
Hibernation with Rhythmicity in the Retina, Brain, and Plasma but Not in the Liver of Hibernating Giant Spiny Frogs (Quasipaa spinosa)




Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Tissue Sampling
2.2. Molecular Cloning
2.3. Quantitative Real-Time PCR
2.4. Measurements of Plasma MT and CORT Concentrations
2.5. Statistical Analyses
3. Results
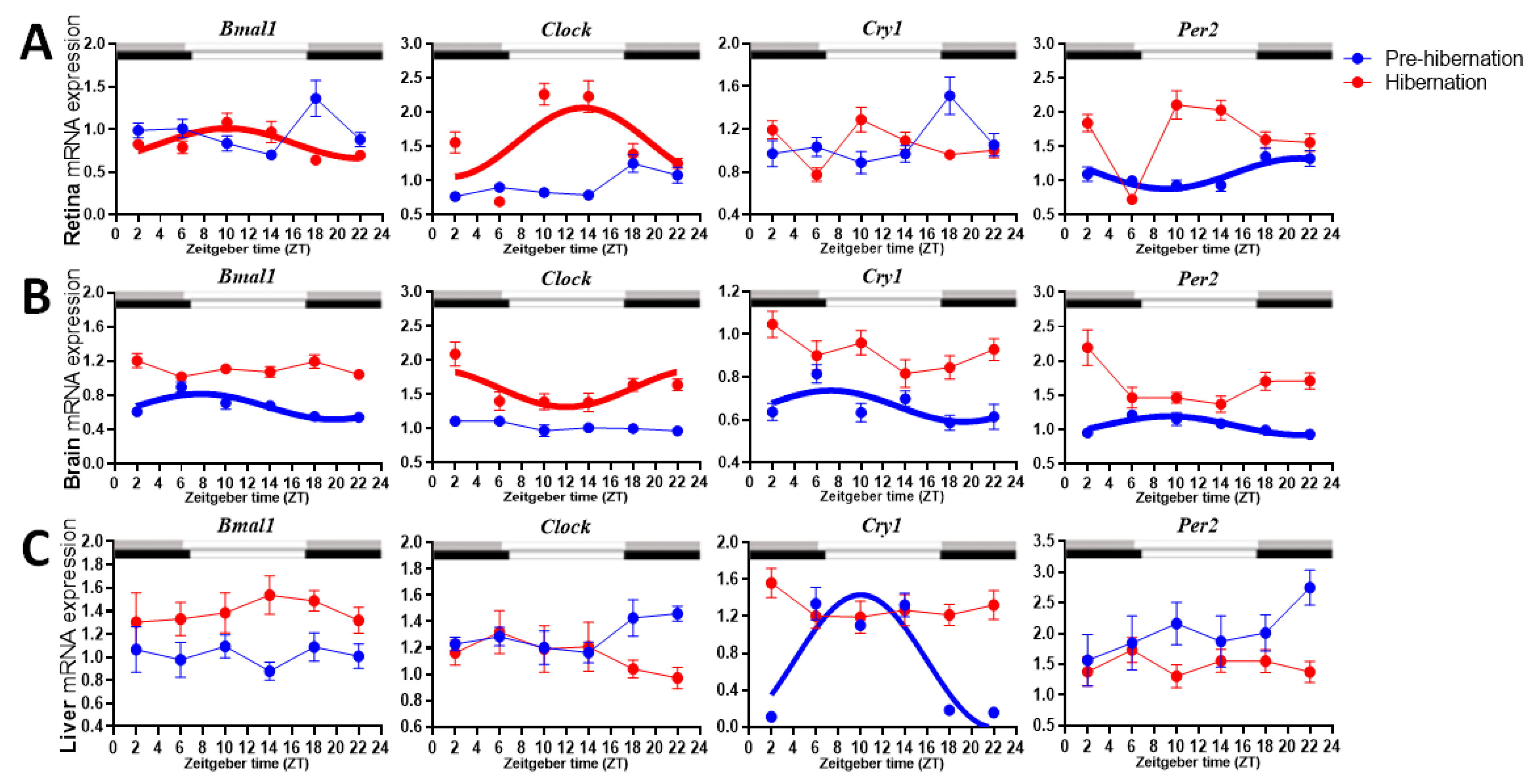
3.1. Rhythmicity of the Clock Genes
3.2. Rhythmicity of Ror-α, AANAT, and Mel-1c Genes
3.3. Rhythmicity of Plasma MT and CORT Levels
3.4. Rhythmicity of the AOEs
4. Discussion
4.1. Rhythmicity of Clock Genes, Clock-Controlled Genes, CORT, and MT
4.2. Rhythmicity of Antioxidant Enzyme Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, V. Biological Rhythms; Springer: Berlin/Heidelberg, Germany, 2002; pp. 99–117. [Google Scholar]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Miller, N. The American toad (Bufo Lentiginosus Americanus, LeConte). II A Study in dynamic biology. Am. Nat. 1909, 43, 730–745. [Google Scholar] [CrossRef][Green Version]
- Ultsch, G.R.; Reese, S.A.; Stewart, E.R. Physiology of hibernation in Rana pipiens: Metabolic rate, critical oxygen tension, and the effects of hypoxia on several plasma variables. J. Exp. Zool. 2004, 301, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Revel, F.G.; Herwig, A.; Garidou, M.-L.; Dardente, H.; Menet, J.S.; Masson-Pévet, M.; Simonneaux, V.; Saboureau, M.; Pévet, P. The circadian clock stops ticking during deep hibernation in the European hamster. Proc. Natl. Acad. Sci. USA 2007, 104, 13816–13820. [Google Scholar] [CrossRef]
- Gautier, C.; Bothorel, B.; Ciocca, D.; Valour, D.; Gaudeau, A.; Dupré, C.; Lizzo, G.; Brasseur, C.; Riest-Fery, I.; Stephan, J.-P.; et al. Gene expression profiling during hibernation in the European hamster. Sci. Rep. 2018, 8, 13167. [Google Scholar] [CrossRef]
- Borah, B.K.; Renthlei, Z.; Trivedi, A.K. Hypothalamus but not liver retains daily expression of clock genes during hibernation in terai tree frog (Polypedates teraiensis). Chronobiol. Int. 2020, 37, 485–492. [Google Scholar] [CrossRef]
- Xie, Z.; Ahmad, I.M.; Zuo, L.; Xiao, F.; Wang, Y.; Li, D. Hibernation with rhythmicity: The circadian clock and hormonal adaptations of the hibernating Asiatic toads (Bufo gargarizans). Integr. Zool. 2021. [Google Scholar] [CrossRef]
- Withers, P.; Cooper, C.E. Dormancy. Encycl. Ecol. 2019, 3, 309–314. [Google Scholar] [CrossRef]
- Cox, K.H.; Takahashi, J.S. Circadian clock genes and the transcriptional architecture of the clock mechanism. J. Mol. Endocrinol. 2019, 63, R93–R102. [Google Scholar] [CrossRef]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef]
- Akashi, M.; Takumi, T. The orphan nuclear receptor RORα regulates circadian transcription of the mammalian core-clock Bmal1. Nat. Struct. Mol. Biol. 2005, 12, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Mazzoccoli, G.; Pazienza, V.; Vinciguerra, M. Clock Genes and Clock-Controlled Genes in the Regulation of Metabolic Rhythms. Chronobiol. Int. 2012, 29, 227–251. [Google Scholar] [CrossRef] [PubMed]
- Gatten, R.E. Cardiovascular and Other Physiological Correlates of Hibernation in Aquatic and Terrestrial Turtles. Am. Zool. 1987, 27, 59–68. [Google Scholar] [CrossRef]
- Magnone, M.C.; Jacobmeier, B.; Bertolucci, C.; Foà, A.; Albrecht, U. Circadian expression of the clock gene Per2 is altered in the ruin lizard (Podarcis sicula) when temperature changes. Mol. Brain Res. 2005, 133, 281–285. [Google Scholar] [CrossRef]
- Vallone, D.; Frigato, E.; Vernesi, C.; Foà, A.; Foulkes, N.S.; Bertolucci, C. Hypothermia modulates circadian clock gene expression in lizard peripheral tissues. Am. J. Physiol. Integr. Comp. Physiol. 2007, 292, R160–R166. [Google Scholar] [CrossRef]
- Ikeno, T.; Williams, C.; Buck, C.L.; Barnes, B.M.; Yan, L. Clock Gene Expression in the Suprachiasmatic Nucleus of Hibernating Arctic Ground Squirrels. J. Biol. Rhythm. 2017, 32, 246–256. [Google Scholar] [CrossRef]
- Yearicks, E.F.; Wood, R.C.; Johnson, W.S. Hibernation of the Northern Diamondback Terrapin, Malaclemys terrapin terrapin. Estuaries 1981, 4, 78–80. [Google Scholar] [CrossRef]
- Ultsch, G.R. Ecology and physiology of hibernation and overwintering among freshwater fishes, turtles, and snakes. Biol. Rev. 1989, 64, 435–515. [Google Scholar] [CrossRef]
- Ikegami, K.; Refetoff, S.; Van Cauter, E.; Yoshimura, T. Interconnection between circadian clocks and thyroid function. Nat. Rev. Endocrinol. 2019, 15, 590–600. [Google Scholar] [CrossRef]
- Neumann, A.-M.; Schmidt, C.X.; Brockmann, R.M.; Oster, H. Circadian regulation of endocrine systems. Auton. Neurosci. 2019, 216, 1–8. [Google Scholar] [CrossRef]
- Oster, H. The interplay between stress, circadian clocks, and energy metabolism. J. Endocrinol. 2020, 247, R13–R25. [Google Scholar] [CrossRef] [PubMed]
- Gamble, K.L.; Berry, R.; Frank, S.J.; Young, M.E. Circadian clock control of endocrine factors. Nat. Rev. Endocrinol. 2014, 10, 466–475. [Google Scholar] [CrossRef]
- So, A.Y.-L.; Bernal, T.U.; Pillsbury, M.L.; Yamamoto, K.R.; Feldman, B.J. Glucocorticoid regulation of the circadian clock modulates glucose homeostasis. Proc. Natl. Acad. Sci. USA 2009, 106, 17582–17587. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.L. Melatonin, diel rhythms, and metamorphosis in anuran amphibians. Gen. Comp. Endocrinol. 2002, 126, 251–254. [Google Scholar] [CrossRef]
- Isorna, E.; Besseau, L.; Boeuf, G.; Desdevises, Y.; Vuilleumier, R.; Alonso-Gómez, A.L.; Delgado, M.J.; Falcón, J. Retinal, pineal and diencephalic expression of frog arylalkylamine N-acetyltransferase-1. Mol. Cell. Endocrinol. 2006, 252, 11–18. [Google Scholar] [CrossRef]
- Venegas, C.; García, J.A.; Doerrier, C.; Volt, H.; Escames, G.; López, L.C.; Reiter, R.J.; Acuña-Castroviejo, D. Analysis of the daily changes of melatonin receptors in the rat liver. J. Pineal Res. 2013, 54, 313–321. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin and retinoid orphan receptors: Demand for new interpretations after their exclusion as nuclear melatonin receptors. Melatonin Res. 2018, 1, 78–93. [Google Scholar] [CrossRef]
- Li, D.Y.; Smith, D.G.; Hardeland, R.; Yang, M.Y.; Xu, H.L.; Zhang, L.; Yin, H.D.; Zhu, Q. Melatonin Receptor Genes in Vertebrates. Int. J. Mol. Sci. 2013, 14, 11208–11223. [Google Scholar] [CrossRef]
- Wiechmann, A.F.; Wirsig-Wiechmann, C.R. Melatonin Receptor mRNA and Protein Expression in Xenopus laevis Nonpigmented Ciliary Epithelial Cells. Exp. Eye Res. 2001, 73, 617–623. [Google Scholar] [CrossRef]
- Wiechmann, A.F.; Smith, A.R. Melatonin receptor RNA is expressed in photoreceptors and displays a diurnal rhythm in Xenopus retina. Mol. Brain Res. 2001, 91, 104–111. [Google Scholar] [CrossRef]
- Owino, S.; Buonfiglio, D.D.C.; Tchio, C.; Tosini, G. Melatonin Signaling a Key Regulator of Glucose Homeostasis and Energy Metabolism. Front. Endocrinol. 2019, 10, 488. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Pérez, J.; Míguez, J.M.; Librán-Pérez, M.; Otero-Rodiño, C.; Naderi, F.; Soengas, J.L.; López-Patiño, M.A. Daily rhythms in activity and mRNA abundance of enzymes involved in glucose and lipid metabolism in liver of rainbow trout, Oncorhynchus mykiss. Influence of light and food availability. Chronobiol. Int. 2015, 32, 1391–1408. [Google Scholar] [CrossRef] [PubMed]
- Cahill, G.M. Circadian regulation of melatonin production in cultured zebrafish pineal and retina. Brain Res. 1996, 708, 177–181. [Google Scholar] [CrossRef]
- Falso, P.G.; Noble, C.A.; Diaz, J.M.; Hayes, T.B. The effect of long-term corticosterone treatment on blood cell differentials and function in laboratory and wild-caught amphibian models. Gen. Comp. Endocrinol. 2015, 212, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Cahill, G.M.; Besharse, J.C. Circadian clock functions localized in xenopus retinal photoreceptors. Neuron 1993, 10, 573–577. [Google Scholar] [CrossRef]
- Hill, S.M.; Frasch, T.; Xiang, S.; Yuan, L.; Duplessis, T.; Mao, L. Molecular Mechanisms of Melatonin Anticancer Effects. Integr. Cancer Ther. 2009, 8, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Zapico, C.; Coto-Montes, A. A proposed mechanism to explain the stimulatory effect of melatonin on antioxidative enzymes. J. Pineal Res. 2005, 39, 99–104. [Google Scholar] [CrossRef]
- Jang, Y.-S.; Lee, M.-H.; Lee, S.-H.; Bae, K. Cu/Zn superoxide dismutase is differentially regulated in period gene-mutant mice. Biochem. Biophys. Res. Commun. 2011, 409, 22–27. [Google Scholar] [CrossRef]
- Sani, M.; Sebaï, H.; Gadacha, W.; Boughattas, N.A.; Reinberg, A.; Mossadok, B.A. Catalase activity and rhythmic patterns in mouse brain, kidney and liver. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 145, 331–337. [Google Scholar] [CrossRef]
- Kim, E.-J.; Yoo, Y.-G.; Yang, W.-K.; Lim, Y.-S.; Na, T.-Y.; Lee, I.-K.; Lee, M.-O. Transcriptional Activation of HIF-1 by RORα and its Role in Hypoxia Signaling. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1796–1802. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, L.; Ding, S.; Lin, N.; Ji, Q.; Gao, L.; Su, Y.; He, B.; Pu, J. Novel protective role of the circadian nuclear receptor retinoic acid-related orphan receptor-α in diabetic cardiomyopathy. J. Pineal Res. 2017, 62, e12378. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Mayo, J.C.; Sainz, R.M.; Leon, J.; Czarnocki, Z. Melatonin as an antioxidant: Biochemical mechanisms and pathophysiological implications in humans. Acta Biochim. Pol. 2003, 50, 1129–1146. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Manchester, L.C.; Esteban-Zubero, E.; Zhou, Z.; Reiter, R.J. Melatonin as a Potent and Inducible Endogenous Antioxidant: Synthesis and Metabolism. Molecules 2015, 20, 18886–18906. [Google Scholar] [CrossRef] [PubMed]
- Ward, C.K.; Fontes, C.; Breuner, C.W.; Mendonça, M.T. Characterization and quantification of corticosteroid-binding globulin in a southern toad, Bufo terrestris, exposed to coal-combustion-waste. Gen. Comp. Endocrinol. 2007, 152, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Joanisse, D.R.; Storey, K. Oxidative damage and antioxidants in Rana sylvatica, the freeze-tolerant wood frog. Am. J. Physiol. Integr. Comp. Physiol. 1996, 271, R545–R553. [Google Scholar] [CrossRef]
- Ren, X.; Zhang, J.; Wang, L.; Wang, Z.; Wang, Y. Diel variation in cortisol, glucose, lactic acid and antioxidant system of black sea bass Centropristis striata under natural photoperiod. Chronobiol. Int. 2020, 37, 176–188. [Google Scholar] [CrossRef]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Herrera, F.; Antol, I. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef]
- Boutilier, R.G.; Donohoe, P.H.; Tattersall, G.; West, T.G. Hypometabolic homeostasis in overwintering aquatic amphibians. J. Exp. Biol. 1997, 200, 387–400. [Google Scholar] [CrossRef]
- Tattersall, G.J.; Ultsch, G.R. Physiological Ecology of Aquatic Overwintering in Ranid Frogs. Biol. Rev. 2008, 83, 119–140. [Google Scholar] [CrossRef]
- Holenweg, A.K.; Reyer, H.U. Hibernation behavior of Rana lessonae and R. esculenta in their natural habitat. Oecologia 2000, 123, 41–47. [Google Scholar] [CrossRef]
- Milsom, W.K.; Jackson, D.C. Hibernation and Gas Exchange. Compr. Physiol. 2011, 1, 397–420. [Google Scholar] [CrossRef]
- Stinner, J.; Zarlinga, N.; Orcutt, S. Overwintering behavior of adult bullfrogs, Rana catesbeiana, in northeastern Ohio. Ohio J. Sci. 1994, 94, 8–13. [Google Scholar]
- Costanzo, J.P.; Lee, R.E. Avoidance and tolerance of freezing in ectothermic vertebrates. J. Exp. Biol. 2013, 216, 1961–1967. [Google Scholar] [CrossRef] [PubMed]
- Kiss, A.J.; Muir, T.J.; Lee, J.R.E.; Costanzo, J.P. Seasonal Variation in the Hepatoproteome of the Dehydration- and Freeze-Tolerant Wood Frog, Rana sylvatica. Int. J. Mol. Sci. 2011, 12, 8406–8414. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.-K.; Shoemaker, K.T.; Karraker, N.E. Demography of Quasipaa frogs in China reveals high vulnerability to widespread harvest pressure. Biol. Conserv. 2014, 170, 3–9. [Google Scholar] [CrossRef]
- Yu, Y.; Hu, Y.; Zhang, Q.; Zheng, R.; Shen, B.; Kong, S.; Li, K. Female Preferences for Call Properties of Giant Spiny Frog (Quasipaa spinosa). Pak. J. Zool. 2020, 52, 825–1224. [Google Scholar] [CrossRef]
- Zheng, R.-Q.; Liu, C.-T. Giant spiny-frog (Paa spinosa) from different populations differ in thermal preference but not in thermal tolerance. Aquat. Ecol. 2010, 44, 723–729. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Hughes, M.E.; HogenEsch, J.B.; Kornacker, K. JTK_CYCLE: An Efficient Nonparametric Algorithm for Detecting Rhythmic Components in Genome-Scale Data Sets. J. Biol. Rhythm. 2010, 25, 372–380. [Google Scholar] [CrossRef]
- Lamia, K.A.; Papp, S.J.; Yu, R.T.; Barish, G.D.; Uhlenhaut, H.; Jonker, J.; Downes, M.; Evans, R.M. Cryptochromes mediate rhythmic repression of the glucocorticoid receptor. Nature 2011, 480, 552–556. [Google Scholar] [CrossRef]
- Serino, I.; D’Istria, M.; Monteleone, P. A comparative study of melatonin production in the retina, pineal gland and Harderian gland of Bufo viridis and Rana esculenta. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1993, 106, 189–193. [Google Scholar] [CrossRef]
- Neufeld-Cohen, A.; Robles, M.S.; Aviram, R.; Manella, G.; Adamovich, Y.; Ladeuix, B.; Nir, D.; Rousso-Noori, L.; Kuperman, Y.; Golik, M.; et al. Circadian control of oscillations in mitochondrial rate-limiting enzymes and nutrient utilization by PERIOD proteins. Proc. Natl. Acad. Sci. USA 2016, 113, E1673–E1682. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.G.; Zhu, J.-H.; Cheng, W.-H.; Bao, Y.; Ho, Y.-S.; Reddi, A.R.; Holmgren, A.; Arnér, E. Paradoxical Roles of Antioxidant Enzymes: Basic Mechanisms and Health Implications. Physiol. Rev. 2016, 96, 307–364. [Google Scholar] [CrossRef] [PubMed]
- Reinke, H.; Asher, G. Crosstalk between metabolism and circadian clocks. Nat. Rev. Mol. Cell Biol. 2019, 20, 227–241. [Google Scholar] [CrossRef]
- Hardeland, R.; Coto-Montes, A.; Poeggeler, B. Circadian rhythms, oxidative stress, and antioxidative defense mechanisms. Chronobiol. Int. 2003, 20, 921–962. [Google Scholar] [CrossRef]
- Ruby, N.F.; Dark, J.; Burns, D.E.; Heller, H.C.; Zucker, I. The Suprachiasmatic Nucleus Is Essential for Circadian Body Temperature Rhythms in Hibernating Ground Squirrels. J. Neurosci. 2002, 22, 357–364. [Google Scholar] [CrossRef]
- Williams, C.T.; Barnes, B.M.; Richter, M.; Buck, C.L. Hibernation and Circadian Rhythms of Body Temperature in Free-Living Arctic Ground Squirrels. Physiol. Biochem. Zool. 2012, 85, 397–404. [Google Scholar] [CrossRef]
- Williams, C.T.; Radonich, M.; Barnes, B.M.; Buck, C.L. Seasonal loss and resumption of circadian rhythms in hibernating arctic ground squirrels. J. Comp. Physiol. B 2017, 187, 693–703. [Google Scholar] [CrossRef]
- Florant, G.L.; Rivera, M.L.; Lawrence, A.K.; Tamarkin, L. Plasma melatonin concentrations in hibernating marmots: Absence of a plasma melatonin rhythm. Am. J. Physiol. Integr. Comp. Physiol. 1984, 247, R1062–R1066. [Google Scholar] [CrossRef]
- Delgado, M.; Vivien-Roels, B. Effect of environmental temperature and photoperiod on the melatonin levels in the pineal, lateral eye, and plasma of the frog, Rana perezi: Importance of ocular melatonin. Gen. Comp. Endocrinol. 1989, 75, 46–53. [Google Scholar] [CrossRef]
- Chong, N.W.; Bernard, M.; Klein, D.C. Characterization of the Chicken Serotonin N-Acetyltransferase Gene: Activation via Clock gene heterodimer/E box interaction. J. Biol. Chem. 2000, 275, 32991–32998. [Google Scholar] [CrossRef] [PubMed]
- Son, G.H.; Chung, S.; Choe, H.K.; Kim, H.-D.; Baik, S.-M.; Lee, H.; Lee, H.-W.; Choi, S.; Sun, W.; Kim, H.; et al. Adrenal peripheral clock controls the autonomous circadian rhythm of glucocorticoid by causing rhythmic steroid production. Proc. Natl. Acad. Sci. USA 2008, 105, 20970–20975. [Google Scholar] [CrossRef] [PubMed]
- Silveira, E.J.D.; Filho, C.H.V.N.; Yujra, V.Q.; Webber, L.P.; Castilho, R.M.; Squarize, C.H. BMAL1 Modulates Epidermal Healing in a Process Involving the Antioxidative Defense Mechanism. Int. J. Mol. Sci. 2020, 21, 901. [Google Scholar] [CrossRef] [PubMed]
- Chhunchha, B.; Kubo, E.; Singh, D.P. Clock Protein Bmal1 and Nrf2 Cooperatively Control Aging or Oxidative Response and Redox Homeostasis by Regulating Rhythmic Expression of Prdx6. Cells 2020, 9, 1861. [Google Scholar] [CrossRef]
- Niu, Y.; Zhang, X.; Zhang, H.; Xu, T.; Zhu, L.; Storey, K.B.; Chen, Q. Metabolic responses of plasma to extreme environments in overwintering Tibetan frogs Nanorana parkeri: A metabolome integrated analysis. Front. Zool. 2021, 18, 41. [Google Scholar] [CrossRef]
- Niu, Y.; Cao, W.; Zhao, Y.; Zhai, H.; Zhao, Y.; Tang, X.; Chen, Q. The levels of oxidative stress and antioxidant capacity in hibernating Nanorana parkeri. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 219–220, 19–27. [Google Scholar] [CrossRef]
- Niu, Y.; Cao, W.; Storey, K.B.; He, J.; Wang, J.; Zhang, T.; Tang, X.; Chen, Q. Metabolic characteristics of overwintering by the high-altitude dwelling Xizang plateau frog, Nanorana parkeri. J. Comp. Physiol. B 2020, 190, 433–444. [Google Scholar] [CrossRef]
- Niu, Y.; Chen, Q.; Storey, K.B.; Teng, L.; Li, X.; Xu, T.; Zhang, H. Physiological Ecology of Winter Hibernation by the High-Altitude Frog Nanorana parkeri. Physiol. Biochem. Zool. 2022, 95, 201–211. [Google Scholar] [CrossRef]
- Petrović, V.M.; Miliič, B.; Spasic, M.; Saičić, Z. Copper-Zinc Containing and Manganese Containing Superoxide Dismutase in the Ground Squirrel/Citellus Citellus/—THE Effect of Hibernation. Free Radic. Res. Commun. 1986, 1, 339–346. [Google Scholar] [CrossRef]
- Gattoni, G.; Bernocchi, G. Calcium-Binding Proteins in the Nervous System during Hibernation: Neuroprotective Strategies in Hypometabolic Conditions? Int. J. Mol. Sci. 2019, 20, 2364. [Google Scholar] [CrossRef]
- Bagnyukova, T.; Storey, K.; Lushchak, V. Induction of oxidative stress in Rana ridibunda during recovery from winter hibernation. J. Therm. Biol. 2003, 28, 21–28. [Google Scholar] [CrossRef]
- Buzadžić, B.; Blagojević, D.; Korać, B.; Saicic, Z.; Spasić, M.; Petrović, V. Seasonal Variation in the Antioxidant Defense System of the Brain of the Ground Squirrel (Citellus citellus) and Response to Low Temperature Compared with Rat. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1997, 117, 141–149. [Google Scholar] [CrossRef]
- Mattice, J.J.L. Regulation of Glutathione-Based Antioxidant Defenses in Response to Dehydration Stress in the African Clawed Frog, Xenopus laevis. Ph.D. Thesis, Carleton University, Ottawa, ONT, Canada, 2018. [Google Scholar] [CrossRef]
- Mauvoisin, D. Circadian rhythms and proteomics: It’s all about posttranslational modifications! WIREs Syst. Biol. Med. 2019, 11, e1450. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Pre-Hibernation | Hibernation | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Tissue | Variable | Mesor | Amplitude | Acrophase | ADJ.P | Mesor | Amplitude | Acrophase | ADJ.P |
| Retina | Bmal1 | 0.97 | 0.03 | 0 | 0.659 | 0.84 | 0.13 | 8 | 0.004 |
| Clock | 0.94 | 0.11 | 20 | 0.144 | 1.59 | 0.56 | 14 | 0.009 | |
| Cry1 | 1.07 | 0.11 | 18 | 0.194 | 1.05 | 0.09 | 12 | 1.000 | |
| Per2 | 1.10 | 0.21 | 20 | 0.011 | 1.65 | 0.36 | 14 | 0.135 | |
| Brain | Bmal1 | 0.67 | 0.13 | 8 | <0.001 | 1.12 | 0.04 | 12 | 1.000 |
| Clock | 1.03 | 0.04 | 2 | 0.360 | 1.59 | 0.13 | 0 | 0.027 | |
| Cry1 | 0.66 | 0.04 | 6 | 0.031 | 0.92 | 0.06 | 2 | 0.102 | |
| Per2 | 1.05 | 0.13 | 8 | 0.002 | 1.65 | 0.18 | 0 | 0.057 | |
| Liver | Bmal1 | 1.02 | 0.07 | 22 | 1.000 | 1.59 | 0.13 | 12 | 0.816 |
| Clock | 1.30 | 0.09 | 22 | 0.384 | 1.15 | 0.09 | 10 | 1.000 | |
| Cry1 | 0.68 | 0.72 | 10 | <0.001 | 1.29 | 0.10 | 0 | 0.851 | |
| Per2 | 2.63 | 0.32 | 18 | 1.000 | 1.48 | 0.09 | 10 | 1.000 | |
| Brain | SOD1 | 0.54 | 0.06 | 6 | 0.033 | 1.33 | 0.21 | 0 | 0.022 |
| SOD2 | 0.48 | 0.07 | 8 | 0.035 | 0.87 | 0.09 | 4 | 0.570 | |
| CAT | 0.75 | 0.04 | 6 | 0.022 | 1.43 | 0.03 | 0 | 0.881 | |
| GPx | 0.70 | 0.13 | 8 | 0.014 | 1.00 | 0.04 | 6 | 1.000 | |
| Liver | SOD1 | 2.03 | 0.25 | 10 | 0.748 | 2.82 | 0.64 | 22 | 0.450 |
| SOD2 | 1.17 | 0.10 | 10 | 1.000 | 1.72 | 0.33 | 20 | 0.558 | |
| CAT | 1.59 | 0.17 | 0 | 1.000 | 0.92 | 0.10 | 18 | 0.160 | |
| GPx | 1.60 | 0.10 | 16 | 1.000 | 0.94 | 0.13 | 22 | 1.000 | |
| Brain | Ror-α | 0.93 | 0.08 | 4 | 0.270 | 1.50 | 0.16 | 0 | 0.007 |
| Liver | Ror-α | 1.54 | 0.22 | 18 | 0.194 | 1.04 | 0.13 | 20 | 0.310 |
| Retina | Mel-1c | 2.38 | 0.12 | 18 | 1.000 | 0.64 | 0.36 | 8 | <0.001 |
| Brain | Mel-1c | 0.41 | 0.10 | 6 | 0.007 | 0.79 | 0.07 | 8 | 0.191 |
| Retina | AANAT | 1.39 | 0.19 | 18 | 0.816 | 1.05 | 0.23 | 10 | 0.659 |
| Plasma | MT | 170.93 | 4.76 | 2 | 0.305 | 190.41 | 14.22 | 8 | 0.006 |
| CORT | 139.54 | 4.55 | 6 | 1.000 | 151.95 | 2.08 | 16 | 0.620 | |
| Parameters | Variables | Retina | Brain | Liver | |||
|---|---|---|---|---|---|---|---|
| T Value | p Value | t Value | p Value | t Value | p Value | ||
| Clock genes | Bmal1 | 1.947 | 0.055 | −11.963 | <0.001 | −5.761 | <0.001 |
| Clock | −5.672 | <0.001 | −8.598 | <0.001 | 2.139 | 0.035 | |
| Cry1 | 0.118 | 0.906 | −7.739 | <0.001 | −5.328 | <0.001 | |
| Per2 | −5.423 | <0.001 | −7.741 | <0.001 | 4.896 | <0.001 | |
| Clock controlled genes | Ror-α | −8.987 | <0.001 | 4.325 | <0.001 | ||
| Mel-1c | 13.294 | <0.001 | −5.63 | <0.001 | |||
| AANAT | 1.832 | 0.07 | |||||
| Antioxidants | SOD1 | −7.994 | <0.001 | −2.784 | 0.007 | ||
| SOD2 | −7.779 | <0.001 | −3.805 | <0.001 | |||
| CAT | −13.957 | <0.001 | 4.573 | <0.001 | |||
| GPx | −7.99 | <0.001 | 3.657 | 0.001 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Z.; Ahmad, I.M.; Zuo, L.; Wang, H.; Li, D. Hibernation with Rhythmicity in the Retina, Brain, and Plasma but Not in the Liver of Hibernating Giant Spiny Frogs (Quasipaa spinosa). Biology 2022, 11, 722. https://doi.org/10.3390/biology11050722
Xie Z, Ahmad IM, Zuo L, Wang H, Li D. Hibernation with Rhythmicity in the Retina, Brain, and Plasma but Not in the Liver of Hibernating Giant Spiny Frogs (Quasipaa spinosa). Biology. 2022; 11(5):722. https://doi.org/10.3390/biology11050722
Chicago/Turabian StyleXie, Zhigang, Ibrahim M. Ahmad, Lirong Zuo, Hui Wang, and Dongming Li. 2022. "Hibernation with Rhythmicity in the Retina, Brain, and Plasma but Not in the Liver of Hibernating Giant Spiny Frogs (Quasipaa spinosa)" Biology 11, no. 5: 722. https://doi.org/10.3390/biology11050722
APA StyleXie, Z., Ahmad, I. M., Zuo, L., Wang, H., & Li, D. (2022). Hibernation with Rhythmicity in the Retina, Brain, and Plasma but Not in the Liver of Hibernating Giant Spiny Frogs (Quasipaa spinosa). Biology, 11(5), 722. https://doi.org/10.3390/biology11050722