
Nutrient Utilization during Male Maturation and Protein Digestion in the Oriental Hornet

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Hornet Colonies
2.2. Preparation of Labelled Diets
2.3. Experimental Procedure for Measuring Nutrient Allocation and Incorporation in Males
2.4. Experimental Procedure for Measuring Protein Digestion in Adult Hornets
2.5. δ13C Analysis in Tissues
2.6. Statistical Analysis
3. Results
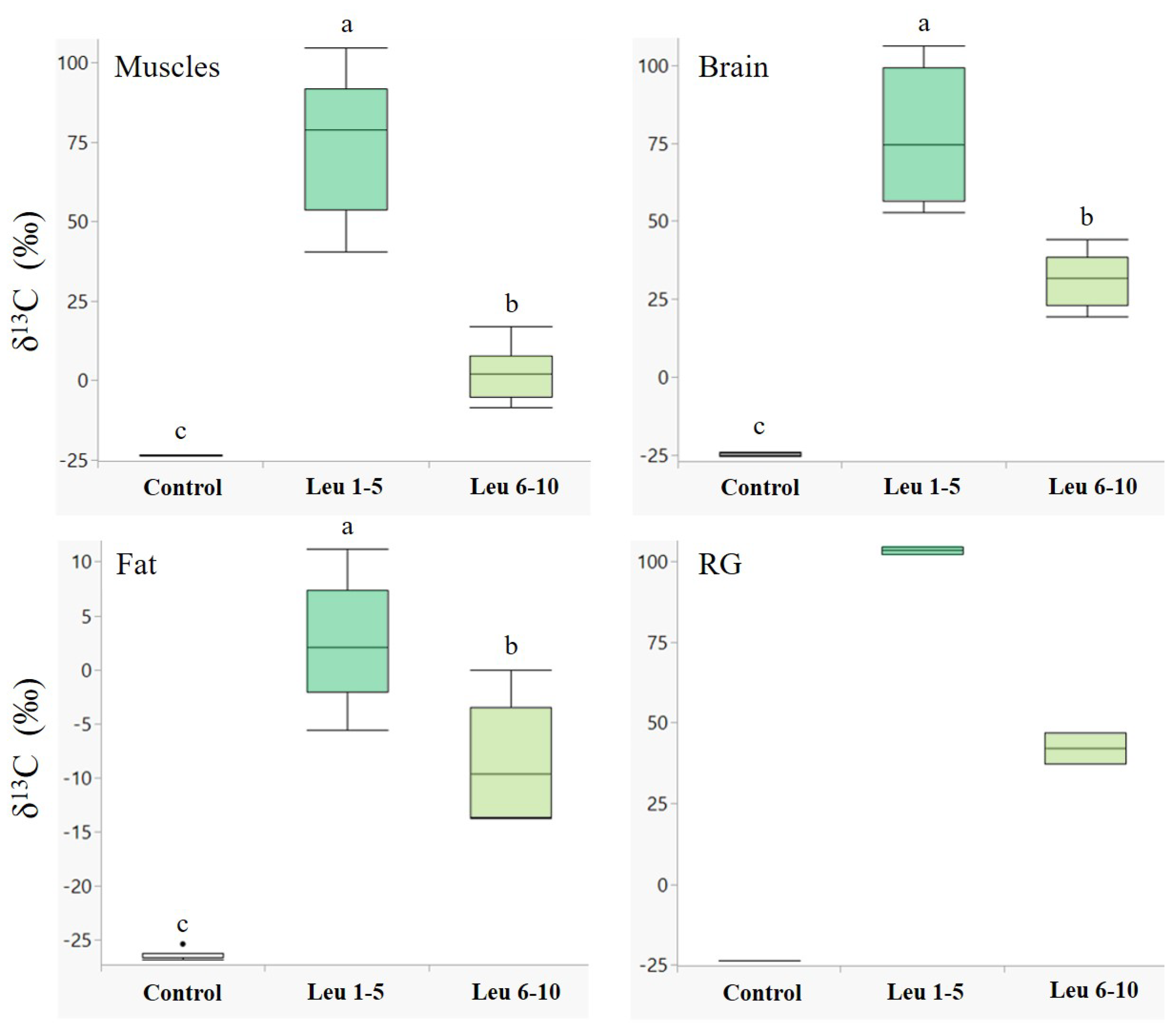
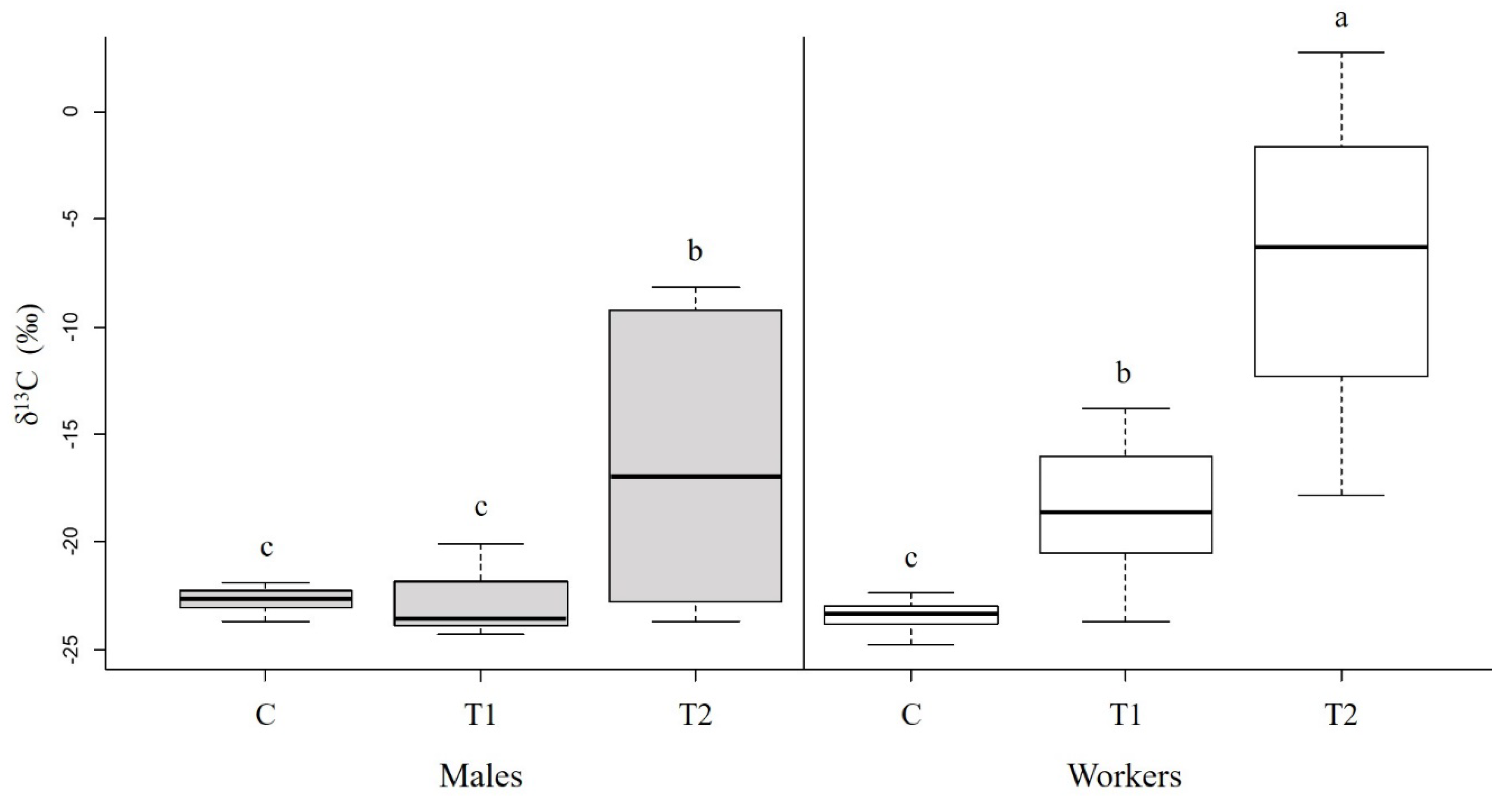
3.1. Nutrient Allocation and Incorporation in Males
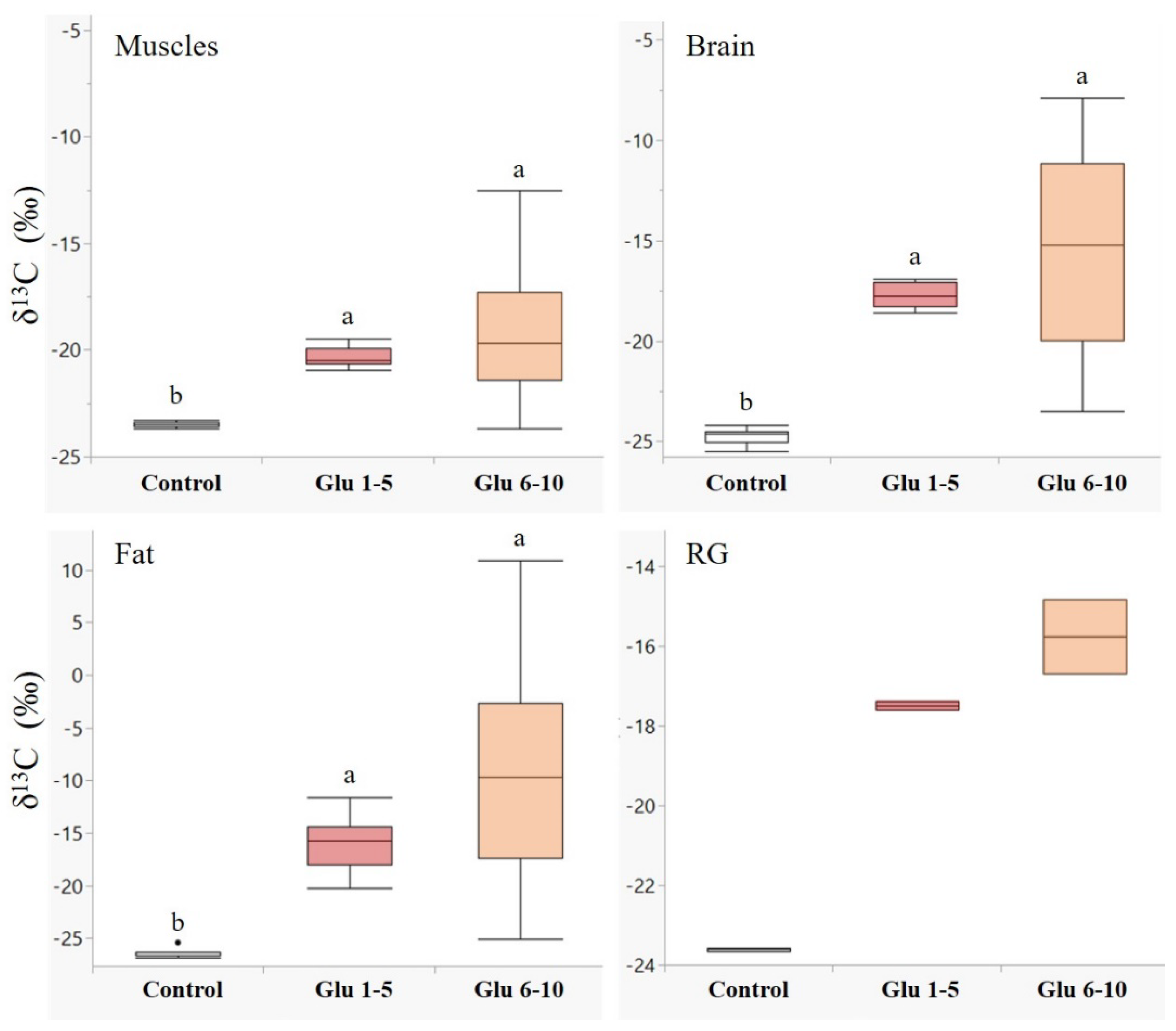
3.2. Protein Digestion in Adult Hornets
4. Discussion
4.1. Nutrient Allocation and Incorporation in Males
4.2. Protein Digestion in Adult Hornets
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rangel, J.; Fisher, A. Factors Affecting the Reproductive Health of Honey Bee (Apis mellifera) Drones—A Review. Apidologie 2019, 50, 759–778. [Google Scholar] [CrossRef]
- Belsky, J.E.; Camp, A.A.; Lehmann, D.M. The Importance of Males to Bumble Bee (Bombus Species) Nest Development and Colony Viability. Insects 2020, 11, 506. [Google Scholar] [CrossRef] [PubMed]
- Martin, S. A Simulation Model for Colony Development of the Hornet Vespa simillima (Hymenoptera: Vespidae). Jpn. J. Entomol. 1991, 59, 105–124. [Google Scholar]
- Martin, S.J. Weight Changes in Adult Hornets, Vespa affinis (Hymenoptera: Vespidae). Insectes Sociaux 1993, 40, 363–368. [Google Scholar] [CrossRef]
- Harano, K.-I.; Sasaki, K. Sexual Maturation, Mating Strategies and Neuroendocrinology in Social Insects. In Human and Animal Mating; Kakamura, M., Ito, T., Eds.; Novinka: New York, NY, USA, 2013; pp. 31–54. [Google Scholar]
- Stürup, M.; Baer-Imhoof, B.; Nash, D.R.; Boomsma, J.J.; Baer, B. When Every Sperm Counts: Factors Affecting Male Fertility in the Honeybee Apis mellifera. Behav. Ecol. 2013, 24, 1192–1198. [Google Scholar] [CrossRef]
- Hunt, J. Nourishment and the Evolution of the Social Vespidae. In The Social Biology of Wasps; Ross, K.G., Matthews, R.W., Eds.; Cornell University Press: Ithaca, NY, USA, 1991; pp. 426–450. ISBN 978-1-5017-1867-0. [Google Scholar]
- Abe, T.; Yoshiya, T.; Hiromitsu, M.; Yasuko, Y.K. Comparative Study of the Composition of Hornet Larval Saliva, its Effect on Behaviour and Role of Trophallaxis. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1991, 99, 79–84. [Google Scholar] [CrossRef]
- Ishay, J.; Ikan, R. Gluconeogenesis in the Oriental hornet Vespa orientalis F. Ecology 1968, 49, 169–171. [Google Scholar] [CrossRef]
- Ishay, J.; Ikan, R. Food Exchange between Adults and Larvae in Vespa orientalis F. Anim. Behav. 1968, 16, 298–303. [Google Scholar] [CrossRef]
- Jeong, H.; Kim, J.M.; Kim, B.; Nam, J.-O.; Hahn, D.; Choi, M.B. Nutritional Value of the Larvae of the Alien Invasive Wasp Vespa velutina nigrithorax and Amino Acid Composition of the Larval Saliva. Foods 2020, 9, 885. [Google Scholar] [CrossRef]
- Ikan, R.; Bergmann, E.D.; Ishay, J.; Gitter, S. Proteolytic Enzyme Activity in the Various Colony Members of the Oriental hornet, Vespa Orientalis, F. Life Sci. 1968, 7, 929–934. [Google Scholar] [CrossRef]
- Grogan, D.E.; Hunt, J.H. Midgut Endopeptidase Activities of the Hornet Vespa crabro germana Christ (Hymenoptera: Vespidae). Insectes Sociaux 1986, 33, 486–489. [Google Scholar] [CrossRef]
- Bourke, A.F.G. Worker Reproduction in the Higher Eusocial Hymenoptera. Q. Rev. Biol. 1988, 63, 291–311. [Google Scholar] [CrossRef]
- Bodenheimer, F.S. Über Die Aktivität von Vespa orientalis L. im Jahresverlauf in Palaestina. Zool. Anz. 1933, 102, 135–140. [Google Scholar]
- Carpenter, J.; Kojima, J. Checklist of the Species in the Subfamily Vespinae (Insecta: Hymenoptera: Vespidae). Nat. Hist. Bull. Ibaraki Univ. 1997, 1, 51–92. [Google Scholar]
- Perrard, A.; Pickett, K.; Villemant, C.; Kojima, J.; Carpenter, J. Phylogeny of Hornets: A Total Evidence Approach (Hymenoptera, Vespidae, Vespinae, Vespa). J. Hymenopt. Res 2013, 32, 1–15. [Google Scholar] [CrossRef]
- Volov, M.; Cohen, N.; Bodner, L.; Dubiner, S.; Hefetz, A.; Bouchebti, S.; Levin, E. The Effect of Climate and Diet on Body Lipid Composition in the Oriental Hornet (Vespa orientalis). Front. Ecol. Evol. 2021, 9, 669. [Google Scholar] [CrossRef]
- Ishay, J.; Bytinski-Salz, H.; Shulov, A. Contributions to the Bionomics of the Oriental hornet (Vespa orientalis Fab.). Isr. J. Entomol. 1967, 2, 45–106. [Google Scholar]
- Dehghani, R.; Kassiri, H.; Mazaheri-Tehrani, A.; Hesam, M.; Valazadi, N.; Mohammadzadeh, M. A Study on Habitats and Behavioral Characteristics of Hornet Wasp (Hymenoptera: Vespidae: Vespa orientalis), an Important Medical-Health Pest. Biomed. Res. 2019, 30, 6–61. [Google Scholar]
- Havron, A.; Margalith, Y. Parasitization of Vespa orientalis Nests by Sphecophaga vesparum Curtis in Southern Israel (Hymenoptera: Vespidae, Ichneumonidae). Phytoparasitica 1995, 23, 19–25. [Google Scholar] [CrossRef]
- Temreshev, I.I. On the expansion of the areas of Vespa orientalis and Polistes wattii (Hymenoptera: Vespidae) on the territory of the Republic of Kazakhstan. Acta Biol. Sib. 2018, 4, 38. [Google Scholar] [CrossRef]
- Sánchez, I.; Fajardo, M.; Castro, M. Primeras Citas Del Avispón Oriental Vespa orientalis Linnaeus 1771 (Hymenoptera: Vespidae) Para Andalucía (España). Rev. Soc. Gaditana Hist. Nat. 2019, 13, 11–14. [Google Scholar]
- Bressi, N.; Colla, A.; Tomasin, G. Orientali Verso Nord: Insediamento Di Una Popolazione Urbana Di Calabrone Orientale (Vespa orientalis) a Trieste (NE Italy). Atti del Mus. Civ. di Stor. Nat. di Trieste 2019, 60, 273–275. [Google Scholar]
- Gereys, B.; Coache, A.; Filippi, G. Présence En France Métropolitaine d’un Frelon Allochtone: Vespa orientalis Linnaeus, 1771 (Le Frelon Oriental) (Hymenoptera, Vespidae, Vespinae). Faunitaxys 2021, 9, 1–5. [Google Scholar]
- Zachi, M.; Ruicănescu, A. Vespa orientalis, a New Alien Species in Romania. Travaux 2021, 64, 67–72. [Google Scholar] [CrossRef]
- Dvořák, L. Oriental hornet Vespa orientalis Linnaeus, 1771 Found in Mexico (Hymenoptera, Vespidae, Vespinae). Entomol. Probl. 2006, 36, 80. [Google Scholar]
- Ríos, M.; Barrera-Medina, R.; Contreras, J.M. Primer Reporte Del Género Vespa Linnaeus (Hymenoptera: Vespidae: Vespinae) En Chile. Rev. Chil. Entomol. 2020, 46, 237–242. [Google Scholar] [CrossRef]
- Werenkraut, V.; Arbetman, M.P.; Fergnani, P.N. The Oriental hornet (Vespa orientalis L.): A Threat to the Americas? Neotrop. Entomol. 2021, 1–9. [Google Scholar] [CrossRef]
- McCue, M.D. Tracking the Oxidative and Nonoxidative Fates of Isotopically Labeled Nutrients in Animals. BioScience 2011, 61, 217–230. [Google Scholar] [CrossRef]
- Poidatz, J.; Bressac, C.; Bonnard, O.; Thiéry, D. Delayed Sexual Maturity in Males of Vespa velutina. Insect Sci. 2018, 25, 679–689. [Google Scholar] [CrossRef]
- Crosson, E.R.; Ricci, K.N.; Richman, B.A.; Chilese, F.C.; Owano, T.G.; Provencal, R.A.; Todd, M.W.; Glasser, J.; Kachanov, A.A.; Paldus, B.A.; et al. Stable Isotope Ratios Using Cavity Ring-Down Spectroscopy: Determination of 13C/12C for Carbon Dioxide in Human Breath. Anal. Chem. 2002, 74, 2003–2007. [Google Scholar] [CrossRef]
- Levin, E.; Mitra, C.; Davidowitz, G. Fed Males Increase Oviposition in Female Hawkmoths via Non-Nutritive Direct Benefits. Anim. Behav. 2016, 112, 111–118. [Google Scholar] [CrossRef]
- Levin, E.; McCue, M.D.; Davidowitz, G. Sex Differences in the Utilization of Essential and Non-Essential Amino Acids in Lepidoptera. J. Exp. Biol. 2017, 220, 2743–2747. [Google Scholar] [CrossRef] [PubMed]
- Werner, R.A.; Brand, W.A. Referencing Strategies and Techniques in Stable Isotope Ratio Analysis. Rapid Commun. Mass Spectrom. 2001, 15, 501–519. [Google Scholar] [CrossRef]
- Hrassnigg, N.; Crailsheim, K. Differences in Drone and Worker Physiology in Honeybees (Apis mellifera). Apidologie 2005, 36, 255–277. [Google Scholar] [CrossRef]
- Berger, B.; Crailsheim, K.; Leonhard, B. Proline, Leucine and Phenylalanine Metabolism in Adult Honeybee Drones (Apis mellifica carnica Pollm). Insect Biochem. Mol. Biol. 1997, 27, 587–593. [Google Scholar] [CrossRef]
- Colonello, N.A.; Hartfelder, K. Protein Content and Pattern during Mucus Gland Maturation and Its Ecdysteroid Control in Honey Bee Drones. Apidologie 2003, 34, 257–267. [Google Scholar] [CrossRef]
- Fahrbach, S.E.; Giray, T.; Farris, S.M.; Robinson, G.E. Expansion of the Neuropil of the Mushroom Bodies in Male Honey Bees Is Coincident with Initiation of Flight. Neurosci. Lett. 1997, 236, 135–138. [Google Scholar] [CrossRef]
- Panzenböck, U.; Crailsheim, K. Glycogen in Honeybee Queens, Workers and Drones (Apis mellifera carnica Pollm.). J. Insect Physiol. 1997, 43, 155–165. [Google Scholar] [CrossRef]
- Levin, E.; Lopez-Martinez, G.; Fane, B.; Davidowitz, G. Hawkmoths Use Nectar Sugar to Reduce Oxidative Damage from Flight. Science 2017, 355, 733–735. [Google Scholar] [CrossRef]
- Tsacopoulos, M.; Veuthey, A.L.; Saravelos, S.G.; Perrottet, P.; Tsoupras, G. Glial Cells Transform Glucose to Alanine, Which Fuels the Neurons in the Honeybee Retina. J. Neurosci. 1994, 14, 1339–1351. [Google Scholar] [CrossRef]
- Tsacopoulos, M.; Veuthey, A.-L. The Nutritive Function of Glia in a Crystal-Like Nervous Tissue: The Retina of the Honeybee Drone. Dev. Neurosci. 1993, 15, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Arrese, E.L.; Soulages, J.L. Insect Fat Body: Energy, Metabolism, and Regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Landim, C.; Dallacqua, R.P. Morphology and Protein Patterns of Honey Bee Drone Accessory Glands. Genet. Mol. Res. Electron. Resour. GMR 2005, 4, 473–481. [Google Scholar]
- Moors, L.; Schoeters, E.; Coudron, K.; Billen, J. Morphological Changes in the Male Accessory Glands and Testes in Vespula vulgaris (Hymenoptera, Vespidae) during Sexual Maturation. Invertebr. Biol. 2009, 128, 364–371. [Google Scholar] [CrossRef]
- Moors, L.; Spaas, O.; Koeniger, G.; Billen, J. Morphological and Ultrastructural Changes in the Mucus Glands of Apis mellifera Drones during Pupal Development and Sexual Maturation. Apidologie 2005, 36, 245–254. [Google Scholar] [CrossRef][Green Version]
- Alumot, E.; Lensky, Y.; Holstein, P. Sugars and Trehalase in the Reproductive Organs and Hemolymph of the Queen and Drone Honey Bees (Apis mellifica L. var. Ligustica spin.). Comp. Biochem. Physiol. 1969, 28, 1419–1425. [Google Scholar] [CrossRef]
- Avila, F.W.; Sirot, L.K.; LaFlamme, B.A.; Rubinstein, C.D.; Wolfner, M.F. Insect Seminal Fluid Proteins: Identification and Function. Annu. Rev. Entomol. 2011, 56, 21–40. [Google Scholar] [CrossRef]
- Baer, B.; Morgan, E.D.; Schmid-Hempel, P. A Nonspecific Fatty Acid within the Bumblebee Mating Plug Prevents Females from Remating. Proc. Natl. Acad. Sci. USA 2001, 98, 3926–3928. [Google Scholar] [CrossRef]
- Baer, B.; Heazlewood, J.L.; Taylor, N.L.; Eubel, H.; Millar, A.H. The Seminal Fluid Proteome of the Honeybee Apis mellifera. Proteomics 2009, 9, 2085–2097. [Google Scholar] [CrossRef]
- Blum, M.S.; Glowska, Z.; Taber, S., III. Chemistry of the Drone Honey Bee Reproductive System. II. Carbohydrates in the Reproductive Organs and Semen. Ann. Entomol. Soc. Am. 1962, 55, 135–139. [Google Scholar] [CrossRef]
- Jasper, W.C.; Brutscher, L.M.; Grozinger, C.M.; Niño, E.L. Injection of Seminal Fluid into the Hemocoel of Honey Bee Queens (Apis mellifera) Can Stimulate Post-Mating Changes. Sci. Rep. 2020, 10, 11990. [Google Scholar] [CrossRef] [PubMed]
- King, M.; Eubel, H.; Millar, A.H.; Baer, B. Proteins within the Seminal Fluid Are Crucial to Keep Sperm Viable in the Honeybee Apis mellifera. J. Insect Physiol. 2011, 57, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Liberti, J.; Görner, J.; Welch, M.; Dosselli, R.; Schiøtt, M.; Ogawa, Y.; Castleden, I.; Hemmi, J.M.; Baer-Imhoof, B.; Boomsma, J.J.; et al. Seminal Fluid Compromises Visual Perception in Honeybee Queens Reducing Their Survival during Additional Mating Flights. eLife 2019, 8, e45009. [Google Scholar] [CrossRef]
- Poole, H.K.; Edwards, J.F. Induction of Motility in Honey Bee (Apis mellifera L.) Spermatozoa by Sugars. Experientia 1970, 26, 859–860. [Google Scholar] [CrossRef] [PubMed]
- Verma, L.R.; Shuel, R.W. Respiratory Metabolism of the Semen of the Honey-Bee, Apis mellifera. J. Insect Physiol. 1973, 19, 97–103. [Google Scholar] [CrossRef]
- Czekońska, K.; Chuda-Mickiewicz, B.; Samborski, J. Quality of Honeybee Drones Reared in Colonies with Limited and Unlimited Access to Pollen. Apidologie 2015, 46, 1–9. [Google Scholar] [CrossRef]
- Rousseau, A.; Giovenazzo, P. Optimizing Drone Fertility with Spring Nutritional Supplements to Honey Bee (Hymenoptera: Apidae) Colonies. J. Econ. Entomol. 2016, 109, 1009–1014. [Google Scholar] [CrossRef]
- Szentgyörgyi, H.; Czekońska, K.; Tofilski, A. The Effects of Starvation of Honey Bee Larvae on Reproductive Quality and Wing Asymmetry of Honey Bee Drones. J. Apic. Sci. 2017, 61, 233–243. [Google Scholar] [CrossRef]
- Bishop, G.H. Fertilization in the Honey-Bee. I. The Male Sexual Organs: Their Histological Structure and Physiological Functioning. J. Exp. Zool. 1920, 31, 224–265. [Google Scholar] [CrossRef]
- Hoage, T.R.; Kessel, R.G. An Electron Microscope Study of the Process of Differentiation during Spermatogenesis in the Drone Honey Bee (Apis mellifera L.) with Special Reference to Centriole Replication and Elimination. J. Ultrastruct. Res. 1968, 24, 6–32. [Google Scholar] [CrossRef]
- Montagner, H. Etudes du comportement alimentaire et des relations trophallactiques du mâle au sein de la société de guêpes. Insectes Sociaux 1964, 11, 301–316. [Google Scholar] [CrossRef]
- Grogan, D.E.; Hunt, J.H. Digestive Proteases of Two Species of Wasps of the Genus Vespula. Insect Biochem. 1977, 7, 191–196. [Google Scholar] [CrossRef]
- Cassill, D.L.; Butler, J.; Vinson, S.B.; Wheeler, D.E. Cooperation during Prey Digestion between Workers and Larvae in the Ant, Pheidole spadonia. Insectes Sociaux 2005, 52, 339–343. [Google Scholar] [CrossRef][Green Version]
- Petralia, R.S.; Sorensen, A.A.; Vinson, S.B. The Labial Gland System of Larvae of the Imported Fire Ant, Solenopsis invicta Buren: Ulfrastructure and Enzyme Analysis. Cell Tissue Res. 1980, 206, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, A.A.; Kamas, R.S.; Vinson, S.B. The Influence of Oral Secretions from Larvae on Levels of Proteinases in Colony Members of Solenopsis invicta Buren (Hymenoptera: Formicidae). J. Insect Physiol. 1983, 29, 163–168. [Google Scholar] [CrossRef]
- Ikan, R.; Ishay, J. Larval Wasps Secretions and Honeydew of the Aphids, Chaitophorus populi, Feeding on Populus euphratica as Sources of Sugars in the Diet of the Oriental hornet, Vespa orientalis, F. Isr. J. Zool. 1966, 15, 64–68. [Google Scholar] [CrossRef]
- Fateryga, A.V. Trophic Relations between Vespid Wasps (Hymenoptera, Vespidae) and Flowering Plants in the Crimea. Entomol. Rev. 2010, 90, 698–705. [Google Scholar] [CrossRef]
- Matsuura, M.; Yamane, S. Biology of the Vespine Wasps. In Biology of the Vespine Wasps; Springer: Berlin, Germany, 1990. [Google Scholar]
- Ueno, T. Flower-Visiting by the Invasive Hornet Vespa velutina nigrithorax (Hymenoptera: Vespidae). Int. J. Chem. Environ. Biol. Sci. 2015, 3, 444–448. [Google Scholar]
- Baker, H.G. Non-sugar Chemical Constituents of Nectar. Apidologie 1977, 8, 349–356. [Google Scholar] [CrossRef]
- Nicolson, S.; Thornburg, R. Nectar Chemistry. In Nectaries and Nectar; Springer: Dordrecht, The Netherlands, 2007; pp. 215–264. ISBN 978-1-4020-5936-0. [Google Scholar]
- Hunt, J.H.; Baker, I.; Baker, H.G. Similarity of Amino Acids in Nectar and Larval Saliva: The Nutritional Basis for Trophallaxis in Social Wasps. Evolution 1982, 36, 1318–1322. [Google Scholar] [CrossRef]
- Bodner, L.; Bouchebti, S.; Levin, E. Allocation and Metabolism of Naturally Occurring Dietary Amino Acids in the Oriental hornet. Insect Biochem. Mol. Biol. 2021, 139, 103675. [Google Scholar] [CrossRef] [PubMed]
- Bursell, E. Aspects of the Metabolism of Amino Acids in the Tsetse Fly, Glossina (Diptera). J. Insect Physiol. 1963, 9, 439–452. [Google Scholar] [CrossRef]
- Micheu, S.; Crailsheim, K.; Leonhard, B. Importance of Proline and Other Amino Acids during Honeybee Flight. Amino Acids 2000, 18, 157–175. [Google Scholar] [CrossRef]
- Teulier, L.; Weber, J.-M.; Crevier, J.; Darveau, C.-A. Proline as a Fuel for Insect Flight: Enhancing Carbohydrate Oxidation in Hymenopterans. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160333. [Google Scholar] [CrossRef] [PubMed]
- Ishay, J. Observations sur la biologie de la guêpe orientale Vespa orientalis F. Insectes Sociaux 1964, 11, 193–206. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bodner, L.; Bouchebti, S.; Watted, O.; Seltzer, R.; Drabkin, A.; Levin, E. Nutrient Utilization during Male Maturation and Protein Digestion in the Oriental Hornet. Biology 2022, 11, 241. https://doi.org/10.3390/biology11020241
Bodner L, Bouchebti S, Watted O, Seltzer R, Drabkin A, Levin E. Nutrient Utilization during Male Maturation and Protein Digestion in the Oriental Hornet. Biology. 2022; 11(2):241. https://doi.org/10.3390/biology11020241
Chicago/Turabian StyleBodner, Levona, Sofia Bouchebti, Omar Watted, Rya Seltzer, Ariel Drabkin, and Eran Levin. 2022. "Nutrient Utilization during Male Maturation and Protein Digestion in the Oriental Hornet" Biology 11, no. 2: 241. https://doi.org/10.3390/biology11020241
APA StyleBodner, L., Bouchebti, S., Watted, O., Seltzer, R., Drabkin, A., & Levin, E. (2022). Nutrient Utilization during Male Maturation and Protein Digestion in the Oriental Hornet. Biology, 11(2), 241. https://doi.org/10.3390/biology11020241