
An Environmentally Friendly Approach for the Release of Essential Fatty Acids from Cereal By-Products Using Cellulose-Degrading Enzymes



Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Plant Material Preparation for Alkaline and Enzymatic Hydrolysis and Analysis of Hydroxycinnamates
2.3. Chemicals and Reagents
2.4. Enzymes
2.5. Soxhlet Extraction
2.6. Preparation of the Lipid Fraction by Enzyme-Assisted Hydrolysis of Wheat, Rye, and Oat Bran with Subsequent Liquid–Liquid Extraction
2.7. Preparation of the Lipid Fraction by Alkaline-Assisted Hydrolysis of Wheat, Rye, and Oat Bran with Subsequent Liquid–Liquid Extraction
2.8. Preparation of Fatty Acids for GC/MS Analysis Using 0.5 M TMPAH Solution
2.9. The GC Conditions for FAME Analysis
2.10. The MS Conditions for FAME Detection
2.11. The HPLC-RID Conditions for Carbohydrates Analysis
2.12. Scanning Electron Microscopy (SEM)
2.13. Statistical Analysis
3. Results and Discussion
3.1. Structural Changes in Wheat, Rye, and Oat Bran Morphology Induced by Cellulose-Degrading Enzymes
3.2. Release of Mono- and Disaccharides from Wheat, Rye, and Oat Bran Using Enzyme-Assisted Hydrolysis
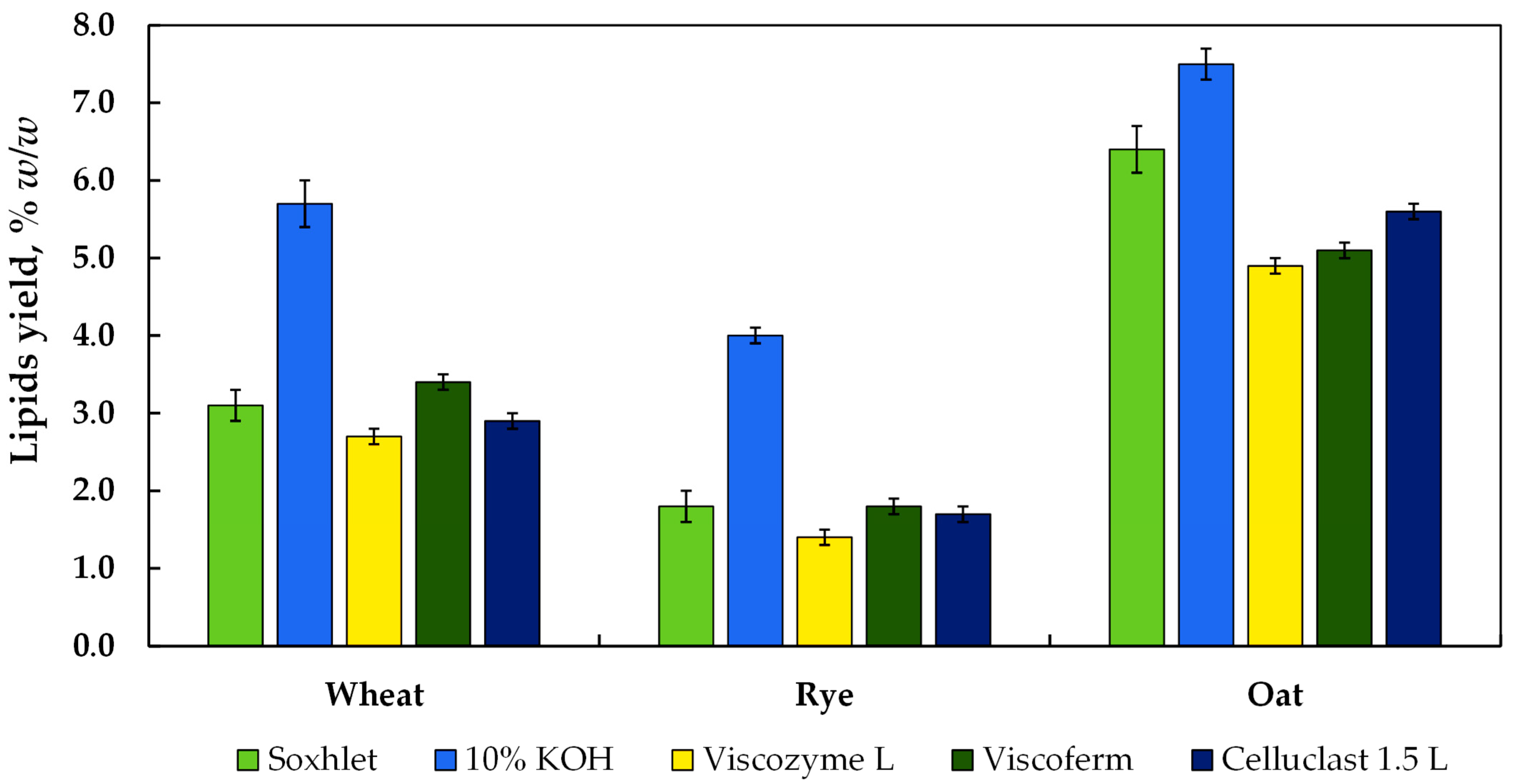
3.3. Effect of Alkaline- and Enzyme-Assisted Hydrolysis on Recovery of Lipids from Wheat, Rye, and Oat Bran
3.4. Effect of Alkaline- and Enzyme-Assisted Hydrolysis on Recovery of Fatty Acids from Wheat, Rye, and Oat Bran
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adamberg, S.; Sumeri, I.; Uusna, R.; Ambalam, P.; Kondepudi, K.K.; Adamberg, K.; Wadström, T.; Ljungh, Å. Survival and Synergistic Growth of Mixed Cultures of Bifidobacteria and Lactobacilli Combined with Prebiotic Oligosaccharides in a Gastrointestinal Tract Simulator. Microb. Ecol. Health Dis. 2014, 25, 23062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radenkovs, V.; Juhnevica-Radenkova, K.; Górnaś, P.; Seglina, D. Non-Waste Technology through the Enzymatic Hydrolysis of Agro-Industrial by-Products. Trends Food Sci. Technol. 2018, 77, 64–76. [Google Scholar] [CrossRef]
- Heiniö, R.L.; Noort, M.W.J.; Katina, K.; Alam, S.A.; Sozer, N.; de Kock, H.L.; Hersleth, M.; Poutanen, K. Sensory Characteristics of Wholegrain and Bran-Rich Cereal Foods—A Review. Trends Food Sci. Technol. 2016, 47, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Onipe, O.O.; Ramashia, S.E.; Jideani, A.I.O. Wheat Bran Modifications for Enhanced Nutrition and Functionality in Selected Food Products. Molecules 2021, 26, 3918. [Google Scholar] [CrossRef]
- Jefremova, O.; Radenkovs, V.; Kunkulberga, D.; Klava, D. Technological Properties of Dough from Wheat Flour and Fermented Bran. Chem. Technol. 2015, 66, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Aravind, N.; Sissons, M.; Egan, N.; Fellows, C. Effect of Insoluble Dietary Fibre Addition on Technological, Sensory, and Structural Properties of Durum Wheat Spaghetti. Food Chem. 2012, 130, 299–309. [Google Scholar] [CrossRef]
- Lebesi, D.M.; Tzia, C. Effect of the Addition of Different Dietary Fiber and Edible Cereal Bran Sources on the Baking and Sensory Characteristics of Cupcakes. Food Bioprocess Technol. 2011, 4, 710–722. [Google Scholar] [CrossRef]
- Grigor, J.M.; Brennan, C.S.; Hutchings, S.C.; Rowlands, D.S. The Sensory Acceptance of Fibre-Enriched Cereal Foods: A Meta-Analysis. Int. J. Food Sci. Technol. 2016, 51, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Seal, C.J.; Courtin, C.M.; Venema, K.; de Vries, J. Health Benefits of Whole Grain: Effects on Dietary Carbohydrate Quality, the Gut Microbiome, and Consequences of Processing. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2742–2768. [Google Scholar] [CrossRef]
- Kirwan, J.P.; Malin, S.K.; Scelsi, A.R.; Kullman, E.L.; Navaneethan, S.D.; Pagadala, M.R.; Haus, J.M.; Filion, J.; Godin, J.P.; Kochhar, S.; et al. A Whole-Grain Diet Reduces Cardiovascular Risk Factors in Overweight and Obese Adults: A Randomized Controlled Trial. J. Nutr. 2016, 146, 2244–2251. [Google Scholar] [CrossRef]
- Philip Karl, J.; Meydani, M.; Barnett, J.B.; Vanegas, S.M.; Goldin, B.; Kane, A.; Rasmussen, H.; Saltzman, E.; Vangay, P.; Knights, D.; et al. Substituting Whole Grains for Refined Grains in a 6-Wk Randomized Trial Favorably Affects Energy-Balance Metrics in Healthy Men and Postmenopausal Women1-3. Am. J. Clin. Nutr. 2017, 105, 589–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.S.; Qi, L.; Fahey, G.C.; Klurfeld, D.M. Consumption of Cereal Fiber, Mixtures of Whole Grains and Bran, and Whole Grains and Risk Reduction in Type 2 Diabetes, Obesity, and Cardiovascular Disease. Am. J. Clin. Nutr. 2013, 98, 594–619. [Google Scholar] [CrossRef] [PubMed]
- Zamaratskaia, G.; Mhd Omar, N.A.; Brunius, C.; Hallmans, G.; Johansson, J.E.; Andersson, S.O.; Larsson, A.; Åman, P.; Landberg, R. Consumption of Whole Grain/Bran Rye Instead of Refined Wheat Decrease Concentrations of TNF-R2, e-Selectin, and Endostatin in an Exploratory Study in Men with Prostate Cancer. Clin. Nutr. 2020, 39, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Barati, Z.; Iravani, M.; Karandish, M.; Haghighizadeh, M.H.; Masihi, S. The Effect of Oat Bran Consumption on Gestational Diabetes: A Randomized Controlled Clinical Trial. BMC Endocr. Disord. 2021, 21, 67. [Google Scholar] [CrossRef]
- Jensen, M.K.; Koh-Banerjee, P.; Franz, M.; Sampson, L.; Grønbæk, M.; Rimm, E.B. Whole Grains, Bran, and Germ in Relation to Homocysteine and Markers of Glycemic Control, Lipids, and Inflammation. Am. J. Clin. Nutr. 2006, 83, 275–283. [Google Scholar] [CrossRef]
- Pihlajaniemi, V.; Mattila, O.; Koitto, T.; Nikinmaa, M.; Heiniö, R.L.; Sorsamäki, L.; Siika-aho, M.; Nordlund, E. Production of Syrup Rich in Arabinoxylan Oligomers and Antioxidants from Wheat Bran by Alkaline Pretreatment and Enzymatic Hydrolysis, and Applicability in Baking. J. Cereal Sci. 2020, 95, 103043. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, Y.; Zhang, M.; Wang, D. Integrating Bran Starch Hydrolysates with Alkaline Pretreated Soft Wheat Bran to Boost Sugar Concentration. Bioresour. Technol. 2020, 302, 122826. [Google Scholar] [CrossRef]
- Gómez, M.; Jiménez, S.; Ruiz, E.; Oliete, B. Effect of Extruded Wheat Bran on Dough Rheology and Bread Quality. LWT Food Sci. Technol. 2011, 44, 2231–2237. [Google Scholar] [CrossRef]
- Sui, W.; Xie, X.; Liu, R.; Wu, T.; Zhang, M. Effect of Wheat Bran Modification by Steam Explosion on Structural Characteristics and Rheological Properties of Wheat Flour Dough. Food Hydrocoll. 2018, 84, 571–580. [Google Scholar] [CrossRef]
- Kumar, A.K.; Sharma, S. Recent Updates on Different Methods of Pretreatment of Lignocellulosic Feedstocks: A Review. Bioresour. Bioprocess. 2017, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Radenkovs, V.; Juhnevica-radenkova, K.; Kviesis, J.; Lazdina, D.; Valdovska, A.; Vallejo, F.; Lacis, G. Lignocellulose-Degrading Enzymes : A Biotechnology Platform for Ferulic Acid Production from Agro-Industrial Side Streams. Foods 2021, 10, 3056. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; Liao, A.M.; Thakur, K.; Huang, J.H.; Zhang, J.G.; Wei, Z.J. Modification of Wheat Bran Insoluble Dietary Fiber with Carboxymethylation, Complex Enzymatic Hydrolysis and Ultrafine Comminution. Food Chem. 2019, 297, 124983. [Google Scholar] [CrossRef] [PubMed]
- Bertacchi, S.; Pagliari, S.; Cantù, C.; Bruni, I.; Labra, M.; Branduardi, P. Enzymatic Hydrolysate of Cinnamon Waste Material as Feedstock for the Microbial Production of Carotenoids. Int. J. Environ. Res. Public Health 2021, 18, 1146. [Google Scholar] [CrossRef] [PubMed]
- Călinoiu, L.F.; Vodnar, D.C. Thermal Processing for the Release of Phenolic Compounds from Wheat and Oat Bran. Biomolecules 2020, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Iftikhar, M.; Zhang, H.; Iftikhar, A.; Raza, A.; Khan, M.; Sui, M.; Wang, J. Comparative Assessment of Functional Properties, Free and Bound Phenolic Profile, Antioxidant Activity, and in Vitro Bioaccessibility of Rye Bran and Its Insoluble Dietary Fiber. J. Food Biochem. 2020, 44, e13388. [Google Scholar] [CrossRef]
- Juhnevica-Radenkova, K.; Kviesis, J.; Moreno, D.A.; Seglina, D.; Vallejo, F.; Valdovska, A.; Radenkovs, V. Highly-Efficient Release of Ferulic Acid from Agro-Industrial by-Products via Enzymatic Hydrolysis with Cellulose-Degrading Enzymes: Part I–The Superiority of Hydrolytic Enzymes versus Conventional Hydrolysis. Foods 2021, 10, 782. [Google Scholar] [CrossRef]
- Gopalan, N.; Nampoothiri, K.M. Biorefining of Wheat Bran for the Purification of Ferulic Acid. Biocatal. Agric. Biotechnol. 2018, 15, 304–310. [Google Scholar] [CrossRef]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary Fibre in Foods: A Review. J. Food Sci. Technol. 2012, 49, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Maina, N.H.; Rieder, A.; De Bondt, Y.; Mäkelä-Salmi, N.; Sahlstrøm, S.; Mattila, O.; Lamothe, L.M.; Nyström, L.; Courtin, C.M.; Katina, K.; et al. Process-Induced Changes in the Quantity and Characteristics of Grain Dietary Fiber. Foods 2021, 10, 2566. [Google Scholar] [CrossRef]
- Wang, T.H.; Lu, S. Production of Xylooligosaccharide from Wheat Bran by Microwave Assisted Enzymatic Hydrolysis. Food Chem. 2013, 138, 1531–1535. [Google Scholar] [CrossRef]
- Siepmann, F.B.; Canan, C.; de Jesus, M.M.M.; Pazuch, C.M.; Colla, E. Release Optimization of Fermentable Sugars from Defatted Rice Bran for Bioethanol Production. Acta Sci.-Technol. 2018, 40, e35000. [Google Scholar] [CrossRef]
- Chen, Y.; Xie, Y.; Ajuwon, K.M.; Zhong, R.; Li, T.; Chen, L.; Zhang, H.; Beckers, Y.; Everaert, N. Xylo-Oligosaccharides, Preparation and Application to Human and Animal Health: A Review. Front. Nutr. 2021, 8, 638. [Google Scholar] [CrossRef] [PubMed]
- Di Gioia, D.; Sciubba, L.; Setti, L.; Luziatelli, F.; Ruzzi, M.; Zanichelli, D.; Fava, F. Production of Biovanillin from Wheat Bran. Enzyme Microb. Technol. 2007, 41, 498–505. [Google Scholar] [CrossRef]
- Wang, G.; Huang, D.; Li, Y.; Wen, J.; Jia, X. A Metabolic-Based Approach to Improve Xylose Utilization for Fumaric Acid Production from Acid Pretreated Wheat Bran by Rhizopus Oryzae. Bioresour. Technol. 2015, 180, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Kaseke, T.; Opara, U.L.; Olaniyi, A.F. Effects of Enzymatic Pretreatment of Seeds on the Physicochemical Properties, Bioactive Compounds, and Antioxidant Activity of Pomegranate Seed Oil. Molecules 2021, 26, 4575. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Z.; Cai, Z.Q. Optimization and Quality Analysis of Different Extraction Methods of Palm Seed Oil. Complexity 2021, 2021, 9925974. [Google Scholar] [CrossRef]
- Jia, Y.; Kumar, D.; Winkler-Moser, J.K.; Dien, B.; Singh, V. Recoveries of Oil and Hydrolyzed Sugars from Corn Germ Meal by Hydrothermal Pretreatment: A Model Feedstock for Lipid-Producing Energy Crops. Energies 2020, 13, 6022. [Google Scholar] [CrossRef]
- Price, P.B.; Parsons, J. Distribution of Lipids in Embryonic Axis, Bran-Endosperm, and Hull Fractions of Hulless Barley and Hulless Oat Grain. J. Agric. Food Chem. 1979, 27, 813–815. [Google Scholar] [CrossRef]
- Hargin, K.D.; Morrison, W.R. The Distribution of Acyl Lipids in the Germ, Aleurone, Starch and Non-starch Endosperm of Four Wheat Varieties. J. Sci. Food Agric. 1980, 31, 877–888. [Google Scholar] [CrossRef]
- González-Thuillier, I.; Salt, L.; Chope, G.; Penson, S.; Skeggs, P.; Tosi, P.; Powers, S.J.; Ward, J.L.; Wilde, P.; Shewry, P.R.; et al. Distribution of Lipids in the Grain of Wheat (Cv. Hereward) Determined by Lipidomic Analysis of Milling and Pearling Fractions. J. Agric. Food Chem. 2015, 63, 10705–10716. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, R.P. Gravimetric Measurements of Water. In Handbook of Food Analytical Chemistry, Water, Proteins, Enzymes, Lipids, and Carbohydrates; Wrolstad, R.E., Acree, T.E., Decker, E.A., Penner, M.H., Reid, D.S., Schwartz, S.J., Shoemaker, C.F., Smith, D., Sporns, P., Eds.; John Wiley and Sons Inc.: Hoboken, NJ, USA, 2005; pp. 5–33. ISBN 9780471709084. [Google Scholar]
- Abdolshahi, A.; Majd, M.H.; Rad, J.S.; Taheri, M.; Shabani, A.; Teixeira da Silva, J.A. Choice of Solvent Extraction Technique Affects Fatty Acid Composition of Pistachio (Pistacia vera L.) Oil. J. Food Sci. Technol. 2015, 52, 2422–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, D.; Chen, X.; Wen, P.; Zhang, J. Insight into the Role of α-Arabinofuranosidase in Biomass Hydrolysis: Cellulose Digestibility and Inhibition by Xylooligomers. Biotechnol. Biofuels 2019, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Taherzadeh, M.J.; Niklasson, C. Ethanol from Lignocellulosic Materials: Pretreatment, Acid and Enzymatic Hydrolyses, and Fermentation. In Lignocellulose Bodegradation; Saha, B.C., Kyoshi, H., Eds.; American Chemical Society: Washington, DC, USA, 2004; pp. 49–68. [Google Scholar]
- Bautista-Expósito, S.; Tomé-Sánchez, I.; Martín-Diana, A.B.; Frias, J.; Peñas, E.; Rico, D.; Casas, M.J.G.; Martínez-Villaluenga, C. Enzyme Selection and Hydrolysis under Optimal Conditions Improved Phenolic Acid Solubility, and Antioxidant and Anti-inflammatory Activities of Wheat Bran. Antioxidants 2020, 9, 984. [Google Scholar] [CrossRef] [PubMed]
- Gama, R.; Van Dyk, J.S.; Pletschke, B.I. Optimisation of Enzymatic Hydrolysis of Apple Pomace for Production of Biofuel and Biorefinery Chemicals Using Commercial Enzymes. 3 Biotech 2015, 5, 1075–1087. [Google Scholar] [CrossRef] [Green Version]
- Lovegrove, A.; Edwards, C.H.; De Noni, I.; Patel, H.; El, S.N.; Grassby, T.; Zielke, C.; Ulmius, M.; Nilsson, L.; Butterworth, P.J.; et al. Role of Polysaccharides in Food, Digestion, and Health. Crit. Rev. Food Sci. Nutr. 2017, 57, 237–253. [Google Scholar] [CrossRef] [Green Version]
- Alvira, P.; Tomás-Pejó, E.; Ballesteros, M.; Negro, M.J. Pretreatment Technologies for an Efficient Bioethanol Production Process Based on Enzymatic Hydrolysis: A Review. Bioresour. Technol. 2010, 101, 4851–4861. [Google Scholar] [CrossRef]
- Kamal-Eldin, A.; Lærke, H.N.; Knudsen, K.E.B.; Lampi, A.M.; Piironen, V.; Adlercreutz, H.; Katina, K.; Poutanen, K.; Åman, P. Physical, Microscopic and Chemical Characterisation of Industrial Rye and Wheat Brans from the Nordic Countries. Food Nutr. Res. 2009, 53, 1912. [Google Scholar] [CrossRef] [Green Version]
- Narducci, V.; Finotti, E.; Galli, V.; Carcea, M. Lipids and Fatty Acids in Italian Durum Wheat (Triticum durum Desf.) Cultivars. Foods 2019, 8, 223. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.S. Comparison of Lipid Content and Fatty Acid Composition and Their Distribution within Seeds of 5 Small Grain Species. J. Food Sci. 2011, 76, C334–C342. [Google Scholar] [CrossRef]
- Díaz-Suárez, P.; Rosales-Quintero, A.; Fernandez-Lafuente, R.; Pola-Sánchez, E.; Hernández-Cruz, M.C.; Ovando-Chacón, S.L.; Rodrigues, R.C.; Tacias-Pascacio, V.G. Aqueous Enzymatic Extraction of Ricinus Communis Seeds Oil Using Viscozyme L. Ind. Crops Prod. 2021, 170, 113811. [Google Scholar] [CrossRef]
- Yilmaz, I.; Daǧlioǧlu, O. The Effect of Replacing Fat with Oat Bran on Fatty Acid Composition and Physicochemical Properties of Meatballs. Meat Sci. 2003, 65, 819–823. [Google Scholar] [CrossRef]
- Rasane, P.; Jha, A.; Sabikhi, L.; Kumar, A.; Unnikrishnan, V.S. Nutritional Advantages of Oats and Opportunities for Its Processing as Value Added Foods—A Review. J. Food Sci. Technol. 2015, 52, 662–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wikiera, A.; Mika, M.; Grabacka, M. Multicatalytic Enzyme Preparations as Effective Alternative to Acid in Pectin Extraction. Food Hydrocoll. 2015, 44, 156–161. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Major Nutrient Profiles, g 100 g−1 DW | ||||||
|---|---|---|---|---|---|---|
| Type of Bran | Moisture, % | Crude Carbohydrates | Starch | Crude Lipids | Crude Proteins | Dietary Fiber |
| Wheat | 11.9 ± 0.2 a | 20.3 ± 0.4 c | 8.7 ± 0.0 c | 4.5 ± 0.1 b | 16.2 ± 0.4 a | 46.5 ± 2.1 a |
| Rye | 11.7 ± 0.2 a | 30.9 ± 0.5 b | 18.6 ± 0.0 b | 3.8 ± 0.1 c | 16.9 ± 0.5 a | 36.0 ± 1.9 b |
| Oat | 12.4 ± 0.3 a | 50.0 ± 0.9 a | 47.6 ± 0.9 a | 6.7 ± 0.5 a | 14.0 ± 0.7 b | 14.0 ± 1.7 c |
| Commercial Enzyme | Declared Activity | Enzyme Activity | Source | EC Number |
|---|---|---|---|---|
| Viscozyme® L | 100 FBG g−1 | Endo-1,3-(1,4)-β-d-glucanase, Endo-1,4-β-xylanase, Non-reducing end α-L-arabinofuranosidase | Aspergillus aculeatus | 3.2.1.6 3.2.1.8 3.2.1.55 |
| Viscoferm® | 222 FBG g−1 | Endo-1,3-(1,4)-β-d-glucanase, Endo-1,4-β-xylanase | Aspergillus spp. | 3.2.1.6 3.2.1.8 |
| Celluclast® 1.5 L | 700 EGU g−1 | Endo-1,4-β-d-glucanase | Trichoderma reesei | 3.2.1.4 |
| Wheat Bran | Rye Bran | Oat Bran | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Carbohydrate | Control | Vzym | Vferm | Cell | Control | Vzym | Vferm | Cell | Control | Vzym | Vferm | Cell |
| Xyl | 0.4 ± 0.0 d | 4.6 ± 0.0 c | 6.1 ± 0.2 b | 8.7 ± 0.1 a | 0.2 ± 0.0 d | 5.9 ± 0.2 c | 7.8 ± 0.3 b | 8.4 ± 0.1 a | n.d. | 0.8 ± 0.0 b | 1.2 ± 0.0 b | 2.1 ± 0.0 a |
| Ara | 0.5 ± 0.0 b | 2.7 ± 0.0 a | 3.2 ± 0.0 a | 2.8 ± 0.0 a | 0.6 ± 0.0 d | 6.2 ± 0.2 a | 4.9 ± 0.1 b | 1.5 ± 0.0 c | n.d. | 0.8 ± 0.0 a | 0.9 ± 0.0 a | n.d. |
| Fru | n.d. | 2.1 ± 0.0 a | 0.7 ± 0.0 b | 0.4 ± 0.0 b | 0.2 ± 0.0 c | 1.7 ± 0.0 a | 0.8 ± 0.0 b | 0.5 ± 0.0 b | n.d. | 1.6 ± 0.1 a | 0.3 ± 0.0 b | n.d. |
| Glu | n.d. | 17.9 ± 0.2 a | 7.9 ± 0.1 c | 11.3 ± 0.3 b | n.d. | 32.4 ± 0.4 a | 12.3 ± 0.2 c | 15.9 ± 0.3 b | n.d. | 47.6 ± 1.3 a | 16.1 ± 0.4 c | 18.4 ± 0.5 b |
| Suc | 1.9 ± 0.1 | n.d. | n.d. | n.d. | 2.9 ± 0.1 | n.d. | n.d. | n.d. | 0.9 ± 0.1 | n.d. | n.d. | n.d. |
| Mal | 0.3 ± 0.0 c | 0.9 ± 0.0 b | 0.9 ± 0.0 b | 3.1 ± 0.0 a | 0.2 ± 0.0 c | 1.9 ± 0.0 b | 0.7 ± 0.0 c | 2.6 ± 0.0 a | n.d. | 1.9 ± 0.0 a | 1.1 ± 0.0 b | 1.8 ± 0.0 ab |
| Tot | 3.1 ± 0.1 c | 28.2 ± 0.2 a | 18.8 ± 0.3 b | 28.6 ± 0.4 a | 4.1 ± 0.1 d | 48.6 ± 0.6 a | 26.5 ± 0.6 c | 28.9 ± 0.4 b | 0.9 ± 0.1 c | 52.9 ± 1.4 a | 19.7 ± 0.4 c | 22.9 ± 0.5 b |
| Wheat | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fatty Acid | Soxhlet | 10% KOH | Vzym | Vferm | Cell | |||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Myristate C14:0 | 1.0 | 0.0 | 0.8 | 0.1 | 1.5 | 0.0 | 1.8 | 0.2 | 0.9 | 0.0 |
| Pentadecanoate C15:0 | 0.8 | 0.0 | 1.0 | 0.1 | 0.8 | 0.0 | 0.8 | 0.0 | 0.7 | 0.0 |
| Palmitate C16:0 | 127.0 | 1.9 | 132.3 | 3.1 | 143.9 | 1.2 | 142.9 | 1.9 | 126.4 | 2.0 |
| Heptadecanoate C17:0 | 1.1 | 0.2 | 1.4 | 0.1 | 1.4 | 0.0 | 1.4 | 0.0 | 1.1 | 0.0 |
| Stearate C18:0 | 8.5 | 0.3 | 8.6 | 0.3 | 9.6 | 0.1 | 9.7 | 0.0 | 8.5 | 0.0 |
| Oleate C18:1 (c9) | 104.5 | 1.1 | 106.2 | 2.5 | 121.3 | 2.3 | 117.2 | 0.9 | 104.8 | 1.9 |
| Vaccenate C18:1 (t11) | 2.6 | 0.4 | 2.3 | 0.0 | 2.8 | 0.2 | 3.0 | 0.0 | 2.0 | 0.1 |
| Linoleate C18:2 (c9,c12) | 365.8 | 0.0 | 379.9 | 7.3 | 408.6 | 6.1 | 417.9 | 4.2 | 377.6 | 10.8 |
| Linolenate C18:3 (c9,c12,c15) | 22.0 | 0.9 | 21.9 | 0.4 | 24.9 | 0.6 | 25.7 | 0.5 | 23.6 | 0.8 |
| Arachidate C20:0 | 2.1 | 0.0 | 1.6 | 0.1 | 1.2 | 0.2 | 1.1 | 0.2 | 0.9 | 0.0 |
| CLA linoleate C18:2 (c9,t11) | 0.4 | 0.0 | 2.7 | 0.1 | 0.8 | 0.1 | 1.1 | 0.0 | 0.6 | 0.0 |
| CLA linoleate C18:2 (t10,c12) | 0.4 | 0.1 | 1.9 | 0.0 | 0.5 | 0.0 | 0.8 | 0.0 | 0.4 | 0.0 |
| 11-eicosenoate C20:1 (c11) | 3.3 | 0.3 | 3.1 | 0.1 | 3.7 | 0.0 | 3.7 | 0.1 | 3.3 | 0.1 |
| Eicosadienoate C20:2 (c11,c14) | BLQ | − | 0.5 | 0.1 | 0.5 | 0.1 | 0.6 | 0.1 | 0.5 | 0.0 |
| Behenate C22:0 | 3.1 | 0.7 | 3.3 | 0.2 | 2.2 | 0.2 | 2.0 | 0.1 | 1.7 | 0.1 |
| Erucate C22:1 (c9,c11) | 0.1 | 0.1 | 0.1 | 0.0 | 0.1 | 0.0 | 0.1 | 0.0 | 0.1 | 0.0 |
| Tricosanoate C23:0 | 1.2 | 0.2 | 10.1 | 1.2 | 1.0 | 0.2 | 0.5 | 0.0 | 0.3 | 0.1 |
| Tetracosanoate C24:0 | 0.8 | 0.2 | 1.1 | 0.1 | 0.8 | 0.1 | 0.6 | 0.0 | 1.3 | 0.1 |
| SFA | 145.6 | 3.6 | 160.3 | 5.2 | 162.5 | 2.1 | 160.8 | 2.5 | 141.7 | 2.4 |
| MUFA | 110.6 | 1.8 | 111.7 | 2.6 | 127.9 | 2.5 | 124.0 | 1.0 | 110.3 | 2.1 |
| PUFA | 388.6 | 1.0 | 406.9 | 7.9 | 435.4 | 6.9 | 446.1 | 4.9 | 402.7 | 11.7 |
| ∑MUFA + PUFA | 499.2 | 2.9 | 518.6 | 10.5 | 563.3 | 9.4 | 570.1 | 5.9 | 513.0 | 13.7 |
| ∑SFA + MUFA + PUFA | 644.8 | 6.5 | 678.9 | 15.7 | 725.8 | 11.5 | 730.9 | 8.4 | 654.7 | 16.1 |
| CLA | 0.8 | 0.1 | 4.6 | 0.1 | 1.4 | 0.1 | 1.9 | 0.0 | 1.0 | 0.0 |
| Rye | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fatty Acid | Soxhlet | 10% KOH | Vzym | Vferm | Cell | |||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Myristate C14:0 | 1.3 | 0.0 | 0.9 | 0.0 | 1.4 | 0.1 | 0.9 | 0.0 | 1.0 | 0.1 |
| Pentadecanoate C15:0 | 0.8 | 0.0 | 1.2 | 0.0 | 0.8 | 0.1 | 0.9 | 0.1 | 0.8 | 0.1 |
| Palmitate C16:0 | 90.2 | 1.5 | 125.5 | 1.3 | 117.1 | 0.6 | 114.0 | 2.0 | 121.5 | 3.3 |
| Heptadecanoate C17:0 | 1.2 | 0.1 | 1.5 | 0.1 | 1.3 | 0.0 | 1.4 | 0.0 | 1.1 | 0.0 |
| Stearate C18:0 | 6.9 | 0.2 | 7.2 | 0.2 | 6.8 | 0.3 | 6.6 | 0.0 | 7.5 | 0.4 |
| Oleate C18:1 (c9) | 64.2 | 1.4 | 76.5 | 1.1 | 71.2 | 0.1 | 72.8 | 0.5 | 87.2 | 3.3 |
| Vaccenate C18:1 (t11) | 1.7 | 0.0 | 2.3 | 0.1 | 2.0 | 0.2 | 2.0 | 0.3 | 2.3 | 0.2 |
| Linoleate C18:2 (c9,c12) | 282.7 | 6.4 | 380.6 | 9.1 | 341.4 | 4.6 | 357.1 | 5.5 | 421.4 | 8.8 |
| Linolenate C18:3 (c9,c12,c15) | 21.6 | 0.5 | 27.2 | 0.7 | 25.0 | 0.2 | 26.5 | 0.6 | 33.9 | 0.9 |
| Arachidate C20:0 | 1.4 | 0.1 | 1.7 | 0.1 | 0.7 | 0.0 | 0.8 | 0.1 | 0.8 | 0.1 |
| CLA linoleate C18:2 (c9,t11) | 0.1 | 0.0 | 2.1 | 0.1 | 0.6 | 0.0 | 1.7 | 0.1 | 0.9 | 0.1 |
| CLA linoleate C18:2 (t10,c12) | 0.2 | 0.0 | 1.6 | 0.0 | 0.5 | 0.0 | 1.1 | 0.0 | 0.5 | 0.0 |
| 11-eicosenoate C20:1 (c11) | 3.9 | 0.3 | 4.3 | 0.1 | 4.0 | 0.1 | 4.1 | 0.2 | 5.2 | 0.1 |
| Eicosadienoate C20:2 (c11,c14) | BLQ | − | 0.8 | 0.1 | 0.5 | 0.1 | 0.8 | 0.0 | 0.8 | 0.0 |
| Behenate C22:0 | 2.7 | 0.1 | 2.9 | 0.1 | 2.0 | 0.1 | 1.8 | 0.0 | 1.6 | 0.1 |
| Erucate C22:1 (c9,c11) | 0.4 | 0.0 | 0.4 | 0.0 | 0.3 | 0.0 | 0.4 | 0.0 | 0.5 | 0.0 |
| Tricosanoate C23:0 | 0.9 | 0.1 | 12.9 | 1.1 | 0.7 | 0.0 | 0.4 | 0.0 | 0.3 | 0.0 |
| Tetracosanoate C24:0 | 1.1 | 0.1 | 1.1 | 0.1 | 0.7 | 0.1 | 0.6 | 0.1 | 4.6 | 0.0 |
| SFA | 106.4 | 2.2 | 154.9 | 3.1 | 131.6 | 1.2 | 127.2 | 2.3 | 139.2 | 4.1 |
| MUFA | 70.3 | 1.8 | 83.5 | 1.4 | 77.6 | 0.4 | 79.3 | 1.0 | 95.2 | 3.6 |
| PUFA | 304.6 | 6.9 | 412.3 | 9.9 | 368.1 | 4.9 | 387.2 | 6.2 | 457.3 | 9.9 |
| ∑MUFA + PUFA | 374.9 | 8.6 | 495.8 | 0.0 | 445.7 | 5.4 | 466.5 | 7.2 | 552.5 | 13.5 |
| ∑SFA + MUFA + PUFA | 481.3 | 10.9 | 650.7 | 14.3 | 577.3 | 6.5 | 593.7 | 9.5 | 691.7 | 17.6 |
| CLA | 0.3 | 0.0 | 3.7 | 0.1 | 1.2 | 0.0 | 2.9 | 0.1 | 1.3 | 0.1 |
| Oat | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fatty Acid | Soxhlet | 10% KOH | Vzym | Vferm | Cell | |||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Myristate C14:0 | 2.6 | 0.0 | 1.8 | 0.0 | 2.3 | 0.1 | 2.1 | 0.1 | 1.4 | 0.1 |
| Pentadecanoate C15:0 | BLQ | − | 0.2 | 0.0 | BLQ | − | BLQ | − | BLQ | − |
| Palmitate C16:0 | 150.5 | 0.1 | 149.0 | 0.7 | 144.4 | 0.4 | 137.0 | 2.4 | 143.1 | 0.5 |
| Heptadecanoate C17:0 | 1.0 | 0.0 | 1.0 | 0.0 | 0.7 | 0.0 | 1.0 | 0.1 | 1.0 | 0.0 |
| Stearate C18:0 | 16.6 | 0.2 | 14.4 | 0.2 | 14.0 | 0.2 | 12.4 | 0.2 | 15.8 | 0.1 |
| Oleate C18:1 (c9) | 343.9 | 4.8 | 300.0 | 1.0 | 270.6 | 1.8 | 266.6 | 1.3 | 332.6 | 0.8 |
| Vaccenate C18:1 (t11) | 4.2 | 0.1 | 4.7 | 0.1 | 3.7 | 0.0 | 2.4 | 0.3 | 3.2 | 0.4 |
| Linoleate C18:2 (c9,c12) | 323.7 | 4.5 | 321.4 | 2.9 | 272.9 | 1.0 | 272.0 | 1.3 | 314.1 | 3.0 |
| Linolenate C18:3 (c9,c12,c15) | 7.7 | 0.2 | 7.4 | 0.2 | 6.1 | 0.0 | 6.2 | 0.2 | 7.3 | 0.0 |
| Arachidate C20:0 | 0.9 | 0.8 | 0.7 | 0.1 | 0.5 | 0.0 | 0.7 | 0.1 | 0.7 | 0.1 |
| CLA linoleate C18:2 (c9,t11) | 0.4 | 0.0 | 1.7 | 0.1 | 0.3 | 0.0 | 0.9 | 0.0 | 0.3 | 0.0 |
| CLA linoleate C18:2 (t10,c12) | 0.1 | 0.0 | 1.1 | 0.1 | 0.3 | 0.0 | 0.5 | 0.0 | 0.2 | 0.0 |
| 11-eicosenoate C20:1 (c11) | 5.7 | 0.1 | 4.7 | 0.0 | 3.9 | 0.0 | 4.4 | 0.1 | 5.7 | 0.0 |
| Eicosadienoate C20:2 (c11,c14) | BLQ | − | BLQ | − | BLQ | − | 0.1 | 0.0 | 0.1 | 0.0 |
| Behenate C22:0 | 1.7 | 0.2 | 1.3 | 1.7 | 1.6 | 0.1 | 1.3 | 0.1 | 1.3 | 0.0 |
| Erucate C22:1 (c9,c11) | 0.2 | 0.0 | 0.1 | 0.0 | 0.1 | 0.0 | 0.1 | 0.0 | 0.1 | 0.0 |
| Tricosanoate C23:0 | 1.0 | 0.1 | 1.6 | 0.7 | 0.7 | 0.2 | 0.7 | 0.0 | 0.6 | 0.0 |
| Tetracosanoate C24:0 | 0.5 | 0.0 | 0.4 | 0.0 | 0.4 | 0.0 | 0.3 | 0.0 | 0.4 | 0.0 |
| SFA | 174.8 | 1.4 | 170.5 | 3.7 | 164.6 | 1.0 | 155.5 | 2.9 | 164.3 | 0.9 |
| MUFA | 353.9 | 5.1 | 309.4 | 1.1 | 278.4 | 1.9 | 273.5 | 1.7 | 341.7 | 1.2 |
| PUFA | 331.8 | 4.7 | 331.7 | 3.3 | 279.6 | 1.1 | 279.7 | 1.6 | 322.0 | 3.1 |
| ∑MUFA + PUFA | 685.7 | 9.8 | 641.1 | 4.4 | 558.0 | 3.0 | 553.2 | 3.3 | 663.6 | 4.3 |
| ∑SFA + MUFA + PUFA | 860.6 | 11.2 | 811.6 | 8.1 | 722.6 | 4.0 | 708.7 | 6.2 | 828.0 | 5.2 |
| CLA | 0.4 | 0.0 | 2.8 | 0.2 | 0.6 | 0.0 | 1.3 | 0.1 | 0.4 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radenkovs, V.; Juhnevica-Radenkova, K.; Kviesis, J.; Valdovska, A. An Environmentally Friendly Approach for the Release of Essential Fatty Acids from Cereal By-Products Using Cellulose-Degrading Enzymes. Biology 2022, 11, 721. https://doi.org/10.3390/biology11050721
Radenkovs V, Juhnevica-Radenkova K, Kviesis J, Valdovska A. An Environmentally Friendly Approach for the Release of Essential Fatty Acids from Cereal By-Products Using Cellulose-Degrading Enzymes. Biology. 2022; 11(5):721. https://doi.org/10.3390/biology11050721
Chicago/Turabian StyleRadenkovs, Vitalijs, Karina Juhnevica-Radenkova, Jorens Kviesis, and Anda Valdovska. 2022. "An Environmentally Friendly Approach for the Release of Essential Fatty Acids from Cereal By-Products Using Cellulose-Degrading Enzymes" Biology 11, no. 5: 721. https://doi.org/10.3390/biology11050721
APA StyleRadenkovs, V., Juhnevica-Radenkova, K., Kviesis, J., & Valdovska, A. (2022). An Environmentally Friendly Approach for the Release of Essential Fatty Acids from Cereal By-Products Using Cellulose-Degrading Enzymes. Biology, 11(5), 721. https://doi.org/10.3390/biology11050721