
Do We Need to Use Bats as Bioindicators?

 ,
,  ,
, 
 and
and 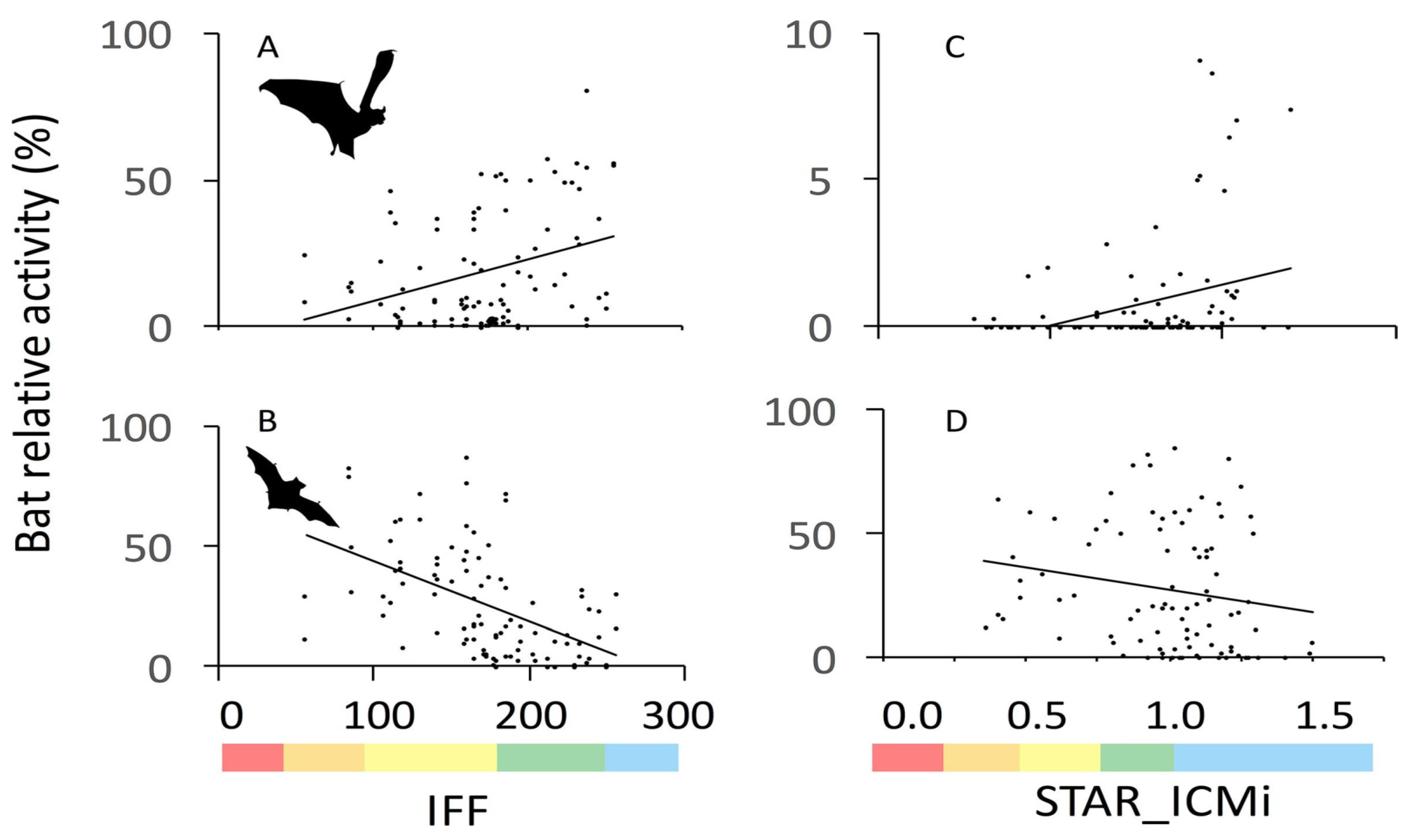{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Testing Bats as Bioindicators: Where Are We Now?
2.1. River Quality
2.2. Farming Practices
2.3. Forest Structure and Management
2.4. Urbanisation
2.5. Bioaccumulation
2.6. Climate Change
2.7. Surrogate Taxa
3. Potential Limitations to the Use of Bats as Bioindicators
3.1. Taxonomical Issues
3.2. Sampling Limitations
3.3. Disentangling Cause-Effect Relationships
3.4. Responses May Be Influenced by Local Adaptations
3.5. Delayed Responses to Environmental Stressors?
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allaby, M.A. Dictionary of Zoology, 5th ed.; Oxford University Press: London, UK, 2020; p. 736. [Google Scholar] [CrossRef]
- McGeoch, M. The selection, testing and application of terrestrial insects as bioindicators. Biol. Rev. 1998, 73, 181–201. [Google Scholar] [CrossRef]
- Macher, J.N.; Salis, R.K.; Blakemore, K.S.; Tollrian, R.; Matthaei, C.D.; Leese, F. Multiple-stressor effects on stream invertebrates: DNA barcoding reveals contrasting responses of cryptic mayfly species. Ecol. Indic. 2016, 61, 159–169. [Google Scholar] [CrossRef]
- Rice, J.C.; Rochet, M.J. A framework for selecting a suite of indicators for fisheries management. ICES J. Mar. Sci. 2005, 62, 516–527. [Google Scholar] [CrossRef]
- Mammal Diversity Database. Mammal Diversity Database (Version 1.4), [Data download May/2021]. Zenodo 2021. [Google Scholar] [CrossRef]
- Russo, D.; Bosso, L.; Ancillotto, L. Novel perspectives on bat insectivory highlight the value of this ecosystem service in farmland: Research frontiers and management implications. Agric. Ecosyst. Environ. 2018, 266, 31–38. [Google Scholar] [CrossRef]
- Kunz, T.H.; Braun de Torrez, E.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef]
- Frick, W.F.; Kingston, T.; Flanders, J. A review of the major threats and challenges to global bat conservation. Ann. N. Y. Acad. Sci. 2019, 1469, 5–25. [Google Scholar] [CrossRef]
- Jones, G.; Jacobs, D.S.; Kunz, T.H.; Willig, M.R.; Racey, P.A. Carpe noctem: The importance of bats as bioindicators. Endanger. Species Res. 2009, 8, 93–115. [Google Scholar] [CrossRef]
- Flaquer, C.; Puig-Montserrat, X. Proceedings of the International Symposium on the Importance of Bats as Bioindicators; Museum of Natural Sciences Edicions: Granollers, Spain, 2012; p. 95. [Google Scholar]
- Russo, D.; Jones, G. Bats as bioindicators. Mamm. Biol. 2015, 80, 157–246. [Google Scholar] [CrossRef]
- Department for Environment, Food and Rural Affairs. UK Biodiversity Indicators 2020; Department for Environment, Food and Rural Affairs: London, UK, 2020; p. 60.
- Van der Meij, T.; Van Strien, A.J.; Haysom, K.A.; Dekker, J.; Russ, J.; Biala, K.; Bihari, Z.; Jansen, E.; Langton, S.; Kurali, A.; et al. Return of the bats? A prototype indicator of trends in European bat populations in underground hibernacula. Mamm. Biol. 2015, 80, 170–177. [Google Scholar] [CrossRef]
- Mas, M.; Flaquer, C.; Rebelo, H.; López-Baucells, A. Bats and wetlands: Synthesising gaps in current knowledge and future opportunities for conservation. Mamm. Rev. 2021. [Google Scholar] [CrossRef]
- Langton, S.D.; Briggs, P.A.; Haysom, K.A. Daubenton’s bat distribution along rivers–developing and testing a predictive model. Aquat. Conserv. 2010, 20, S45–S54. [Google Scholar] [CrossRef]
- López-Baucells, A.; Casanova, L.; Puig-Montserrat, X.; Espinal, A.; Páramo, F.; Flaquer, C. Evaluating the use of Myotis daubentonii as an ecological indicator in Mediterranean riparian habitats. Ecol. Ind. 2017, 74, 19–27. [Google Scholar] [CrossRef]
- Li, H.; Kalcounis-Rueppell, M. Separating the effects of water quality and urbanization on temperate insectivorous bats at the landscape scale. Ecol. Evol. 2018, 8, 667–678. [Google Scholar] [CrossRef]
- De Conno, C.; Nardone, V.; Ancillotto, L.; De Bonis, S.; Guida, M.; Jorge, I.; Scarpa, U.; Russo, D. Testing the performance of bats as indicators of riverine ecosystem quality. Ecol. Ind. 2018, 95, 741–750. [Google Scholar] [CrossRef]
- Park, K.J. Mitigating the impacts of agriculture on biodiversity: Bats and their potential role as bioindicators. Mamm. Biol. 2015, 80, 191–204. [Google Scholar] [CrossRef]
- Olimpi, E.M.; Philpott, S.M. Agroecological farming practices promote bats. Agric. Ecosyst. Environ. 2018, 265, 282–291. [Google Scholar] [CrossRef]
- Wickramasinghe, L.P.; Harris, S.; Jones, G.; Vaughan, N. Bat activity and species richness on organic and conventional farms: Impact of agricultural intensification. J. Appl. Ecol. 2003, 40, 984–993. [Google Scholar] [CrossRef]
- Fuller, R.J.; Norton, L.R.; Feber, R.E.; Johnson, P.J.; Chamberlain, D.E.; Joys, A.C.; Mathews, F.; Stuart, R.C.; Townsend, M.C.; Manley, W.J.; et al. Benefits of organic farming to biodiversity vary among taxa. Biol. Lett. 2005, 1, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Davy, C.M.; Russo, D.; Fenton, M.B. Use of native woodlands and traditional olive groves by foraging bats on a Mediterranean island: Consequences for conservation. J. Zool. 2007, 273, 397–405. [Google Scholar] [CrossRef]
- Put, J.E.; Mitchell, G.W.; Fahrig, L. Higher bat and prey abundance at organic than conventional soybean fields. Biol. Cons. 2018, 226, 177–185. [Google Scholar] [CrossRef]
- Pocock, M.J.; Jennings, N. Testing biotic indicator taxa: The sensitivity of insectivorous mammals and their prey to the intensification of lowland agriculture. J. Appl. Ecol. 2008, 45, 151–160. [Google Scholar] [CrossRef]
- Long, B.L.; Kurta, A. Activity and diet of bats in conventional versus organic apple orchards in southern Michigan. Can. Field Nat. 2014, 128, 158–164. [Google Scholar] [CrossRef][Green Version]
- Froidevaux, J.S.; Louboutin, B.; Jones, G. Does organic farming enhance biodiversity in Mediterranean vineyards? A case study with bats and arachnids. Agric. Ecosyst. Environ. 2017, 249, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Denzinger, A.; Schnitzler, H.U. Bat guilds, a concept to classify the highly diverse foraging and echolocation behaviors of microchiropteran bats. Front. Physiol. 2013, 4, 164. [Google Scholar] [CrossRef]
- Russo, D.; Billington, G.; Bontadina, F.; Dekker, J.; Dietz, M.; Jones, G.; Meschede, A.; Rebelo, H.; Reiter, G. Identifying key research objectives to make European forests greener for bats. Front. Ecol. Evol. 2016, 4. [Google Scholar] [CrossRef]
- Lacki, M.; Hayes, J.; Kurta, A. Bats in Forests: Conservation and Management; Baltimore, M.D., Ed.; The John Hopkins University Press: Baltimore, MD, USA, 2007; p. 329. [Google Scholar]
- Cistrone, L.; Altea, T.; Matteucci, G.; Posillico, M.; De Cinti, B.; Russo, D. The effect of thinning on bat activity in Italian high forests: The LIFE + “ManFor C. BD” experience. Hystrix It. J. Mamm. 2015, 26, 125–131. [Google Scholar]
- Steel, Z.L.; Campos, B.; Frick, W.F.; Burnett, R.; Safford, H.D. The effects of wildfire severity and pyrodiversity on bat occupancy and diversity in fire-suppressed forests. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bosso, L.; Ancillotto, L.; Smeraldo, S.; D’Arco, S.; Migliozzi, A.; Conti, P.; Russo, D. Loss of potential bat habitat following a severe wildfire: A model-based rapid assessment. Int. J. Wildland Fire 2018, 27, 756–769. [Google Scholar] [CrossRef]
- López-Baucells, A.; Flaquer, C.; Mas, M.; Pons, P.; Puig-Montserrat, X. Recurring fires in Mediterranean habitats and their impact on bats. Biodivers. Conserv. 2021, 30, 385–402. [Google Scholar] [CrossRef]
- Santini, L.; González-Suárez, M.; Russo, D.; Gonzalez-Voyer, A.; von Hardenberg, A.; Ancillotto, L. One strategy does not fit all: Determinants of urban adaptation in mammals. Ecol. Lett. 2019, 22, 365–376. [Google Scholar] [CrossRef]
- Ancillotto, L.; Bosso, L.; Salinas-Ramos, V.B.; Russo, D. The importance of ponds for the conservation of bats in urban landscapes. Landsc. Urban Plan. 2019, 190, 103607. [Google Scholar] [CrossRef]
- Stone, E.L.; Harris, S.; Jones, G. Impacts of artificial lighting on bats: A review of challenges and solutions. Mammal. Biol. 2015, 80, 213–219. [Google Scholar] [CrossRef]
- Finch, D.; Schofield, H.; Mathews, F. Traffic noise playback reduces the activity and feeding behaviour of free-living bats. Environ. Pollut. 2020, 263, 114405. [Google Scholar] [CrossRef] [PubMed]
- Zukal, J.; Pikula, J.; Bandouchova, H. Bats as bioindicators of heavy metal pollution: History and prospect. Mammal. Biol. 2015, 80, 220–227. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Protection Products and their Residues (PPR); Hernandez-Jerez, A.; Adriaanse, P.; Aldrich, A.; Berny, P.; Coja, T.; Duquesne, S.; Gimsing, A.L.; Marina, M.; Millet, M.; et al. Scientific statement on the coverage of bats by the current pesticide risk assessment for birds and mammals. EFSA J. 2019, 17, e05758. [Google Scholar]
- Heiker, L.M.; Adams, R.A.; Ramos, C.V. Mercury bioaccumulation in two species of insectivorous bats from urban China: Influence of species, age, and land use type. Arch. Environ. Contam. Toxicol. 2018, 75, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Flache, L.; Ekschmitt, K.; Kierdorf, U.; Czarnecki, S.; Düring, R.A.; Encarnação, J.A. Reduction of metal exposure of Daubenton’s bats (Myotis daubentonii) following remediation of pond sediment as evidenced by metal concentrations in hair. Sci. Total Environ. 2016, 547, 182–189. [Google Scholar] [CrossRef]
- Altringham, J.D. Bats: From Evolution to Conservation; Oxford University Press: New York, NY, USA, 2011; p. 331. [Google Scholar]
- Smeraldo, S.; Bosso, L.; Salinas-Ramos, V.B.; Ancillotto, L.; Sánchez-Cordero, V.; Gazaryan, S.; Russo, D. Generalists yet different: Distributional responses to climate change may vary in opportunistic bat species sharing similar ecological traits. Mamm. Rev. 2021. [Google Scholar] [CrossRef]
- Newson, S.E.; Mendes, S.; Crick, H.Q.; Dulvy, N.K.; Houghton, J.D.; Hays, G.C.; Robinson, R.A. Indicators of the impact of climate change on migratory species. Endanger. Species Res. 2009, 7, 101–113. [Google Scholar] [CrossRef]
- Lewandowski, A.S.; Noss, R.F.; Parsons, D.R. The effectiveness of surrogate taxa for the representation of biodiversity. Conserv. Biol. 2010, 24, 1367–1377. [Google Scholar] [CrossRef]
- Sauberer, N.; Zulka, K.P.; Abensperg-Traun, M.; Berg, H.M.; Bieringer, G.; Milasowszky, N.; Grabherr, G. Surrogate taxa for biodiversity in agricultural landscapes of eastern Austria. Biol. Conserv. 2004, 117, 181–190. [Google Scholar] [CrossRef]
- Wunderle, J.M., Jr. The role of animal seed dispersal in accelerating native forest regeneration on degraded tropical lands. For. Ecol. Manag. 1997, 99, 223–235. [Google Scholar] [CrossRef]
- Lund, M.P.; Rahbek, C. Cross-taxon congruence in complementarity and conservation of temperate biodiversity. Animal Conserv. 2002, 5, 163–171. [Google Scholar] [CrossRef]
- Larrieu, L.; Gosselin, F.; Archaux, F.; Chevalier, R.; Corriol, G.; Dauffy-Richard, E.; Bouget, C. Cost-efficiency of cross-taxon surrogates in temperate forests. Ecol. Ind. 2018, 87, 56–65. [Google Scholar] [CrossRef]
- Van Weerd, M.; De Haes, H.A.U. Cross-taxon congruence in tree, bird and bat species distributions at a moderate spatial scale across four tropical forest types in the Philippines. Biod. Conserv. 2010, 19, 3393–3411. [Google Scholar] [CrossRef]
- Franke, S.; Brandl, R.; Heibl, C.; Mattivi, A.; Müller, J.; Pinkert, S.; Thorn, S. Predicting regional hotspots of phylogenetic diversity across multiple species groups. Diver. Distrib. 2020, 26, 1305–1314. [Google Scholar] [CrossRef]
- Mayer, F.; Helversen, O.V. Cryptic diversity in European bats. P. Roy Soc. B Biol. Sci. 2001, 268, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Demos, T.C.; Webala, P.W.; Kerbis Peterhans, J.C.; Goodman, S.M.; Bartonjo, M.; Patterson, B.D. Molecular phylogenetics of slit-faced bats (Chiroptera: Nycteridae) reveal deeply divergent African lineages. J. Zool. Syst. Evol. Res. 2019, 57, 1019–1038. [Google Scholar] [CrossRef] [PubMed]
- Barratt, E.M.; Deaville, R.; Burland, T.M.; Bruford, M.W.; Jones, G.; Racey, P.A.; Wayne, R.K. DNA answers the call of pipistrelle bat species. Nature 1997, 387, 138–139. [Google Scholar] [CrossRef]
- Ancillotto, L.; Bosso, L.; Mori, E.; Mazza, G.; Herkt, M.; Galimberti, A.; Ramazzotti, F.; Russo, D. An African bat in Europe, Plecotus gaisleri: Biogeographic and ecological insights from molecular taxonomy and Species Distribution Models. Ecol. Evol. 2020, 10, 5785–5800. [Google Scholar] [CrossRef] [PubMed]
- Davidson-Watts, I.; Walls, S.; Jones, G. Differential habitat selection by Pipistrellus pipistrellus and Pipistrellus pygmaeus identifies distinct conservation needs for cryptic species of echolocating bats. Biol. Conserv. 2006, 133, 118–127. [Google Scholar] [CrossRef]
- Flaquer, C.; Torre, I.; Arrizabalaga, A. Comparison of sampling methods for inventory of bat communities. J. Mammal. 2007, 88, 526–533. [Google Scholar] [CrossRef]
- Bogdanowicz, W.; Hulva, P.; Bolfíková, B.C.; Buś, M.M.; Rychlicka, E.; Sztencel-Jabłonka, A.; Cistrone, L.; Russo, D. Cryptic diversity of Italian bats and the role of the Apennine refugium in the phylogeography of the western Palaearctic. Zool. J. Linn. Soc. 2015, 174, 635–648. [Google Scholar] [CrossRef]
- Obrist, M. Bat Echolocation Research. A Handbook for Planning and Conducting Acoustic Studies, 2nd ed.; Fraser, E.E., Silvis, A., Brigham, R.M., Czenze, Z.J., Eds.; Bat Conservation International: Tucson, AZ, USA, 2020; p. 122. [Google Scholar]
- Griffin, D.R.; Webster, F.A.; Michael, C.R. The echolocation of flying insects by bats. Anim. Behav. 1960, 8, 141–154. [Google Scholar] [CrossRef]
- Russo, D.; Ancillotto, L.; Jones, G. Bats are still not birds in the digital era: Echolocation call variation and why it matters for bat species identification. Can. J. Zool. 2018, 96, 63–78. [Google Scholar] [CrossRef]
- Russo, D.; Voigt, C.C. The use of automated identification of bat echolocation calls in acoustic monitoring: A cautionary note for a sound analysis. Ecol. Indic. 2016, 66, 598–602. [Google Scholar] [CrossRef]
- Ellis, D. Taxonomic sufficiency in pollution assessment. Mar. Pollut. Bull. 1985, 16, 459. [Google Scholar] [CrossRef]
- Resh, V.H.; Rosenberg, D.M. Freshwater Biomonitoring and Benthic Macroinvertebrates (No. 504.4 FRE); Chapman & Hall: New York, NY, USA, 2003; p. 790. [Google Scholar]
- Wright, I.A.; Ryan, M.M. Impact of mining and industrial pollution on stream macroinvertebrates: Importance of taxonomic resolution, water geochemistry and EPT indices for impact detection. Hydrobiologia 2016, 772, 103–115. [Google Scholar] [CrossRef]
- Guillera-Arroita, G.; Lahoz-Monfort, J.J.; van Rooyen, A.R.; Weeks, A.R.; Tingley, R. Dealing with false-positive and false-negative errors about species occurrence at multiple levels. Methods Ecol. Evol. 2017, 8, 1081–1091. [Google Scholar] [CrossRef]
- Ancillotto, L.; Bosso, L.; Conti, P.; Russo, D. Resilient responses by bats to a severe wildfire: Conservation implications. Anim. Conserv. 2021. [Google Scholar] [CrossRef]
- Salinas-Ramos, V.B.; Ancillotto, L.; Cistrone, L.; Nastasi, C.; Bosso, L.; Smeraldo, S.; Cordero, V.S.; Russo, D. Artificial illumination influences niche segregation in bats. Environ. Pollut. 2021, 284, 117187. [Google Scholar] [CrossRef] [PubMed]
- Russo, D.; Cistrone, L.; Jones, G.; Mazzoleni, S. Roost selection by barbastelle bats (Barbastella barbastellus, Chiroptera: Vespertilionidae) in beech woodlands of central Italy: Consequences for conservation. Biol. Conserv. 2004, 117, 73–81. [Google Scholar] [CrossRef]
- Russo, D.; Di Febbraro, M.; Cistrone, L.; Jones, G.; Smeraldo, S.; Garonna, A.P.; Bosso, L. Protecting one, protecting both? Scale-dependent ecological differences in two species using dead trees, the rosalia longicorn beetle and the barbastelle bat. J. Zool. 2015, 297, 165–175. [Google Scholar] [CrossRef]
- Ancillotto, L.; Rydell, J.; Nardone, V.; Russo, D. Coastal cliffs on islands as foraging habitat for bats. Acta Chiropt. 2014, 16, 103–108. [Google Scholar] [CrossRef]
- Ancillotto, L.; Cistrone, L.; Mosconi, F.; Jones, G.; Boitani, L.; Russo, D. The importance of non-forest landscapes for the conservation of forest bats: Lessons from barbastelles (Barbastella barbastellus). Biodivers. Conserv. 2015, 24, 171–185. [Google Scholar] [CrossRef]
- Russo, D.; Cistrone, L.; Garonna, A.P.; Jones, G. Reconsidering the importance of harvested forests for the conservation of tree-dwelling bats. Biodivers. Conserv. 2010, 19, 2501–2515. [Google Scholar] [CrossRef]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mammal. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Kolkert, H.; Smith, R.; Rader, R.; Reid, N. Insectivorous bats provide significant economic value to the Australian cotton industry. Ecosyst. Serv. 2021, 49, 101280. [Google Scholar] [CrossRef]
- Arrizabalaga-Escudero, A.; Garin, I.; García-Mudarra, J.L.; Alberdi, A.; Aihartza, J.; Goiti, U. Trophic requirements beyond foraging habitats: The importance of prey source habitats in bat conservation. Biol. Conserv. 2015, 191, 512–519. [Google Scholar] [CrossRef]
- Russo, D.; Cistrone, L.; Jones, G. Spatial and temporal patterns of roost use by tree-dwelling barbastelle bats Barbastella barbastellus. Ecography 2005, 28, 769–776. [Google Scholar] [CrossRef]
- Russo, D.; Cistrone, L.; Jones, G. Emergence time in forest bats: The influence of canopy closure. Acta Oecol. 2007, 31, 119–126. [Google Scholar] [CrossRef]
- Russo, D.; Cistrone, L.; Budinski, I.; Console, G.; Della Corte, M.; Milighetti, C.; Di Salvo, I.; Nardone, V.; Brigham, R.M.; Ancillotto, L. Sociality influences thermoregulation and roost switching in a forest bat using ephemeral roosts. Ecol. Evol. 2017, 7, 5310–5321. [Google Scholar] [CrossRef] [PubMed]
- Mendes, E.S.; Fonseca, C.; Marques, S.F.; Maia, D.; Pereira, M.J.R. Bat richness and activity in heterogeneous landscapes: Guild-specific and scale-dependent? Landsc. Ecol. 2017, 32, 295–311. [Google Scholar] [CrossRef]
- O’Donnell, C.F. Influence of season, habitat, temperature, and invertebrate availability on nocturnal activity of the New Zealand long-tailed bat (Chalinolobus tuberculatus). N. Z. J. Zool. 2000, 27, 207–221. [Google Scholar] [CrossRef]
- Verboom, B.; Spoelstra, K. Effects of food abundance and wind on the use of tree lines by an insectivorous bat, Pipistrellus pipistrellus. Can. J. Zool. 1999, 77, 1393–1401. [Google Scholar] [CrossRef]
- Saldaña-Vázquez, R.A.; Munguía-Rosas, M.A. Lunar phobia in bats and its ecological correlates: A meta-analysis. Mamm. Biol. 2013, 78, 216–219. [Google Scholar] [CrossRef]
- Razgour, O.; Forester, B.; Taggart, J.B.; Bekaert, M.; Juste, J.; Ibáñez, C.; Puechmaille, S.J.; Novella-Fernandez, R.; Alberdi, A.; Manel, S. Considering adaptive genetic variation in climate change vulnerability assessment reduces species range loss projections. Proc. Natl. Acad. Sci. USA 2019, 116, 10418–10423. [Google Scholar] [CrossRef]
- Rodhouse, T.J.; Rodriguez, R.M.; Banner, K.M.; Ormsbee, P.C.; Barnett, J.; Irvine, K.M. Evidence of region-wide bat population decline from long-term monitoring and Bayesian occupancy models with empirically informed priors. Ecol. Evol. 2019, 9, 11078–11088. [Google Scholar] [CrossRef]
- Yamanaka, S.; Akasaka, T.; Yamaura, Y.; Kaneko, M.; Nakamura, F. Time-lagged responses of indicator taxa to temporal landscape changes in agricultural landscapes. Ecol. Indic. 2015, 48, 593–598. [Google Scholar] [CrossRef]
- Russo, D.; Ancillotto, L.; Hughes, A.C.; Galimberti, A.; Mori, E. Collection of voucher specimens for bat research: Conservation, ethical implications, reduction, and alternatives. Mamm. Rev. 2017, 47, 237–246. [Google Scholar] [CrossRef]
- Hill, A.P.; Prince, P.; Snaddon, J.L.; Doncaster, C.P.; Rogers, A. AudioMoth: A low-cost acoustic device for monitoring biodiversity and the environment. HardwareX 2019, 6, e00073. [Google Scholar] [CrossRef]
- Revilla-Martín, N.; Budinski, I.; Puig-Montserrat, X.; Flaquer, C.; López-Baucells, A. Monitoring cave-dwelling bats using remote passive acoustic detectors: A new approach for cave monitoring. Bioacoustics 2020, 1–16. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russo, D.; Salinas-Ramos, V.B.; Cistrone, L.; Smeraldo, S.; Bosso, L.; Ancillotto, L. Do We Need to Use Bats as Bioindicators? Biology 2021, 10, 693. https://doi.org/10.3390/biology10080693
Russo D, Salinas-Ramos VB, Cistrone L, Smeraldo S, Bosso L, Ancillotto L. Do We Need to Use Bats as Bioindicators? Biology. 2021; 10(8):693. https://doi.org/10.3390/biology10080693
Chicago/Turabian StyleRusso, Danilo, Valeria B. Salinas-Ramos, Luca Cistrone, Sonia Smeraldo, Luciano Bosso, and Leonardo Ancillotto. 2021. "Do We Need to Use Bats as Bioindicators?" Biology 10, no. 8: 693. https://doi.org/10.3390/biology10080693
APA StyleRusso, D., Salinas-Ramos, V. B., Cistrone, L., Smeraldo, S., Bosso, L., & Ancillotto, L. (2021). Do We Need to Use Bats as Bioindicators? Biology, 10(8), 693. https://doi.org/10.3390/biology10080693