernsel, A.; Bertin, P.N.; Cheung, F.; Cruveiller, S.; D’Amico, S.; Duilio, A.; et al. Coping with Cold: The Genome of the Versatile Marine Antarctica Bacterium Pseudoalteromonas Haloplanktis TAC125. Genome Res. 2005, 15, 1325–1335. [Google Scholar] [CrossRef]
- Zheng, K.; Dong, Y.; Liang, Y.; Liu, Y.; Zhang, X.; Zhang, W.; Wang, Z.; Shao, H.; Sung, Y.Y.; Mok, W.J.; et al. Genomic Diversity and Ecological Distribution of Marine Pseudoalteromonas Phages. Mar. Life Sci. Technol. 2023, 5, 271–285. [Google Scholar] [CrossRef]
- Yu, Y.; Li, H.; Zeng, Y.; Chen, B. Extracellular Enzymes of Cold-Adapted Bacteria from Arctic Sea Ice, Canada Basin. Polar Biol. 2009, 32, 1539–1547. [Google Scholar] [CrossRef]
- Wietz, M.; Gram, L.; Jørgensen, B.; Schramm, A. Latitudinal Patterns in the Abundance of Major Marine Bacterioplankton Groups. Aquat. Microb. Ecol. 2010, 61, 179–189. [Google Scholar] [CrossRef]
- López-Pérez, M.; Rodriguez-Valera, F. The Family Alteromonadaceae. In The Prokaryotes: Gammaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 69–92. ISBN 978-3-642-38922-1. [Google Scholar]
- Francis, B.; Urich, T.; Mikolasch, A.; Teeling, H.; Amann, R. North Sea Spring Bloom-Associated Gammaproteobacteria Fill Diverse Heterotrophic Niches. Environ. Microb. 2021, 16, 15. [Google Scholar] [CrossRef]
- Xue, C.; Xie, Z.-X.; Li, Y.-Y.; Chen, X.-H.; Sun, G.; Lin, L.; Giovannoni, S.J.; Wang, D.-Z. Polysaccharide Utilization by a Marine Heterotrophic Bacterium from the SAR92 Clade. FEMS Microbiol. Ecol. 2021, 97, fiab120. [Google Scholar] [CrossRef] [PubMed]
- Carini, P.; Steindler, L.; Beszteri, S.; Giovannoni, S.J. Nutrient Requirements for Growth of the Extreme Oligotroph ‘Candidatus Pelagibacter Ubique’ HTCC1062 on a Defined Medium. ISME J. 2013, 7, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.M.; Rappé, M.S.; Connon, S.A.; Vergin, K.L.; Siebold, W.A.; Carlson, C.A.; Giovannoni, S.J. SAR11 Clade Dominates Ocean Surface Bacterioplankton Communities. Nature 2002, 420, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Pernthaler, J. Roseobacter and SAR11 Dominate Microbial Glucose Uptake in Coastal North Sea Waters. Environ. Microbiol. 2006, 8, 2022–2030. [Google Scholar] [CrossRef]
- Liu, Y.; Debeljak, P.; Rembauville, M.; Blain, S.; Obernosterer, I. Diatoms Shape the Biogeography of Heterotrophic Prokaryotes in Early Spring in the Southern Ocean. Environ. Microbiol. 2019, 21, 1452–1465. [Google Scholar] [CrossRef]
- Milici, M.; Vital, M.; Tomasch, J.; Badewien, T.H.; Giebel, H.-A.; Plumeier, I.; Wang, H.; Pieper, D.H.; Wagner-Döbler, I.; Simon, M. Diversity and Community Composition of Particle-Associated and Free-Living Bacteria in Mesopelagic and Bathypelagic Southern Ocean Water Masses: Evidence of Dispersal Limitation in the Bransfield Strait. Limnol. Oceanogr. 2017, 62, 1080–1095. [Google Scholar] [CrossRef]
- Bowman, J.P. The Family Cryomorphaceae. In The Prokaryotes: Other Major Lineages of Bacteria and The Archaea; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 539–550. ISBN 978-3-642-38954-2. [Google Scholar]
- Morris, R.M.; Spietz, R.L. The Physiology and Biogeochemistry of SUP05. Annu. Rev. Mar. Sci. 2022, 14, 261–275. [Google Scholar] [CrossRef]
- Marshall, K.T.; Morris, R.M. Isolation of an Aerobic Sulfur Oxidizer from the SUP05/Arctic96BD-19 Clade. ISME J. 2013, 7, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Ozhigin, V.K.; Ivshin, V.A.; Trofimov, A.G.; Karsakov, A.L.; Antsiferov, M.Y. The Barents Sea Water: Structure, Circulation, Variability; Polar Research Institute of Marine Fisheries and Oceanography: Murmansk, Russia, 2016; ISBN 978-5-86349-215-5. [Google Scholar]
- Dobrovolsky, A.D.; Zalogin, B.S. Seas of USSR; Moscow University Press: Moscow, USSR, 1982. [Google Scholar]
- Zalogin, B.; Kosarev, A. The Seas; Mysl: Moscow, Russia, 1999; ISBN 5-244-00624-X. [Google Scholar]
- Salazar, G.; Cornejo-Castillo, F.M.; Benítez-Barrios, V.; Fraile-Nuez, E.; Álvarez-Salgado, X.A.; Duarte, C.M.; Gasol, J.M.; Acinas, S.G. Global Diversity and Biogeography of Deep-Sea Pelagic Prokaryotes. ISME J. 2016, 10, 596–608. [Google Scholar] [CrossRef] [PubMed]
- Hanson, C.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Martiny, J.B.H. Beyond Biogeographic Patterns: Processes Shaping the Microbial Landscape. Nat. Rev. Microbiol. 2012, 10, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Gaisin, V.A.; Grouzdev, D.S.; Namsaraev, Z.B.; Sukhacheva, M.V.; Gorlenko, V.M.; Kuznetsov, B.B. Biogeography of Thermophilic Phototrophic Bacteria Belonging to Roseiflexus Genus. FEMS Microbiol. Ecol. 2016, 92, fiw012. [Google Scholar] [CrossRef]
- De Wit, R.; Bouvier, T. ‘Everything Is Everywhere, but, the Environment Selects’; What Did Baas Becking and Beijerinck Really Say? Environ. Microbiol. 2006, 8, 755–758. [Google Scholar] [CrossRef]
- Richter, D.J.; Watteaux, R.; Vannier, T.; Leconte, J.; Frémont, P.; Reygondeau, G.; Maillet, N.; Henry, N.; Benoit, G.; Da Silva, O.; et al. Genomic Evidence for Global Ocean Plankton Biogeography Shaped by Large-Scale Current Systems. eLife 2022, 11, e78129. [Google Scholar] [CrossRef]
- Ibarbalz, F.M.; Henry, N.; Mahé, F.; Ardyna, M.; Zingone, A.; Scalco, E.; Lovejoy, C.; Lombard, F.; Jaillon, O.; Iudicone, D.; et al. Pan-Arctic Plankton Community Structure and Its Global Connectivity. Elem. Sci. Anthr. 2023, 11, 00060. [Google Scholar] [CrossRef]
- Årthun, M.; Eldevik, T.; Smedsrud, L.H.; Skagseth, Ø.; Ingvaldsen, R.B. Quantifying the Influence of Atlantic Heat on Barents Sea Ice Variability and Retreat. J. Clim. 2012, 25, 4736–4743. [Google Scholar] [CrossRef]
- Oziel, L.; Sirven, J.; Gascard, J.-C. The Barents Sea Frontal Zones and Water Masses Variability (1980–2011). Ocean. Sci. 2016, 12, 169–184. [Google Scholar] [CrossRef]
- Karcher, M.; Kulakov, M.; Pivovarov, S.; Schauer, U.; Kauker, F.; Schlitzer, R. Atlantic Water Flow to the Kara Sea—Comparing Model Results with Observations. In Siberian River Runoff in the Kara Sea: Characterisation, Quantification, Variability and Environmental Significance; Stein, R., Fahl, K., Fütterer, D.K., Galimov, E.M., Stepanets, O.V., Eds.; Proceedings in Marine Science; Elsevier: Amsterdam, The Netherlands, 2003; pp. 47–69. [Google Scholar]
- Maslowski, W.; Marble, D.; Walczowski, W.; Schauer, U.; Clement, J.L.; Semtner, A.J. On Climatological Mass, Heat, and Salt Transports through the Barents Sea and Fram Strait from a Pan-Arctic Coupled Ice-Ocean Model Simulation. J. Geophys. Res. Ocean. 2004, 109, C03032. [Google Scholar] [CrossRef]
- Kirchman, D.L. Growth Rates of Microbes in the Oceans. Annu. Rev. Mar. Sci. 2016, 8, 285–309. [Google Scholar] [CrossRef] [PubMed]
- Kirchman, D.L.; Morán, X.A.G.; Ducklow, H. Microbial Growth in the Polar Oceans—Role of Temperature and Potential Impact of Climate Change. Nat. Rev. Microbiol. 2009, 7, 451–459. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Namsaraev, Z.; Kozlova, A.; Tuzov, F.; Krylova, A.; Izotova, A.; Makarov, I.; Bezgreshnov, A.; Melnikova, A.; Trofimova, A.; Kuzmin, D.; et al. Biogeographic Analysis Suggests Two Types of Planktonic Prokaryote Communities in the Barents Sea. Biology 2023, 12, 1310. https://doi.org/10.3390/biology12101310
Namsaraev Z, Kozlova A, Tuzov F, Krylova A, Izotova A, Makarov I, Bezgreshnov A, Melnikova A, Trofimova A, Kuzmin D, et al. Biogeographic Analysis Suggests Two Types of Planktonic Prokaryote Communities in the Barents Sea. Biology. 2023; 12(10):1310. https://doi.org/10.3390/biology12101310
Chicago/Turabian StyleNamsaraev, Zorigto, Aleksandra Kozlova, Fedor Tuzov, Anastasia Krylova, Anna Izotova, Ivan Makarov, Andrei Bezgreshnov, Anna Melnikova, Anna Trofimova, Denis Kuzmin, and et al. 2023. "Biogeographic Analysis Suggests Two Types of Planktonic Prokaryote Communities in the Barents Sea" Biology 12, no. 10: 1310. https://doi.org/10.3390/biology12101310
APA StyleNamsaraev, Z., Kozlova, A., Tuzov, F., Krylova, A., Izotova, A., Makarov, I., Bezgreshnov, A., Melnikova, A., Trofimova, A., Kuzmin, D., Patrushev, M., & Toshchakov, S. (2023). Biogeographic Analysis Suggests Two Types of Planktonic Prokaryote Communities in the Barents Sea. Biology, 12(10), 1310. https://doi.org/10.3390/biology12101310