
Contrasting Marine Microbial Communities of the Fram Strait with the First Confirmed Record of Cyanobacteria Prochlorococcus marinus in the Arctic Region

 , and
, and
Abstract
:Simple Summary
Abstract
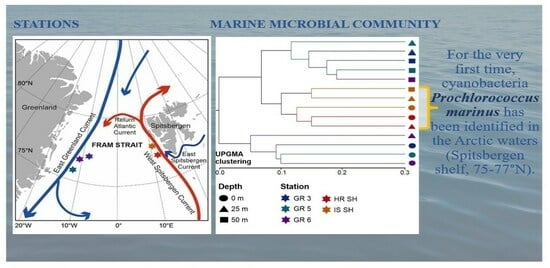
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. DNA Extraction
2.3. 16S rRNA Amplicon Library Generation and Taxonomic Classification
2.4. Analyses of the Marine Microbial Community
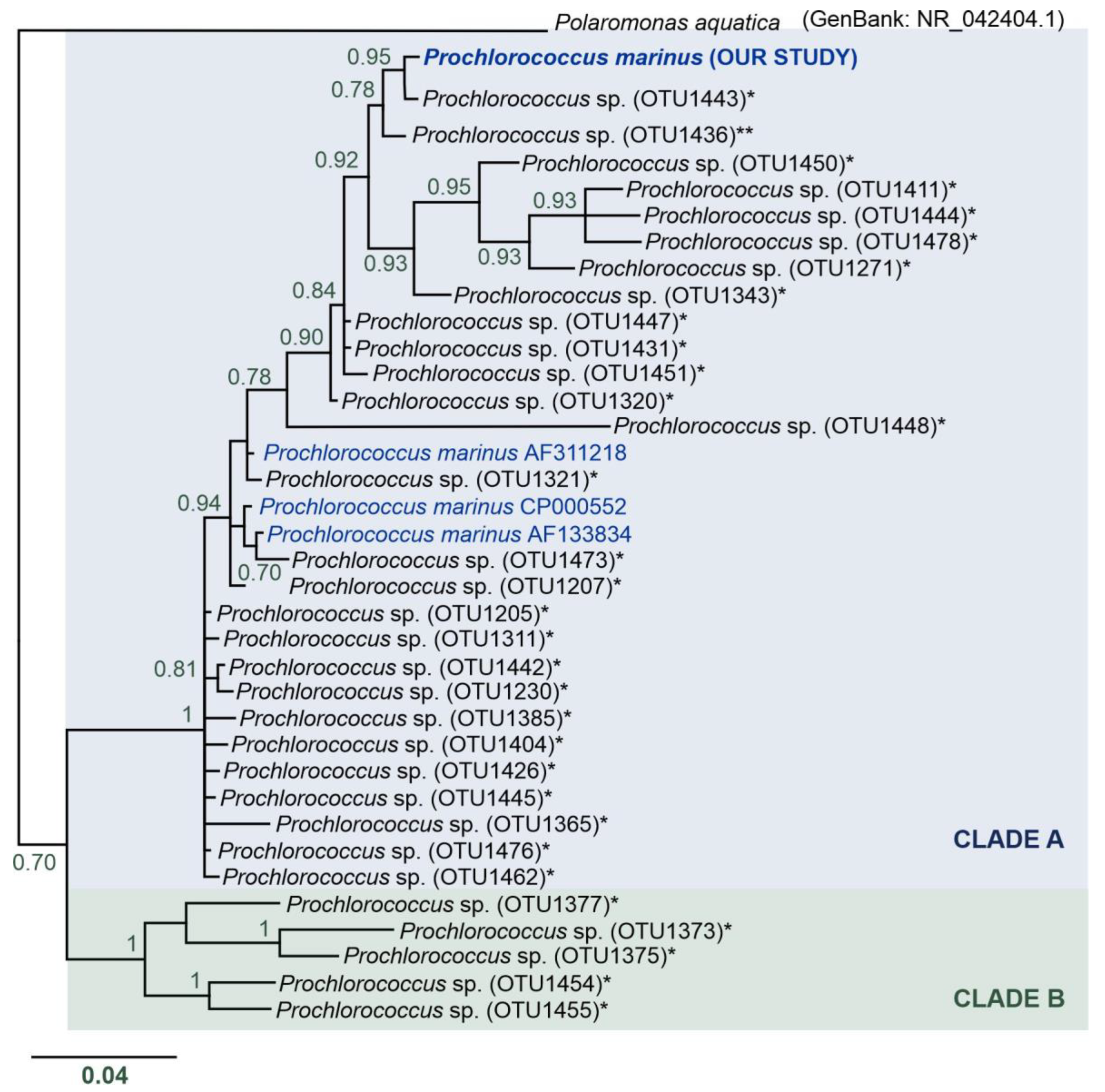
2.5. Phylogenetical Analysis of Prochlorococcus marinus OTU
3. Results
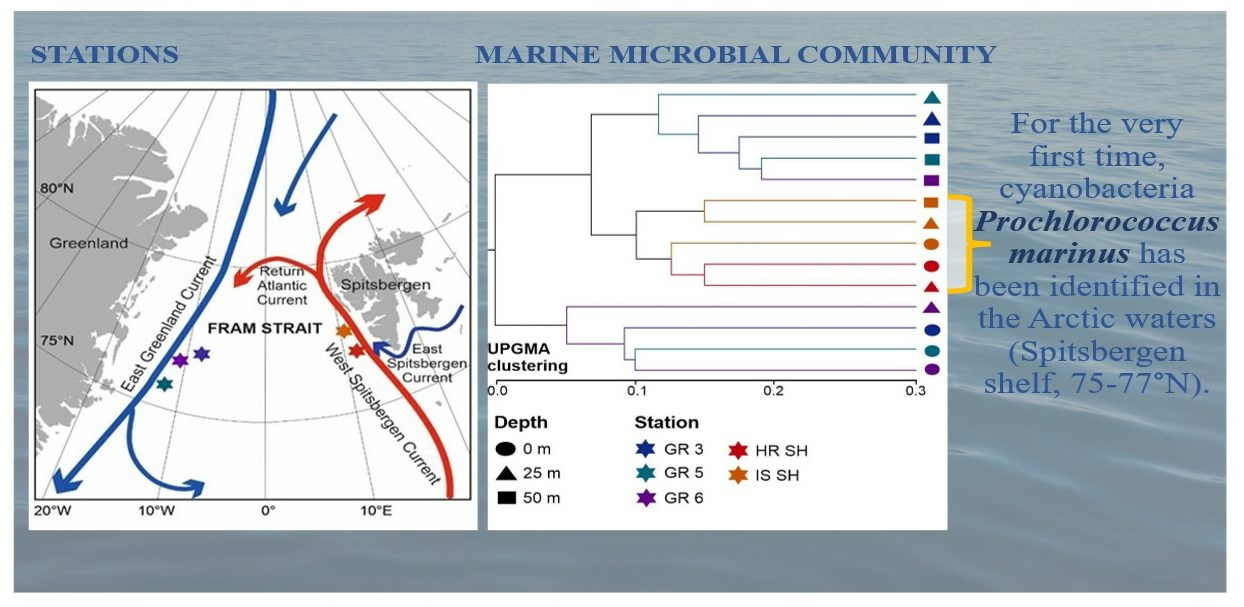
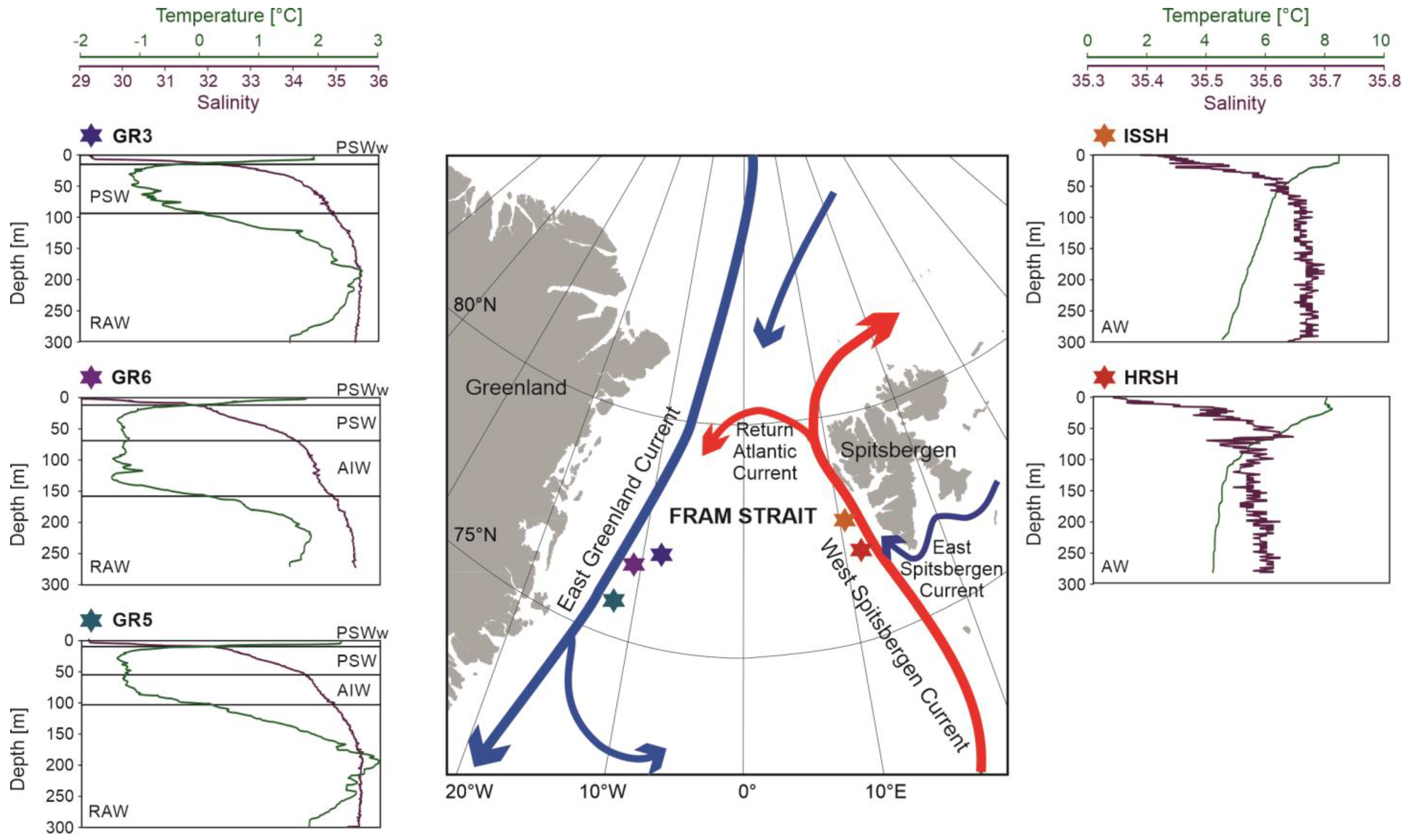
3.1. Hydrographical Conditions
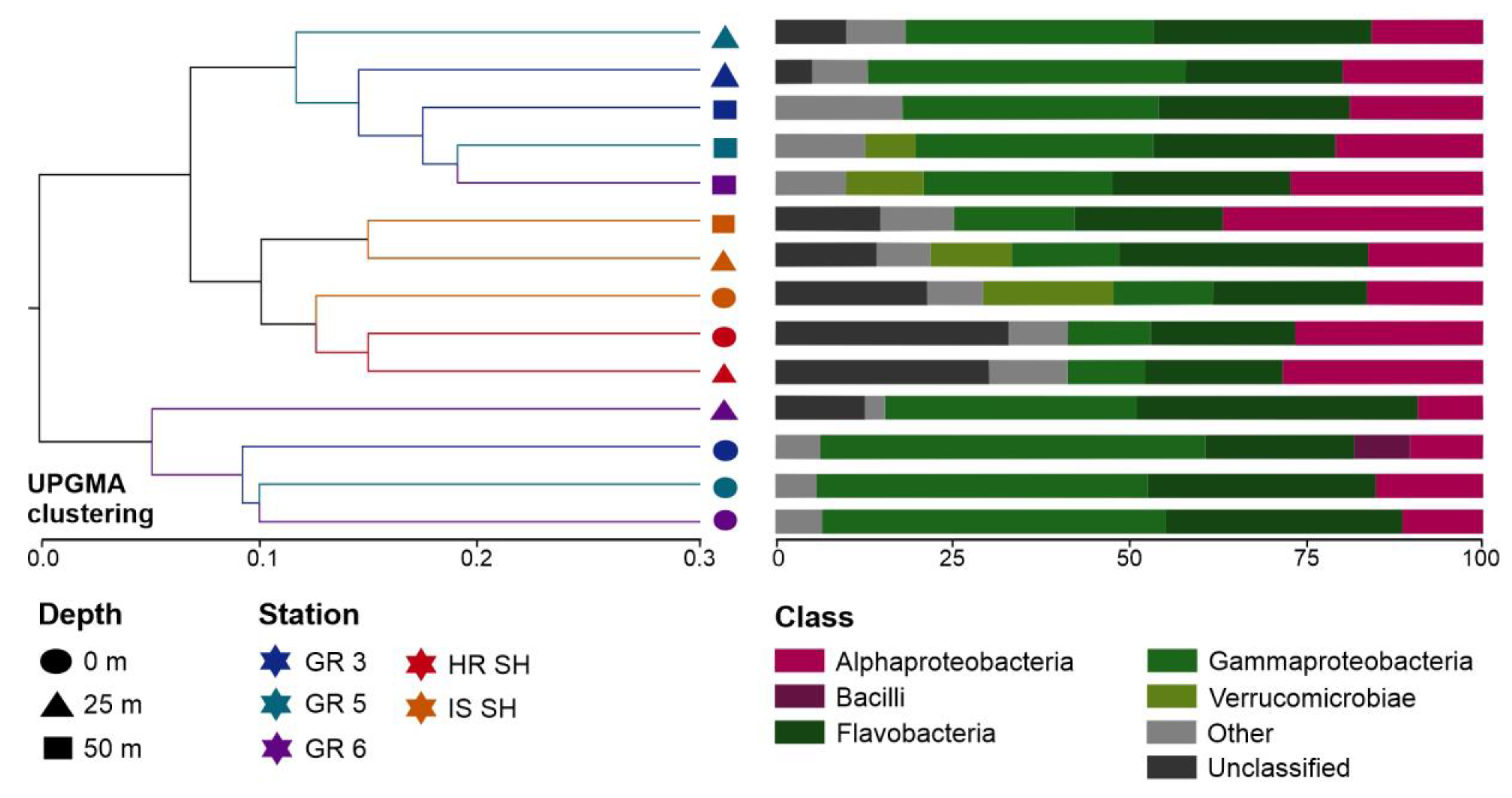
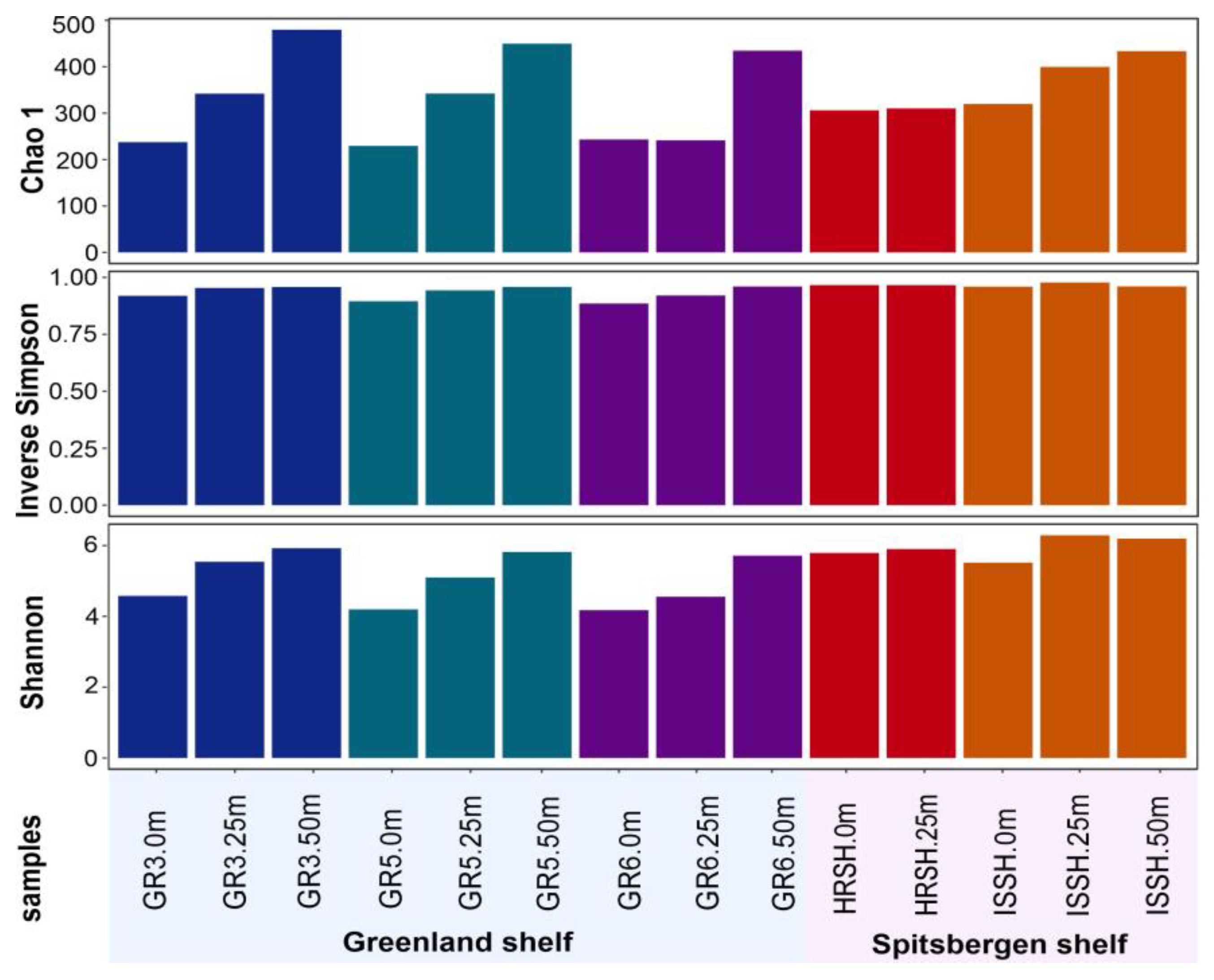
3.2. General Characteristic of 16S rRNA Gene Metagenetic Library and Marine Microbiome Diversity
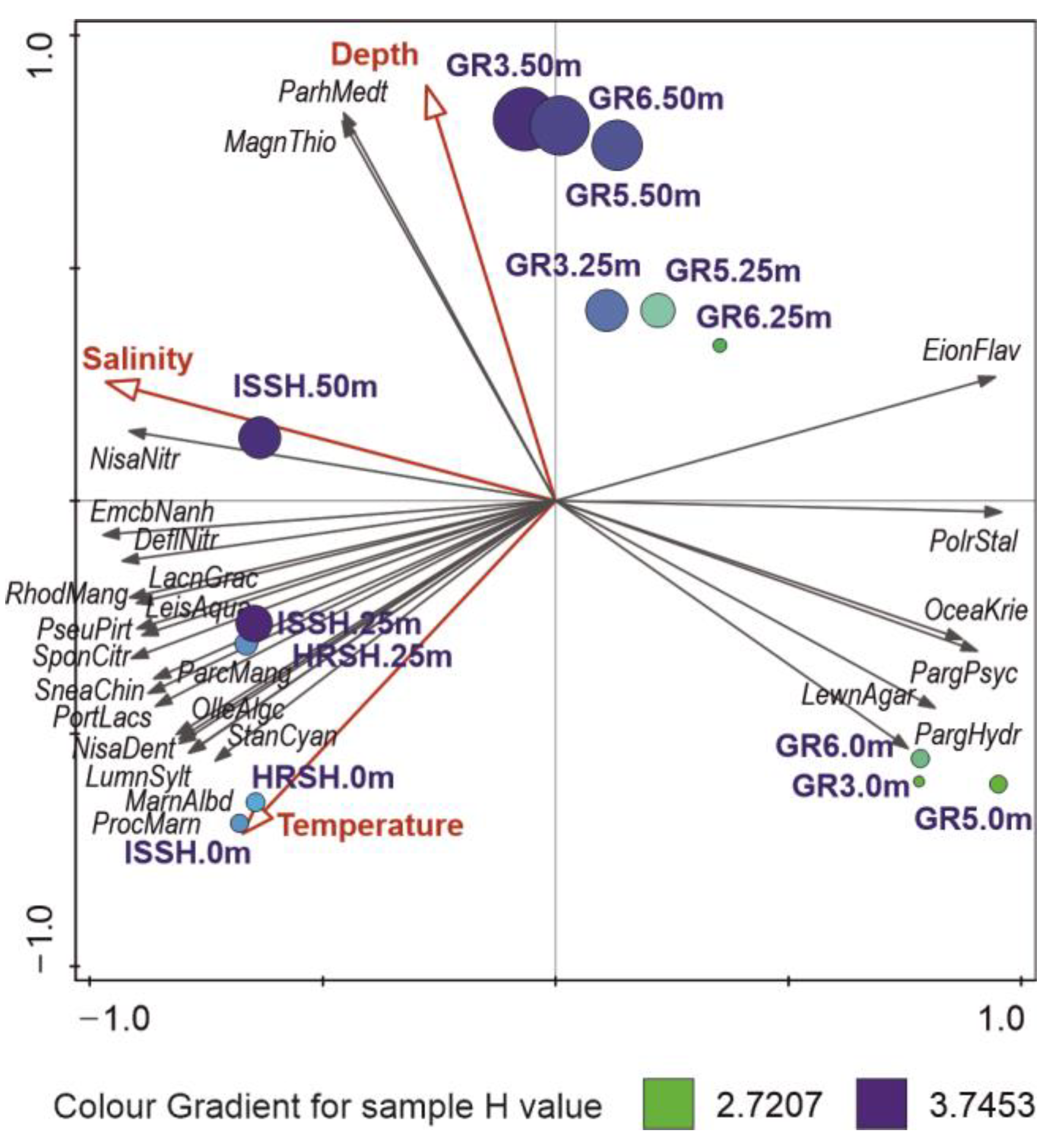
3.3. The Influence of Hydrographic Conditions on Bacterial Communities
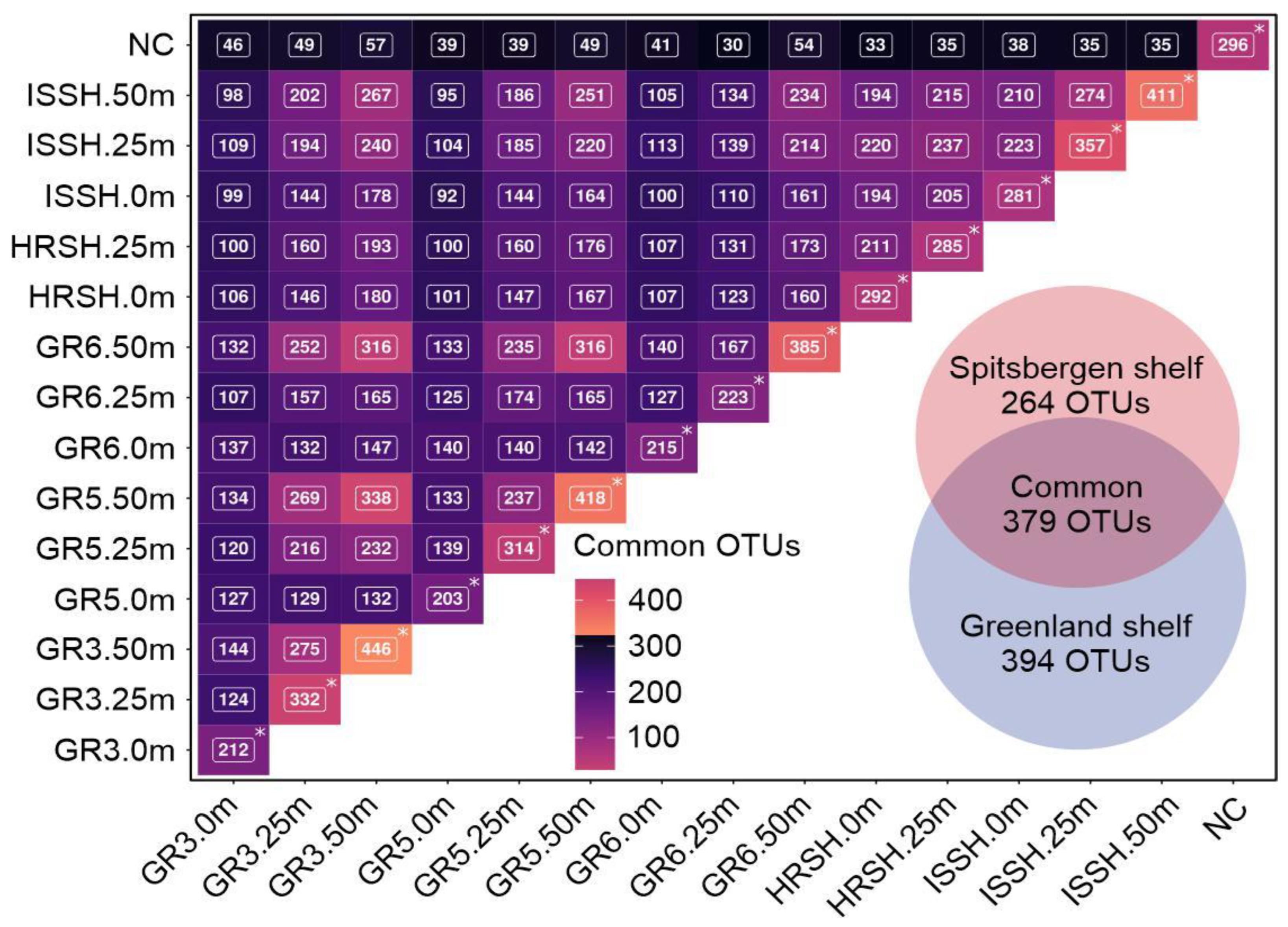
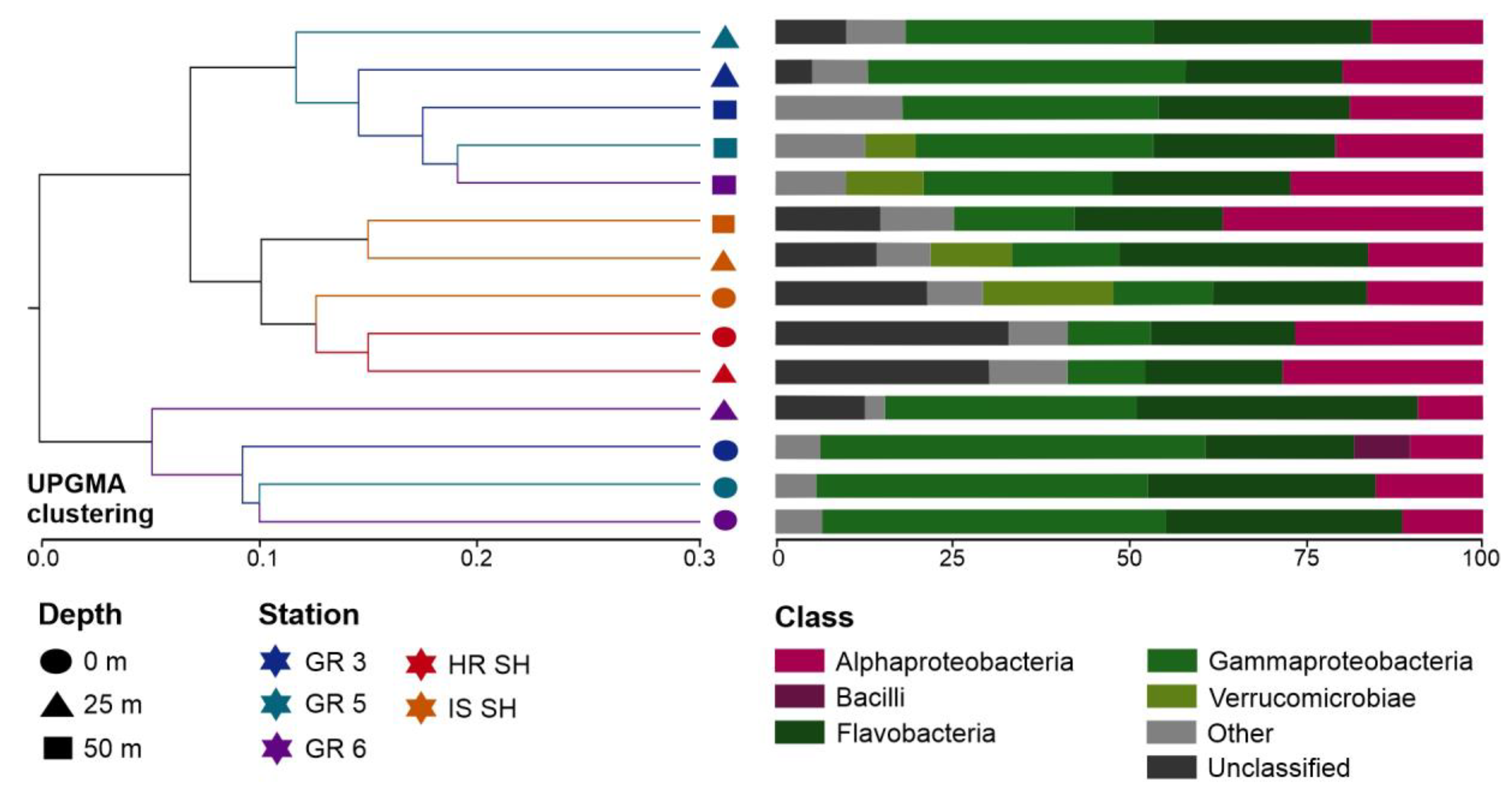
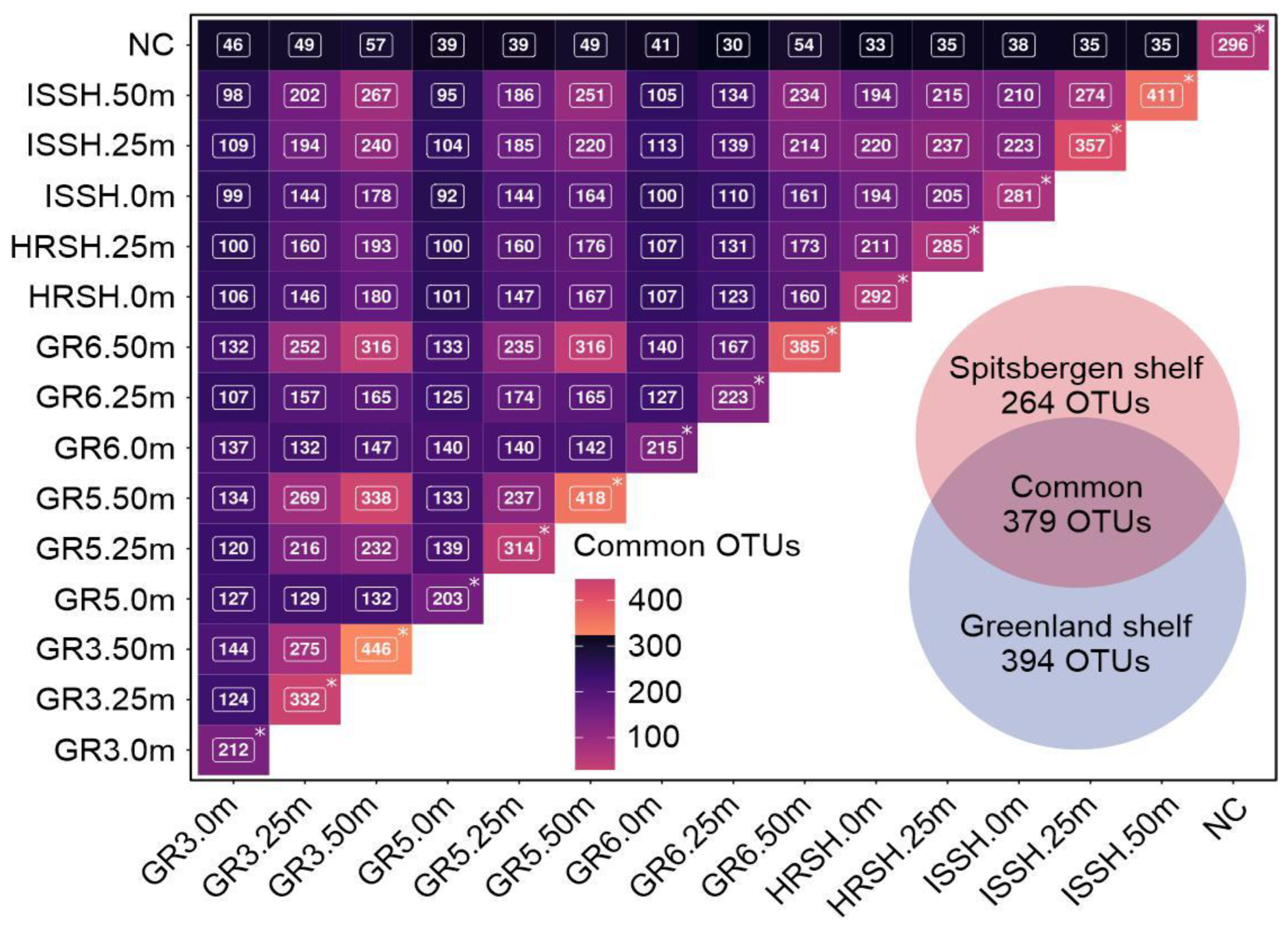
3.4. Core Marine Microbial Communities and Taxonomic Bacterial Specificity in the Greenland and Spitsbergen Shelves
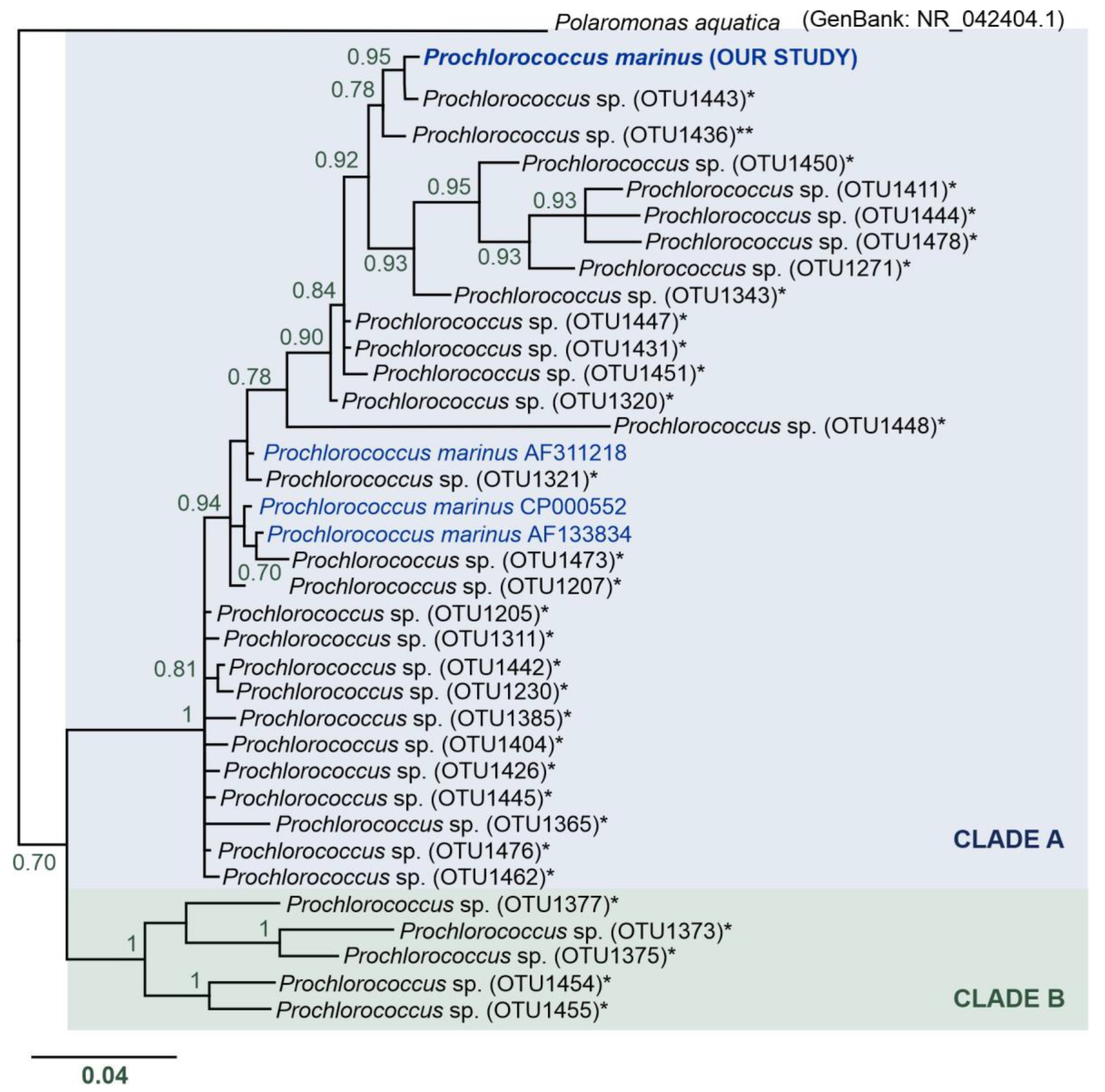
3.5. Unexpected Discovery of Prochlorococcus marinus Cyanobacteria in Arctic Waters
4. Discussion
5. Conclusions
- Contrasting marine microbial communities on the opposite sides of the Fram Strait were connected to water masses distribution and depth stratification, and salinity was the most important factor shaping these communities.
- The presence of some bacteria was restricted to one area, i.e., Dulcicalothrix necridiiformans, Lewinella xylanilytica, Olleya marilimosa, Rubritalea marina, and Vicingus serpentipes were found only on the Greenland shelf, while others, like Dulcicalothrix necridiiformans, Leisingera aquaemixtae, Luteibaculum oceani, Marinifilum albidiflavum, Olleya algicola, Phaeocystidibacter marisrubri, Portibacter lacus, Porticoccus hydrocarbonoclasticus, Pseudofulvibacter gastropodicola, Pseudohongiella nitratireducens, and Roseibacillus persicicus, were present only on the Spitsbergen shelf.
- The surface waters of the Greenland shelf, dominated by the Polar Surface Water, were characterized by the lowest number of microbial OTUs and the lowest diversity, while the deepest samples from this area showed the opposite trends. The Spitsbergen shelf could also be characterized by the number of OTUs and diversity increasing with depth, and this area, dominated by Atlantic waters with a homogenous depth stratification in terms of salinity and temperature, contained less-unique bacterial sequences.
- Higher similarity was found between the deep waters of the Greenland shelf and samples from the Spitsbergen waters than between the surface and deep waters of the Greenland shelf.
- Two main bacterial symbionts were found: Acaryochloris marina and Endozoicomonas ascidiicola; the latter was absent on the Spitsbergen shelf and in the surface waters of the Greenland shelf.
- The presence of Prochlorococcus marinus on the Spitsbergen shelf, for the very first time as far north as over 77° N, could have been connected with the process of Atlantification of the Arctic, which can have serious ecological consequences.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef]
- Cebrian, E.; Uriz, M.J.; Garrabou, J.; Ballesteros, E. Sponge mass mortalities in a warming mediterranean sea: Are cyanobacteria harboring species worse off? PLoS ONE 2011, 6, e20211. [Google Scholar] [CrossRef] [PubMed]
- Martiny, J.B.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Fenchel, T.; Finlay, B.J. The ubiquity of small species: Patterns of local and global diversity. Bioscience 2004, 54, 777–784. [Google Scholar] [CrossRef]
- Baas-Becking, L.G.M. Geobiologie of Inleiding tot de Milieukunde; W. P. Van Stockum & Zoon: The Hague, The Netherlands, 1934; Volume 263. [Google Scholar]
- Cao, S.; Zhang, W.; Ding, W.; Wang, M.; Fan, S.; Yang, B.; Mcminn, A.; Wang, M.; Xie, B.B.; Qin, Q.L.; et al. Structure and function of the Arctic and Antarctic marine microbiota as revealed by metagenomics. Microbiome 2020, 8, 47. [Google Scholar] [CrossRef]
- Galand, P.E.; Potvin, M.; Casamayor, E.O.; Lovejoy, C. Hydrography shapes bacterial biogeography of the deep Arctic Ocean. ISME J. 2010, 4, 564–576. [Google Scholar] [CrossRef] [PubMed]
- Fadeev, E.; Salter, I.; Schourup-Kristensen, V.; Schourup-Kristensen, S.; Bienhold, C.; Bienhold, C. Microbial communities in the east and west Fram Strait during sea ice melting season. Front. Mar. Sci. 2018, 5, 1–21. [Google Scholar] [CrossRef]
- Tuerena, R.E.; Hopkins, J.; Buchanan, P.J.; Ganeshram, R.S.; Norman, L.; von Appen, W.J.; Tagliabue, A.; Doncila, A.; Graeve, M.; Ludwichowski, K.U.; et al. An Arctic strait of two halves: The changing dynamics of nutrient uptake and limitation across the Fram Strait. Global Biogeochem. Cycles 2021, 35, 1–20. [Google Scholar] [CrossRef]
- Wietz, M.; Bienhold, C.; Metfies, K.; Torres-Valdés, S.; von Appen, W.J.; Salter, I.; Boetius, A. The polar night shift: Seasonal dynamics and drivers of Arctic Ocean microbiomes revealed by autonomous sampling. ISME Commun. 2021, 1, 76. [Google Scholar] [CrossRef]
- Karner, M.B.; DeLong, E.F.; Karl, D.M. Archaeal dominance in the mesopelagic zone of the Pacific Ocean. Nature 2001, 409, 507–510. [Google Scholar] [CrossRef]
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Structure and function of the global ocean microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef]
- Cardozo-Mino, M.G.; Fadeev, E.; Salman-Carvalho, V.; Boetius, A. Spatial distribution of Arctic bacterioplankton abundance is linked to distinct water masses and summertime phytoplankton bloom dynamics (Fram Strait, 79°N). Front. Microbiol. 2021, 12, 658803. [Google Scholar] [CrossRef]
- Quero, G.M.; Celussi, M.; Relitti, F.; Kovačević, V.; Del Negro, P.; Luna, G.M. Inorganic and organic carbon uptake processes and their connection to microbial diversity in meso- and bathypelagic Arctic Waters (Eastern Fram Strait). Microb. Ecol. 2020, 79, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Galand, P.E.; Casamayor, E.O.; Kirchman, D.L.; Lovejoy, C. Ecology of the rare microbial biosphere of the Arctic Ocean. Proc. Natl. Acad. Sci. USA 2009, 106, 22427–22432. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.; Rasmussen, S.; Blom, N.; Deming, J.W.; Rysgaard, S.; Sicheritz-Ponten, T. Microbial community structure of Arctic multiyear sea ice and surface seawater by 454 sequencing of the 16S RNA gene. ISME J. 2012, 6, 11–20. [Google Scholar] [CrossRef]
- Kirchman, D.L.; Cottrell, M.T.; Lovejoy, C. The structure of bacterial communities in the western Arctic Ocean as revealed by pyrosequencing of 16S rRNA genes. Environ. Microbiol. 2010, 12, 1132–1143. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.; Müller, O.; Nordmann, E.L.; Seuthe, L.; Bratbak, G.; Øvreås, L. Changes in marine prokaryote composition with season and depth over an Arctic polar year. Front. Mar. Sci. 2017, 4, 95. [Google Scholar] [CrossRef]
- de Sousa, A.G.G.; Tomasino, M.P.; Duarte, P.; Fernández-Méndez, M.; Assmy, P.; Ribeiro, H.; Surkont, J.; Leite, R.B.; Pereira-Leal, J.B.; Torgo, L.; et al. Diversity and composition of pelagic prokaryotic and protist communities in a thin Arctic sea-ice regime. Microb. Ecol. 2019, 78, 388–408. [Google Scholar] [CrossRef]
- von Jackowski, A.; Grosse, J.; Nöthig, E.M.; Engel, A. Dynamics of organic matter and bacterial activity in the Fram Strait during summer and autumn. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2020, 378, 20190366. [Google Scholar] [CrossRef]
- von Jackowski, A.; Becker, K.W.; Wietz, M.; Bienhold, C.; Zäncker, B.; Nöthig, E.M.; Engel, A. Variations of microbial communities and substrate regimes in the eastern Fram Strait between summer and fall. Environ. Microbiol. 2022, 24, 4124–4136. [Google Scholar] [CrossRef]
- Screen, J.A.; Simmonds, I. The central role of diminishing sea ice in recent Arctic temperature amplification. Nature 2010, 464, 1334–1337. [Google Scholar] [CrossRef]
- Sutherland, D.A.; Pickart, R.S. The East Greenland Coastal Current: Structure, variability, and forcing. Prog. Oceanogr. 2008, 78, 58–77. [Google Scholar] [CrossRef]
- Rudels, B.; Eriksson, P.; Grönvall, H.; Hietala, R.; Launiainen, J. Hydrographic observations in Denmark Strait in fall 1997, and their implications for the entrainment into the overflow plume. Geophys. Res. Lett. 1999, 26, 1325–1328. [Google Scholar] [CrossRef]
- Sternal, B.; Szczuciski, W.; Forwick, M.; Zajączkowski, M.; Lorenc, S.; Przytarska, J. Postglacial variability in near-bottom current speed on the continental shelf off south-west Spitsbergen. J. Quat. Sci. 2014, 29, 767–777. [Google Scholar] [CrossRef]
- Loeng, H. Features of the physical oceanographic conditions of the Barents Sea. Polar Res. 1991, 10, 5–18. [Google Scholar] [CrossRef]
- Carstensen, J.; Weydmann, A. Tipping points in the arctic: Eyeballing or statistical significance? Ambio 2012, 41, 34–43. [Google Scholar] [CrossRef]
- Polyakov, I.V.; Pnyushkov, A.V.; Alkire, M.B.; Ashik, I.M.; Baumann, T.M.; Carmack, E.C.; Goszczko, I.; Guthrie, J.; Ivanov, V.V.; Kanzow, T.; et al. Greater role for Atlantic inflows on sea-ice loss in the Eurasian Basin of the Arctic Ocean. Science 2017, 356, 285–291. [Google Scholar] [CrossRef]
- Andrews, A.J.; Christiansen, J.S.; Bhat, S.; Lynghammar, A.; Westgaard, J.I.; Pampoulie, C.; Præbel, K. Boreal marine fauna from the Barents Sea disperse to Arctic Northeast Greenland. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Weydmann-Zwolicka, A.; Prątnicka, P.; Łącka, M.; Majaneva, S.; Cottier, F.; Berge, J. Zooplankton and sediment fluxes in two contrasting fjords reveal Atlantification of the Arctic. Sci. Total Environ. 2021, 773, 145599. [Google Scholar] [CrossRef]
- Weydmann, A.; Carstensen, J.; Goszczko, I.; Dmoch, K.; Olszewska, A.; Kwasniewski, S. Shift towards the dominance of boreal species in the Arctic: Inter-annual and spatial zooplankton variability in the West Spitsbergen Current. Mar. Ecol. Prog. Ser. 2014, 501, 41–52. [Google Scholar] [CrossRef]
- Mańko, K.M.; Głuchowska, M.; Weydmann-Zwolicka, A. Footprints of Atlantification in the vertical distribution and diversity of gelatinous zooplankton in the Fram Strait (Arctic Ocean). Prog. Oceanogr. 2020, 189, 102414. [Google Scholar] [CrossRef]
- Kortsch, S.; Primicerio, R.; Fossheim, M.; Dolgov, A.V.; Aschan, M. Climate change alters the structure of arctic marine food webs due to poleward shifts of boreal generalists. Proc. R. Soc. B. 2015, 282, 20151546. [Google Scholar] [CrossRef] [PubMed]
- Møller, E.F.; Nielsen, T.G. Borealization of Arctic zooplankton—Smaller and less fat zooplankton species in Disko Bay, Western Greenland. Limnol. Oceanogr. 2020, 65, 1175–1188. [Google Scholar] [CrossRef]
- Stempniewicz, L.; Weydmann-Zwolicka, A.; Strzelewicz, A.; Goc, M.; Głuchowska, M.; Kidawa, D.; Walczowski, W.; Węsławski, J.M.; Zwolicki, A. Advection of Atlantic water masses influences seabird community foraging in a high-Arctic fjord. Prog. Oceanogr. 2021, 193, 102549. [Google Scholar] [CrossRef]
- Vincent, W. Microbial ecosystem responses to rapid climate change in the Arctic. ISME J. 2010, 4, 1087–1090. [Google Scholar] [CrossRef]
- Bowman, J.S. The relationship between sea ice bacterial community structure and biogeochemistry: A synthesis of current knowledge and known unknowns. Elementa-Sci. Anthrop. 2015, 3, 000072. [Google Scholar] [CrossRef]
- Cottier, F.; Tverberg, V.; Inall, M.; Svendsen, H.; Nilsen, F.; Griffiths, C. Water mass modification in an Arctic fjord through cross-shelf exchange: The seasonal hydrography of Kongsfjorden, Svalbard. J. Geophys. Res. Ocean. 2005, 110, 1–18. [Google Scholar] [CrossRef]
- Rudels, B.; Korhonen, M.; Budeus, G.; Beszczynska-Moeller, A.; Schauer, U.; Nummelin, A.; Quadfasel, D.; Valdimarsson, H. The East Greenland Current and its impacts on the Nordic Seas: Observed trends in the past decade. ICES J. Mar. Sci. 2012, 69, 841–851. [Google Scholar] [CrossRef]
- Cherkasheva, A.; Nöthig, E.M.; Bauerfeind, E.; Melsheimer, C.; Bracher, A. From the chlorophyll a in the surface layer to its vertical profile: A Greenland Sea relationship for satellite applications. Ocean Sci. 2013, 9, 431–445. [Google Scholar] [CrossRef]
- Dąbrowska, A.M.; Wiktor, J.M., Jr.; Merchel, M.; Wiktor, J.M. Planktonic protists of the Eastern Nordic Seas and the Fram Strait: Spatial changes related to hydrography during early summer. Front. Mar. Sci. 2020, 7, 557. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplles, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal. RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res 2013, 41, e1. [Google Scholar] [CrossRef]
- Caporaso, J.G.J.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods. 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastq: An ultra fast all in one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Aronesty, E. ea-utils: Command-Line Tools for Processing Biological Sequencing Data. Available online: https://github.com/ExpressionAnalysis/ea-utils (accessed on 14 September 2021).
- Magoc, T.; Salzberg, S. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Li, W.; Fu, L.; Niu, B.; Wu, S.; Wooley, J. Ultrafast clustering algorithms for metagenomic sequence analysis. Brief. Bioinform. 2012, 13, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Lemos, L.N.; Fulthorpe, R.R.; Triplett, E.W.; Roesch, L.F. Rethinking microbial diversity analysis in the high throughput sequencing era. J. Microbiol. Methods 2011, 86, 42–51. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; John Wiley & Sons: Hoboken, NJ, USA, 2013; p. 272. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Lozupone, C.; Hamady, M.; Knight, R. UniFrac—An online tool for comparing microbial community diversity in a phylogenetic context. BMC Bioinform. 2006, 7, 371. [Google Scholar] [CrossRef]
- Wickham, H. Tidyr: Tidy Messy Data. R Package Version 1.1.2. 2020. Available online: https://CRAN.R-project.org/package=tidyr (accessed on 18 January 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Gentleman, R., Hornik, K., Parmigiani, G., Eds.; Springer: Berlin, Germany, 2016; p. 268. [Google Scholar]
- Garnier, S. Viridis: Default Color Maps from ‘matplotlib’. R Package Version 0.5.1. 2018. Available online: https://CRAN.R-project.org/package=viridis (accessed on 18 January 2021).
- ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination. Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012; p. 496. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Bah, T. Inkscape: Guide to a Vector Drawing Program; Prentice Hall: Hoboken, NJ, USA, 2011; Volume 559, p. 473. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Blindheim, J.; Østerhus, S. The Nordic seas, main oceanographic features. Geophys. Monogr. Ser. 2005, 158, 11–37. [Google Scholar] [CrossRef]
- Willis, A.D. Rarefaction, alpha diversity, and statistics. Front. Microbiol. 2019, 10, 2407. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Delmont, T.O.; Quince, C.; Shaiber, A.; Esen, Ö.C.; Lee, S.T.M.; Rappé, M.S.; McLellan, S.L.; Lücker, S.; Eren, A.M. Nitrogen-fixing populations of planctomycetes and proteobacteria are abundant in surface ocean metagenomes. Nat. Microbiol. 2018, 3, 804–813. [Google Scholar] [CrossRef]
- Auguet, J.C.; Barberan, A.; Casamayor, E.O. Global ecological patterns in uncultured Archaea. ISME J. 2010, 4, 182–190. [Google Scholar] [CrossRef]
- Follows, M.J.; Dutkiewicz, S.; Grant, S.; Chisholm, S.W. Emergent biogeography of microbial communities in a model ocean. Science 2007, 315, 1843–1846. [Google Scholar] [CrossRef] [PubMed]
- Hansman, R.L.; Griffin, S.; Watson, J.T.; Aluwihare, L.I. The radiocarbon signature of microorganisms in the mesopelagic ocean. Proc. Natl. Acad. Sci. USA 2009, 106, 6513–6518. [Google Scholar] [CrossRef] [PubMed]
- Pita, L.; López-Legentil, S.; Erwin, P.M. Biogeography and host fidelity of bacterial communities in Ircinia spp. from the Bahamas. Microb. Ecol. 2013, 66, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, H.; Ikemoto, H.; Kurano, N.; Adachi, K.; Chihara, M.; Miyachi, S. Chlorophyll d as a major pigment. Nature 1996, 383, 402. [Google Scholar] [CrossRef]
- Kühl, M.; Chen, M.; Ralph, P.J.; Schreiber, U.; Larkum, A.W.D. A niche for cyanobacteria containing chlorophyll d. Nature 2005, 433, 820. [Google Scholar] [CrossRef] [PubMed]
- Hirose, E.; Maruyama, T. What are the benefits in the ascidian-Prochloron symbiosis? Endocytobiosis Cell Res. 2004, 15, 51–62. [Google Scholar]
- Schreiber, L.; Kjeldsen, K.U.; Funch, P.; Jensen, J.; Obst, M.; López-Legentil, S.; Schramm, A. Endozoicomonas are specific, facultative symbionts of sea squirts. Front. Microbiol. 2016, 7, 1042. [Google Scholar] [CrossRef]
- Schreiber, L.; Kjeldsen, K.U.; Obst, M.; Funch, P.; Schramm, A. Description of Endozoicomonas ascidiicola sp. nov., isolated from Scandinavian ascidians. Syst. Appl. Microbiol. 2016, 39, 313–318. [Google Scholar] [CrossRef]
- Pedrós-Alió, C.; Potvin, M.; Lovejoy, C. Diversity of planktonic microorganisms in the Arctic Ocean. Prog. Oceanogr. 2015, 139, 233–243. [Google Scholar] [CrossRef]
- Becker, J.W.; Hogle, S.L.; Rosendo, K.; Chisholm, S.W. Co-culture and biogeography of Prochlorococcus and SAR11. ISME J. 2019, 13, 1506–1519. [Google Scholar] [CrossRef] [PubMed]
- Bano, N.; Hollibaugh, J.T. Phylogenetic composition of bacterioplankton assemblages from the Arctic Ocean. Appl. Environ. Microbiol. 2002, 68, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Biller, S.J.; Berube, P.M.; Lindell, D.; Chisholm, S.W. Prochlorococcus: The structure and function of collective diversity. Nat. Rev. Microbiol. 2014, 13, 13–27. [Google Scholar] [CrossRef]
- Kettler, C.G.; Martiny, A.C.; Huang, K.; Zucker, J.; Coleman, M.L.; Rodrigue, S.; Chen, F.; Lapidus, A.; Ferriera, S.; Johnson, J.; et al. Patterns and implications of gene gain and loss in the evolution of Prochlorococcus. PLoS Genet. 2007, 3, e231. [Google Scholar] [CrossRef]
- West, N.J.; Schönhuber, W.A.; Fuller, N.J.; Amann, R.I.; Rippka, R.; Post, A.F.; Scanlan, D.J. Closely related Prochlorococcus genotypes show remarkably different depth distributions in two oceanic regions as revealed by in situ hybridization using 16S rRNA-targeted oligonucleotides. Microbiology 2001, 147, 1731–1744. [Google Scholar] [CrossRef]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Martiny, A.C. Present and future global distributions of the marine cyanobacteria Prochlrococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, M.L.; Doré, H.; Garczarek, L.; Seuthe, L.; Müller, O.; Sandaa, R.A.; Bratbak, G.; Larsen, A. Synechococcus in the Atlantic gateway to the Arctic Ocean. Front. Mar. Sci. 2016, 3, 191. [Google Scholar] [CrossRef]
- Mańko, M.K.; Merchel, M.; Kwasniewski, S.; Weydmann-Zwolicka, A. Oceanic fronts shape biodiversity of gelatinous zooplankton in the European Arctic. Front. Mar. Sci. 2022, 9, 941025. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Station | Latitude | Longitude | Sampling Date | Depth [m] |
|---|---|---|---|---|---|
| Spitsbergen | ISSH | 77°31,349′ N | 10°44,314′ E | 14 August 2018 | 980 |
| HRSH | 76°57,348′ N | 12°59,901′ E | 14 August 2018 | 287 | |
| Greenland | GR3 | 76°53,604′ N | 7°04,895′ W | 16 August 2018 | 299 |
| GR5 | 75°53,488′ N | 12°56,915′ W | 17 August 2018 | 275 | |
| GR6 | 76°23,954′ N | 10°38,947′ W | 17 August 2018 | 304 |
| Geographical Region | Sample ID | No. of Bacterial Reads (% Passing QC) | No. of Observed OTUs (at 97% Sequence Similarity Cut-Off) | No of Sequences for Which No Homology Sequences Were Found in NCBI Database |
|---|---|---|---|---|
| Greenland shelf | GR3.0m | 111,709 (98.23%) | 212 | 239 |
| GR3.25m | 97,920 (98.27%) | 332 | 2580 | |
| GR3.50m | 106,871 (98.28%) | 446 | 1614 | |
| GR5.0m | 100,812 (98.35%) | 203 | 383 | |
| GR5.25m | 121,097 (98.35%) | 314 | 1649 | |
| GR5.50m | 105,060 (98.28%) | 418 | 1421 | |
| GR6.0m | 107,538 (98.23%) | 215 | 622 | |
| GR6.25m | 111,056 (98.27%) | 223 | 236 | |
| GR6.50m | 93,150 (98.38%) | 385 | 1024 | |
| Spitsbergen shelf | HRSH.0m | 135,715 (98.45%) | 292 | 1409 |
| HRSH.25m | 107,635 (98.31%) | 285 | 1037 | |
| ISSH.0m | 109,873 (98.20%) | 281 | 1522 | |
| ISSH.25m | 107,576 (98.27%) | 357 | 1414 | |
| ISSH.50m | 119,237 (98.41%) | 411 | 2163 |
| Variable | Explains % | Contribution % | Pseudo-F | P | P (adj) |
|---|---|---|---|---|---|
| Salinity | 45.9 | 57.5 | 10.2 | 0.001 | 0.00333 |
| Temperature | 21.5 | 26.9 | 7.3 | 0.001 | 0.001 |
| Depth | 5.4 | 6.8 | 2 | 0.025 | 0.04167 |
| Bacterial Species | Samples | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GR3 | GR5 | GR6 | ISSH | HRSH | ||||||||||
| 0 | 25 | 50 | 0 | 25 | 50 | 0 | 25 | 50 | 0 | 25 | 50 | 0 | 25 | |
| Specific bacteria for geographical region | ||||||||||||||
| Dulcicalothrix necridiiformans | + | + | - | + | + | - | + | + | - | - | - | - | - | - |
| Lewinella xylanilytica | + | - | - | + | + | - | + | + | + | - | - | - | - | - |
| Olleya marilimosa | - | + | + | - | + | + | - | - | + | - | - | - | - | - |
| Rubritalea marina | + | + | + | + | + | + | + | + | + | - | - | - | - | - |
| Vicingus serpentipes | + | + | + | + | + | + | + | - | + | - | - | - | - | - |
| Dulcicalothrix necridiiformans | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Leisingera aquaemixtae | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Luteibaculum oceani | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Marinifilum albidiflavum | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Olleya algicola | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Phaeocystidibacter marisrubri | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Portibacter lacus | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Porticoccus hydrocarbonoclasticus | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Pseudofulvibacter gastropodicola | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Pseudohongiella nitratireducens | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Roseibacillus persicicus | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Bacterial symbiont | ||||||||||||||
| Acaryochloris marina | + | - | - | + | + | - | + | - | - | - | - | - | + | + |
| Endozoicomonas ascidiicola | - | + | + | - | + | + | - | + | + | - | - | - | - | - |
| Cyanobacteria | ||||||||||||||
| Brasilonema tolantongensis | + | - | - | + | - | - | + | - | - | - | - | - | + | - |
| Chamaesiphon minutus | - | - | - | - | - | - | - | - | - | + | - | - | - | - |
| Cylindrospermum stagnale | + | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Prochlorococcus marinus | - | - | - | - | - | - | - | - | - | + | + | + | + | + |
| Scytonema hofmannii | - | - | - | - | + | - | - | + | - | - | - | - | - | - |
| Stations | Spitsbergen Shelf Station ISSH | Spitsbergen Shelf Station HRSH | |||
|---|---|---|---|---|---|
| Depth | 0 m | 25 m | 50 m | 0 m | 25 m |
| Number of identified P. marinus sequences | 4000 | 829 | 283 | 2811 | 1826 |
| Abundance of identified P. marinus in microbial communities | 10% | 2% | 1% | 6% | 5% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mioduchowska, M.; Pawłowska, J.; Mazanowski, K.; Weydmann-Zwolicka, A. Contrasting Marine Microbial Communities of the Fram Strait with the First Confirmed Record of Cyanobacteria Prochlorococcus marinus in the Arctic Region. Biology 2023, 12, 1246. https://doi.org/10.3390/biology12091246
Mioduchowska M, Pawłowska J, Mazanowski K, Weydmann-Zwolicka A. Contrasting Marine Microbial Communities of the Fram Strait with the First Confirmed Record of Cyanobacteria Prochlorococcus marinus in the Arctic Region. Biology. 2023; 12(9):1246. https://doi.org/10.3390/biology12091246
Chicago/Turabian StyleMioduchowska, Monika, Joanna Pawłowska, Karol Mazanowski, and Agata Weydmann-Zwolicka. 2023. "Contrasting Marine Microbial Communities of the Fram Strait with the First Confirmed Record of Cyanobacteria Prochlorococcus marinus in the Arctic Region" Biology 12, no. 9: 1246. https://doi.org/10.3390/biology12091246
APA StyleMioduchowska, M., Pawłowska, J., Mazanowski, K., & Weydmann-Zwolicka, A. (2023). Contrasting Marine Microbial Communities of the Fram Strait with the First Confirmed Record of Cyanobacteria Prochlorococcus marinus in the Arctic Region. Biology, 12(9), 1246. https://doi.org/10.3390/biology12091246