
Transcriptomic Properties of HER2+ Ductal Carcinoma In Situ of the Breast Associate with Absence of Immune Cells

 , , , , ,
, , , , , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Patients and Methods
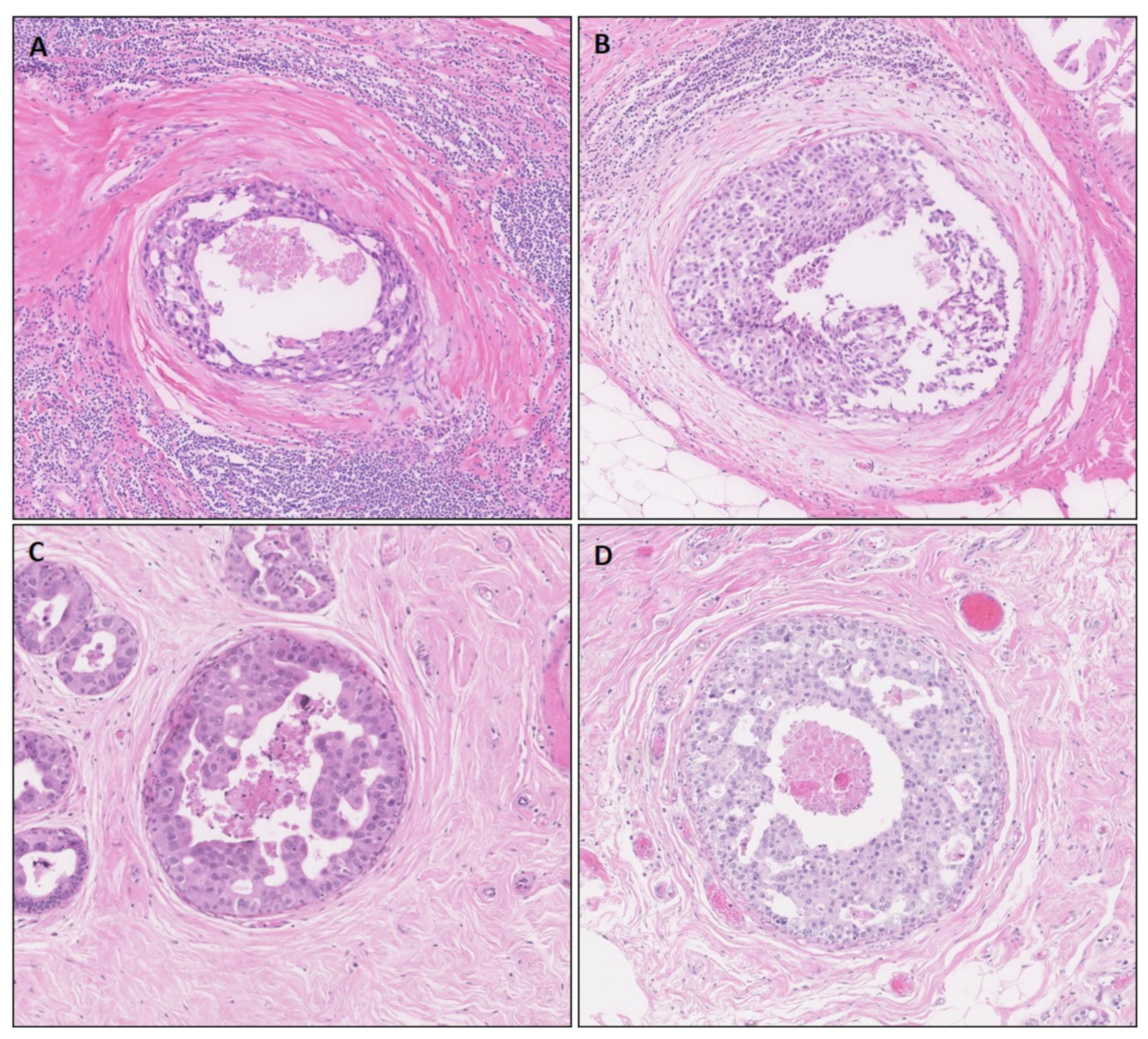
2.1. Study Cohort and Histopathological Assessment
2.2. Micro-Dissection and RNA Isolation
2.3. Targeted RNA Sequencing
2.4. Data Processing and Analysis
2.5. Immunohistochemistry
2.6. Statistical Analysis
3. Results
3.1. Clinicopathological Patient Characteristics
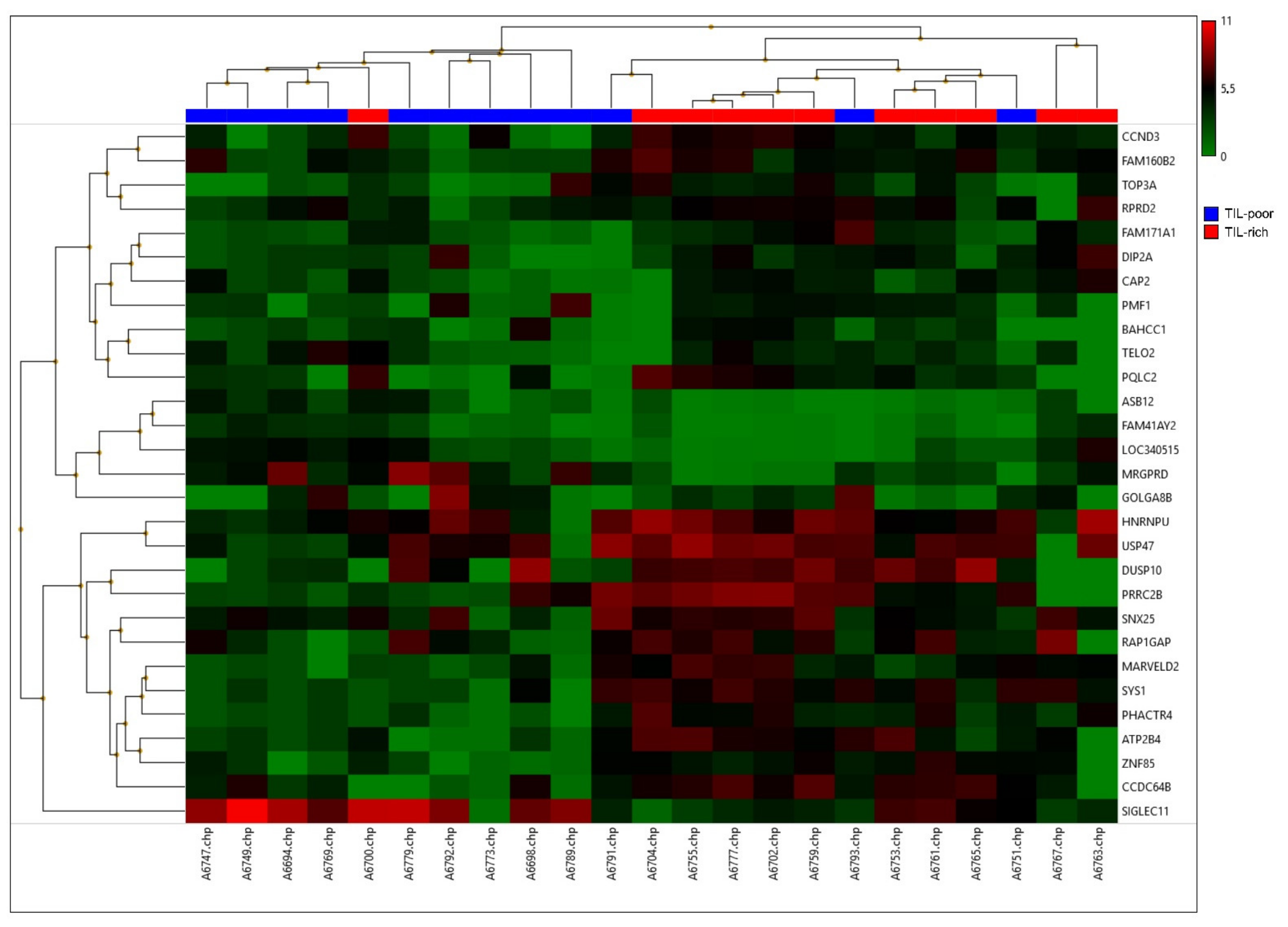
3.2. Differentially Expressed Genes on RNA Level
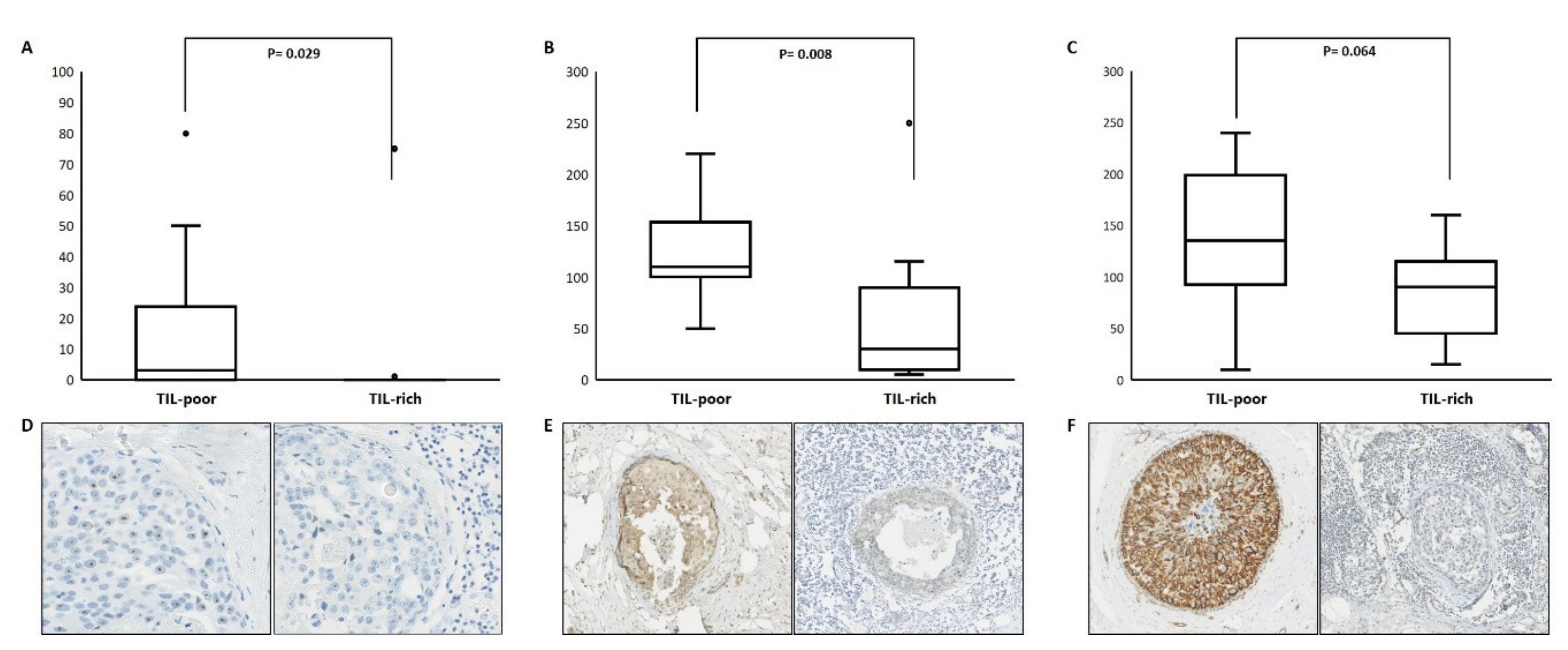
3.3. Differentially Expressed Genes on Protein Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Barnes, N.L.P.; Ooi, J.L.; Yarnold, J.R.; Bundred, N.J. Ductal carcinoma in situ of the breast. BMJ 2012, 344, e797. [Google Scholar] [CrossRef] [PubMed]
- Virnig, B.A.; Wang, S.-Y.; Shamilyan, T.; Kane, R.L.; Tuttle, T. Ductal Carcinoma In Situ: Risk Factors and Impact of Screening. J. Natl. Cancer Inst. Monogr. 2010, 2010, 113–116. [Google Scholar] [CrossRef]
- Cedolini, C.; Bertozzi, S.; Londero, A.P.; Seriau, L.; Andretta, M.; Agakiza, D.; Fongione, S.; Uzzau, A.; Risaliti, A. Impact of the presence and quantity of ductal carcinoma in situ component on the outcome of invasive breast cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 13304–13313. [Google Scholar]
- Hanna, W.M.; Parra-Herran, C.; Lu, F.-I.; Slodkowska, E.; Rakovitch, E.; Nofech-Mozes, S. Ductal carcinoma in situ of the breast: An update for the pathologist in the era of individualized risk assessment and tailored therapies. Mod. Pathol. 2019, 32, 896–915. [Google Scholar] [CrossRef] [PubMed]
- Meijnen, P.; Peterse, J.L.; Antonini, N.; Rutgers, E.J.T.; van de Vijver, M. Immunohistochemical categorisation of ductal carcinoma in situ of the breast. Br. J. Cancer 2007, 98, 137–142. [Google Scholar] [CrossRef]
- Van Seijen, M.; Lips, E.H.; Thompson, A.M.; Nik-Zainal, S.; Futreal, A.; Hwang, E.S.; Verschuur, E.; Lane, J.; Jonkers, J.; Rea, D.W.; et al. Ductal carcinoma in situ: To treat or not to treat, that is the question. Br. J. Cancer 2019, 121, 285–292. [Google Scholar] [CrossRef]
- Liao, N.; Zhang, G.-C.; Liu, Y.-H.; Li, X.-R.; Yao, M.; Xu, F.-P.; Li, L.; Wu, Y.-L. HER2-positive status is an independent predictor for coexisting invasion of ductal carcinoma in situ of the breast presenting extensive DCIS component. Pathol. Res. Pr. 2011, 207, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Doebar, S.; De Monyé, C.; Stoop, H.; Rothbarth, J.; Willemsen, S.; Van Deurzen, C. Ductal carcinoma in situ diagnosed by breast needle biopsy: Predictors of invasion in the excision specimen. Breast 2016, 27, 15–21. [Google Scholar] [CrossRef]
- Van Bockstal, M.; Lambein, K.; Denys, H.; Braems, G.; Nuyts, A.; Broecke, R.V.D.; Cocquyt, V.; De Wever, O.; Libbrecht, L. Histopathological characterization of ductal carcinoma in situ (DCIS) of the breast according to HER2 amplification status and molecular subtype. Virchows Arch. 2014, 465, 275–289. [Google Scholar] [CrossRef]
- Roses, R.E.; Paulson, E.C.; Sharma, A.; Schueller, J.E.; Nisenbaum, H.; Weinstein, S.; Fox, K.R.; Zhang, P.J.; Czerniecki, B.J. HER-2/neu Overexpression as a Predictor for the Transition from In situ to Invasive Breast Cancer. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1386–1389. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, R.E.; DeStefano, L.M.; Bahng, J.; Yoon-Flannery, K.; Fisher, C.S.; Zhang, P.J.; Tchou, J.; Czerniecki, B.J.; De La Cruz, L.M. Evaluating the Risk of Upstaging HER2-Positive DCIS to Invasive Breast Cancer. Ann. Surg. Oncol. 2017, 24, 2999–3003. [Google Scholar] [CrossRef]
- Davis, J.; Nemesure, B.; Mehmood, S.; Nayi, V.; Burke, S.; Brzostek, S.R.; Singh, M. Her2 and Ki67 Biomarkers Predict Recurrence of Ductal Carcinoma in Situ. Appl. Immunohistochem. Mol. Morphol. 2016, 24, 20–25. [Google Scholar] [CrossRef]
- Harada, S.; Ms, R.M.; Roses, R.E.; Graves, H.; Niu, H.; Sharma, A.; Ba, J.E.S.; Nisenbaum, H.; Czerniecki, B.J.; Zhang, P.J. The significance of HER-2/neu receptor positivity and immunophenotype in ductal carcinoma in situ with early invasive disease. J. Surg. Oncol. 2011, 104, 458–465. [Google Scholar] [CrossRef]
- Czerniecki, B.J.; Roses, R.E.; Koski, G.K. Development of Vaccines for High-Risk Ductal Carcinoma In situ of the Breast. Cancer Res. 2007, 67, 6531–6534. [Google Scholar] [CrossRef]
- Sharma, A.; Koldovsky, U.; Xu, S.; Ms, R.M.; Roses, R.; Bs, E.F.; Weinstein, S.; Nisenbaum, H.; Levine, B.L.; Fox, K.; et al. HER-2 pulsed dendritic cell vaccine can eliminate HER-2 expression and impact ductal carcinoma in situ. Cancer 2012, 118, 4354–4362. [Google Scholar] [CrossRef] [PubMed]
- Fracol, M.; Xu, S.; Mick, R.; Fitzpatrick, E.; Nisenbaum, H.; Roses, R.; Fisher, C.; Tchou, J.; Fox, K.; Zhang, P.; et al. Response to HER-2 Pulsed DC1 Vaccines is Predicted by Both HER-2 and Estrogen Receptor Expression in DCIS. Ann. Surg. Oncol. 2013, 20, 3233–3239. [Google Scholar] [CrossRef]
- Lowenfeld, L.; Mick, R.; Datta, J.; Xu, S.; Fitzpatrick, E.; Fisher, C.S.; Fox, K.R.; DeMichele, A.; Zhang, P.J.; Weinstein, S.P.; et al. Dendritic Cell Vaccination Enhances Immune Responses and Induces Regression of HER2pos DCIS Independent of Route: Results of Randomized Selection Design Trial. Clin. Cancer Res. 2017, 23, 2961–2971. [Google Scholar] [CrossRef] [PubMed]
- Czerniecki, B.J.; Koski, G.K.; Koldovsky, U.; Xu, S.; Cohen, P.A.; Mick, R.; Nisenbaum, H.; Pasha, T.; Xu, M.; Fox, K.R.; et al. Targeting HER-2/neu in Early Breast Cancer Development Using Dendritic Cells with Staged Interleukin-12 Burst Secretion. Cancer Res. 2007, 67, 1842–1852. [Google Scholar] [CrossRef] [PubMed]
- Pruneri, G.; Lazzeroni, M.; Bagnardi, V.; Tiburzio, G.B.; Rotmensz, N.; DeCensi, A.; Gonzaga, A.G.; Vingiani, A.; Curigliano, G.; Zurrida, S.; et al. The prevalence and clinical relevance of tumor-infiltrating lymphocytes (TILs) in ductal carcinoma in situ of the breast. Ann. Oncol. 2017, 28, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; Eynden, G.V.D.; Baehner, F.L.; Penault-Llorca, F.; et al. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. 2015, 26, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Denkert, C.; Loibl, S.; Noske, A.; Roller, M.; Müller, B.M.; Komor, M.; Budczies, J.; Darb-Esfahani, S.; Kronenwett, R.; Hanusch, C.; et al. Tumor-Associated Lymphocytes as an Independent Predictor of Response to Neoadjuvant Chemotherapy in Breast Cancer. J. Clin. Oncol. 2010, 28, 105–113. [Google Scholar] [CrossRef]
- Loi, S.; Michiels, S.; Salgado, R.; Sirtaine, N.; Jose, V.; Fumagalli, D.; Kellokumpu-Lehtinen, P.-L.; Bono, P.; Kataja, V.; Desmedt, C.; et al. Tumor infiltrating lymphocytes are prognostic in triple negative breast cancer and predictive for trastuzumab benefit in early breast cancer: Results from the FinHER trial. Ann. Oncol. 2014, 25, 1544–1550. [Google Scholar] [CrossRef]
- Stanton, S.E.; Disis, M.L. Clinical significance of tumor-infiltrating lymphocytes in breast cancer. J. Immunother. Cancer 2016, 4, 59. [Google Scholar] [CrossRef] [PubMed]
- Denkert, C.; Von Minckwitz, G.; Darb-Esfahani, S.; Lederer, B.; Heppner, B.I.; Weber, K.E.; Budczies, J.; Huober, J.; Klauschen, F.; Furlanetto, J.; et al. Tumour-infiltrating lymphocytes and prognosis in different subtypes of breast cancer: A pooled analysis of 3771 patients treated with neoadjuvant therapy. Lancet Oncol. 2018, 19, 40–50. [Google Scholar] [CrossRef]
- Pelekanou, V.; Barlow, W.E.; Nahleh, Z.A.; Wasserman, B.; Lo, Y.-C.; Von Wahlde, M.-K.; Hayes, D.F.; Hortobagyi, G.N.; Gralow, J.R.; Tripathy, D.; et al. Tumor-Infiltrating Lymphocytes and PD-L1 Expression in Pre- and Posttreatment Breast Cancers in the SWOG S0800 Phase II Neoadjuvant Chemotherapy Trial. Mol. Cancer Ther. 2018, 17, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Agahozo, M.C.; Hammerl, D.; Debets, R.; Kok, M.; Van Deurzen, C.H.M. Tumor-infiltrating lymphocytes and ductal carcinoma in situ of the breast: Friends or foes? Mod. Pathol. 2018, 31, 1012–1025. [Google Scholar] [CrossRef]
- Toss, M.S.; Miligy, I.; Al-Kawaz, A.; Alsleem, M.; Khout, H.; Rida, P.C.; Aneja, R.; Green, A.; Ellis, I.; Rakha, E.A. Prognostic significance of tumor-infiltrating lymphocytes in ductal carcinoma in situ of the breast. Mod. Pathol. 2018, 31, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-Y.; Yeong, J.; Thike, A.A.; Bay, B.H.; Tan, P.H. Prognostic role of immune infiltrates in breast ductal carcinoma in situ. Breast Cancer Res. Treat. 2019, 177, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Semeraro, M.; Adam, J.; Stoll, G.; Louvet, E.; Chaba, K.; Poirier-Colame, V.; Sauvat, A.; Senovilla, L.; Vacchelli, E.; Bloy, N.; et al. The ratio of CD8+/FOXP3 T lymphocytes infiltrating breast tissues predicts the relapse of ductal carcinoma in situ. OncoImmunology 2016, 5, e1218106. [Google Scholar] [CrossRef]
- Kim, M.; Chung, Y.R.; Kim, H.J.; Woo, J.W.; Ahn, S.; Park, S.Y. Immune microenvironment in ductal carcinoma in situ: A comparison with invasive carcinoma of the breast. Breast Cancer Res. 2020, 22, 32. [Google Scholar] [CrossRef]
- Agahozo, M.; Van Bockstal, M.; Groenendijk, F.; Bosch, T.V.D.; Westenend, P.; Van Deurzen, C. Ductal carcinoma in situ of the breast: Immune cell composition according to subtype. Ann. Oncol. 2019, 30, iii5. [Google Scholar] [CrossRef]
- Livasy, C.A.; Perou, C.; Karaca, G.; Cowan, D.W.; Maia, D.; Jackson, S.; Tse, C.-K.; Nyante, S.; Millikan, R.C. Identification of a basal-like subtype of breast ductal carcinoma in situ. Hum. Pathol. 2007, 38, 197–204. [Google Scholar] [CrossRef]
- E Clark, S.; Warwick, J.; Carpenter, R.; Bowen, R.L.; Duffy, S.W.; Jones, J.L. Molecular subtyping of DCIS: Heterogeneity of breast cancer reflected in pre-invasive disease. Br. J. Cancer 2010, 104, 120–127. [Google Scholar] [CrossRef]
- Morita, M.; Yamaguchi, R.; Tanaka, M.; Tse, G.; Yamaguchi, M.; Kanomata, N.; Naito, Y.; Akiba, J.; Hattori, S.; Minami, S.; et al. CD 8 + tumor-infiltrating lymphocytes contribute to spontaneous “healing” in HER 2-positive ductal carcinoma in situ. Cancer Med. 2016, 5, 1607–1618. [Google Scholar] [CrossRef]
- Parise, C.A.; Caggiano, V. Breast Cancer Survival Defined by the ER/PR/HER2 Subtypes and a Surrogate Classification according to Tumor Grade and Immunohistochemical Biomarkers. J. Cancer Epidemiol. 2014, 2014, 469251. [Google Scholar] [CrossRef] [PubMed]
- Doebar, S.C.; Broek, E.C.V.D.; Koppert, L.B.; Jager, A.; Baaijens, M.H.A.; Obdeijn, I.-M.A.M.; Van Deurzen, C.H.M. Extent of ductal carcinoma in situ according to breast cancer subtypes: A population-based cohort study. Breast Cancer Res. Treat. 2016, 158, 179–187. [Google Scholar] [CrossRef]
- Onitilo, A.A.; Engel, J.M.; Greenlee, R.T.; Mukesh, B.N. Breast Cancer Subtypes Based on ER/PR and Her2 Expression: Comparison of Clinicopathologic Features and Survival. Clin. Med. Res. 2009, 7, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Esteva, F.J.; Hubbard-Lucey, V.M.; Tang, J.; Pusztai, L. Immunotherapy and targeted therapy combinations in metastatic breast cancer. Lancet Oncol. 2019, 20, e175–e186. [Google Scholar] [CrossRef]
- García-Aranda, M.; Redondo, M. Immunotherapy: A Challenge of Breast Cancer Treatment. Cancers 2019, 11, 1822. [Google Scholar] [CrossRef] [PubMed]
- Hammerl, D.; Smid, M.; Timmermans, A.; Sleijfer, S.; Martens, J.W.; Debets, R. Breast cancer genomics and immu-no-oncological markers to guide immune therapies. Semin. Cancer Biol. 2018, 52, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Borstkanker Algemeen Richtlijn Richtlijnendatabase. Available online: https://richtlijnendatabase.nl/richtlijn/borstkanker/algemeen.html (accessed on 11 August 2021).
- Wolff, A.C.; Hammond, M.E.H.; Allison, K.H.; Harvey, B.E.; Mangu, P.B.; Bartlett, J.M.S.; Bilous, M.; Ellis, I.O.; Fitzgibbons, P.; Hanna, W.; et al. Human Epidermal Growth Factor Receptor 2 Testing in Breast Cancer: American Society of Clinical Oncology/College of American Pathologists Clinical Practice Guideline Focused Update. J. Clin. Oncol. 2018, 36, 2105–2122. [Google Scholar] [CrossRef]
- FEDERA. Human Tissue and Medical Research: Code of Conduct for Responsible Use (2011). Rotterdam. 2011. Available online: https://www.federa.org/sites/default/files/digital_version_first_part_code_of_conduct_in_uk_2011_12092012.pdf (accessed on 12 September 2018).
- Sieuwerts, A.M.; Schrijver, W.A.M.E.; Dalm, S.U.; De Weerd, V.; Moelans, C.B.; Ter Hoeve, N.; Van Diest, P.J.; Martens, J.W.M.; Van Deurzen, C.H.M. Progressive APOBEC3B mRNA expression in distant breast cancer metastases. PLoS ONE 2017, 12, e0171343. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.; Li, C.; Rabinovic, A. Adjusting batch effects in microarray expression data using empirical Bayes methods. Biostatistics 2006, 8, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Massink, M.P.G.; Kooi, I.E.; Martens, J.W.M.; Waisfisz, Q.; Meijers-Heijboer, H. Genomic profiling of CHEK2*1100delC-mutated breast carcinomas. BMC Cancer 2015, 15, 877. [Google Scholar] [CrossRef] [PubMed]
- Hammerl, D.; Massink, M.P.; Smid, M.; Van Deurzen, C.H.; Meijers-Heijboer, H.E.; Waisfisz, Q.; Debets, R.; Martens, J.W. Clonality, Antigen Recognition, and Suppression of CD8+ T Cells Differentially Affect Prognosis of Breast Cancer Subtypes. Clin. Cancer Res. 2019, 26, 505–517. [Google Scholar] [CrossRef]
- Chi, Y.; Huang, S.; Liu, M.; Guo, L.; Shen, X.; Wu, J. Cyclin D3 predicts disease-free survival in breast cancer. Cancer Cell Int. 2015, 15, 1–9. [Google Scholar] [CrossRef][Green Version]
- Cato, M.H.; Chintalapati, S.K.; Yau, I.W.; Omori, S.A.; Rickert, R.C. Cyclin D3 Is Selectively Required for Proliferative Expansion of Germinal Center B Cells. Mol. Cell. Biol. 2010, 31, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Hrstka, R.; Bouchalova, P.; Michalová, E.; Matoulkova, E.; Muller, P.; Coates, P.J.; Vojtěšek, B. AGR2 oncoprotein inhibits p38 MAPK and p53 activation through a DUSP10-mediated regulatory pathway. Mol. Oncol. 2015, 10, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Endo, Y.; Onodera, A.; Hirahara, K.; Asou, H.K.; Nakajima, T.; Kanno, T.; Ouchi, Y.; Uematsu, S.; Nishimasu, H.; et al. DUSP10 constrains innate IL-33-mediated cytokine production in ST2hi memory-type pathogenic Th2 cells. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Shah, S.; Brock, E.J.; Jackson, R.M.; Ji, K.; Boerner, J.L.; Sloane, B.F.; Mattingly, R.R. Downregulation of Rap1Gap: A Switch from DCIS to Invasive Breast Carcinoma via ERK/MAPK Activation. Neoplasia 2018, 20, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Sakr, R.A.; Weigelt, B.; Chandarlapaty, S.; Andrade, V.P.; Rocco, E.G.; Giri, D.; Ng, C.K.Y.; Cowell, C.F.; Rosen, N.; Reis-Filho, J.S.; et al. PI3K Pathway Activation in High-Grade Ductal Carcinoma In Situ—Implications for Progression to Invasive Breast Carcinoma. Clin. Cancer Res. 2014, 20, 2326–2337. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; Van Jaarsveld, M.T.M.; Hollestelle, A.; Der Smissen, W.J.C.P.-V.; Heine, A.A.J.; Boersma, A.W.M.; Liu, J.; Helmijr, J.; Ozturk, B.; Smid, M.; et al. miRNA expression profiling of 51 human breast cancer cell lines reveals subtype and driver mutation-specific miRNAs. Breast Cancer Res. 2013, 15, R33. [Google Scholar] [CrossRef]
- Darb-Esfahani, S.; Denkert, C.; Stenzinger, A.; Salat, C.; Sinn, B.; Schem, C.; Endris, V.; Klare, P.; Schmitt, W.; Blohmer, J.-U.; et al. Role ofTP53mutations in triple negative and HER2-positive breast cancer treated with neoadjuvant anthracycline/taxane-based chemotherapy. Oncotarget 2016, 7, 67686–67698. [Google Scholar] [CrossRef] [PubMed]
- Tőkés, T.; Tőkés, A.M.; Szentmártoni, G.; Kiszner, G.; Mühl, D.; Molnár, B.Á.; Kulka, J.; Krenács, T.; Dank, M. Prognostic and Clinicopathological Correlations of Cell Cycle Marker Expressions before and after the Primary Systemic Therapy of Breast Cancer. Pathol. Oncol. Res. 2020, 26, 1499–1510. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Park, I.A.; Heo, S.-H.; Kim, Y.-A.; Gong, G.; Lee, H.J. Association between p53 Expression and Amount of Tumor-Infiltrating Lymphocytes in Triple-Negative Breast Cancer. J. Pathol. Transl. Med. 2019, 53, 180–187. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Z.; Han, L.; Gong, S.; Wang, Y.; He, Z.; Feng, Y.; Yang, Z. Prognostic significance of cyclin D3 expression in malignancy patients: A meta-analysis. Cancer Cell Int. 2019, 19, 158. [Google Scholar] [CrossRef]
- Smid, M.; Gonzalez, F.G.R.; Sieuwerts, A.M.; Salgado, R.; Der Smissen, W.J.C.P.-V.; Van Der Vlugt-Daane, M.; Van Galen, A.; Nik-Zainal, S.; Staaf, J.; Brinkman, A.; et al. Breast cancer genome and transcriptome integration implicates specific mutational signatures with immune cell infiltration. Nat. Commun. 2016, 7, 12910. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Li, R.; Zhang, Q.; Wang, Y.; Jiang, Y.; Meng, Q.; Xi, Q.; Wu, G. Prognostic role of cyclin D2/D3 in multiple human malignant neoplasms: A systematic review and meta-analysis. Cancer Med. 2019, 8, 2717–2729. [Google Scholar] [CrossRef]
- Chen, B.B.; Glasser, J.R.; A Coon, T.; Mallampalli, R.K. F-box protein FBXL2 exerts human lung tumor suppressor-like activity by ubiquitin-mediated degradation of cyclin D3 resulting in cell cycle arrest. Oncogene 2011, 31, 2566–2579. [Google Scholar] [CrossRef][Green Version]
- Vargas, T.R.; Boudoukha, S.; E Simon, A.; Souidi, M.; Cuvellier, S.; Pinna, G.; Polesskaya, A. Post-transcriptional regulation of cyclins D1, D3 and G1 and proliferation of human cancer cells depend on IMP-3 nuclear localization. Oncogene 2014, 33, 2866–2875. [Google Scholar] [CrossRef]
- Goel, S.; DeCristo, M.J.; Watt, A.C.; BrinJones, H.; Sceneay, J.; Li, B.B.; Khan, N.; Ubellacker, J.M.; Xie, S.; Metzger-Filho, O.; et al. CDK4/6 inhibition triggers anti-tumour immunity. Nat. Cell Biol. 2017, 548, 471–475. [Google Scholar] [CrossRef]
- Deng, J.; Wang, E.S.; Jenkins, R.W.; Li, S.; Dries, R.; Yates, K.; Chhabra, S.; Huang, W.; Liu, H.; Aref, A.R.; et al. CDK4/6 Inhibition Augments Antitumor Immunity by Enhancing T-cell Activation. Cancer Discov. 2018, 8, 216–233. [Google Scholar] [CrossRef]
- Schaer, D.A.; Beckmann, R.P.; Dempsey, J.A.; Huber, L.; Forest, A.; Amaladas, N.; Li, Y.; Wang, Y.C.; Rasmussen, E.R.; Chin, D.; et al. The CDK4/6 Inhibitor Abemaciclib Induces a T Cell Inflamed Tumor Microenvironment and Enhances the Efficacy of PD-L1 Checkpoint Blockade. Cell Rep. 2018, 22, 2978–2994. [Google Scholar] [CrossRef]
- Manzano, R.G.; Martinez-Navarro, E.M.; Forteza, J.; Brugarolas, A. Microarray phosphatome profiling of breast cancer patients unveils a complex phosphatase regulatory role of the MAPK and PI3K pathways in estrogen receptor-negative breast cancers. Int. J. Oncol. 2014, 45, 2250–2266. [Google Scholar] [CrossRef]
- Lucci, M.A.; Orlandi, R.; Triulzi, T.; Tagliabue, E.; Balsari, A.; Villa-Moruzzi, E. Expression profile of tyrosine phosphatases in HER2 breast cancer cells and tumors. Cell. Oncol. 2010, 32, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Martínez, M.; Stamatakis, K.; Fresno, M. The Dual-Specificity Phosphatase 10 (DUSP10): Its Role in Cancer, Inflammation, and Immunity. Int. J. Mol. Sci. 2019, 20, 1626. [Google Scholar] [CrossRef] [PubMed]
- Manley, G.C.A.; Stokes, C.A.; Marsh, E.K.; Sabroe, I.; Parker, L.C. DUSP10 Negatively Regulates the Inflammatory Response to Rhinovirus through Interleukin-1β Signaling. J. Virol. 2019, 93, e01659-18. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, P.; Gao, K.; Tang, Y.; Jin, X.; Zhang, Y.; Yi, Q.; Wang, C.; Yu, L. PLK1 and β-TrCP-Dependent Ubiquitination and Degradation of Rap1GAP Controls Cell Proliferation. PLoS ONE 2014, 9, e110296. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, T.; Asaoka, M.; Katsuta, E.; Photiadis, S.J.; Narayanan, S.; Yan, L.; Takabe, K. High expression of polo-like kinase 1 is associated with TP53 inactivation, DNA repair deficiency, and worse prognosis in ER positive Her2 negative breast cancer. Am. J. Transl. Res. 2019, 11, 6507–6521. [Google Scholar] [PubMed]
- Li, M.; Liu, Z.; Wang, X. Exploration of the Combination of PLK1 Inhibition with Immunotherapy in Cancer Treatment. J. Oncol. 2018, 2018, 3979527. [Google Scholar] [CrossRef] [PubMed]
- King, S.I.; Purdie, C.A.; Bray, S.E.; Quinlan, P.R.; Jordan, L.B.; Thompson, A.M.; Meek, D.W. Immunohistochemical detection of Polo-like kinase-1 (PLK1) in primary breast cancer is associated with TP53mutation and poor clinical outcome. Breast Cancer Res. 2012, 14, R40. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | n/Median | %/Range |
|---|---|---|
| Age | 56 | 37.0–73.0 |
| DCIS size (cm) | 3.8 | 0.9–9.0 |
| Grade | ||
| Low | 0 | 0 |
| Intermediate | 1 | 4.3 |
| High | 22 | 95.7 |
| Comedonecrosis | ||
| Absent | 3 | 13 |
| Present | 20 | 87 |
| Growth Pattern | ||
| Solid | 15 | 65.2 |
| Cribriform | 7 | 30.4 |
| Papillary | 1 | 4.3 |
| p53 | ||
| Wild type | 7 | 30.4 |
| Aberrant | 11 | 47.8 |
| Null | 5 | 21.7 |
| Ki67 (%) | 14.2 | 3.0–25.0 |
| Gene | p-Value | Fold Change (High–Low) | Role in (Breast) Cancer | Reference | General Function (Uniprot.org, accessed on 11 July 2021) |
|---|---|---|---|---|---|
| HNRNPU | 0.02 | 4.07 | Upregulated in pancreatic ductal adenocarcinoma, mediates invasion and migration in pancreatic ductal adenocarcinoma cell lines | Shen et al., Med Sci Monit 2018, Sutaria et al., Noncoding RNA 2017 | DNA/RNA-binding protein, cell cycle/transcription |
| MRGPRD | 0.01 | −5.09 | Unkown | n.a. | Transcription |
| RPRD2 | 0.04 | 4.05 | Mutated in Burkitt lymphoma | Kaymaz et al., Mol Cancer Res. 2017 | Regulatory protein, transcription |
| TOP3A | 0.02 | 4.89 | Unknown | n.a. | DNA/RNA-binding protein, cell cycle/transcription |
| ZNF85 | 0.02 | 4.12 | Unknown, overexperssed in SCLC cell lines | Loiselle et al. Heliyon 2016 | DNA/RNA binding protein, transcription |
| BAHCC1 | 0.01 | 4.47 | Predicts survival in melanoma, upregulated in hepatocellular carcinoma | Gao et al., Biomed Res Int 2020, Nalesnik et al., Am J Pathol 2012 | Chromatin-binding protein, cell cycle |
| CCND3 | 0.02 | 4.29 | Amplified in basal-like breast cancer, correlates with reduced overall suvival in breast cancer, discriminates inflammatory breast cancer from non-inflammatory breast cancer | Smid et al., Nat Comm 2016, Ding et al., Cancer Medicine 2019, Keyomarsi et al., N Engl J Med 2002, Lerebours et al., BMC 2008 | Regulatory protein, cell cycle |
| PHACTR4 | 0.02 | 5.61 | Suggested tumorsupressor in various cancers including breast cancer, overexpression inhibits cell proliferation and invasion in hepatocellular carcinoma by inhibiting IL6/Stat3 pathway | Solimini et al., PNAS 2012, Cao et al., Eur Rev Med Pharmacol Sci. 2016 | Regulatory protein, cell cycle |
| PMF1 | 0.04 | 5.65 | Regulates the expression of SSAT in breast cancer cell lines, methilation associated with bladder cancer progression | Husbeck et al., Biochem Biophys Res Commun 2003, Aleman et al., Clin Cancer Res. 2008 | Involved in cell cycle |
| TELO2 | 0.05 | 4.52 | Associated with oncogenic profile in breast cancer cell line | Morais-Rodrigues et al., Gene 2020 | Cell cycle |
| USP47 | 0.03 | 4.33 | Promotes EMT (mortality and disasociation) in breast cancer cells | Silvestrini et al., J Proteomics 2020 | Ubiquitin-specific protease, negative regulator of cell cycle |
| DUSP10 | 0.01 | 6.62 | Mediates immune response, increased DUSP10 downregulates inflammation and overexpressed in HER2+ breast cancer, high expression associated with reduced relapse-free survival in ER+ wt P53 breast cancer | Jiménez-Martínez et al., Int J Mol Sci. 2019, Hrstka et al., Mol Onc 2015 | Enzyme, proliferation and differentiation |
| PRRC2B | 0.02 | 11.98 | Unknown, somatic variant found in T-cell lymphoma | Donner et al., Fam Cancer 2019 | RNA binding, cell differentiation |
| RAP1GAP | 0.03 | 6.18 | Tumorsupressive in several cancers, inhibits progression in endometrial cancer, increased in ductal carcinoma in situ compared to invasive breast cancer, reduced expression enhances invasion | Tamate et al., Biochem Biophys Res Commun. 2017, Shah et al., Neoplasia 2018 | Regulatory protein, differentiation and proliferation |
| ASB12 | 0.05 | −4.28 | Unknown | n.a. | E3 ubiquitin protein, translation |
| DIP2A | 0.05 | 4.68 | Promotes FSTL1 immune resistance and correlates with poor prognosis in non-small cell lung cancer patients | Kudo-Saito et al., Cell Rep 2018 | Regulatory protein, developmental protein |
| FAM171A1 | 0.00 | 5.16 | Increased expression in invasive vs. in situ breast carcinoma, correlates with loss of ER and formation of mammospheres (in cell lines) (suggested to increase metastatic potential) in triple negative breast cancer, suggested prognostic marker in triple negative breast cancer (cell lines), ref. | Rsila et al., The American Joural of Pathology 2019, Sanawar et al., Ongegenesis 2019, Bao et al., Cell Death Dis. 2019 | Cell shape/mortality |
| LOC340515 | 0.04 | −4.02 | Unkown | n.a. | |
| MARVELD2 | 0.02 | 5.49 | Generally described in pancreas and liver carcinoma. Overexpression (tricellulin) associated with unfavorable pronnosis in primary liver carcinomas, decreased expression correlates with poor prognosis in pancreatic adenocarcionma | Somoracz et al., Pathol Oncol Res. 2014 | Cell–cell junction |
| SIGLEC11 | 0.03 | −8.59 | Unknown | n.a. | Regulatory protein |
| ATP2B4 | 0.01 | 8.89 | Metastasis surpressor of BRAF mutated melanoma cells, overexpression plays a role in chronic lymphocytic leukemia pathogenesis, lower ATP2B4 mRNA expression in invasive breast cancer tissue samples compared to normal breast tissue | Hegudus et al., Int J Cancer 2017, Johnston et al., Mol Cell Proteomics 2018, Varga et al., BMC Cancer 2028 | Enzyme/catalyzes calcium transport |
| CCDC64B | 0.01 | 4.15 | Unknown | n.a. | Rab GTPase binding, transport |
| GOLGA8B | 0.04 | −4.27 | Associated with shorter overall survival in patients with renal cell carcinoma, associated with tumor progression and prognosis in prostate cancer | Wang et al., Journal of Cellular Biochemistry 2018, Cheng et al., J Cell Mol Med. 2020 | Protein transport |
| PQLC2 | 0.00 | 4.13 | Overexpression promotes cell growth and tumor formation of gastric cancer in nude mice. Suppression/inhibition causes cell death of cancer cells and suppressed growth | Jeung et al., Cancer Sci 2019 | Protein transport |
| SNX25 | 0.01 | 4.44 | Unknown | n.a. | Protein transport |
| SYS1 | 0.04 | 8.32 | Overexpressed in cervical cancer | Wu et al., Mol Med Rep 2018 | Protein transport |
| CAP2 | 0.02 | 4.70 | Associated with PR expression and decreased overall survival in breast cancer, suggested prognostic marker in gastric cancer | Xu et al., Oncol Rep 2016, Li et al., Pathol Oncol Res. 2020 | Regulatory protein, unknown |
| FAM160B2 | 0.03 | 4.26 | Enhances tumorigenesis in hepatocellular carcinoma (RAI16) | Wang et al., Carcinogenesis 2012 | Unknown |
| FAM41AY2 | 0.02 | −4.38 | Unknown | n.a. | Unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agahozo, M.C.; Smid, M.; van Marion, R.; Hammerl, D.; van den Bosch, T.P.P.; Timmermans, M.A.M.; Heijerman, C.J.; Westenend, P.J.; Debets, R.; Martens, J.W.M.; et al. Transcriptomic Properties of HER2+ Ductal Carcinoma In Situ of the Breast Associate with Absence of Immune Cells. Biology 2021, 10, 768. https://doi.org/10.3390/biology10080768
Agahozo MC, Smid M, van Marion R, Hammerl D, van den Bosch TPP, Timmermans MAM, Heijerman CJ, Westenend PJ, Debets R, Martens JWM, et al. Transcriptomic Properties of HER2+ Ductal Carcinoma In Situ of the Breast Associate with Absence of Immune Cells. Biology. 2021; 10(8):768. https://doi.org/10.3390/biology10080768
Chicago/Turabian StyleAgahozo, Marie Colombe, Marcel Smid, Ronald van Marion, Dora Hammerl, Thierry P. P. van den Bosch, Mieke A. M. Timmermans, Chayenne J. Heijerman, Pieter J. Westenend, Reno Debets, John W. M. Martens, and et al. 2021. "Transcriptomic Properties of HER2+ Ductal Carcinoma In Situ of the Breast Associate with Absence of Immune Cells" Biology 10, no. 8: 768. https://doi.org/10.3390/biology10080768
APA StyleAgahozo, M. C., Smid, M., van Marion, R., Hammerl, D., van den Bosch, T. P. P., Timmermans, M. A. M., Heijerman, C. J., Westenend, P. J., Debets, R., Martens, J. W. M., & van Deurzen, C. H. M. (2021). Transcriptomic Properties of HER2+ Ductal Carcinoma In Situ of the Breast Associate with Absence of Immune Cells. Biology, 10(8), 768. https://doi.org/10.3390/biology10080768