
An Integrated Peptidomics and In Silico Approach to Identify Novel Anti-Diabetic Peptides in Parmigiano-Reggiano Cheese

 ,
,  ,
, 


Abstract
:Simple Summary
Abstract
1. Introduction
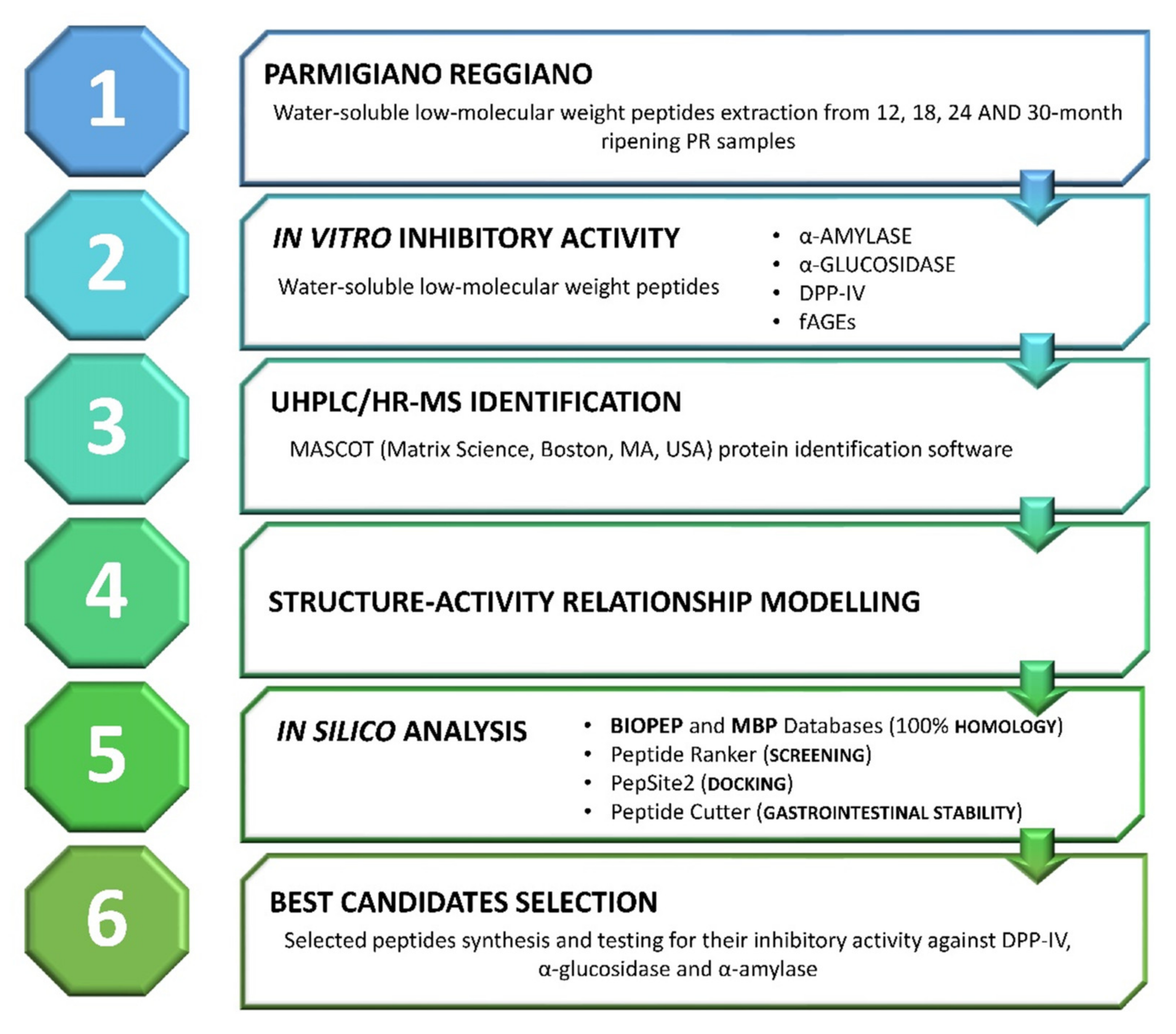
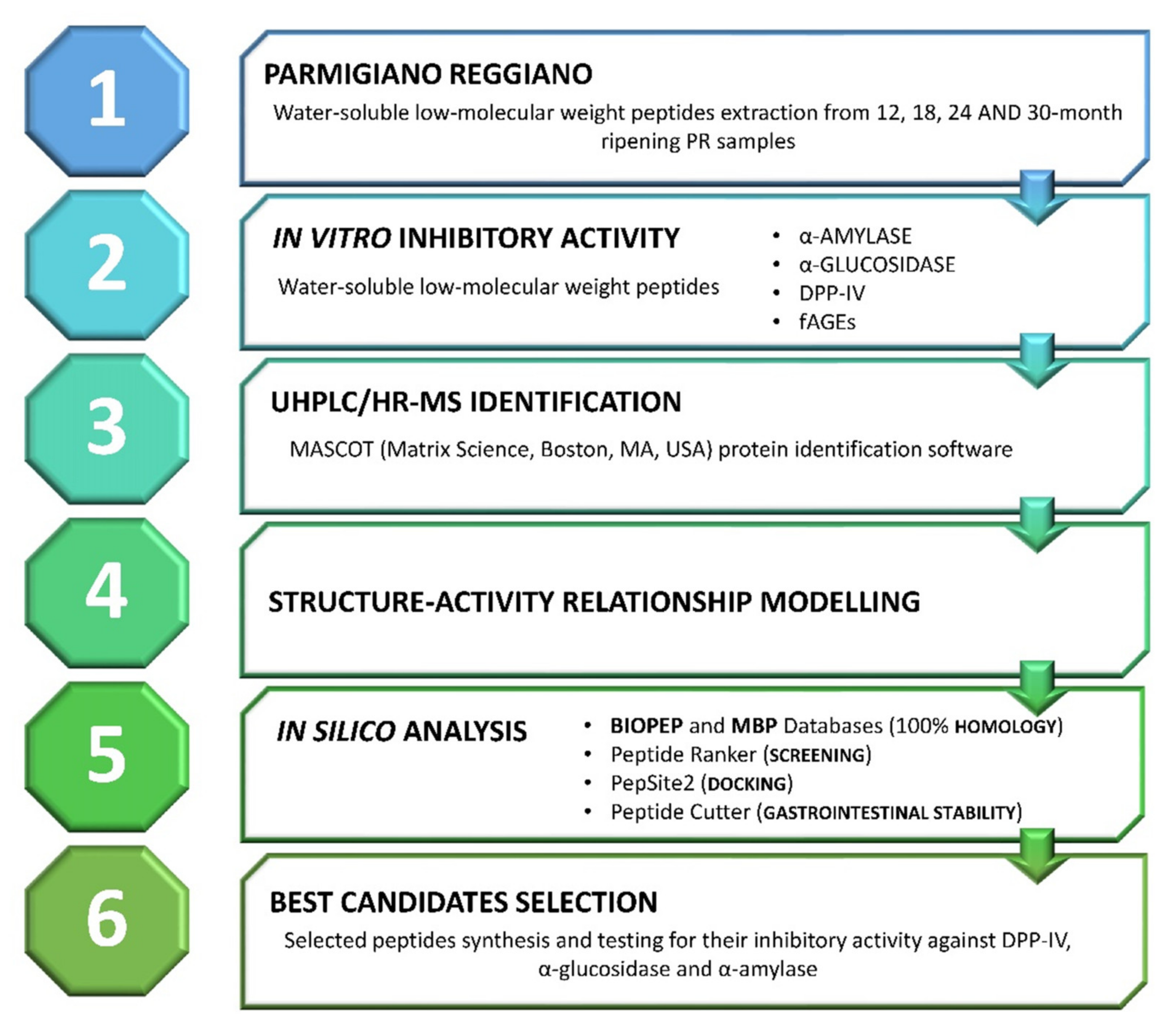
2. Materials and Methods
2.1. Materials
2.2. Extraction of Water-Soluble Low-Molecular Weight Peptides from Parmigiano-Reggiano Samples
2.3. Analysis of the Enzymatic Inhibitory Activity of Water-Soluble Low-Molecular Weight Peptides Extracted form PR Samples
2.3.1. α-Amylase Assay
2.3.2. α-Glucosidase Assay
2.3.3. Dipeptidyl Peptidase IV (DPP-IV) Assay
2.4. Advanced Glycation End-Products (AGEs) Inhibitory Assay
2.5. Identification of Low Molecular Weight Peptides by Ultra-High-Performance Liquid Chromatography/High-Resolution Mass Spectrometry (UHPLC/HR-MS)
2.6. In Silico Analysis
2.6.1. Identification of Previously Reported Bioactive Peptides
2.6.2. Identification of New Potential Bioactive Peptides
2.7. Statistical Analysis
3. Results and Discussion
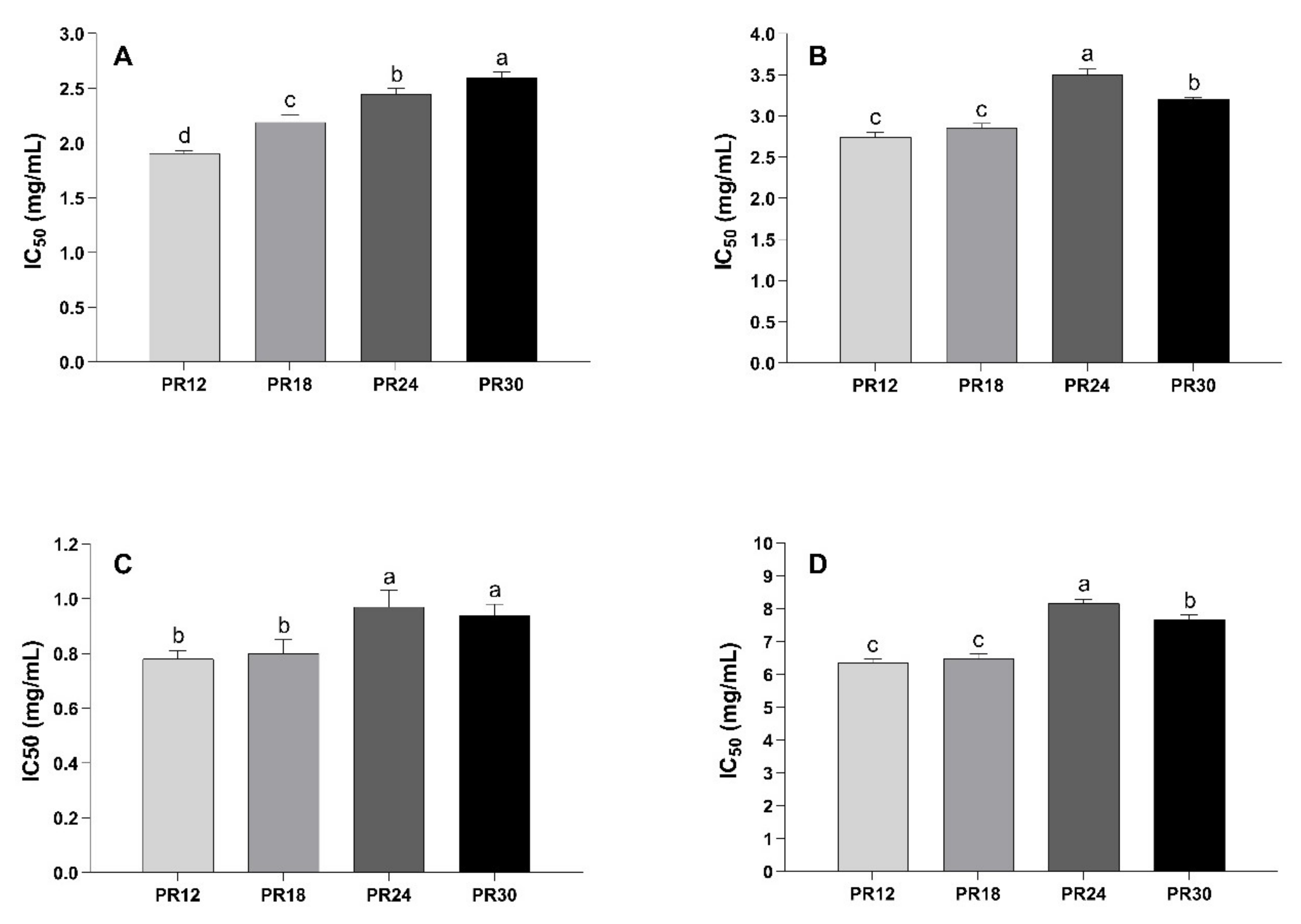
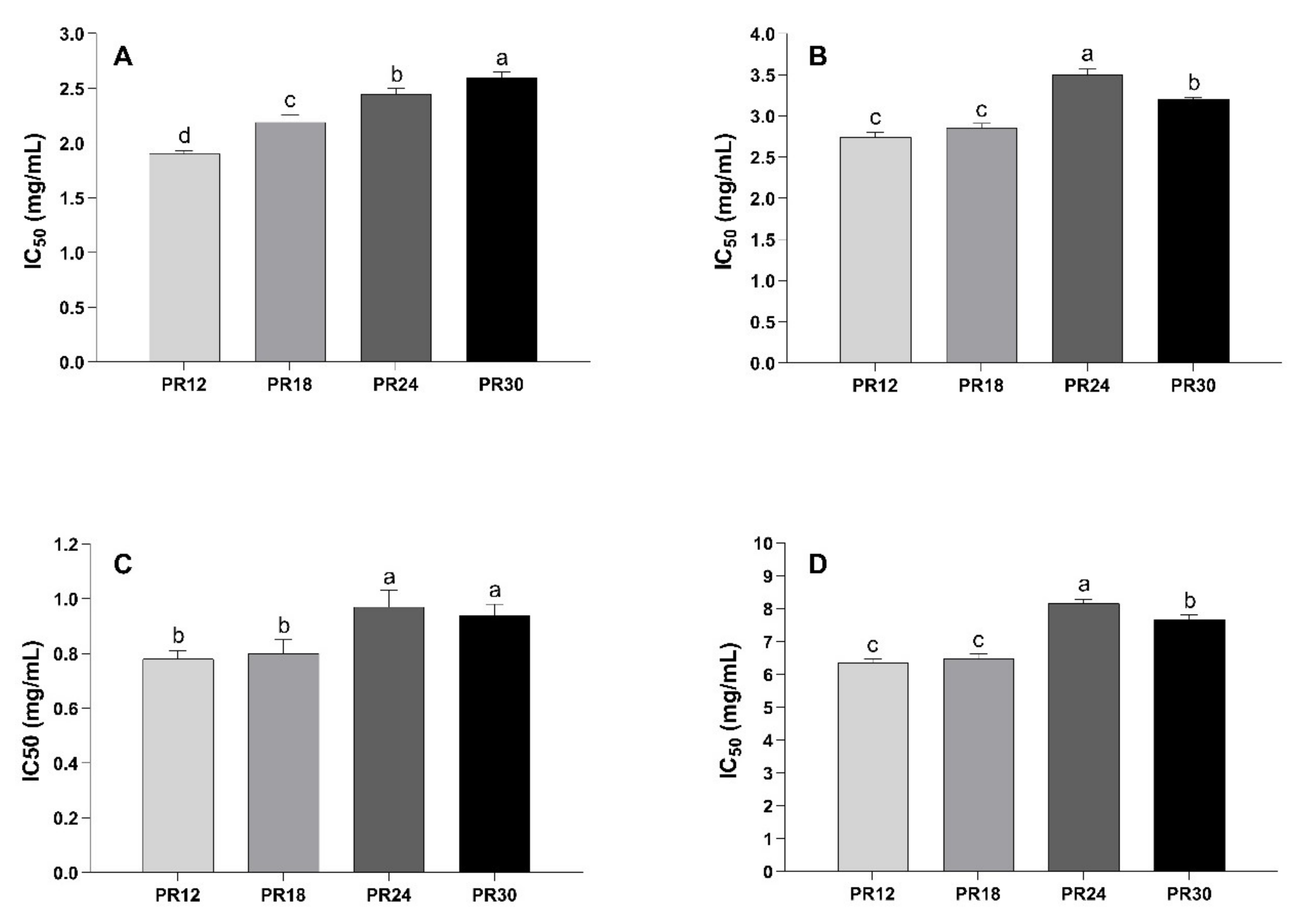
3.1. Inhibitory Effect of Water-Soluble Low-Molecular Weight Peptide Extracts of Parmigiano-Reggiano (PR) Samples on α-Amylase, α-Glucosidase, and Dipeptidyl Peptidase-IV (DPP-IV) Activities and Fluorescent Advanced Glycation End-Product (AGEs) Formation
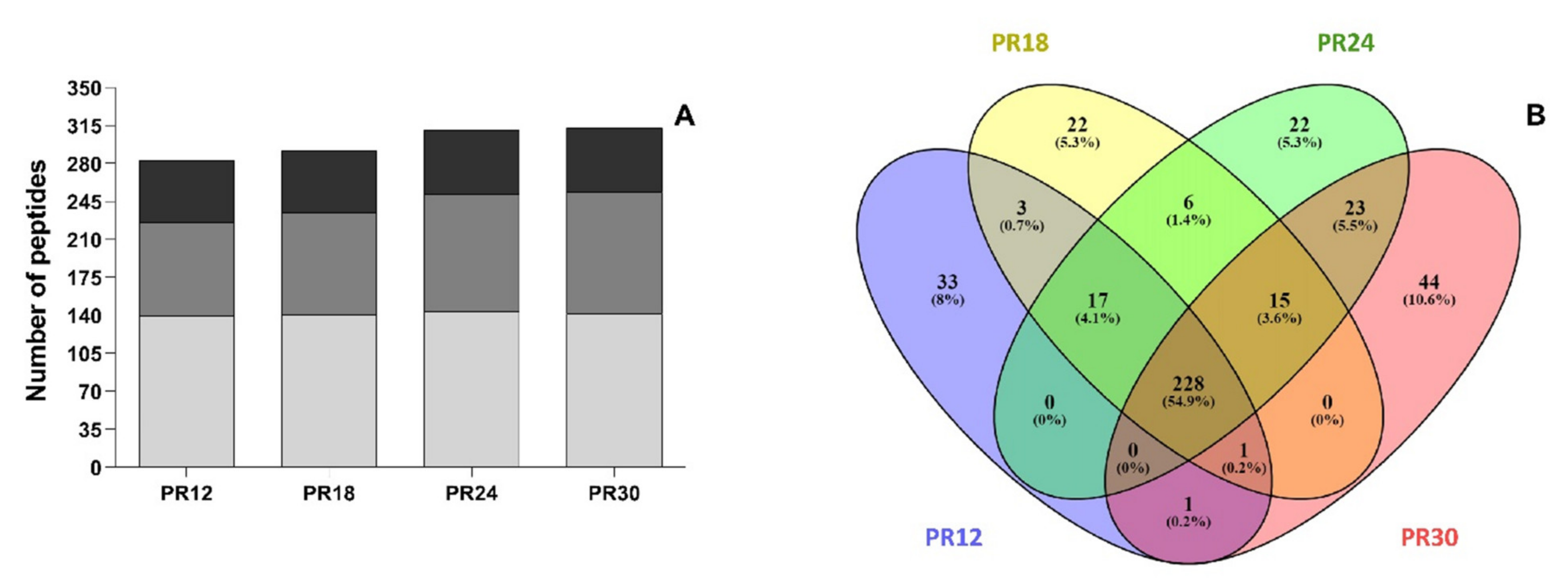
3.2. Peptidomics Profiles of Water-Soluble Low-Molecular Weight Peptide Extracts of Parmigiano-Reggiano (PR) Samples
3.3. Identification of Bioactive Peptides in Comparison with Databases
3.4. Selection of Potential Anti-Diabetic Peptides by In Silico Approach and Structure–Activity Relationship Modeling
3.5. In Vitro Inhibitory Activity of Synthetic Selected Peptides
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.; Ren, Z.H.; Qiang, H.; Wu, J.; Shen, M.; Zhang, L.; Lyu, J. Trends in the incidence of diabetes mellitus: Results from the Global Burden of Disease Study 2017 and implications for diabetes mellitus prevention. BMC Public Health 2020, 20, 1415. [Google Scholar] [CrossRef]
- Gerich, J.E. Clinical significance, pathogenesis, and management of post-prandial hyperglycemia. Arch. Intern. Med. 2003, 163, 1306–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalot, F.; Pagliarino, A.; Valle, M.; Di Martino, L.; Bonomo, K.; Massucco, P.; Anfossi, G.; Trovati, M. Postprandial blood glucose predicts cardiovascular events and all-cause mortality in type 2 diabetes in a 14-year follow-up: Lessons from the San Luigi Gonzaga Diabetes study. Diabetes Care 2011, 34, 2237–2243. [Google Scholar] [CrossRef] [Green Version]
- Cox, D.J.; Fang, K.; McCall, A.L.; Conaway, M.R.; Banton, T.A.; Moncrief, M.A.; Diamond, A.M.; Taylor, A.G. Behavioral strategies to lower postprandial glucose in those with type 2 diabetes may also lower risk of coronary heart disease. Diabetes Ther. 2019, 10, 277–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennerz, B.S.; Koutink, A.P.; Azova, S.; Wolfsdorf, J.I.; Ludwig, D.S. Carbohydrate restriction for diabetes: Rediscovering centuries-old wisdom. J. Clin. Investig. 2021, 131, e142246. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.G.; Gan, C.Y. In vivo challenges of anti-diabetic peptide therapeutics: Gastrointestinal stability, toxicity and allergenicity. Trends Food Sci Technol. 2020, 105, 161–175. [Google Scholar] [CrossRef]
- Rivero-Pino, F.; Espejo-Carpio, F.J.; Guadix, E.M. Antidiabetic food-derived peptides for functional feeding: Production, functionality and in vivo evidences. Foods 2020, 9, 983. [Google Scholar] [CrossRef]
- Joshi, S.R.; Standl, E.; Tong, N.; Shah, P.; Kalra, S.; Rathod, R. Therapeutic potential of α-glucosidase inhibitors in type 2 diabetes mellitus: An evidence-based review. Expert Opin. Pharmacother. 2015, 16, 1959–1981. [Google Scholar] [CrossRef]
- Li, W.; Yuan, G.; Pan, Y.; Wang, C.; Chen, H. Network pharmacology studies on the bioactive compounds and action mechanisms of natural products for the treatment of diabetes mellitus: A review. Front. Pharmacol. 2017, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- Iwaniak, A.; Mogut, D. Metabolic syndrome-preventive peptides derived from milk proteins and their presence in cheeses: A review. Appl. Sci. 2020, 10, 2772. [Google Scholar] [CrossRef]
- Deacom, C.F. Dipeptidyl peptidase 4 inhibitors in the treatment of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2020, 16, 642–653. [Google Scholar] [CrossRef]
- Wautier, J.L.; Guillausseasu, P.J. Advanced glycation end products, their receptors and diabetic angiopathy. Diabetes Metab. 2001, 27, 535–542. [Google Scholar] [CrossRef]
- Rhee, S.Y.; Kim, Y.S. The role of advanced glycation end products in diabetic vascular complications. Diabetes Metab. J. 2018, 42, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Jud, P.; Sourij, H. Therapeutic options to reduce advanced glycation end products in patients with diabetes mellitus: A review. Diabetes Res. Clin. Pract. 2019, 148, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Givens, D.I.; Astrup, A.; Bakker, S.J.L.; Goossens, G.H.; Kratz, M.; Marette, A.; Pijl, A.; Soedamah-Muthu, S.S. The impact of dairy products in the development of type 2 diabetes: Where does the evidence stand in 2019? Adv. Nutr. 2019, 10, 1066–1075. [Google Scholar] [CrossRef] [Green Version]
- Mitri, J.; Yusof, B.N.M.; Maryniuk, M.; Schrager, C.; Hamdy, O.; Salsberg, V. Dairy intake and type 2 diabetes risk factors: A narrative review. Diabetes Metab. Syndr. 2019, 13, 2879–2887. [Google Scholar] [CrossRef] [PubMed]
- Drehmer, M.; Pereira, M.A.; Schmidt, M.I.; Molina, M.D.C.B.; Alvim, S.; Lotufo, P.A.; Duncan, B.B. Associations of dairy intake with glycemia and insulinemia, independent of obesity, in Brazilian adults: The Brazilian Longitudinal Study of Adult Health (ELSA-Brasil). Am. J. Clin. Nutr. 2015, 101, 775–782. [Google Scholar] [CrossRef] [Green Version]
- Tagliazucchi, D.; Martini, S.; Solieri, L. Bioprospecting for bioactive peptide production by lactic acid bacteria isolated from fermented dairy food. Fermentation 2019, 5, 96. [Google Scholar] [CrossRef] [Green Version]
- Grom, L.C.; Coutinho, N.M.; Guimarães, J.T.; Balthazar, C.F.; Silva, R.; Rocha, R.S.; Freitas, M.Q.; Duarte, M.C.K.H.; Pimentel, C.T.; Esmerino, E.A.; et al. Probiotic dairy foods and postprandial glycemia: A mini-review. Trends Food Sci. Technol. 2020, 101, 165–171. [Google Scholar] [CrossRef]
- Summer, A.; Formaggioni, P.; Franceschi, P.; Di Frangia, F.; Righi, F.; Malacarne, M. Cheese as functional food: The example of Parmigiano-Reggiano and Grana Padano. Food Technol. Biotechnol. 2017, 55, 277–289. [Google Scholar]
- Solieri, L.; Baldaccini, A.; Martini, S.; Bianchi, A.; Pizamiglio, V.; Tagliazucchi, D. Peptide profiling and biological activities of 12-month ripened Parmigiano Reggiano cheese. Biology 2020, 9, 170. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Tagliazucchi, D.; Babini, E.; Rutella, G.S.; Taneyo Saa, D.L.; Gianotti, A. Bioactive peptides from vegetable food matrices: Research trends and novel biotechnologies for synthesis and recovery. J. Funct. Foods 2016, 27, 549–569. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Baldaccini, A.; Martini, S.; Bianchi, A.; Pizzamiglio, V.; Solieri, L. Cultivable non-starter lactobacilli from ripened Parmigiano Reggiano cheeses with different salt content and their potential to release anti-hypertensive peptides. Int. J. Food Microbiol. 2020, 330, 108688. [Google Scholar] [CrossRef] [PubMed]
- Sforza, S.; Cavatorta, V.; Lambertini, F.; Galaverna, G.; Dossena, A.; Marchelli, R. Cheese peptidomics: A detailed study on the evolution of the oligopeptide fraction in Parmigiano-Reggiano cheese from curd to 24 months of aging. J. Dairy Sci. 2012, 95, 3514–3526. [Google Scholar] [CrossRef] [Green Version]
- Basiricò, L.; Catalani, E.; Morera, P.; Cattaneo, S.; Stuknyte, M.; Bernabucci, U.; De Noni, I.; Nardone, A. Release of angiotensin converting enzyme-inhibitor peptides during in vitro gastrointestinal digestion of Parmigiano-Reggiano PDO cheese and their absorption through an in vitro model of intestinal epithelium. J. Dairy Sci. 2015, 98, 7595–7601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini, S.; Conte, A.; Tagliazucchi, D. Effect of ripening and in vitro digestion on the evolution and fate of bioactive peptides in Parmigiano-Reggiano cheese. Int. Dairy J. 2020, 105, 104668. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Shamsia, S.; Helal, A.; Conte, A. Angiotensin-converting enzyme inhibitory peptides from goats’ milk released by in vitro gastro-intestinal digestion. Int. Dairy J. 2017, 71, 6–16. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzensulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- McDougall, G.J.; Shapiro, F.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Different polyphenolic components of soft fruits inhibit a-amylase and a-glucosidase. J. Agric. Food Chem. 2005, 53, 2760–2766. [Google Scholar] [CrossRef]
- Bellesia, A.; Verzelloni, E.; Tagliazucchi, D. Pomegranate ellagitannins inhibit α-glucosidase activity in vitro and reduce starch digestibility under simulated gastrointestinal conditions. Int. J. Food Sci. Nutr. 2015, 66, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliazucchi, D.; Martini, S.; Shamsia, S.; Helal, A.; Conte, A. Biological activity and peptidomic profile of in vitro digested cow, camel, goat and sheep milk. Int. Dairy J. 2018, 81, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Tagliazucchi, D.; Martini, S.; Conte, A. Protocatechuic and 3,4-dihydroxyphenylacetic acids inhibit protein glycation by binding lysine through a metal-catalyzed oxidative mechanism. J. Agric. Food Chem. 2019, 67, 7821–7831. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, S.D.; Beverly, R.L.; Qu, Y.; Dallas, D.C. Milk bioactive peptide database: A comprehensive database of milk protein-derived bioactive peptides and novel visualization. Food Chem. 2017, 232, 673–682. [Google Scholar] [CrossRef]
- Mooney, C.; Haslam, N.J.; Pollastri, G.; Shields, D.C. Towards the improved discovery and design of functional peptides: Common features of diverse classes permit generalized prediction of bioactivity. PLoS ONE 2012, 7, e45012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trabuco, L.G.; Lise, S.; Petsalaki, E.; Russell, R.B. PepSite: Prediction of peptide-binding sites from protein surfaces. Nucleic Acids Res. 2012, 40, W423–W427. [Google Scholar] [CrossRef] [Green Version]
- Al-Dhaheri, A.S.; Al-Hemeiri, R.; Kizhakkayil, J.; Al-Nabulsi, A.; Abushelaibi, A.; Shah, N.P.; Ayyash, M. Health-promoting benefits of low-fat akawi cheese made by exopolysaccharide-producing probiotic Lactobacillus plantarum isolated from camel milk. J. Dairy Sci. 2017, 100, 7771–7779. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, M.; Gani, A.; Masoodi, F.A. Himalayan cheese (Kalari/Kradi) fermented with different probiotic strains: In vitro investigation of nutraceutical properties. LWT 2019, 104, 53–60. [Google Scholar] [CrossRef]
- Mudgil, P.; Kamal, H.; Yuen, G.C.; Maqsood, S. Characterization and identification of novel antidiabetic and anti-obesity peptides from camel milk protein hydrolysates. Food Chem. 2018, 259, 46–54. [Google Scholar] [CrossRef]
- Admassu, H.; Gasmalla, M.A.A.; Yang, R.; Zhao, W. Identification of bioactive peptides with α-amylase inhibitory potential from enzymatic protein hydrolysates of red seaweed (Porphyra spp.). J. Agric. Food Chem. 2018, 66, 4872–4882. [Google Scholar] [CrossRef]
- Uenishi, H.; Kabuki, T.; Seto, Y.; Serizawa, A.; Nakajima, H. Isolation and identification of casein-derived dipeptidyl-peptidase 4 (DPP-4)-inhibitory peptide LPQNIPPL from gouda-type cheese and its effect on plasma glucose in rats. Int. Dairy J. 2012, 22, 24–30. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food-an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Shi, F.; Bai, B.; Ma, S.; Ji, S.; Liu, L. The inhibitory effects of γ-glutamylcysteine derivatives from fresh garlic on glycation radical formation. Food Chem. 2016, 194, 538–544. [Google Scholar] [CrossRef]
- Freund, M.A.; Chen, B.; Decker, E.A. The inhibition of advanced glycation end products by carnosine and other natural dipeptides to reduce diabetic and age-related complications. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1367–1378. [Google Scholar] [CrossRef] [Green Version]
- Han, C.H.; Lin, Y.S.; Lin, S.Y.; Hou, W.C. Antioxidant and antiglycation activities of the synthesised dipeptide, Asn-Trp, derived from computer-aided simulation of yam dioscorin hydrolysis and its analogue, Gln-Trp. Food Chem. 2014, 147, 195–202. [Google Scholar] [CrossRef]
- Bottari, B.; Levante, A.; Bancalari, E.; Sforza, S.; Bottesini, C.; Prandi, B.; De Filippis, S.; Ercolini, D.; Nocetti, M.; Gatti, M. The interrelationship between microbiota and peptides during ripening as a driver for Parmigiano Reggiano cheese quality. Front. Microbiol. 2020, 11, 581658. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Fogacci, F.; Colletti, A. Potential role of bioactive peptides in prevention and treatment of chronic diseases: A narrative review. Br. J. Pharmacol. 2017, 174, 1378–1394. [Google Scholar] [CrossRef]
- Miguel, M.; Recio, I.; Ramos, M.; Delgado, M.A.; Aleixandre, M.A. Antihypertensive effect of peptides obtained from Enterococcus faecalis-fermented milk in rats. J. Dairy Sci. 2006, 89, 3352–3359. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Rivera, L.; Ferreira Santos, P.; Miralles, B.; Carrón, R.; Montero, M.J.; Recio, I. Peptide fragments from β-casein f(134–138), HLPLP, generated by the action of rat blood plasma peptidases show potent antihypertensive activity. Food Res. Int. 2016, 88, 348–353. [Google Scholar] [CrossRef] [Green Version]
- Lahov, E.; Regelson, W. Antibacterial and inmunostimulating casein-derived substances from milk: Casecidin, isracidin peptides. Food Chem. Toxicol. 1996, 34, 131–145. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Inhibition of dipeptidyl peptidase IV (DPP-IV) by proline containing casein-derived peptides. J. Funct. Foods 2013, 5, 1909–1917. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; FitzGerald, R.J. Structure activity relationship modelling of milk protein-derived peptides with dipeptidyl peptidase IV (DPP-IV) inhibitory activity. Peptides 2016, 79, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.J.; Yi, S.H.; Lee, J.S. Production and characterization of a new α-glucosidase inhibitory peptide from Aspergillus oryzae N159-1. Mycobiology 2013, 41, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Tulipano, G.; Faggi, L.; Nardone, A.; Cocchi, D.; Caroli, A.M. Characterisation of the potential of β-lactoglobulin and α-lactalbumin as sources of bioactive peptides affecting incretin function: In silico and in vitro comparative studies. Int. Dairy J. 2015, 48, 66–72. [Google Scholar] [CrossRef]
- Power, O.; Nongonierma, A.B.; Jakeman, P.; FitzGerald, R.J. Food protein hydrolysates as a source of dipeptidyl peptidase IV inhibitory peptides for the management of type 2 diabetes. Proc. Nutr. Soc. 2014, 73, 34–46. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; FitzGerald, R.J. An in silico model to predict the potential of dietary proteins as sources of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides. Food Chem. 2014, 165, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.H.; Gaspar, A.R.M. Structural properties of bioactive peptides with α-glucosidase inhibitory activity. Chem. Biol. Drug Des. 2018, 91, 370–379. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Bu, T.; Zheng, J.; Liu, L.; He, G.; Wu, J. Preparation, bioavailability, and mechanism of emerging activities of Ile-Pro-Pro and Val-Pro-Pro. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1097–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.H.; Gaspar, A.R.M. Rational in silico design of novel α-glucosidase inhibitory peptides and in vitro evaluation of promising candidates. Biomed. Pharmacother. 2018, 107, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Peptide Sequence | Protein Precursor | Sample | Bioactivity a |
|---|---|---|---|
| DKIHP | β-casein f(47–51) | PR24 | ACE-inhibition |
| DKIHPF | β-casein f(47–52) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| DVPSERYLG | αS1-casein f(85–93) | PR30 | ACE-inhibition |
| EMPFPK | β-casein f(108–113) | PR24, PR30 | ACE-inhibition Anti-microbial |
| ENLLRF | αS1-casein f(18–23) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| FFVAP | αS1-casein f(23–27) | PR24 | ACE-inhibition |
| FGK | αS1-casein f(32–34) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| FVAP | αS1-casein f(24–27) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| GTQY | αS1-casein f(170–172) | PR18, PR24, PR30 | ACE-inhibition Antioxidant |
| IPP | β-casein f(74–76) | PR12, PR18, PR24, PR30 | ACE-inhibition Antioxidant Anti-inflammatory |
| IPPL | β-casein f(74–77) | PR12, PR18, PR24, PR30 | DPP-IV-inhibition |
| IPPLTQTPV | β-casein f(74–82) | PR18, PR24 | DPP-IV-inhibition |
| IVP | αS1-casein f(71–73) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| LEE | β-casein f(3–5) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| LHLPLP | β-casein f(133–138) | PR12, PR18 | ACE-inhibition |
| LLY | β-casein f(191–193) | PR18 | Antioxidant Immunomodulation Anti-inflammatory |
| LNF | αS2-casein f(161–163) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| LPLP | β-casein f(135–138) | PR12, PR18, PR24 | ACE-inhibition |
| LPQ | β-casein f(70–72) αS1-casein f(11–13) αS2-casein f(176–178) | PR18, PR24, PR30 | DPP-IV-inhibition |
| LVYPFP | β-casein f(58–63) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| LVYPFPGP | β-casein f(58–65) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| PEL | αS1-casein f(147–149) | PR12, PR18, PR24, PR30 | Antioxidant |
| PFP | β-casein f(61–63) | PR12, PR18, PR24, PR30 | ACE-inhibition α-glucosidase inhibition |
| PFPE | αS1-casein f(27–30) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| PGPIP | β-casein f(63–67) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| PGPIPN | β-casein f(63–68) | PR12, PR18, PR24, PR30 | ACE-inhibition Immunomodulation Anti-inflammatory |
| PLW | αS1-casein f(197–199) | PR30 | ACE-inhibition |
| QEPV | β-casein f(194–197) | PR12, PR18, PR24, PR30 | Immunomodulation |
| QGP | αS2-casein f(101–103) | PR18, PR24 | ACE-inhibition |
| QGPIVLNPWDQVKR | αS2-casein f(101–114) | PR30 | Antioxidant |
| RELEEL | β-casein f(1–6) | PR12, PR18, PR24, PR30 | Antioxidant |
| RPKHPIKHQGLPQEVLNENLLRF | αS1-casein f(1–23) | PR30 | Immunomodulation Anti-microbial |
| SLPQ | β-casein f(69–72) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| TEDELQDKIHPF | β-casein f(41–52) | PR24, PR30 | Anti-microbial |
| TKVIPYVRYL | αS2-casein f(198–207) | PR18, PR24, PR30 | Anti-microbial |
| TVY | αS2-casein f(182–184) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| VEP | β-casein f(116–118) | PR24, PR30 | ACE-inhibition |
| VLP | β-casein f(170–172) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| VPP | β-casein f(84–86) | PR12, PR18, PR24, PR30 | ACE-inhibition Antioxidant Anti-inflammatory |
| VVPP | β-casein f(83–86) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| VVVPPF | β-casein f(82–87) | PR12, PR18, PR24, PR30 | ACE-inhibition |
| WIQP | αS2-casein f(193–196) | PR24 | ACE-inhibition DPP-IV-inhibition |
| YLEQLLR | αS1-casein f(94–100) | PR24, PR30 | Anti-microbial |
| YLG | αS1-casein f(91–93) | PR12, PR18, PR24, PR30 | Antioxidant |
| YLGY | αS1-casein f(91–94) | PR12 | ACE-inhibition Antioxidant |
| YQEP | β-casein f(193–196) | PR12 | ACE-inhibition |
| YQEPVLGPVRGPFPIIV | β-casein f(193–209) | PR24, PR30 | ACE-inhibition Anti-microbial |
| YQGPIVLNPWDQVKR | αS2-casein f(100–114) | PR12, PR18, PR24, PR30 | ACE-inhibition Anti-microbial Immunomodulation |
| YQL | αS1-casein f(154–156) | PR18, PR24, PR30 | Antioxidant |
| Peptide Sequence | DPP-IV μmol/L | α-Glucosidase μmol/L | α-Amylase μmol/L |
|---|---|---|---|
| APFPE | 49.5 ± 0.5 | n.a. | n.a. |
| IPP | 168.8 ± 2.2 | 764.5 ± 15.0 | 763.5 ± 18.9 |
| LPPT | 495.3 ± 3.6 | n.a. | n.a. |
| PPF | 141.9 ± 1.7 | >1000 | n.a. |
| VVPPF | 698.7 ± 11.1 | >1000 | n.a. |
| VVVPPF | 609.7 ± 13.6 | >1000 | n.a. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martini, S.; Solieri, L.; Cattivelli, A.; Pizzamiglio, V.; Tagliazucchi, D. An Integrated Peptidomics and In Silico Approach to Identify Novel Anti-Diabetic Peptides in Parmigiano-Reggiano Cheese. Biology 2021, 10, 563. https://doi.org/10.3390/biology10060563
Martini S, Solieri L, Cattivelli A, Pizzamiglio V, Tagliazucchi D. An Integrated Peptidomics and In Silico Approach to Identify Novel Anti-Diabetic Peptides in Parmigiano-Reggiano Cheese. Biology. 2021; 10(6):563. https://doi.org/10.3390/biology10060563
Chicago/Turabian StyleMartini, Serena, Lisa Solieri, Alice Cattivelli, Valentina Pizzamiglio, and Davide Tagliazucchi. 2021. "An Integrated Peptidomics and In Silico Approach to Identify Novel Anti-Diabetic Peptides in Parmigiano-Reggiano Cheese" Biology 10, no. 6: 563. https://doi.org/10.3390/biology10060563
APA StyleMartini, S., Solieri, L., Cattivelli, A., Pizzamiglio, V., & Tagliazucchi, D. (2021). An Integrated Peptidomics and In Silico Approach to Identify Novel Anti-Diabetic Peptides in Parmigiano-Reggiano Cheese. Biology, 10(6), 563. https://doi.org/10.3390/biology10060563