
Revisiting the Tigger Transposon Evolution Revealing Extensive Involvement in the Shaping of Mammal Genomes

 , and
, and 
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Mining of Tigger Transposons
2.2. Structure and Phylogenetic Analysis of Tigger
2.3. Evidence of HT for Tigger across the Animal Kingdom
3. Results
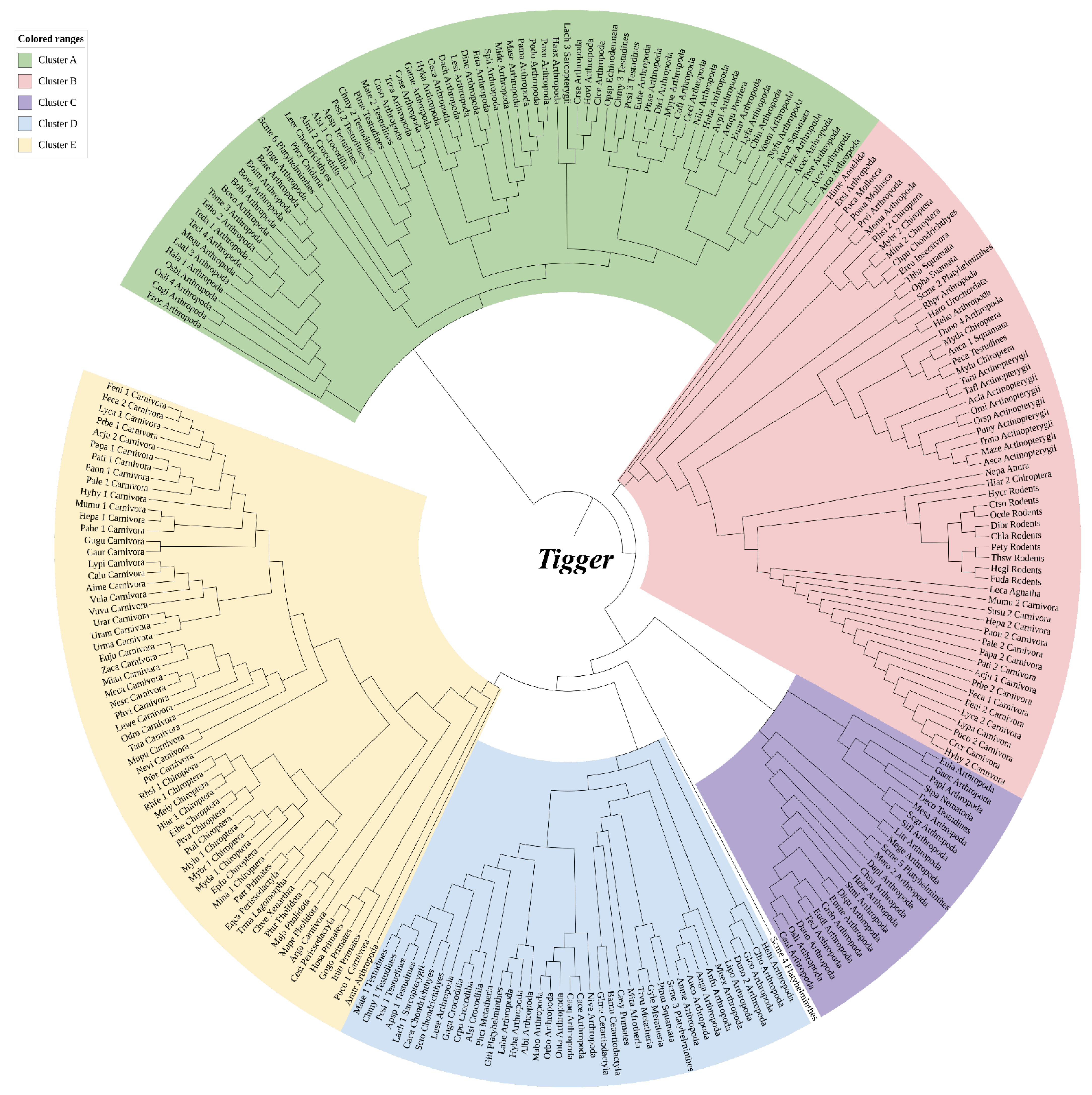
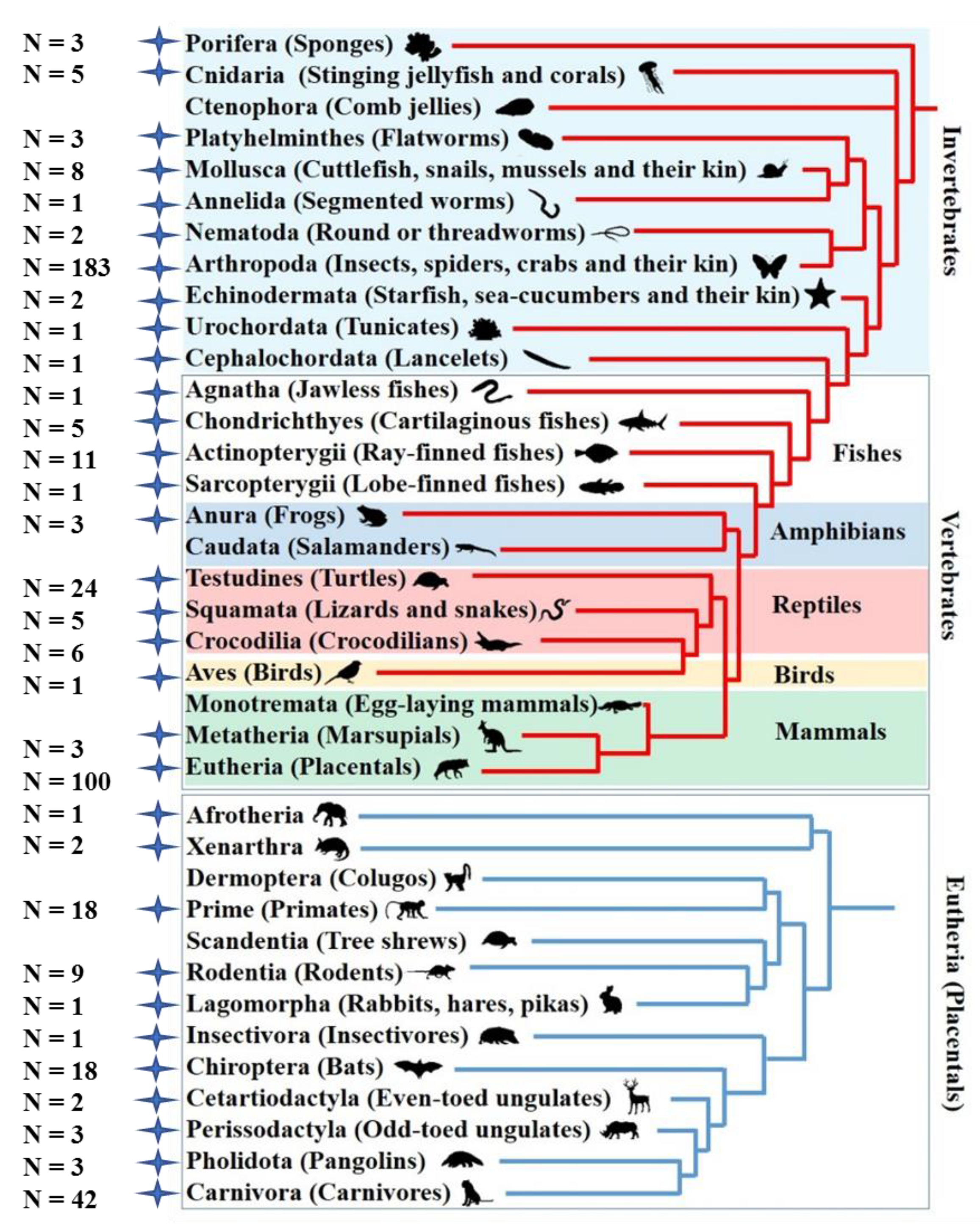
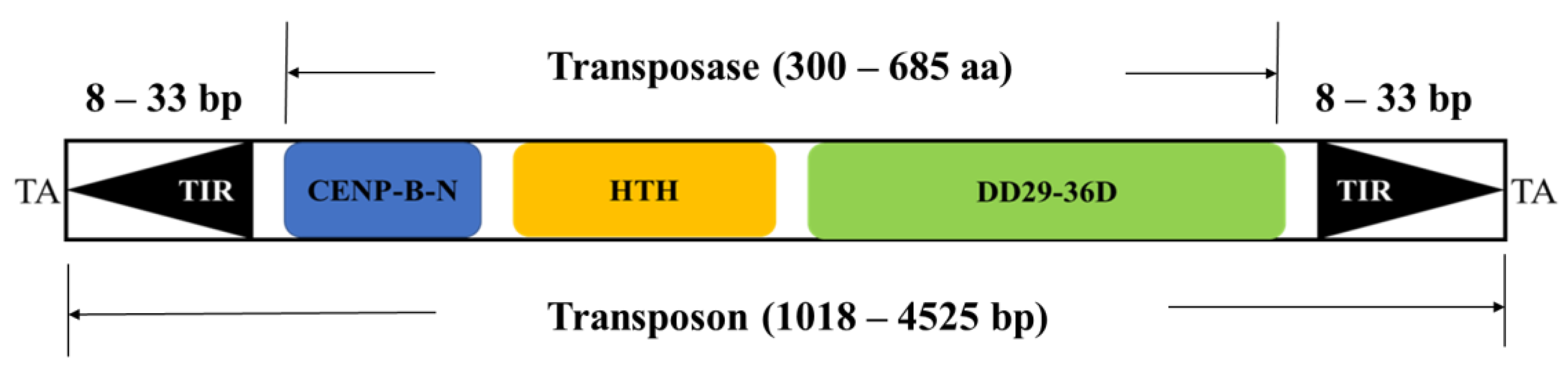
3.1. Taxonomic Distribution and Structure Organization of Tigger
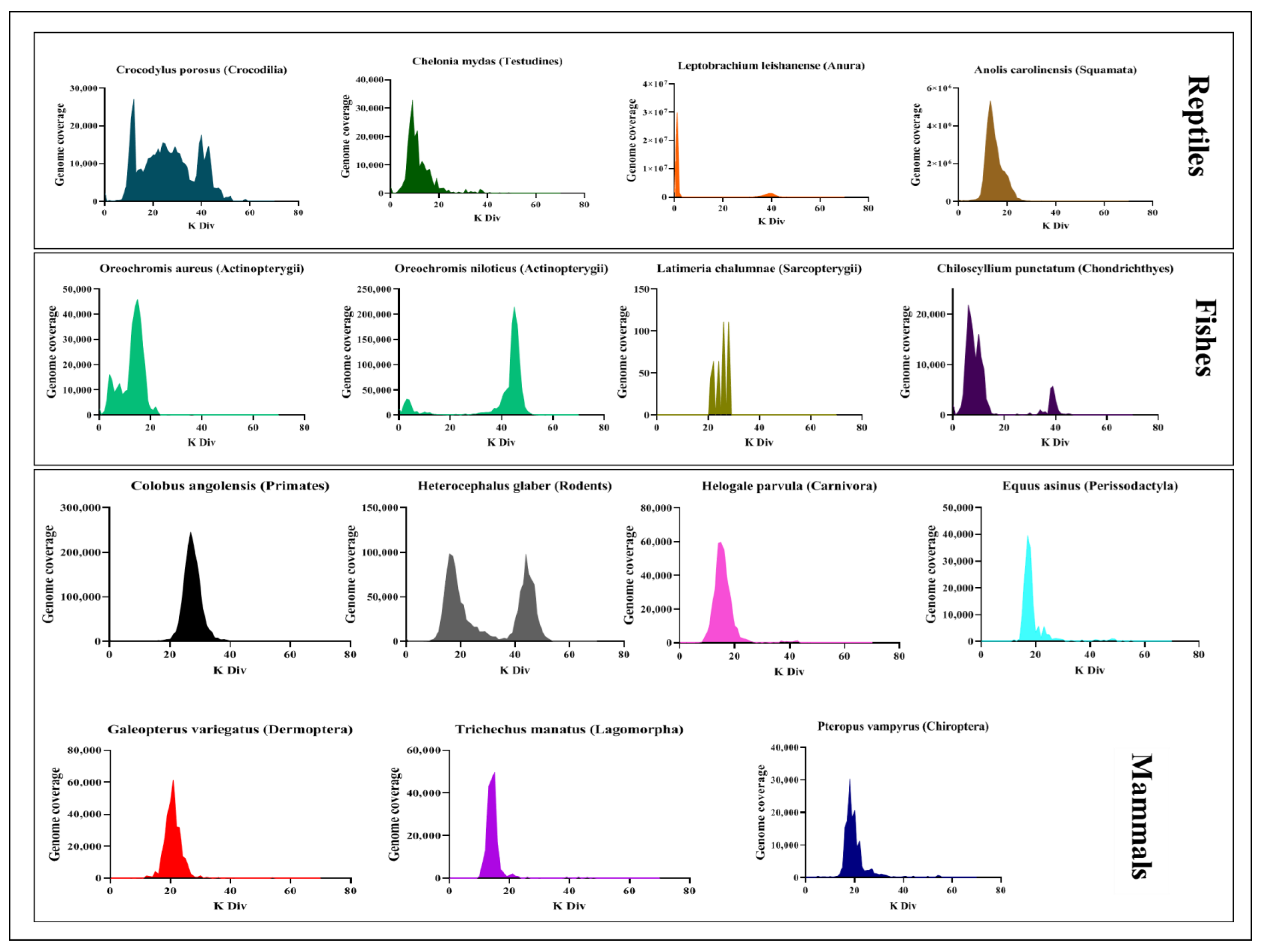
3.2. Tigger Evolutionary Dynamics in Vertebrate Genomes
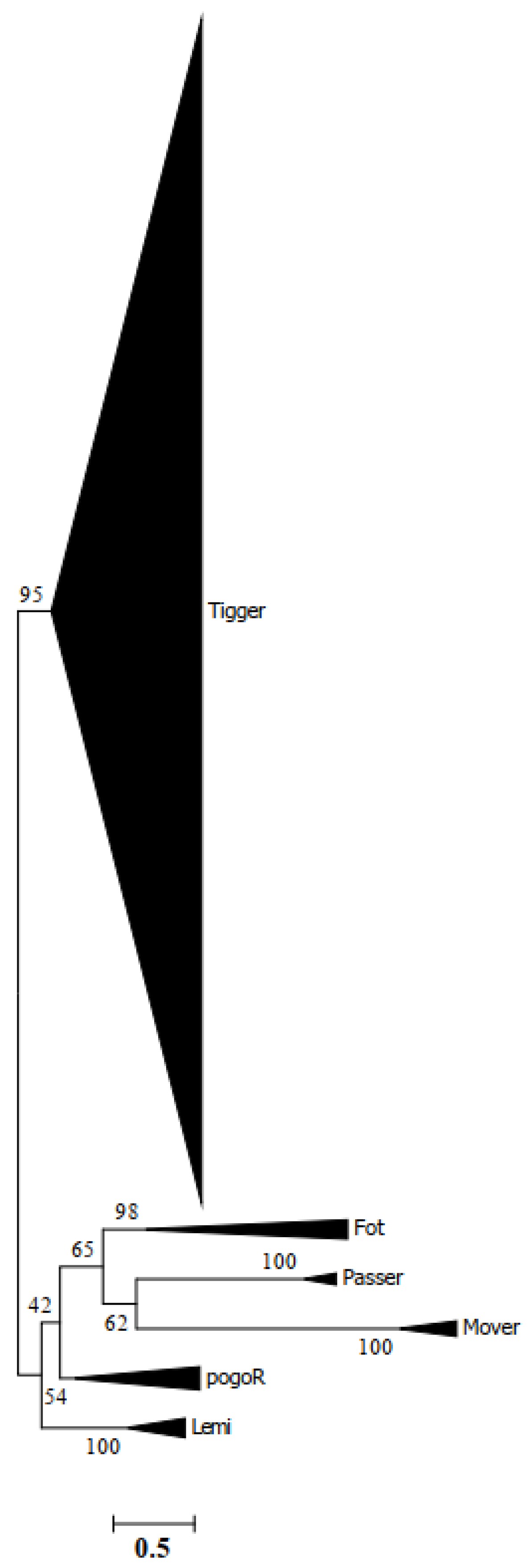
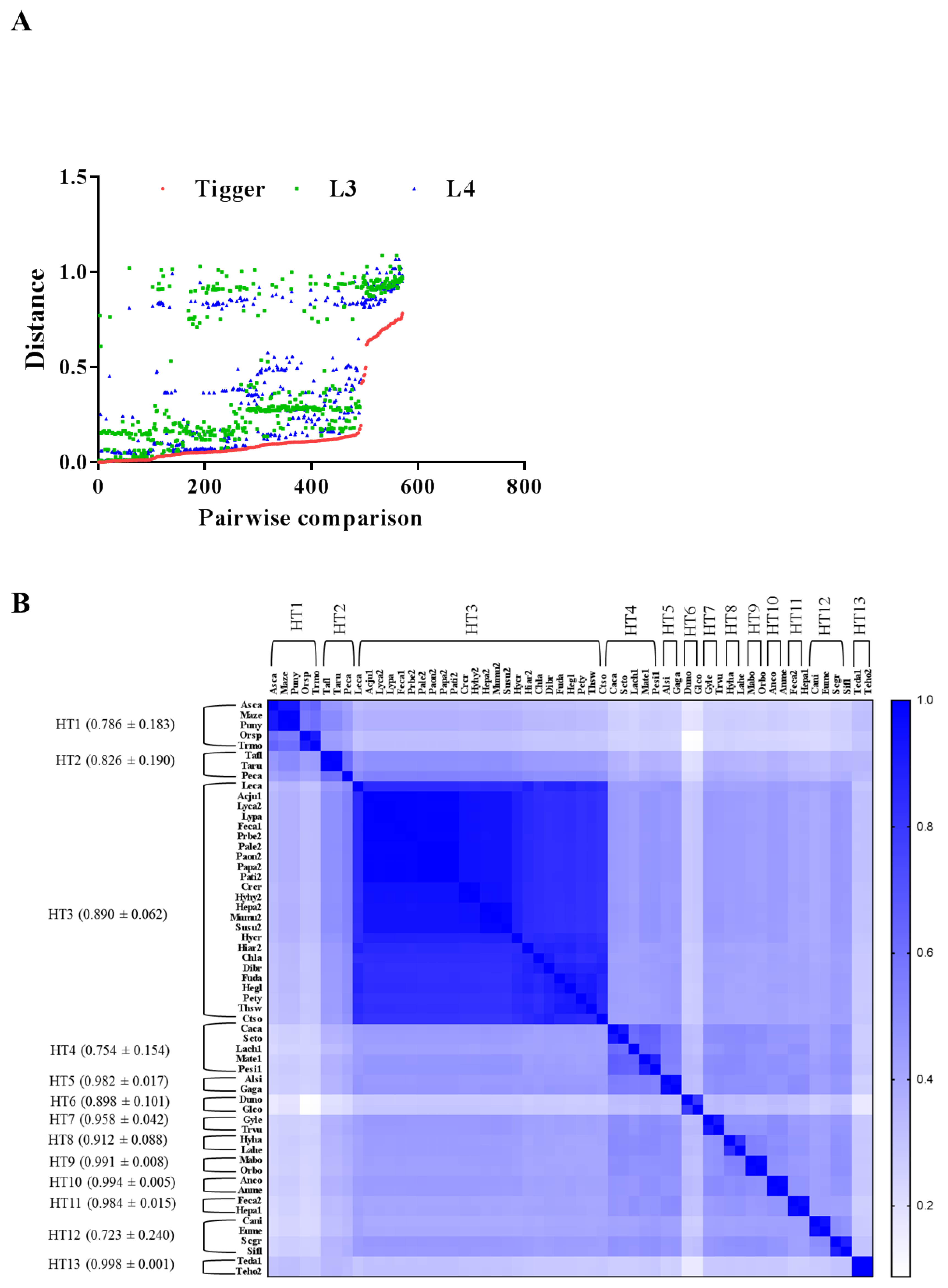
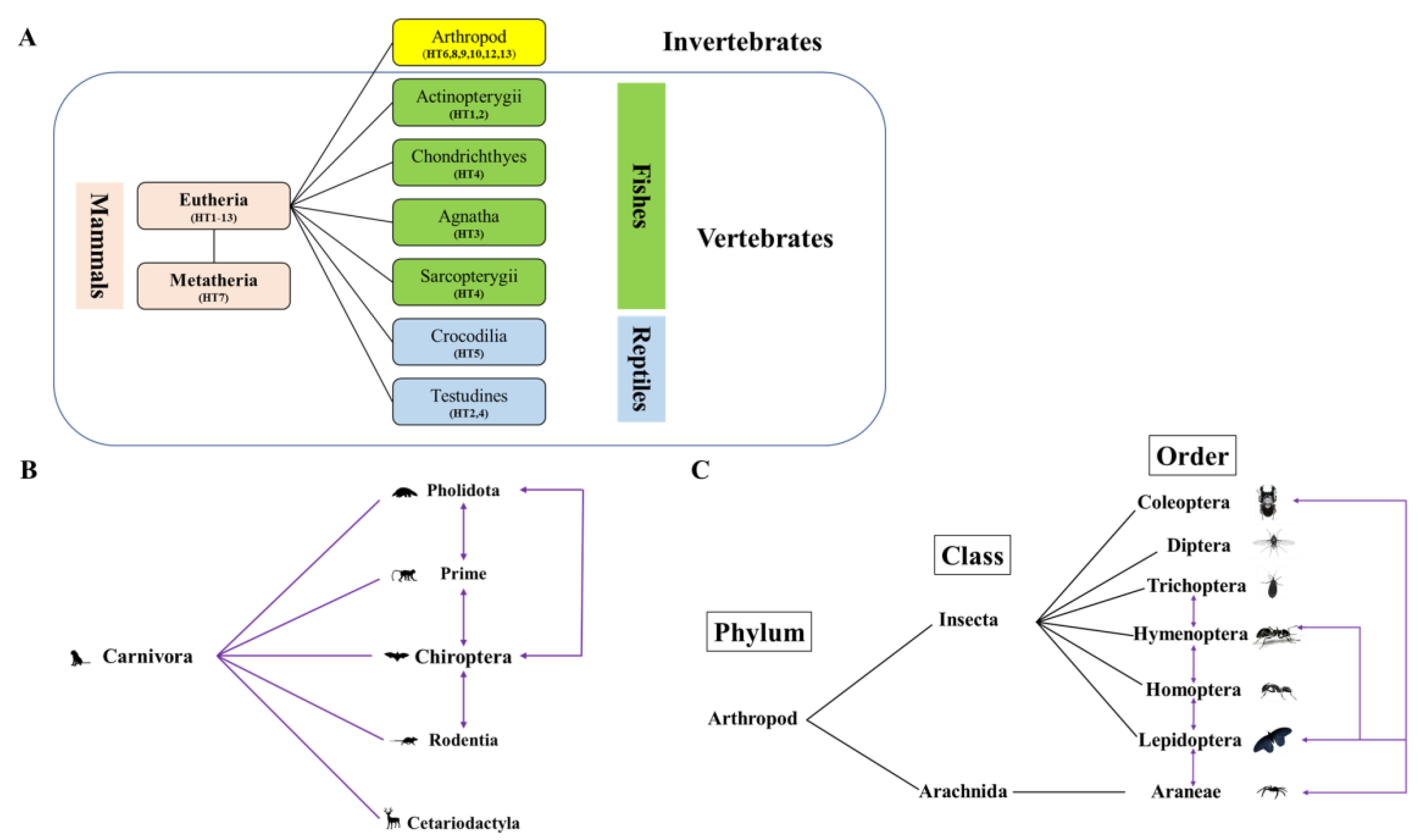
3.3. Evidence of HT Events for Tigger across Animals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ochman, H.; Lawrence, J.G.; Groisman, E.A. Lateral Gene Transfer and the Nature of Bacterial Innovation. Nature 2000, 405, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Makarova, K.S.; Aravind, L. Horizontal Gene Transfer in Prokaryotes: Quantification and Classification. Annu. Rev. Microbiol. 2001, 55, 709–742. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J.; Palmer, J.D. Horizontal Gene Transfer in Eukaryotic Evolution. Nat. Rev. Genet. 2008, 9, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Danchin, E.G.J.; Rosso, M.-N.; Vieira, P.; de Almeida-Engler, J.; Coutinho, P.M.; Henrissat, B.; Abad, P. Multiple Lateral Gene Transfers and Duplications Have Promoted Plant Parasitism Ability in Nematodes. Proc. Natl. Acad. Sci. USA 2010, 107, 17651–17656. [Google Scholar] [CrossRef]
- Marcet-Houben, M.; Gabaldón, T. Acquisition of Prokaryotic Genes by Fungal Genomes. Trends Genet. 2010, 26, 5–8. [Google Scholar] [CrossRef]
- Andersson, J.O. Gene Transfer and Diversification of Microbial Eukaryotes. Annu. Rev. Microbiol. 2009, 63, 177–193. [Google Scholar] [CrossRef]
- Gladyshev, E.A.; Meselson, M.; Arkhipova, I.R. Massive Horizontal Gene Transfer in Bdelloid Rotifers. Science 2008, 320, 1210–1213. [Google Scholar] [CrossRef]
- Moran, N.A.; Jarvik, T. Lateral Transfer of Genes from Fungi Underlies Carotenoid Production in Aphids. Science 2010, 328, 624–627. [Google Scholar] [CrossRef]
- Slot, J.C.; Rokas, A. Horizontal Transfer of a Large and Highly Toxic Secondary Metabolic Gene Cluster between Fungi. Curr. Biol. 2011, 21, 134–139. [Google Scholar] [CrossRef]
- Schaack, S.; Gilbert, C.; Feschotte, C. Promiscuous DNA: Horizontal Transfer of Transposable Elements and Why It Matters for Eukaryotic Evolution. Trends Ecol. Evol. 2010, 25, 537–546. [Google Scholar] [CrossRef]
- Craig, N.L. Tn7. In Mobile DNA II; American Society of Microbiology: Washington, DC, USA, 2002; pp. 423–456. [Google Scholar]
- Lander, E.S.L.; Birren, L.M.; Nusbaum, B.; Zody, C.; Baldwin, M.C.; Devon, J.; Dewar, K.; Doyle, K.; Fitz Hugh, M.; Funke, W.; et al. Initial Sequencing and Analysis of the Human Genome. Nature 2001, 409, 860–921. [Google Scholar] [PubMed]
- Feschotte, C.; Pritham, E.J. DNA Transposons and the Evolution of Eukaryotic Genomes. Annu. Rev. Genet. 2007, 41, 331–368. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Wang, Y.; Diaby, M.; Zong, W.; Shen, D.; Wang, S.; Chen, C.; Wang, X.; Song, C. Evolution of Pogo, a Separate Superfamily of IS630-Tc1-Mariner Transposons, Revealing Recurrent Domestication Events in Vertebrates. Mob. DNA 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Han, M.-J.; Xu, H.-E.; Zhang, H.-H.; Feschotte, C.; Zhang, Z. Spy: A New Group of Eukaryotic DNA Transposons without Target Site Duplications. Genome Biol. Evol. 2014, 6, 1748–1757. [Google Scholar] [CrossRef] [PubMed]
- Jurka, J. Repbase Update: A Database and an Electronic Journal of Repetitive Elements. Trends Genet. 2000, 16, 418–420. [Google Scholar] [CrossRef]
- Silva, J.C.; Loreto, E.L.; Clark, J.B. Factors That Affect the Horizontal Transfer of Transposable Elements. Curr. Issues Mol. Biol. 2004, 6, 57–72. [Google Scholar] [PubMed]
- Syvanen, M. Temporal Patterns of Plant and Metazoan Evolution Suggest Extensive Polyphyly. In Horizontal Gene Transfer; Elsevier: Amsterdam, The Netherlands, 2002; pp. 383–395. [Google Scholar]
- Hartl, D.L.; Lohe, A.R.; Lozovskaya, E.R. Modern Thoughts on an Ancyent Marinere: Function, Evolution, Regulation. Annu. Rev. Genet. 1997, 31, 337–358. [Google Scholar] [CrossRef]
- Yoshiyama, M.; Tu, Z.; Kainoh, Y.; Honda, H.; Shono, T.; Kimura, K. Possible Horizontal Transfer of a Transposable Element from Host to Parasitoid. Mol. Biol. Evol. 2001, 18, 1952–1958. [Google Scholar] [CrossRef]
- Loreto, E.L.S.; Carareto, C.M.A.; Capy, P. Revisiting Horizontal Transfer of Transposable Elements in Drosophila. Heredity 2008, 100, 545–554. [Google Scholar] [CrossRef]
- de Boer, J.G.; Yazawa, R.; Davidson, W.S.; Koop, B.F. Bursts and Horizontal Evolution of DNA Transposons in the Speciation of Pseudotetraploid Salmonids. BMC Genom. 2007, 8, 422. [Google Scholar] [CrossRef]
- Palazzo, A.; Escuder, E.; D’Addabbo, P.; Lovero, D.; Marsano, R.M. A Genomic Survey of Tc1-Mariner Transposons in Nematodes Suggests Extensive Horizontal Transposon Transfer Events. Mol. Phylogenet. Evol. 2021, 158, 107090. [Google Scholar] [CrossRef] [PubMed]
- Diao, X.; Freeling, M.; Lisch, D. Horizontal Transfer of a Plant Transposon. PLoS Biol. 2006, 4, e5. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Konkel, M.K.; Xing, J.; Wang, H.; Lee, J.; Meyer, T.J.; Huang, C.T.; Sandifer, E.; Hebert, K.; Barnes, E.W.; et al. Mobile DNA in Old World Monkeys: A Glimpse through the Rhesus Macaque Genome. Science 2007, 316, 238–240. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gifford, R.; Tristem, M. The Evolution, Distribution and Diversity of Endogenous Retroviruses. Virus Genes 2003, 26, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Yohn, C.T.; Jiang, Z.; McGrath, S.D.; Hayden, K.E.; Khaitovich, P.; Johnson, M.E.; Eichler, M.Y.; McPherson, J.D.; Zhao, S.; Pääbo, S.; et al. Lineage-Specific Expansions of Retroviral Insertions within the Genomes of African Great Apes but Not Humans and Orangutans. PLoS Biol. 2005, 3, e110. [Google Scholar] [CrossRef]
- Tarlinton, R.E.; Meers, J.; Young, P.R. Retroviral Invasion of the Koala Genome. Nature 2006, 442, 79–81. [Google Scholar] [CrossRef]
- Piskurek, O.; Okada, N. Poxviruses as Possible Vectors for Horizontal Transfer of Retroposons from Reptiles to Mammals. Proc. Natl. Acad. Sci. USA 2007, 104, 12046–12051. [Google Scholar] [CrossRef]
- Kordis, D.; Gubensek, F. Unusual Horizontal Transfer of a Long Interspersed Nuclear Element between Distant Vertebrate Classes. Proc. Natl. Acad. Sci. USA 1998, 95, 10704–10709. [Google Scholar] [CrossRef]
- Kidwell, M.G. Horizontal Transfer of P Elements and Other Short Inverted Repeat Transposons. In Transposable Elements and Evolution; Springer: Berlin/Heidelberg, Germany, 1993; pp. 158–172. [Google Scholar]
- Yuan, Y.-W.; Wessler, S.R. The Catalytic Domain of All Eukaryotic Cut-and-Paste Transposase Superfamilies. Proc. Natl. Acad. Sci. USA 2011, 108, 7884–7889. [Google Scholar] [CrossRef]
- Shi, S.; Puzakov, M.; Guan, Z.; Xiang, K.; Diaby, M.; Wang, Y.; Wang, S.; Song, C.; Gao, B. Prokaryotic and Eukaryotic Horizontal Transfer of Sailor (DD82E), a New Superfamily of IS630-Tc1-Mariner DNA Transposons. Biology 2021, 10, 1005. [Google Scholar] [CrossRef]
- Shao, H.; Tu, Z. Expanding the Diversity of the IS630-Tc1-Mariner Superfamily: Discovery of a Unique DD37E Transposon and Reclassification of the DD37D and DD39D Transposons. Genetics 2001, 159, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Tellier, M.; Bouuaert, C.C.; Chalmers, R. Mariner and the ITm Superfamily of Transposons. In Mobile DNA III; ASM Press: Washington, DC, USA, 2015; pp. 753–772. [Google Scholar]
- Gao, B.; Zong, W.; Miskey, C.; Ullah, N.; Diaby, M.; Chen, C.; Wang, X.; Ivics, Z.; Song, C. Intruder (DD38E), a Recently Evolved Sibling Family of DD34E/Tc1 Transposons in Animals. Mob. DNA 2020, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Jurka, M.G.; Kapitonov, V.V.; Jurka, J. New Superfamilies of Eukaryotic DNA Transposons and Their Internal Divisions. Mol. Biol. Evol. 2009, 26, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Coy, M.R.; Tu, Z. Gambol and Tc1 Are Two Distinct Families of DD34E Transposons: Analysis of the Anopheles Gambiae Genome Expands the Diversity of the IS630-Tc1-Mariner Superfamily. Insect Mol. Biol. 2005, 14, 537–546. [Google Scholar] [CrossRef]
- Zong, W.; Gao, B.; Diaby, M.; Shen, D.; Wang, S.; Wang, Y.; Sang, Y.; Chen, C.; Wang, X.; Song, C. Traveler, a New DD35E Family of Tc1/mariner Transposons, Invaded Vertebrates Very Recently. Genome Biol. Evol. 2020, 12, 66–76. [Google Scholar] [CrossRef]
- Zhang, H.-H.; Shen, Y.-H.; Xiong, X.-M.; Han, M.-J.; Zhang, X.-G. Identification and Evolutionary History of the DD41D Transposons in Insects. Genes Genom. 2016, 38, 109–117. [Google Scholar] [CrossRef]
- Zhang, H.H.; Li, G.Y.; Xiong, X.M.; Han, M.J.; Zhang, X.G.; Dai, F.Y. TRT, a Vertebrate and Protozoan tc1-like Transposon: Current Activity and Horizontal Transfer. Genome Biol. Evol. 2016, 8, 2994–3005. [Google Scholar] [CrossRef]
- Wang, S.; Diaby, M.; Puzakov, M.; Ullah, N.; Wang, Y.; Danley, P.; Chen, C.; Wang, X.; Gao, B.; Song, C. Divergent Evolution Profiles of DD37D and DD39D Families of Tc1/mariner Transposons in Eukaryotes. Mol. Phylogenet. Evol. 2021, 161, 107143. [Google Scholar] [CrossRef]
- Shen, D.; Gao, B.; Miskey, C.; Chen, C.; Sang, Y.; Zong, W.; Wang, S.; Wang, Y.; Wang, X.; Ivics, Z.; et al. Multiple Invasions of Visitor, a DD41D Family of Tc1/mariner Transposons, throughout the Evolution of Vertebrates. Genome Biol. Evol. 2020, 12, 1060–1073. [Google Scholar] [CrossRef]
- Sang, Y.; Gao, B.; Diaby, M.; Zong, W.; Chen, C.; Shen, D.; Wang, S.; Wang, Y.; Ivics, Z.; Song, C. Incomer, a DD36E Family of Tc1/mariner Transposons Newly Discovered in Animals. Mob. DNA 2019, 10, 45. [Google Scholar] [CrossRef]
- Muñoz-López, M.; Garcia-Pérez, J.L. DNA Transposons: Nature and Applications in Genomics. Curr. Genom. 2010, 11, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.H.; Hermann, D.; Caruso, A.; Tastard, E.; Marchand, J.; Rouault, J.D.; Denis, F.; Thiriet-Rupert, S.; Casse, N.; Morant-Manceau, A. First Evidence of Mariner-like Transposons in the Genome of the Marine Microalga Amphora Acutiuscula (Bacillariophyta). Protist 2014, 165, 730–744. [Google Scholar] [CrossRef] [PubMed]
- Tudor, M.; Lobocka, M.; Goodell, M.; Pettitt, J.; O’Hare, K. The Pogo Transposable Element Family of Drosophila Melanogaster. MGG Mol. Gen. Genet. 1992, 232, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Smit, A.F.; Riggs, A.D. Tiggers and DNA Transposon Fossils in the Human Genome. Proc. Natl. Acad. Sci. USA 1996, 93, 1443–1448. [Google Scholar] [CrossRef]
- Liu, Y.; Zong, W.; Diaby, M.; Lin, Z.; Wang, S.; Gao, B.; Ji, T.; Song, C. Diversity and Evolution of Pogo and Tc1/mariner Transposons in the Apoidea Genomes. Biology 2021, 10, 940. [Google Scholar] [CrossRef]
- Dufresne, M.; Lespinet, O.; Daboussi, M.J.; Hua-Van, A. Genome-Wide Comparative Analysis of Pogo-like Transposable Elements in Different Fusarium Species. J. Mol. Evol. 2011, 73, 230–243. [Google Scholar] [CrossRef]
- Hey, P.; Robson, G.; Birch, M.; Bromley, M. Characterisation of Aft1 a Fot1/Pogo Type Transposon of Aspergillus Fumigatus. Fungal Genet. Biol. 2008, 45, 117–126. [Google Scholar] [CrossRef]
- Nyyssönen, E.; Amutan, M.; Enfield, L.; Stubbs, J.; Dunn-Coleman, N.S. The Transposable Element Tan1 of Aspergillus Niger Var. Awamori, a New Member of the Fot1 Family. Mol. Gen. Genet. 1996, 253, 50–56. [Google Scholar] [CrossRef]
- Levis, C.; Fortini, D.; Brygoo, Y. Flipper, a Mobile Fot1-like Transposable Element in Botrytis Cinerea. Mol. Gen. Genet. 1997, 254, 674–680. [Google Scholar] [CrossRef]
- Kachroo, P.; Leong, S.A.; Chattoo, B.B. Pot2, an Inverted Repeat Transposon from the Rice Blast Fungus Magnaporthe Grisea. MGG Mol. Gen. Genet. 1994, 245, 339–348. [Google Scholar] [CrossRef]
- Daboussi, M.-J.; Langin, T.; Brygoo, Y. Fot1, a New Family of Fungal Transposable Elements. Mol. Gen. Genet. MGG 1992, 232, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C.; Mouchès, C. Evidence That a Family of Miniature Inverted-Repeat Transposable Elements (MITEs) from the Arabidopsis Thaliana Genome Has Arisen from a Pogo- like DNA Transposon. Mol. Biol. Evol. 2000, 17, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Chen, W.; Shen, D.; Wang, S.; Chen, C.; Zhang, L.; Wang, W.; Wang, X.; Song, C. Characterization of Autonomous Families of Tc1/mariner Transposons in Neoteleost Genomes. Mar. Genom. 2017, 34, 67–77. [Google Scholar] [CrossRef]
- Gao, B.; Shen, D.; Xue, S.; Chen, C.; Cui, H.; Song, C. The Contribution of Transposable Elements to Size Variations between Four Teleost Genomes. Mob. DNA 2016, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Dupeyron, M.; Baril, T.; Bass, C.; Hayward, A. Phylogenetic Analysis of the Tc1/mariner Superfamily Reveals the Unexplored Diversity of Pogo-like Elements. Mob. DNA 2020, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.D.; Tomii, K.; Katoh, K. Application of the MAFFT Sequence Alignment Program to Large Data—reexamination of the Usefulness of Chained Guide Trees. Bioinformatics 2016, 32, 3246–3251. [Google Scholar] [CrossRef] [PubMed]
- Xia, X. DAMBE7: New and Improved Tools for Data Analysis in Molecular Biology and Evolution. Mol. Biol. Evol. 2018, 35, 1550–1552. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Harris, J.K.; Kelley, S.T.; Spiegelman, G.B.; Pace, N.R. The Genetic Core of the Universal Ancestor. Genome Res. 2003, 13, 407–412. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Gilbert, C.; Cordaux, R. Horizontal Transfer and Evolution of Prokaryote Transposable Elements in Eukaryotes. Genome Biol. Evol. 2013, 5, 822–832. [Google Scholar] [CrossRef]
- Zhang, H.-H.; Li, G.-Y.; Xiong, X.-M.; Han, M.-J.; Dai, F.-Y. Horizontal Transfer of a Novel Helentron in Insects. Mol. Genet. Genomics 2017, 292, 243–250. [Google Scholar] [CrossRef]
- Platt, R.N.; Vandewege, M.W.; Ray, D.A. Mammalian Transposable Elements and Their Impacts on Genome Evolution. Chromosom. Res. 2017, 26, 25–43. [Google Scholar] [CrossRef]
- Chalopin, D.; Naville, M.; Plard, F.; Galiana, D.; Volff, J.N. Comparative Analysis of Transposable Elements Highlights Mobilome Diversity and Evolution in Vertebrates. Genome Biol. Evol. 2015, 7, 567–580. [Google Scholar] [CrossRef]
- Sotero-Caio, C.G.; Platt, R.N.; Suh, A.; Ray, D.A. Evolution and Diversity of Transposable Elements in Vertebrate Genomes. Genome Biol. Evol. 2017, 9, 161–177. [Google Scholar] [CrossRef]
- Dupeyron, M.; Leclercq, S.; Cerveau, N.; Bouchon, D.; Gilbert, C. Horizontal Transfer of Transposons between and within Crustaceans and Insects. Mob. DNA 2014, 5, 4. [Google Scholar] [CrossRef]
- Dunemann, S.M.; Wasmuth, J.D. Horizontal Transfer of a Retrotransposon between Parasitic Nematodes and the Common Shrew. Mob. DNA 2019, 10, 24. [Google Scholar] [CrossRef]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kipling, D.; Warburton, P.E. Centromeres, CENP-B and Tigger Too. Trends Genet. 1997, 13, 141–145. [Google Scholar] [CrossRef]
- Bourgeois, Y.; Boissinot, S. On the Population Dynamics of Junk: A Review on the Population Genomics of Transposable Elements. Genes 2019, 10, 419. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.J.; O’Neill, R.J. Transposable Elements: Genome Innovation, Chromosome Diversity, and Centromere Conflict. Chromosom. Res. 2018, 26, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Mitra, R.; Li, X.; Kapusta, A.; Mayhew, D.; Mitra, R.D.; Feschotte, C.; Craig, N.L. Functional Characterization of piggyBat from the Bat Myotis Lucifugus Unveils an Active Mammalian DNA Transposon. Proc. Natl. Acad. Sci. USA 2013, 110, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Wallau, G.L.; Vieira, C.; Loreto, É.L.S. Genetic Exchange in Eukaryotes through Horizontal Transfer: Connected by the Mobilome. Mob. DNA 2018, 9, 6. [Google Scholar] [CrossRef]
- Tang, Z.; Zhang, H.-H.; Huang, K.; Zhang, X.-G.; Han, M.-J.; Zhang, Z. Repeated Horizontal Transfers of Four DNA Transposons in Invertebrates and Bats. Mob. DNA 2015, 6, 3. [Google Scholar] [CrossRef]
- Novick, P.; Smith, J.; Ray, D.; Boissinot, S. Independent and Parallel Lateral Transfer of DNA Transposons in Tetrapod Genomes. Gene 2010, 449, 85–94. [Google Scholar] [CrossRef]
- Oliveira, S.G.; Bao, W.; Martins, C.; Jurka, J. Horizontal Transfers of Mariner Transposons between Mammals and Insects. Mob. DNA 2012, 3, 14. [Google Scholar] [CrossRef]
- Lin, X.; Faridi, N.; Casola, C. An Ancient Transkingdom Horizontal Transfer of Penelope-like Retroelements from Arthropods to Conifers. Genome Biol. Evol. 2016, 8, 1252–1266. [Google Scholar]
- Suh, A.; Witt, C.C.; Menger, J.; Sadanandan, K.R.; Podsiadlowski, L.; Gerth, M.; Weigert, A.; McGuire, J.A.; Mudge, J.; Edwards, S.V.; et al. Ancient Horizontal Transfers of Retrotransposons between Birds and Ancestors of Human Pathogenic Nematodes. Nat. Commun. 2016, 7, 11396. [Google Scholar] [CrossRef]
- Combes, C. Parasitism: The Ecology and Evolution of Intimate Interactions; University of Chicago Press: Chicago, IL, USA, 2001. [Google Scholar]
- Manser, M.B.; Jansen, D.A.; Graw, B.; Hollén, L.I.; Bousquet, C.A.H.; Furrer, R.D.; Le Roux, A. Vocal Complexity in Meerkats and Other Mongoose Species. In Advances in the Study of Behavior; Elsevier: Amsterdam, The Netherlands, 2014; Volume 46, pp. 281–310. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distribution | Number of Species Containing Tigger | Number of Species Containing Full Tigger (%) a | Number of Species Containing Intact Tigger | Length of the Full Tigger (bp) b | Length of the Intact Tigger (bp) c | Transposase Length of the Intact Tigger | TIR Length of the Intact Tigger (bp) | TSD |
|---|---|---|---|---|---|---|---|---|
| Porifera | 3 | 2/66.6 | 2 | 2288–3872 | 2288–3872 | 334–685 | 20–741 | TA |
| Cnidaria | 5 | 4/80 | 2 | 1385–3309 | 1385–3210 | 374–538 | 8–24 | TA |
| Platyhelminthes | 3 | 3/100 | 3 | 2049–2762 | 2049–2762 | 360–598 | 17–25 | TA |
| Mollusca | 8 | 7/87.5 | 6 | 1609–3452 | 1609–2851 | 532–575 | 15–25 | TA |
| Nematoda | 2 | 1/50 | 1 | 2399–3400 | 3400 | 555 | 23–24 | TA |
| Echinodermata | 2 | 1/50 | 1 | 2988–3068 | 3068 | 540 | 21–23 | TA |
| Urochordata | 1 | 1/100 | 1 | 2381 | 2381 | 519 | 22 | TA |
| Arthropods | 183 | 159/87 | 147 | 1018–4225 | 1428–4225 | 301–607 | 12–33 | TA |
| Annelida | 1 | 1/100 | 1 | 1969 | 1969 | 396 | 24 | TA |
| Cephalochordata | 1 | 1/100 | 1 | 2872 | 2872 | 305 | 18 | TA |
| Actinopterygii | 11 | 10/91 | 9 | 1385–2821 | 1385–2821 | 301–438 | 13–23 | TA |
| Agnatha | 1 | 1/100 | 1 | 2906 | 2906 | 580 | 21 | TA |
| Sarcopterygii | 1 | 1/100 | 1 | 2099–2418 | 2099–2418 | 457–582 | 23 | TA |
| Chondrichthyes | 5 | 4/80 | 4 | 2094–4525 | 2094–4525 | 306–585 | 18–23 | TA |
| Anura | 3 | 2/66.6 | 2 | 1620–2277 | 1620–2277 | 340–597 | 13–29 | TA |
| Squamata | 5 | 4/80 | 4 | 2336–3988 | 2336–3988 | 532–618 | 17–26 | TA |
| Crocodilia | 6 | 3/50 | 3 | 2329–2516 | 2345–2360 | 587–640 | 11–24 | TA |
| Aves | 1 | ND | ND | 3808 | ND | 584 | 20 | TA |
| Testudines | 24 | 21/87.5 | 21 | 1370–2740 | 1370–2740 | 323–601 | 10–27 | TA |
| Metatheria | 3 | 3/100 | 3 | 2245–2379 | 2245–2379 | 541–546 | 20–24 | TA |
| Eutheria | 100 | 46/46 | 46 | 1605–2962 | 1605–2962 | 300–636 | 8–27 | TA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diaby, M.; Guan, Z.; Shi, S.; Sang, Y.; Wang, S.; Wang, Y.; Zong, W.; Ullah, N.; Gao, B.; Song, C. Revisiting the Tigger Transposon Evolution Revealing Extensive Involvement in the Shaping of Mammal Genomes. Biology 2022, 11, 921. https://doi.org/10.3390/biology11060921
Diaby M, Guan Z, Shi S, Sang Y, Wang S, Wang Y, Zong W, Ullah N, Gao B, Song C. Revisiting the Tigger Transposon Evolution Revealing Extensive Involvement in the Shaping of Mammal Genomes. Biology. 2022; 11(6):921. https://doi.org/10.3390/biology11060921
Chicago/Turabian StyleDiaby, Mohamed, Zhongxia Guan, Shasha Shi, Yatong Sang, Saisai Wang, Yali Wang, Wencheng Zong, Numan Ullah, Bo Gao, and Chengyi Song. 2022. "Revisiting the Tigger Transposon Evolution Revealing Extensive Involvement in the Shaping of Mammal Genomes" Biology 11, no. 6: 921. https://doi.org/10.3390/biology11060921
APA StyleDiaby, M., Guan, Z., Shi, S., Sang, Y., Wang, S., Wang, Y., Zong, W., Ullah, N., Gao, B., & Song, C. (2022). Revisiting the Tigger Transposon Evolution Revealing Extensive Involvement in the Shaping of Mammal Genomes. Biology, 11(6), 921. https://doi.org/10.3390/biology11060921