
Diversity and Evolution of DNA Transposons Targeting Multicopy Small RNA Genes from Actinopterygian Fish


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Genome Survey
2.2. Transcriptome Survey
2.3. Phylogenetic Analysis
3. Results
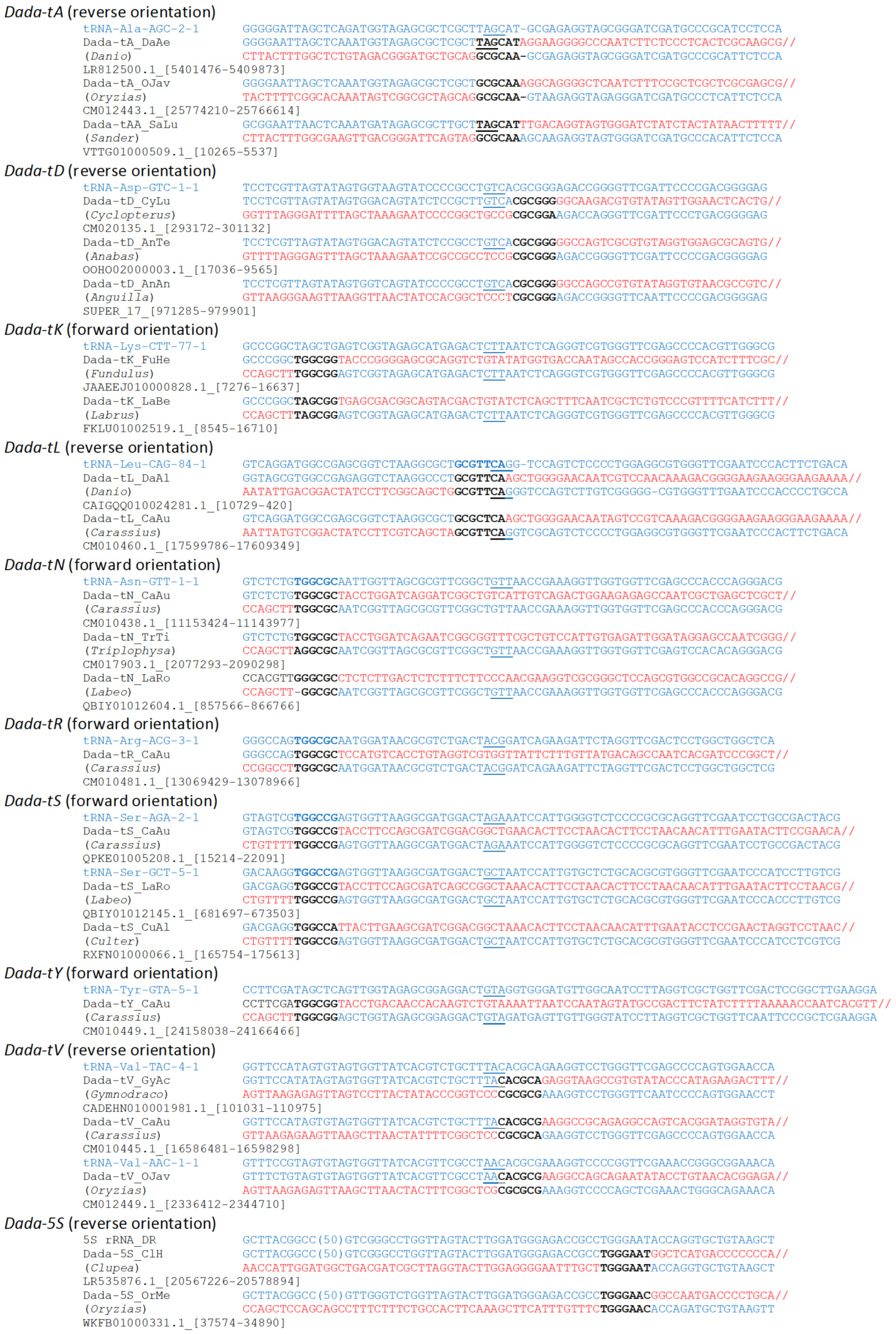
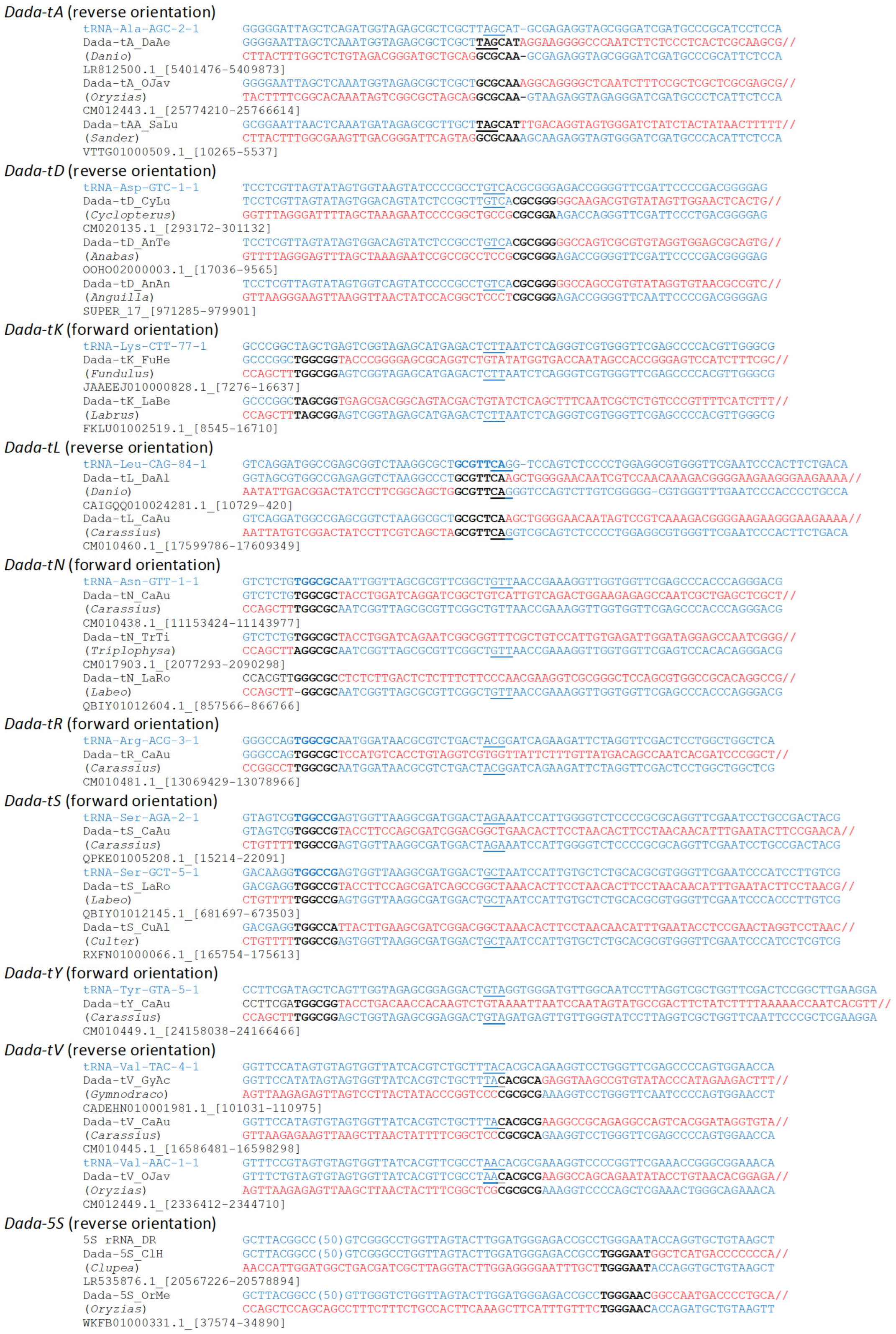
3.1. Wide Distribution and Various Target Specificities in Actinopterygian Dada Families
3.2. Transcriptome Analysis
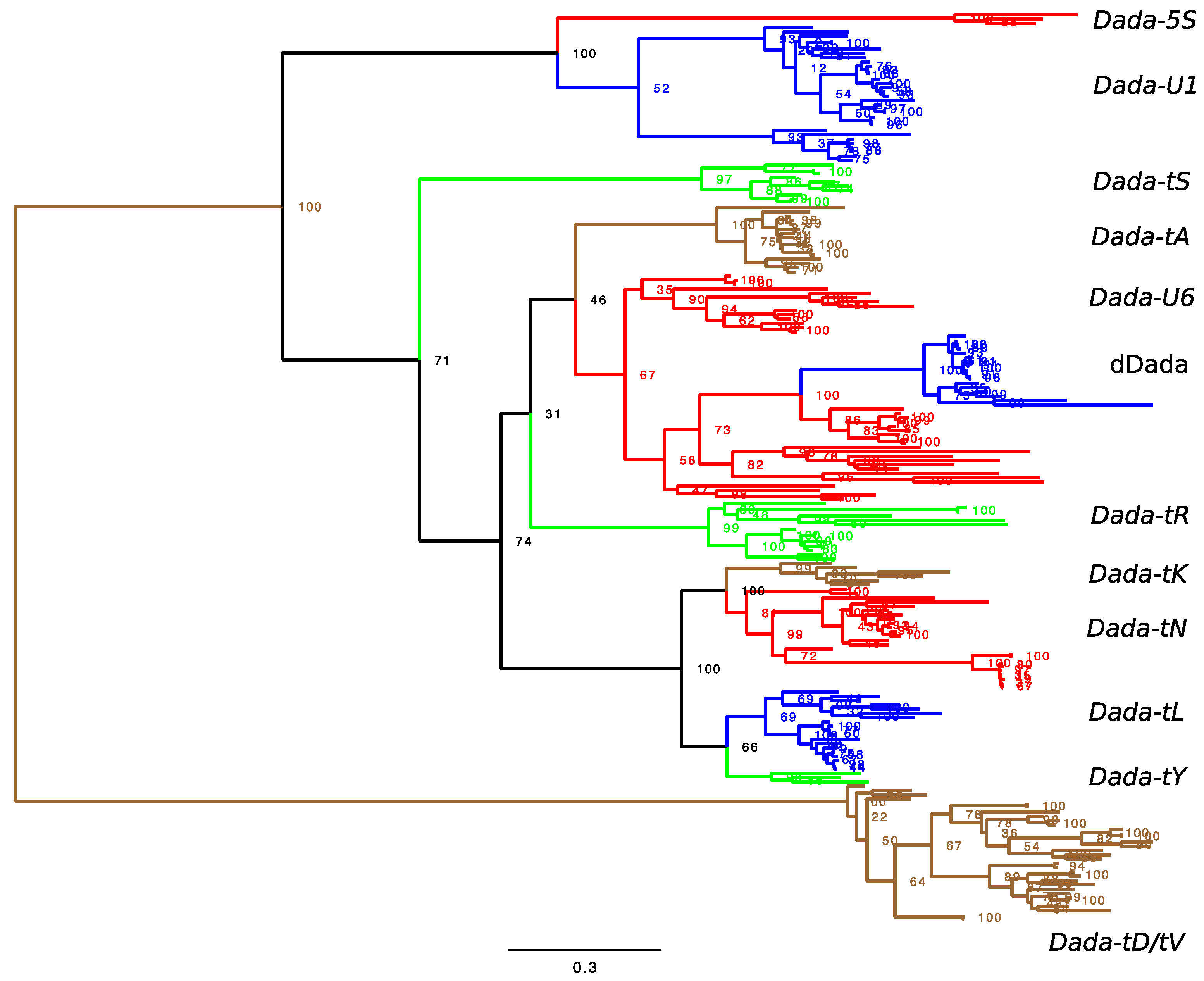
3.3. Phylogenetic Analysis
3.4. dDada, Domesticated Dada-U6 in Cypriniformes
4. Discussion
4.1. Distributions of Target-Specific Dada Lineages
4.2. Target Selection and Evolution of Dada
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Curcio, M.J.; Derbyshire, K.M. The outs and ins of transposition: From mu to kangaroo. Nat. Rev. Mol. Cell Biol. 2003, 4, 865–877. [Google Scholar] [CrossRef]
- Kojima, K.K. Structural and sequence diversity of eukaryotic transposable elements. Genes Genet. Syst. 2020, 94, 233–252. [Google Scholar] [CrossRef] [Green Version]
- Arkhipova, I.R. Using bioinformatic and phylogenetic approaches to classify transposable elements and understand their complex evolutionary histories. Mob. DNA 2017, 8, 19. [Google Scholar] [CrossRef]
- Finnegan, D.J. Eukaryotic transposable elements and genome evolution. Trends Genet. 1989, 5, 103–107. [Google Scholar] [CrossRef]
- Glockner, G.; Szafranski, K.; Winckler, T.; Dingermann, T.; Quail, M.A.; Cox, E.; Eichinger, L.; Noegel, A.A.; Rosenthal, A. The complex repeats of Dictyostelium discoideum. Genome Res. 2001, 11, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, T.J.; Poulter, R.T. A new group of tyrosine recombinase-encoding retrotransposons. Mol. Biol. Evol. 2004, 21, 746–759. [Google Scholar] [CrossRef]
- Evgen’ev, M.B.; Zelentsova, H.; Shostak, N.; Kozitsina, M.; Barskyi, V.; Lankenau, D.H.; Corces, V.G. Penelope, a new family of transposable elements and its possible role in hybrid dysgenesis in Drosophila virilis. Proc. Natl. Acad. Sci. USA 1997, 94, 196–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evgen’ev, M.B.; Arkhipova, I.R. Penelope-like elements--a new class of retroelements: Distribution, function and possible evolutionary significance. Cytogenet. Genome Res. 2005, 110, 510–521. [Google Scholar] [CrossRef]
- Hickman, A.B.; Chandler, M.; Dyda, F. Integrating prokaryotes and eukaryotes: DNA transposases in light of structure. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 50–69. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Breitbart, M.; Edwards, R.A. Transposases are the most abundant, most ubiquitous genes in nature. Nucleic Acids Res. 2010, 38, 4207–4217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, T.J.; Butler, M.I.; Poulter, R.T. Cryptons: A group of tyrosine-recombinase-encoding DNA transposons from pathogenic fungi. Microbiology 2003, 149, 3099–3109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapitonov, V.V.; Jurka, J. Rolling-circle transposons in eukaryotes. Proc. Natl. Acad. Sci. USA 2001, 98, 8714–8719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, W.; Kojima, K.K.; Kohany, O. Repbase Update, a database of repetitive elements in eukaryotic genomes. Mob. DNA 2015, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.W.; Wessler, S.R. The catalytic domain of all eukaryotic cut-and-paste transposase superfamilies. Proc. Natl. Acad. Sci. USA 2011, 108, 7884–7889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, K.K.; Jurka, J. A superfamily of DNA transposons targeting multicopy small RNA genes. PLoS ONE 2013, 8, e68260. [Google Scholar] [CrossRef] [Green Version]
- Babushok, D.V.; Kazazian, H.H., Jr. Progress in understanding the biology of the human mutagen LINE-1. Hum. Mutat. 2007, 28, 527–539. [Google Scholar] [CrossRef]
- Belancio, V.P.; Hedges, D.J.; Deininger, P. Mammalian non-LTR retrotransposons: For better or worse, in sickness and in health. Genome Res. 2008, 18, 343–358. [Google Scholar] [CrossRef] [Green Version]
- Kojima, K.K.; Fujiwara, H. Cross-genome screening of novel sequence-specific non-LTR retrotransposons: Various multicopy RNA genes and microsatellites are selected as targets. Mol. Biol. Evol. 2004, 21, 207–217. [Google Scholar] [CrossRef]
- Kojima, K.K.; Jurka, J. Ancient Origin of the U2 Small Nuclear RNA Gene-Targeting Non-LTR Retrotransposons Utopia. PLoS ONE 2015, 10, e0140084. [Google Scholar] [CrossRef] [Green Version]
- Starnes, J.H.; Thornbury, D.W.; Novikova, O.S.; Rehmeyer, C.J.; Farman, M.L. Telomere-targeted retrotransposons in the rice blast fungus Magnaporthe oryzae: Agents of telomere instability. Genetics 2012, 191, 389–406. [Google Scholar] [CrossRef] [Green Version]
- Malik, H.S.; Eickbush, T.H. NeSL-1, an ancient lineage of site-specific non-LTR retrotransposons from Caenorhabditis elegans. Genetics 2000, 154, 193–203. [Google Scholar] [CrossRef]
- Aksoy, S.; Williams, S.; Chang, S.; Richards, F.F. SLACS retrotransposon from Trypanosoma brucei gambiense is similar to mammalian LINEs. Nucleic Acids Res. 1990, 18, 785–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, S.; Ishikawa, H.; Fujiwara, H. Structural analysis of TRAS1, a novel family of telomeric repeat-associated retrotransposons in the silkworm, Bombyx mori. Mol. Cell Biol. 1995, 15, 4545–4552. [Google Scholar] [CrossRef] [Green Version]
- Christensen, S.; Pont-Kingdon, G.; Carroll, D. Target specificity of the endonuclease from the Xenopus laevis non-long terminal repeat retrotransposon, Tx1L. Mol. Cell Biol. 2000, 20, 1219–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Q.; Schumann, G.; Boeke, J.D. Retrotransposon R1Bm endonuclease cleaves the target sequence. Proc. Natl. Acad. Sci. USA 1998, 95, 2083–2088. [Google Scholar] [CrossRef] [Green Version]
- Christensen, S.M.; Bibillo, A.; Eickbush, T.H. Role of the Bombyx mori R2 element N-terminal domain in the target-primed reverse transcription (TPRT) reaction. Nucleic Acids Res. 2005, 33, 6461–6468. [Google Scholar] [CrossRef] [Green Version]
- Anzai, T.; Takahashi, H.; Fujiwara, H. Sequence-specific recognition and cleavage of telomeric repeat (TTAGG)(n) by endonuclease of non-long terminal repeat retrotransposon TRAS1. Mol. Cell Biol. 2001, 21, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Malik, H.S.; Burke, W.D.; Eickbush, T.H. The age and evolution of non-LTR retrotransposable elements. Mol. Biol. Evol. 1999, 16, 793–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, D.D.; Korman, M.H.; Jakubczak, J.L.; Eickbush, T.H. Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal target site: A mechanism for non-LTR retrotransposition. Cell 1993, 72, 595–605. [Google Scholar] [CrossRef]
- Kojima, K.K.; Fujiwara, H. Long-term inheritance of the 28S rDNA-specific retrotransposon R2. Mol. Biol. Evol. 2005, 22, 2157–2165. [Google Scholar] [CrossRef] [Green Version]
- Burke, W.D.; Malik, H.S.; Lathe, W.C., 3rd; Eickbush, T.H. Are retrotransposons long-term hitchhikers? Nature 1998, 392, 141–142. [Google Scholar] [CrossRef]
- Penton, E.H.; Sullender, B.W.; Crease, T.J. Pokey, a new DNA transposon in Daphnia (cladocera: Crustacea). J. Mol. Evol. 2002, 55, 664–673. [Google Scholar] [CrossRef]
- Kapitonov, V.V.; Jurka, J. CryptonV, a group of target-site specific Crypton DNA transposons from cnidarians. Repbase Rep. 2012, 12, 2034. [Google Scholar]
- Strecker, J.; Ladha, A.; Gardner, Z.; Schmid-Burgk, J.L.; Makarova, K.S.; Koonin, E.V.; Zhang, F. RNA-guided DNA insertion with CRISPR-associated transposases. Science 2019, 365, 48–53. [Google Scholar] [CrossRef]
- Kohany, O.; Gentles, A.J.; Hankus, L.; Jurka, J. Annotation, submission and screening of repetitive elements in Repbase: RepbaseSubmitter and Censor. BMC Bioinform. 2006, 7, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solovyev, V.; Kosarev, P.; Seledsov, I.; Vorobyev, D. Automatic annotation of eukaryotic genes, pseudogenes and promoters. Genome Biol. 2006, 7, S10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stout, C.C.; Tan, M.; Lemmon, A.R.; Lemmon, E.M.; Armbruster, J.W. Resolving Cypriniformes relationships using an anchored enrichment approach. BMC Evol. Biol. 2016, 16, 244. [Google Scholar] [CrossRef] [Green Version]
- Chan, P.P.; Lowe, T.M. GtRNAdb 2.0: An expanded database of transfer RNA genes identified in complete and draft genomes. Nucleic Acids Res. 2016, 44, D184–D189. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Order | Family 1 | Dada Lineages with Confirmed Target Sequences | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| U6 | tA | tY | tL | tN | tK | tR | tS | U1 | 5S | th | tV | ||
| Anguilliformes | Anguillidae (1) | + | |||||||||||
| Clupeiformes | Clupeidae (1) | + | |||||||||||
| Engraulidae (1) | + | + | + | ||||||||||
| Cypriniformes | Cyprinidae (8) | + | + | + | + | + | + | + | + | + | + | ||
| Danionidae (8) | + | + | + | + | + | ||||||||
| Nemacheilidae (1) | + | + | + | ||||||||||
| Characiformes | Characidae (1) | + | |||||||||||
| Siluriformes | Ictaluridae (1) | + | |||||||||||
| Pangasiidae (1) | + | ||||||||||||
| Salmoniformes | Salmonidae (1) | + | |||||||||||
| Gadiformes | Gadidae (1) | + | + | + | + | ||||||||
| Scombriformes | Scombridae (1) | + | |||||||||||
| Anabantiformes | Anabantidae (2) | + | + | ||||||||||
| Carangiformes | Carangidae (1) | + | + | ||||||||||
| Ovalentaria incertae sedis | Ambassidae (1) | + | + | ||||||||||
| Mugiliformes | Mugilidae (1) | + | |||||||||||
| Cichliformes | Cichlidae (2) | + | + | ||||||||||
| Beloniformes | Adrianichthyidae (3) | + | + | + | + | + | + | ||||||
| Cyprinodontiformes | Fundulidae (1) | + | |||||||||||
| Cyprinodontidae (1) | + | ||||||||||||
| Poeciliidae (1) | + | ||||||||||||
| Perciformes | Percidae (5) | + | + | + | + | + | + | + | + | ||||
| Sciaenidae (1) | + | + | |||||||||||
| Cyclopteridae (1) | + | ||||||||||||
| Nototheniidae (2) | + | + | + | ||||||||||
| Gasterosteidae (1) | + | ||||||||||||
| Labriformes | Labridae (2) | + | + | + | |||||||||
| Spariformes | Sparidae (2) | + | + | + | |||||||||
| Tetraodontiformes | Tetraodontidae (1) | + | |||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kojima, K.K. Diversity and Evolution of DNA Transposons Targeting Multicopy Small RNA Genes from Actinopterygian Fish. Biology 2022, 11, 166. https://doi.org/10.3390/biology11020166
Kojima KK. Diversity and Evolution of DNA Transposons Targeting Multicopy Small RNA Genes from Actinopterygian Fish. Biology. 2022; 11(2):166. https://doi.org/10.3390/biology11020166
Chicago/Turabian StyleKojima, Kenji K. 2022. "Diversity and Evolution of DNA Transposons Targeting Multicopy Small RNA Genes from Actinopterygian Fish" Biology 11, no. 2: 166. https://doi.org/10.3390/biology11020166
APA StyleKojima, K. K. (2022). Diversity and Evolution of DNA Transposons Targeting Multicopy Small RNA Genes from Actinopterygian Fish. Biology, 11(2), 166. https://doi.org/10.3390/biology11020166