
Phosphate Solubilizing Microorganism Bacillus sp. MVY-004 and Its Significance for Biomineral Fertilizers’ Development in Agrobiotechnology

 , ,
, ,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Samples Collection
2.2. Phosphate Solubilizing Microorganism Isolation and Phosphate Solubilizing Activity
2.3. Molecular Identification of the Microorganism Strain
2.4. Microorganism and Inoculant Preparation
2.5. The Nutrient Medium Composition and Fermentation Process Parameters’ Optimization
2.6. Fed-Batch Fermentation Process Optimization and Bacterial Broth Samples Collection
2.7. Organic Acids Analysis by Liquid Chromatography Time-of-Flight Mass Spectrometry (LC-TOF/MS)
2.8. Phytohormones Analysis by Liquid Chromatography Time-of-Flight Mass Spectrometry (LC-TOF/MS)
2.9. Mobile Phosphorus (P2O5) Concentration Changes in Loamy Soil Using Mineral Fertilizers (NPS-12:40:10) and Biomineral Fertilizers (NPS-12:40:10 + Bacillus sp. MVY-004)
- Normal (T = 20 °C; relative humidity 60%),
- Hot and dry (T = 30 °C; relative humidity 30%),
- Hot and humid (T = 30 °C; relative humidity 80%).
2.10. Bacillus sp. MVY-004 Cells Viability and Activity in Biomineral Fertilizers
2.11. Statistical Analysis
3. Results and Discussion
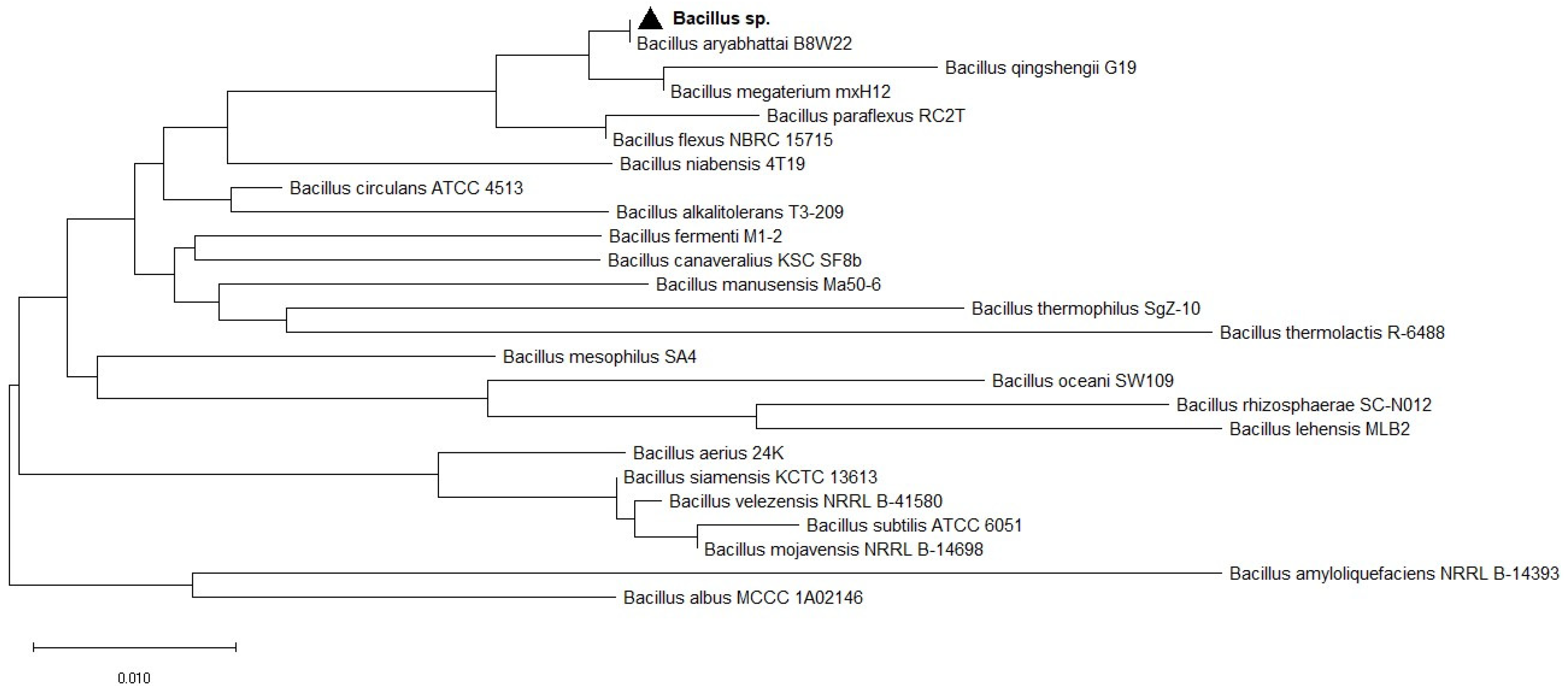
3.1. Isolation, Screening, and Identification of Phosphate Solubilizing Bacteria
3.2. Optimization of the Culture Medium and Growth Conditions of the Bacillus sp. MVY-004
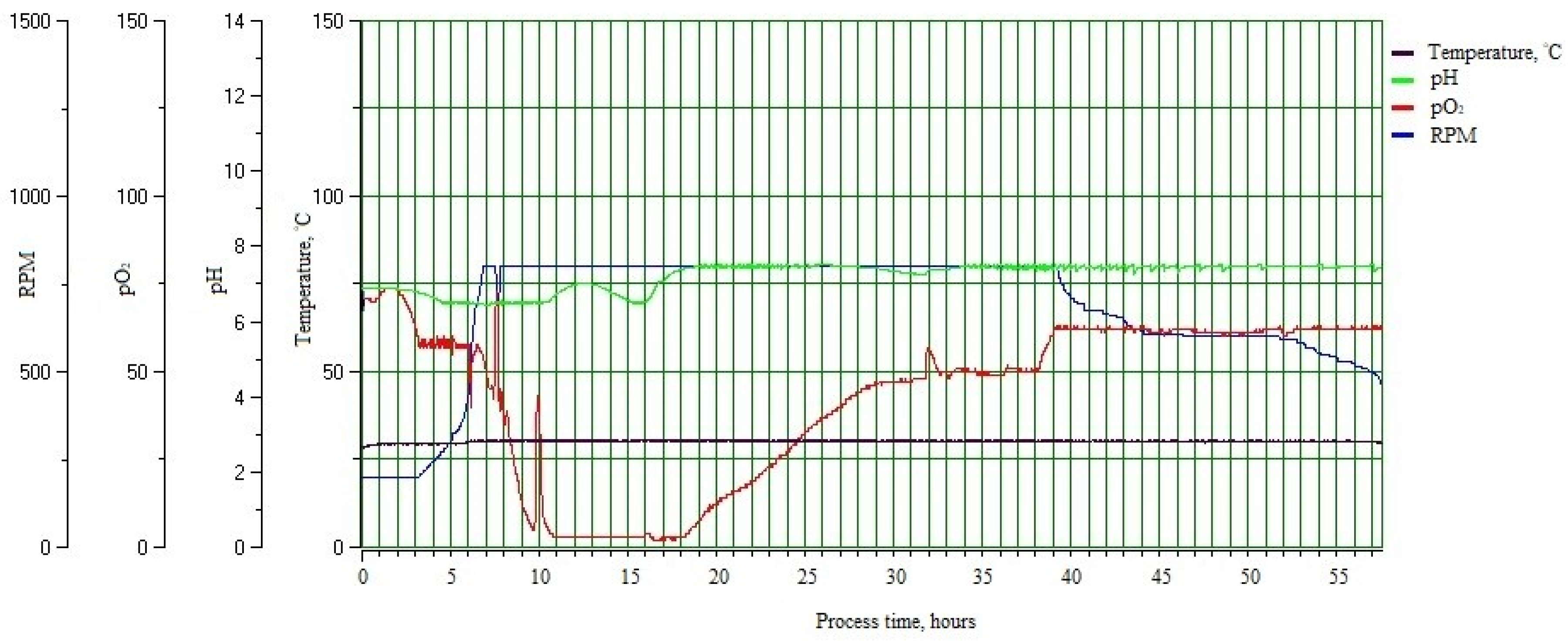
3.3. Optimization of the Industrial Fermentation Process of the Bacillus sp. MVY-004
3.4. Organic Acids and Plant Hormone Detection in Bacterial Supernatant Using Liquid Chromatography Time-of-Flight Mass Spectrometry (LC-TOF/MS) with External Calibration
3.5. Variation of Mobile Phosphorus (P2O5) Concentration in Loamy Soil Using Fertilizers NPS-12:40:10 and NPS-12:40:10 + Bacillus sp. MVY-004
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kalayu, G. Phosphate solubilizing microorganisms: Promising approach as biofertilizers. Int. J. Agron. 2019, 2019, 4917256. [Google Scholar] [CrossRef]
- Malhotra, H.; Vandana Sharma, S.; Pandey, R. Phosphorus nutrition: Plant growth in response to deficiency and excess. Plant Nutr. Abiotic Stress Toler. 2018, 7, 171–190. [Google Scholar]
- Bindraban, P.S.; Dimkpa, C.O.; Pandey, R. Exploring phosphorus fertilizers and fertilization strategies for improved human and environmental health. Biol. Fertil. Soils 2020, 56, 299–317. [Google Scholar] [CrossRef]
- Weeks, J.J.; Hettiarachchi, G.M. A review of the latest in phosphorus fertilizer technology: Possibilities and pragmatism. J. Environ. Qual. 2019, 48, 1300–1313. [Google Scholar] [CrossRef]
- Bargaz, A.; Lyamlouli, K.; Chtouki, M.; Zeroual, Y.; Dhiba, D. Soil microbial resources for improving fertilizers efficiency in an integrated plant nutrient management system. Front. Microbiol. 2018, 9, 1606. [Google Scholar] [CrossRef]
- Qurban, A.P. Isolation and characterization of phosphate-solubilizing bacteria from aerobic rice. Afr. J. Biotechnol. 2012, 11, 2711–2719. [Google Scholar]
- Rouphael, Y.; Colla, G. Toward a sustainable agriculture through plant biostimulants: From experimental data to practical applications. Agronomy 2020, 10, 1461. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- Wei, Y.; Zhao, Y.; Shi, M.; Cao, Z.; Lu, Q.; Yang, T.; Fan, Y.; Wei, Z. Effect of organic acids production and bacterial community on the possible mechanism of phosphorus solubilization during composting with enriched phosphate-solubilizing bacteria inoculation. Bioresour. Technol. 2018, 247, 190–199. [Google Scholar] [CrossRef]
- Pande, A.; Pandey, P.; Mehra, S.; Singh, M.; Kaushik, S. Phenotypic and genotypic characterization of phosphate solubilizing bacteria and their efficiency on the growth of maize. J. Genet. Eng. Biotechnol. 2017, 15, 379–391. [Google Scholar] [CrossRef]
- Migliorini, P.; Wezel, A. Converging and diverging principles and practices of organic agriculture regulations and agroecology. A review. Agron. Sustain. Dev. 2017, 37, 63. [Google Scholar] [CrossRef]
- Antoun, H. Beneficial microorganisms for the sustainable use of phosphates in agriculture. Proc. Procedia Eng. 2012, 46, 62–67. [Google Scholar] [CrossRef]
- Nev, O.A.; Lindsay, R.J.; Jepson, A.; Butt, L.; Beardmore, R.E.; Gudelj, I. Predicting microbial growth dynamics in response to nutrient availability. PLoS Comput. Biol. 2021, 17, e1008817. [Google Scholar] [CrossRef] [PubMed]
- Franco-Duarte, R.; Černáková, L.; Kadam, S.; Kaushik, K.S.; Salehi, B.; Bevilacqua, A.; Corbo, M.R.; Antolak, H.; Dybka-Stępień, K.; Leszczewicz, M.; et al. Advances in chemical and biological methods to identify microorganisms—From past to present. Microorganisms 2019, 7, 130. [Google Scholar] [CrossRef] [PubMed]
- Jornitz, M.W.; Cappia, J.-M.; Rao, G. Industrial biotechnology and commodity products. Compr. Biotechnol. 2011, 3, 641–652. [Google Scholar]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef]
- Formenti, L.R.; Nørregaard, A.; Bolic, A.; Hernandez, D.Q.; Hagemann, T.; Heins, A.L.; Larsson, H.; Mears, L.; Mauricio-Iglesias, M.; Krühne, U.; et al. Challenges in industrial fermentation technology research. Biotechnol. J. 2014, 9, 727–738. [Google Scholar] [CrossRef]
- Pham, J.V.; Yilma, M.A.; Feliz, A.; Majid, M.T.; Maffetone, N.; Walker, J.R.; Kim, E.; Cho, H.J.; Reynolds, J.M.; Song, M.C.; et al. A review of the microbial production of bioactive natural products and biologics. Front. Microbiol. 2019, 10, 1404. [Google Scholar] [CrossRef]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The role of soil microorganisms in plant mineral nutrition—current knowledge and future directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef]
- Ye, L.; Zhao, X.; Bao, E.; Li, J.; Zou, Z.; Cao, K. Bio-organic fertilizer with reduced rates of chemical fertilization improves soil fertility and enhances tomato yield and quality. Sci. Rep. 2020, 10, 177. [Google Scholar] [CrossRef]
- Soumare, A.; Boubekri, K.; Lyamlouli, K.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L. From isolation of phosphate solubilizing microbes to their formulation and use as biofertilizers: Status and needs. Front. Bioeng. Biotechnol. 2020, 7, 425. [Google Scholar] [CrossRef] [PubMed]
- Nosheen, S.; Ajmal, I.; Song, Y. Microbes as biofertilizers, a potential approach for sustainable crop production. Sustainability 2021, 13, 1868. [Google Scholar] [CrossRef]
- Efremova, S.Y.; Kulikova, I.V.; Sukhova, N.V. Agroecological efficiency of biomodified mineral fertilizers. In Proceedings of the International Conference on Efficient Production and Processing (ICEPP-2020), Prague, Czech Republic, 27–28 February 2020; Smyatskaya, J., Ed.; E3S Web of Conferences: Les Ulis, France, 2020; Volume 161, p. 01115. [Google Scholar]
- Onyia, C.E.; Anyanwu, C.U. Comparative study on solubilization of tri-calcium phosphate (TCP) by phosphate solubilizing fungi (PSF) isolated from Nsukka pepper plant rhizosphere and root free soil. J. Yeast Fungal Res. 2013, 4, 52–57. [Google Scholar]
- Boczek, L.A.; Rice, E.W.; Johnson, C.H. Total viable counts: Spread plate technique. In Encyclopedia of Food Microbiology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 3, pp. 636–637. [Google Scholar]
- Paul, D.; Narayan Sinha, S. Phosphate solubilizing activity of some bacterial strains isolated from jute mill effluent exposed water of river Ganga. Indian J. Fundam. Appl. Life Sci. 2013, 3, 39–45. [Google Scholar]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 46, D41–D47. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Macwilliams, M.P.; Liao, M.-K. Luria broth (LB) and Luria agar (LA) media and their uses protocol. Am. Soc. Microbiol. 2006, 3031, 1–4. Available online: https://asm.org/Protocols/Luria-Broth-LB-and-Luria-Agar-LA-Media-and-Their-U (accessed on 12 August 2019).
- Zhang, X.; Wang, Y.; Guo, J.; Yu, Y.; Li, J.; Guo, Y.; Liu, C. Comparing two functions for optical density and cell numbers in bacterial exponential growth phase. J. Pure Appl. Microbiol. 2015, 9, 299–305. [Google Scholar]
- Boukouvalas, D.T.; Prates, R.A.; Lima Leal, C.R.; de Araújo, S.A. Automatic segmentation method for CFU counting in single plate-serial dilution. Chemom. Intell. Lab. Syst. 2019, 195, 103889. [Google Scholar] [CrossRef]
- Kittle, A.J.; Sisler, S.E.; Nguyen, N.; Gill, J.C.; Reed, V. Method-comparison analysis between a Contour Next glucometer and YSI 2900 biochemistry analyzer in equine. J. Equine Vet. Sci. 2019, 76, 127. [Google Scholar] [CrossRef]
- Sargautiene, V.; Nakurte, I.; Nikolajeva, V. Broad prebiotic potential of non-starch polysaccharides from oats (Avena Sativa L.): An in vitro study. Pol. J. Microbiol. 2018, 67, 307–313. [Google Scholar] [CrossRef]
- Castillo, G.; Torrecillas, A.; Nogueiras, C.; Michelena, G.; Sánchez-Bravo, J.; Acosta, M. Simultaneous quantification of phytohormones in fermentation extracts of Botryodiplodia theobromae by liquid chromatography-electrospray tandem mass spectrometry. World J. Microbiol. Biotechnol. 2014, 30, 1937–1946. [Google Scholar] [CrossRef] [PubMed]
- Farahani, S.S.; Rajabipour, A.; Keyhani, A.; Sharifi, M. Energy use and economic analysis of NPK-15:8:15 fertilizer granulation process in Iran. J. Saudi Soc. Agric. Sci. 2017, 16, 265–269. [Google Scholar] [CrossRef]
- Assanga, I. Cell growth curves for different cell lines and their relationship with biological activities. Int. J. Biotechnol. Mol. Biol. Res. 2013, 4, 60–70. [Google Scholar] [CrossRef]
- Lu, C.; Yin, J.; Zhang, C.; Lu, W. Fed-batch fermentation spinosad production in an improved reactor. Trans. Tianjin Univ. 2017, 23, 1–8. [Google Scholar] [CrossRef]
- Watanabe, T.; Irokawa, H.; Ogasawara, A.; Iwai, K.; Kuge, S. Requirement of peroxiredoxin on the stationary phase of yeast cell growth. J. Toxicol. Sci. 2014, 39, 51–58. [Google Scholar] [CrossRef]
- Agnihotri, V.; Adhikari, P.; Pandey, N.; Sati, P.; Pandey, A. Thin layer drying behavior of Ginkgo Biloba L. leaves with respect to ginkgolide A and bilobalide content and microbial load. Heliyon 2020, 4, e03220. [Google Scholar] [CrossRef]
- Stella, M.; Theeba, M.; Illani, Z.I. Organic fertilizer amended with immobilized bacterial cells for extended shelf-life. Biocatal. Agric. Biotechnol. 2019, 20, 101248. [Google Scholar] [CrossRef]
- Eurostat. Statistic Explained Agri-Environmental Indicator-Mineral Fertiliser Consumption-Statistics Explained. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Main_Page (accessed on 10 September 2021).
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; Enshasy, H. El Plant growth promoting rhizobacteria (Pgpr) as green bioinoculants: Recent developments, constraints, and prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Tian, J.; Ge, F.; Zhang, D.; Deng, S.; Liu, X. Roles of phosphate solubilizing microorganisms from managing soil phosphorus deficiency to mediating biogeochemical p cycle. Biology 2021, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2013, 2, 587. [Google Scholar] [CrossRef] [PubMed]
- Kämpfer, P. Taxonomy of phosphate solublizing bacteria. In First International Meeting on Microbial Phosphate Solubilization; Springer: Dordrecht, The Netherlands, 2007; Volume 102, pp. 101–106. [Google Scholar]
- Chen, Q.; Liu, S. Identification and characterization of the phosphate-solubilizing bacterium Pantoea sp. S32 in reclamation soil in Shanxi, China. Front. Microbiol. 2019, 10, 2171. [Google Scholar] [CrossRef]
- Santoyo, G.; Hernández-Pacheco, C.; Hernández-Salmerón, J.; Hernández-León, R. The role of abiotic factors modulating the plant-microbe-soil interactions: Toward sustainable agriculture. A review. Span. J. Agric. Res. 2017, 15, e03R01. [Google Scholar] [CrossRef]
- Liu, C.; Gong, Z.; Shen, B.; Feng, E. Modelling and optimal control for a fed-batch fermentation process. Appl. Math. Model. 2013, 37, 695–706. [Google Scholar] [CrossRef]
- Yang, Z.; Xiong, H.-R. Culture conditions and types of growth media for mammalian cells. Biomed. Tissue Cult. 2012, 1, 1–18. [Google Scholar]
- Han, X.; Zeng, H.; Bartocci, P.; Fantozzi, F.; Yan, Y. Phytohormones and effects on growth and metabolites of microalgae: A review. Fermentation 2018, 4, 25. [Google Scholar] [CrossRef]
- Li, M.; Li, Q.; Yun, J.; Yang, X.; Wang, X.; Lian, B.; Lu, C. Bio-organic-mineral fertilizer can improve soil quality and promote the growth and quality of water spinach. Can. J. Soil Sci. 2017, 97, 552–560. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Hashem, A.; Abd Allah, E.F. Bacillus: A biological tool for crop improvement through bio-molecular changes in adverse environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef]
- Kashyap, B.K.; Solanki, M.K.; Pandey, A.K.; Prabha, S.; Kumar, P.; Kumari, B. Bacillus as plant growth promoting rhizobacteria (PGPR): A promising green agriculture technology. Plant Health Biot. Stress 2019, 2, 219–236. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days of Incubation | Colony Diameter (cm) | Solubilization Zone Diameter (cm) | Phosphate Solubilization Index (PSI) |
|---|---|---|---|
| 2 | 0.4 ± 0.08 | 0.6 ± 0.05 | 2.50 |
| 5 | 0.8 ± 0.08 | 1.2 ± 0.04 | 2.50 |
| 7 | 0.9 ± 0.12 | 1.5 ± 0.05 | 2.67 |
| 10 | 1.0 ± 0.12 | 1.7 ± 0.08 | 2.70 |
| 14 | 1.2 ± 0.16 | 2.2 ± 0.08 | 2.83 |
| Concentration, µg mL−1 (Average from Triplicates ± SD) | ||||||
|---|---|---|---|---|---|---|
| Citric Acid * | Succinic Acid ** | 2-ketogluconic Acid * | Gluconic Acid* | Malic Acid ** | Lactic Acid *** | Oxalic Acid *** |
| 13.5 ± 0.6 | 257.0 ± 3.0 | 68.0 ± 2.0 | 287.0 ± 13.0 | 232.0 ± 7.0 | 419.0 ± 3.0 | 34.0 ± 2.0 |
| Concentration, µg mL−1 (Average from Triplicates ± SD) | ||
|---|---|---|
| Indole-3-acetic Acid * | Jasmonic Acid ** | Gibberellic Acid *** |
| 1.370 ± 0.020 | 0.173 ± 0.009 | 0.800 ± 0.060 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mažylytė, R.; Kaziūnienė, J.; Orola, L.; Valkovska, V.; Lastauskienė, E.; Gegeckas, A. Phosphate Solubilizing Microorganism Bacillus sp. MVY-004 and Its Significance for Biomineral Fertilizers’ Development in Agrobiotechnology. Biology 2022, 11, 254. https://doi.org/10.3390/biology11020254
Mažylytė R, Kaziūnienė J, Orola L, Valkovska V, Lastauskienė E, Gegeckas A. Phosphate Solubilizing Microorganism Bacillus sp. MVY-004 and Its Significance for Biomineral Fertilizers’ Development in Agrobiotechnology. Biology. 2022; 11(2):254. https://doi.org/10.3390/biology11020254
Chicago/Turabian StyleMažylytė, Raimonda, Justina Kaziūnienė, Liana Orola, Valda Valkovska, Eglė Lastauskienė, and Audrius Gegeckas. 2022. "Phosphate Solubilizing Microorganism Bacillus sp. MVY-004 and Its Significance for Biomineral Fertilizers’ Development in Agrobiotechnology" Biology 11, no. 2: 254. https://doi.org/10.3390/biology11020254
APA StyleMažylytė, R., Kaziūnienė, J., Orola, L., Valkovska, V., Lastauskienė, E., & Gegeckas, A. (2022). Phosphate Solubilizing Microorganism Bacillus sp. MVY-004 and Its Significance for Biomineral Fertilizers’ Development in Agrobiotechnology. Biology, 11(2), 254. https://doi.org/10.3390/biology11020254