
Reduced Retinal Degeneration in an Oxidative Stress Organ Culture Model through an iNOS-Inhibitor


 ,
,  and
and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
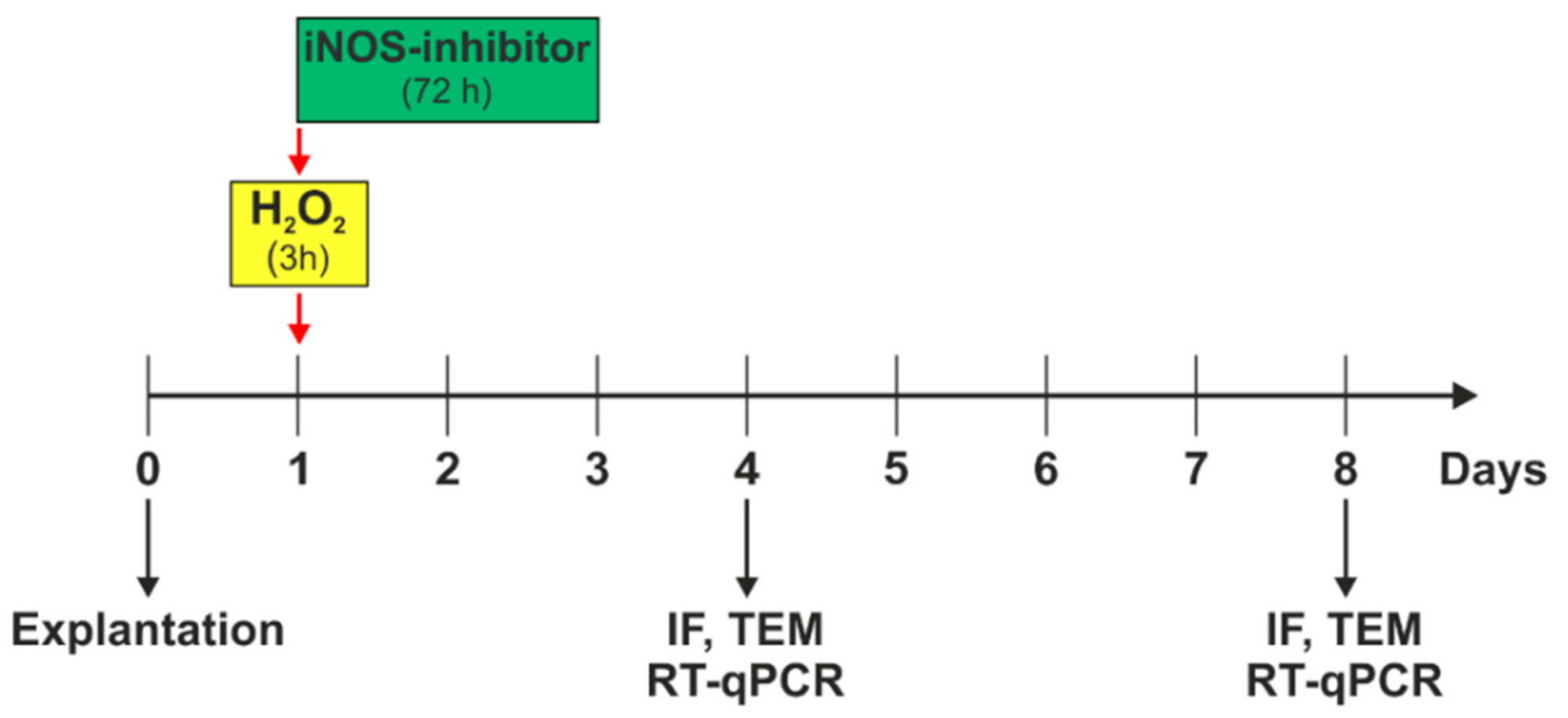
2.1. Subsection Preparation of Porcine Retinal Explants and Treatment Scheme of H2O2-Induced Degeneration
2.2. Analyses of Immunohistological Stainings
2.3. Analyses of Quantitative Real-Time PCR (RT-qPCR)
2.4. Transmission Electron Microscopy
2.5. Statistical Analyses
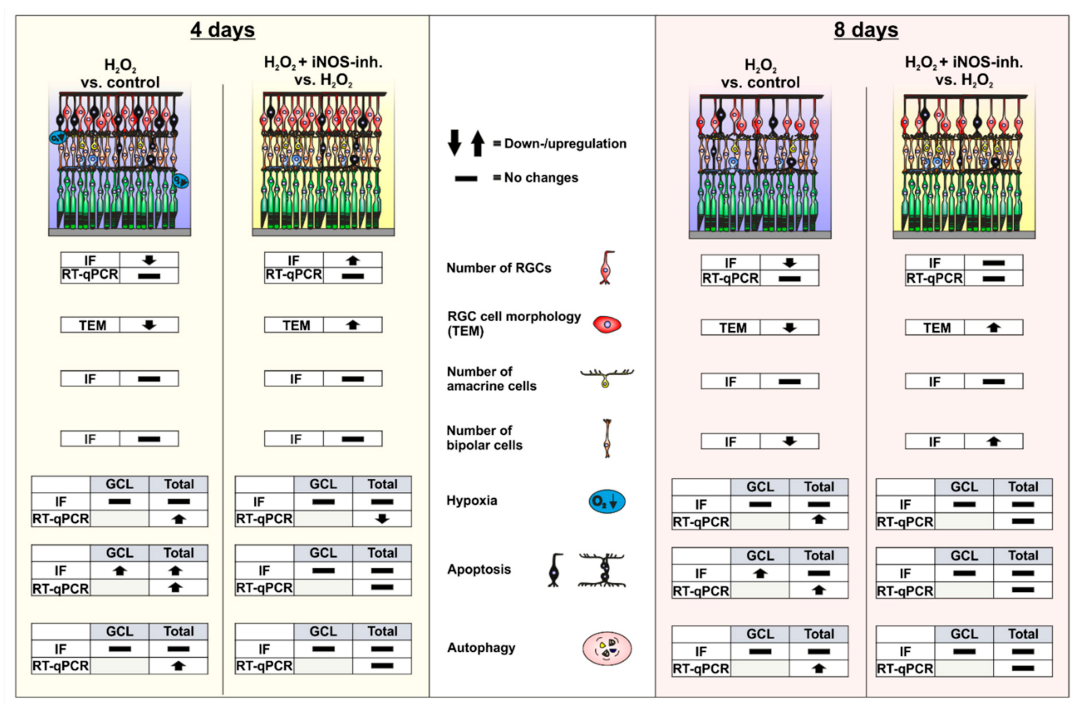
3. Results
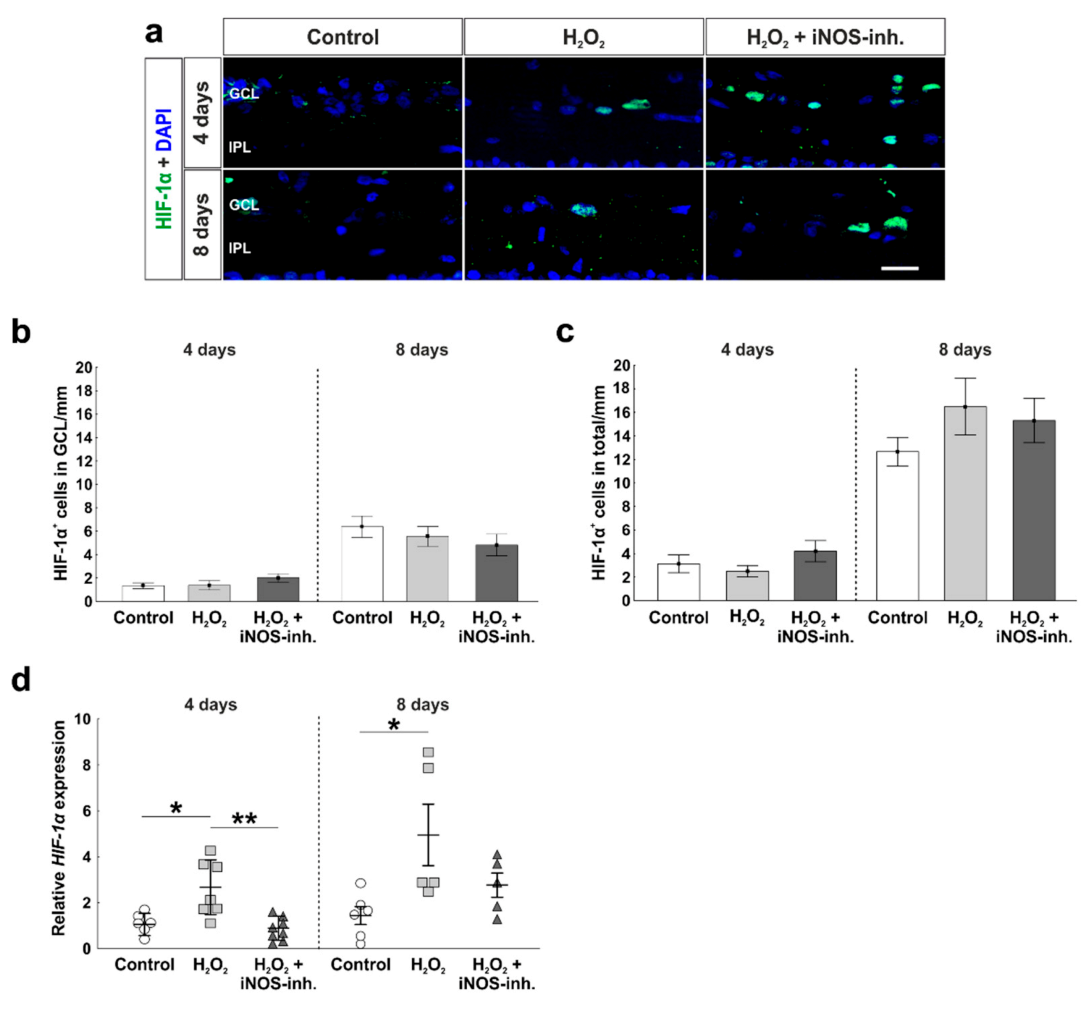
3.1. Early Reduction of Hypoxic Stress Marker HIF-1α after iNOS-Inhibitor Treatment
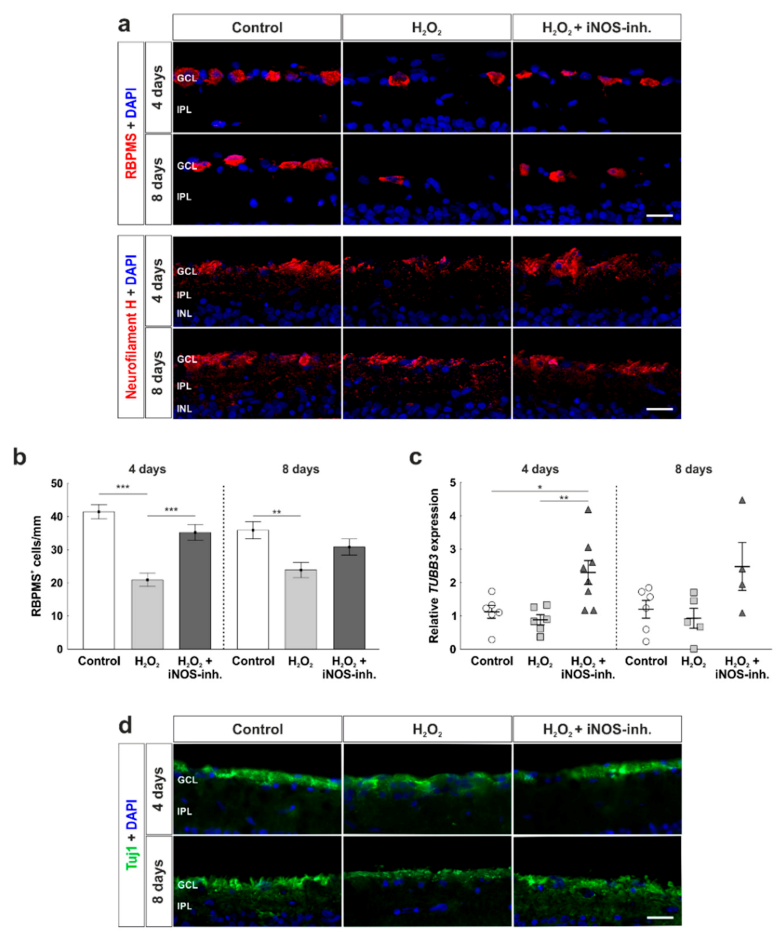
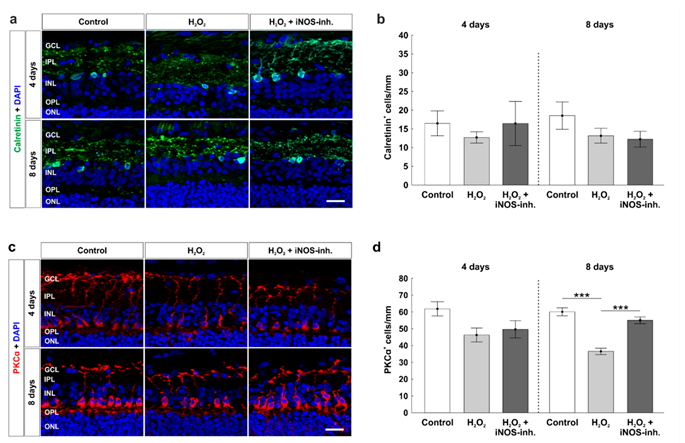
3.2. Protection of Retinal Ganglion Cells and Mitochondria
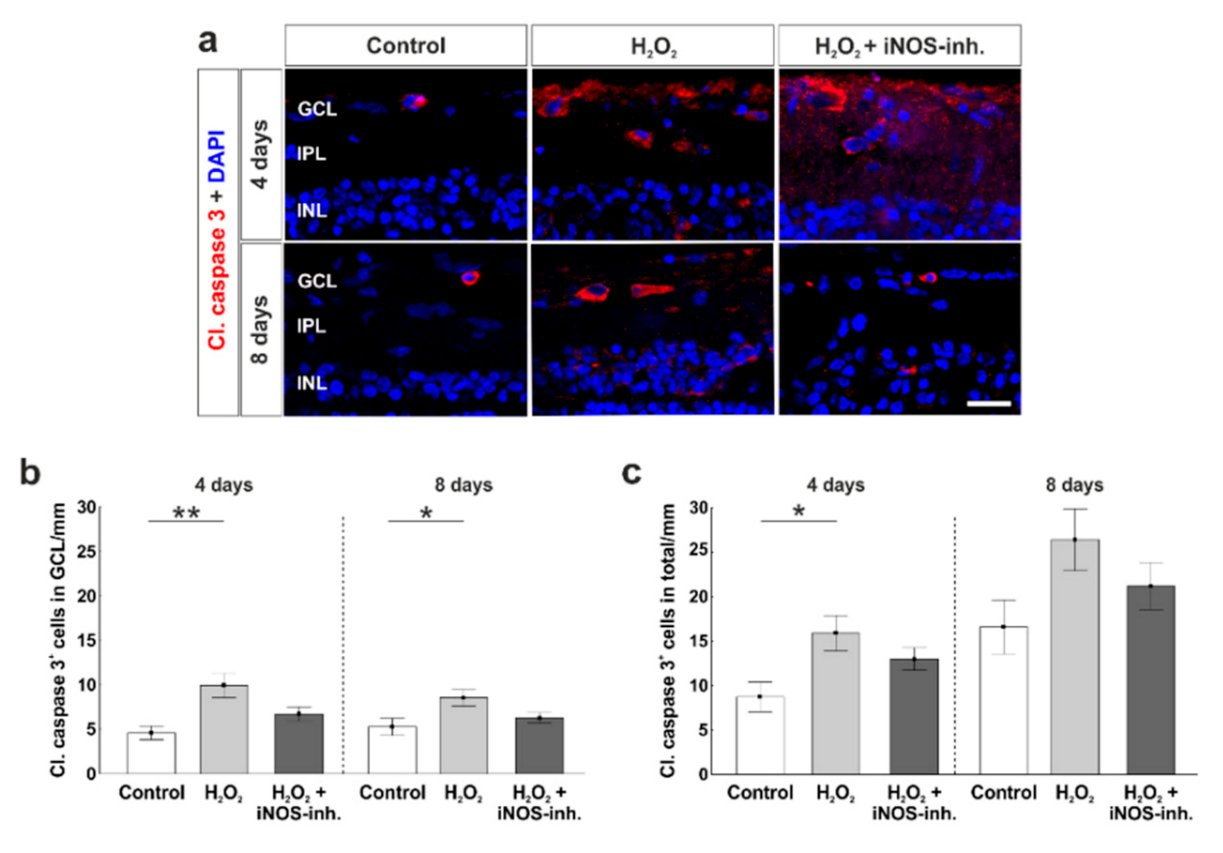
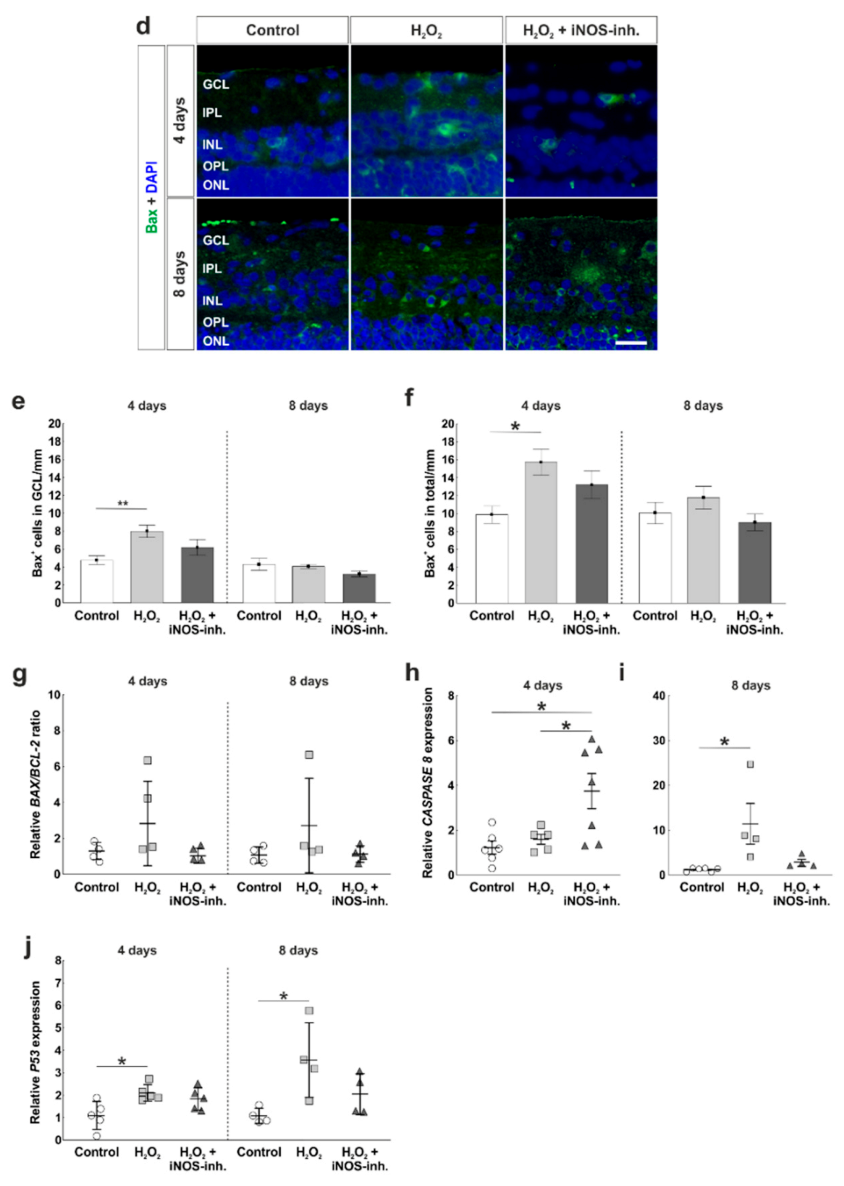
3.3. Slight Apoptosis after Oxidative Stress
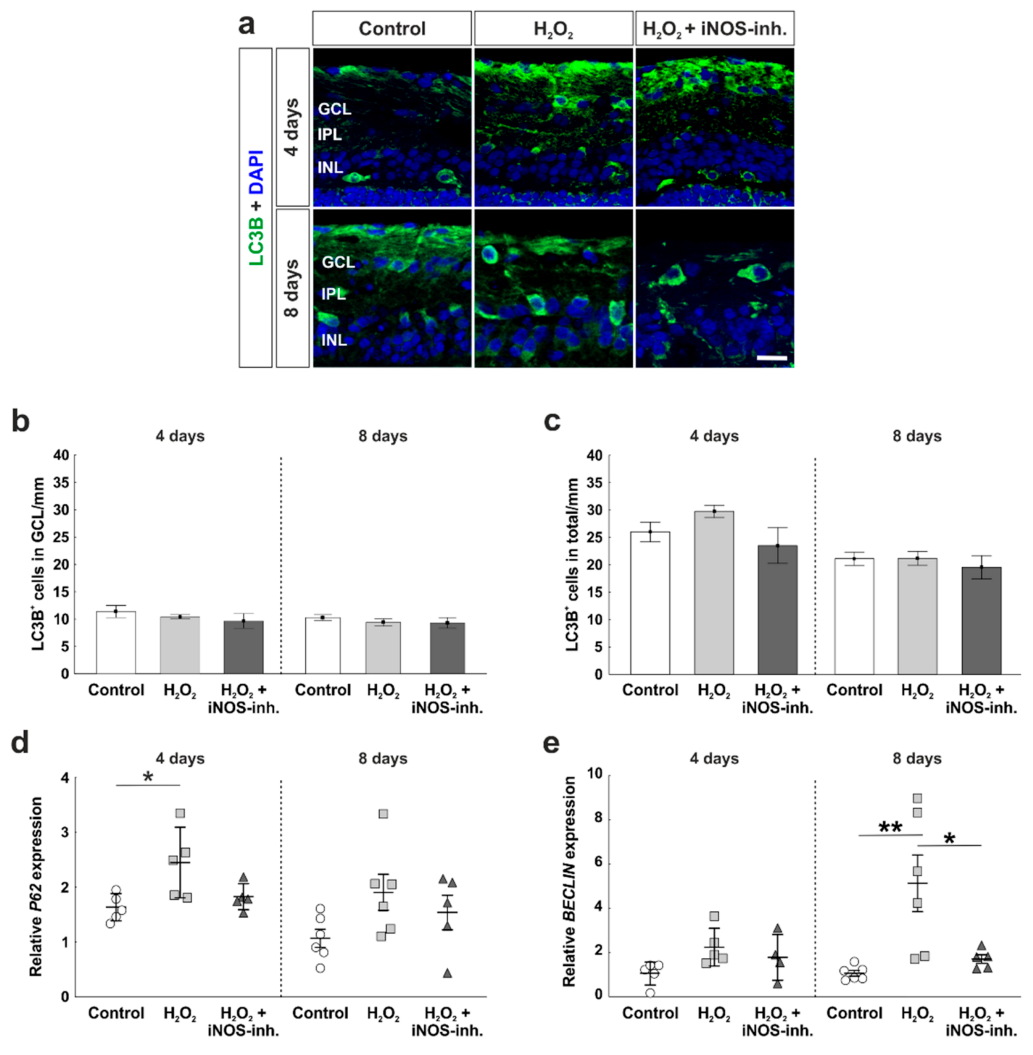
3.4. Early Activation of p62, an Autophagic Marker, through Oxidative Stress
3.5. Late Rescue of Rod Bipolar Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castro, N.; Gillespie, S.R.; Bernstein, A.M. Ex Vivo Corneal Organ Culture Model for Wound Healing Studies. J. Vis. Exp. 2019, 15, e58562. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.S.; Lalgudi, S.; Joshua, A.; Paul, J.; Thambaiah, A.; Wankhar, S.; Chacko, G.; Kuriakose, T.; Korah, S. An experimental study to test the efficacy of Mesenchymal Stem Cells in reducing corneal scarring in an ex-vivo organ culture model. Exp. Eye Res. 2020, 190, 107891. [Google Scholar] [CrossRef] [PubMed]
- Schnichels, S.; Paquet-Durand, F.; Loscher, M.; Tsai, T.; Hurst, J.; Joachim, S.C.; Klettner, A. Retina in a dish: Cell cultures, retinal explants and animal models for common diseases of the retina. Prog. Retin. Eye Res. 2020, 81, 100880. [Google Scholar] [CrossRef] [PubMed]
- Englund-Johansson, U.; Mohlin, C.; Liljekvist-Soltic, I.; Ekstrom, P.; Johansson, K. Human neural progenitor cells promote photoreceptor survival in retinal explants. Exp. Eye Res. 2010, 90, 292–299. [Google Scholar] [CrossRef]
- Hurst, J.; Mueller-Buehl, A.M.; Hofmann, L.; Kuehn, S.; Herms, F.; Schnichels, S.; Joachim, S.C. iNOS-inhibitor driven neuroprotection in a porcine retina organ culture model. J. Cell. Mol. Med. 2020, 24, 4312–4323. [Google Scholar] [CrossRef]
- Mueller-Buehl, A.M.; Doepper, H.; Grauthoff, S.; Kiebler, T.; Peters, L.; Hurst, J.; Kuehn, S.; Bartz-Schmidt, K.U.; Dick, H.B.; Joachim, S.C.; et al. Oxidative stress-induced retinal damage is prevented by mild hypothermia in an ex vivo model of cultivated porcine retinas. Clin. Exp. Ophthalmol. 2020, 48, 666–681. [Google Scholar] [CrossRef]
- Maliha, A.M.; Kuehn, S.; Hurst, J.; Herms, F.; Fehr, M.; Bartz-Schmidt, K.U.; Dick, H.B.; Joachim, S.C.; Schnichels, S. Diminished apoptosis in hypoxic porcine retina explant cultures through hypothermia. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef]
- Hurst, J.; Kuehn, S.; Jashari, A.; Tsai, T.; Bartz-Schmidt, K.U.; Schnichels, S.; Joachim, S.C. A novel porcine ex vivo retina culture model for oxidative stress induced by H2O2. Altern. Lab. Anim. 2017, 45, 11–25. [Google Scholar] [CrossRef]
- Hendrickson, A.; Hicks, D. Distribution and density of medium- and short-wavelength selective cones in the domestic pig retina. Exp. Eye Res. 2002, 74, 435–444. [Google Scholar] [CrossRef]
- Betteridge, D.J. What is oxidative stress? Metabolism 2000, 49, 3–8. [Google Scholar] [CrossRef]
- Richard, D.E.; Berra, E.; Pouyssegur, J. Nonhypoxic pathway mediates the induction of hypoxia-inducible factor 1alpha in vascular smooth muscle cells. J. Biol. Chem. 2000, 275, 26765–26771. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef]
- Christopherson, K.S.; Bredt, D.S. Nitric oxide in excitable tissues: Physiological roles and disease. J. Clin. Investig. 1997, 100, 2424–2429. [Google Scholar] [CrossRef]
- Robbins, R.A.; Grisham, M.B. Nitric oxide. Int. J. Biochem. Cell. Biol. 1997, 29, 857–860. [Google Scholar] [CrossRef]
- Roth, S. Role of nitric oxide in retinal cell death. Clin. Neurosci. 1997, 4, 216–223. [Google Scholar]
- Haefliger, I.O.; Dettmann, E.; Liu, R.; Meyer, P.; Prunte, C.; Messerli, J.; Flammer, J. Potential role of nitric oxide and endothelin in the pathogenesis of glaucoma. Surv. Ophthalmol. 1999, 43 (Suppl. 1), S51–S58. [Google Scholar] [CrossRef]
- Peterson, L.J.; Flood, P.M. Oxidative stress and microglial cells in Parkinson’s disease. Mediat. Inflamm. 2012, 2012, 401264. [Google Scholar] [CrossRef]
- Gella, A.; Durany, N. Oxidative stress in Alzheimer disease. Cell. Adhes. Migr. 2009, 3, 88–93. [Google Scholar] [CrossRef]
- Izzotti, A.; Bagnis, A.; Sacca, S.C. The role of oxidative stress in glaucoma. Mutat. Res. 2006, 612, 105–114. [Google Scholar] [CrossRef]
- Chiou, G.C. Review: Effects of nitric oxide on eye diseases and their treatment. J. Ocul. Pharmacol. Ther. 2001, 17, 189–198. [Google Scholar] [CrossRef]
- Vannini, F.; Kashfi, K.; Nath, N. The dual role of iNOS in cancer. Redox. Biol. 2015, 6, 334–343. [Google Scholar] [CrossRef]
- Carr, B.C.; Emigh, C.E.; Bennett, L.D.; Pansick, A.D.; Birch, D.G.; Nguyen, C. Towards a Treatment for Diabetic Retinopathy: Intravitreal Toxicity and Preclinical Safety Evaluation of Inducible Nitric Oxide Synthase Inhibitors. Retina 2017, 37, 22–31. [Google Scholar] [CrossRef]
- Schnichels, S.; Joachim, S.C. The inducible nitric oxide synthase-inhibitor 1400 W as a potential treatment for retinal diseases. Neural Regen. Res. 2021, 16, 1221–1222. [Google Scholar] [CrossRef]
- Rodriguez-Muela, N.; Germain, F.; Marino, G.; Fitze, P.S.; Boya, P. Autophagy promotes survival of retinal ganglion cells after optic nerve axotomy in mice. Cell. Death Differ. 2012, 19, 162–169. [Google Scholar] [CrossRef]
- Romano, G.L.; Amato, R.; Lazzara, F.; Porciatti, V.; Chou, T.H.; Drago, F.; Bucolo, C. P2X7 receptor antagonism preserves retinal ganglion cells in glaucomatous mice. Biochem. Pharmacol. 2020, 180, 114199. [Google Scholar] [CrossRef]
- Kuehn, S.; Reinehr, S.; Stute, G.; Rodust, C.; Grotegut, P.; Hensel, A.T.; Dick, H.B.; Joachim, S.C. Interaction of complement system and microglia activation in retina and optic nerve in a NMDA damage model. Mol. Cell. Neurosci. 2018, 89, 95–106. [Google Scholar] [CrossRef]
- Eguchi, H.; Fujiwara, N.; Sakiyama, H.; Yoshihara, D.; Suzuki, K. Hydrogen peroxide enhances LPS-induced nitric oxide production via the expression of interferon beta in BV-2 microglial cells. Neurosci. Lett. 2011, 494, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Vielma, A.H.; Retamal, M.A.; Schmachtenberg, O. Nitric oxide signaling in the retina: What have we learned in two decades? Brain. Res. 2012, 1430, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Vitecek, J.; Lojek, A.; Valacchi, G.; Kubala, L. Arginine-based inhibitors of nitric oxide synthase: Therapeutic potential and challenges. Mediat. Inflamm. 2012, 2012, 318087. [Google Scholar] [CrossRef] [PubMed]
- Rokicki, W.; Zaba, M.; Wygledowska-Promienska, D.; Kabiesz, A.; Reichman-Warmusz, E.; Brzozowa, M.; Majewski, W.; Wojnicz, R. Inducible and endothelial nitric synthetase expression and nitrotyrosine accumulation in iris vasculature of patients with primary open-angle glaucoma: A pilot study. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2015, 21, 76–81. [Google Scholar] [CrossRef]
- Koeberle, P.D.; Ball, A.K. Nitric oxide synthase inhibition delays axonal degeneration and promotes the survival of axotomized retinal ganglion cells. Exp. Neurol. 1999, 158, 366–381. [Google Scholar] [CrossRef]
- Garvey, E.P.; Oplinger, J.A.; Furfine, E.S.; Kiff, R.J.; Laszlo, F.; Whittle, B.J.; Knowles, R.G. 1400 W is a slow, tight binding, and highly selective inhibitor of inducible nitric-oxide synthase in vitro and in vivo. J. Biol. Chem. 1997, 272, 4959–4963. [Google Scholar] [CrossRef]
- Parmentier, S.; Bohme, G.A.; Lerouet, D.; Damour, D.; Stutzmann, J.M.; Margaill, I.; Plotkine, M. Selective inhibition of inducible nitric oxide synthase prevents ischaemic brain injury. Br. J. Pharmacol. 1999, 127, 546–552. [Google Scholar] [CrossRef]
- Brzozowski, M.J.; Jenner, P.; Rose, S. Inhibition of i-NOS but not n-NOS protects rat primary cell cultures against MPP+-induced neuronal toxicity. J. Neural. Transm. 2015, 122, 779–788. [Google Scholar] [CrossRef]
- Knels, L.; Worm, M.; Wendel, M.; Roehlecke, C.; Kniep, E.; Funk, R.H. Effects of advanced glycation end products-inductor glyoxal and hydrogen peroxide as oxidative stress factors on rat retinal organ cultures and neuroprotection by UK-14,304. J. Neurochem. 2008, 106, 1876–1887. [Google Scholar] [CrossRef]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural. Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef]
- Lee, J.; Giordano, S.; Zhang, J. Autophagy, mitochondria and oxidative stress: Cross-talk and redox signalling. Biochem. J. 2012, 441, 523–540. [Google Scholar] [CrossRef]
- Kowaltowski, A.J.; Castilho, R.F.; Vercesi, A.E. Mitochondrial permeability transition and oxidative stress. FEBS Lett. 2001, 495, 12–15. [Google Scholar] [CrossRef]
- Wang, C.; Youle, R.J. The role of mitochondria in apoptosis*. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef]
- Tengan, C.H.; Moraes, C.T. NO control of mitochondrial function in normal and transformed cells. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 573–581. [Google Scholar] [CrossRef]
- Sennlaub, F.; Courtois, Y.; Goureau, O. Inducible nitric oxide synthase mediates retinal apoptosis in ischemic proliferative retinopathy. J. Neurosci. 2002, 22, 3987–3993. [Google Scholar] [CrossRef] [PubMed]
- Vinas, J.L.; Sola, A.; Genesca, M.; Alfaro, V.; Pi, F.; Hotter, G. NO and NOS isoforms in the development of apoptosis in renal ischemia/reperfusion. Free Radic. Biol. Med. 2006, 40, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Lakin, N.D.; Jackson, S.P. Regulation of p53 in response to DNA damage. Oncogene 1999, 18, 7644–7655. [Google Scholar] [CrossRef] [PubMed]
- Frake, R.A.; Ricketts, T.; Menzies, F.M.; Rubinsztein, D.C. Autophagy and neurodegeneration. J. Clin. Investig. 2015, 125, 65–74. [Google Scholar] [CrossRef]
- Kabeya, Y.; Mizushima, N.; Ueno, T.; Yamamoto, A.; Kirisako, T.; Noda, T.; Kominami, E.; Ohsumi, Y.; Yoshimori, T. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 2000, 19, 5720–5728. [Google Scholar] [CrossRef]
- Komatsu, M.; Waguri, S.; Koike, M.; Sou, Y.S.; Ueno, T.; Hara, T.; Mizushima, N.; Iwata, J.; Ezaki, J.; Murata, S.; et al. Homeostatic levels of p62 control cytoplasmic inclusion body formation in autophagy-deficient mice. Cell 2007, 131, 1149–1163. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef]
- Song, C.; Mitter, S.K.; Qi, X.; Beli, E.; Rao, H.V.; Ding, J.; Ip, C.S.; Gu, H.; Akin, D.; Dunn, W.A., Jr.; et al. Oxidative stress-mediated NFkappaB phosphorylation upregulates p62/SQSTM1 and promotes retinal pigmented epithelial cell survival through increased autophagy. PLoS ONE 2017, 12, e0171940. [Google Scholar] [CrossRef]
- Wang, L.; Cano, M.; Handa, J.T. p62 provides dual cytoprotection against oxidative stress in the retinal pigment epithelium. Biochim. Biophys. Acta Mol. Cell. Res. 2014, 1843, 1248–1258. [Google Scholar] [CrossRef]
- Li, M.; Gao, P.; Zhang, J. Crosstalk between Autophagy and Apoptosis: Potential and Emerging Therapeutic Targets for Cardiac Diseases. Int. J. Mol. Sci. 2016, 17, 332. [Google Scholar] [CrossRef]
- Kimura, H.; Weisz, A.; Kurashima, Y.; Hashimoto, K.; Ogura, T.; D’Acquisto, F.; Addeo, R.; Makuuchi, M.; Esumi, H. Hypoxia response element of the human vascular endothelial growth factor gene mediates transcriptional regulation by nitric oxide: Control of hypoxia-inducible factor-1 activity by nitric oxide. Blood 2000, 95, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Sogawa, K.; Numayama-Tsuruta, K.; Ema, M.; Abe, M.; Abe, H.; Fujii-Kuriyama, Y. Inhibition of hypoxia-inducible factor 1 activity by nitric oxide donors in hypoxia. Proc. Natl. Acad. Sci. USA 1998, 95, 7368–7373. [Google Scholar] [CrossRef] [PubMed]
- Agurto, A.; Vielma, A.H.; Cadiz, B.; Couve, E.; Schmachtenberg, O. NO signaling in retinal bipolar cells. Exp. Eye Res. 2017, 161, 30–35. [Google Scholar] [CrossRef]
- Kuehn, S.; Hurst, J.; Rensinghoff, F.; Tsai, T.; Grauthoff, S.; Satgunarajah, Y.; Dick, H.B.; Schnichels, S.; Joachim, S.C. Degenerative effects of cobalt-chloride treatment on neurons and microglia in a porcine retina organ culture model. Exp. Eye Res. 2017, 155, 107–120. [Google Scholar] [CrossRef]
- Palmhof, M.; Frank, V.; Rappard, P.; Kortenhorn, E.; Demuth, J.; Biert, N.; Stute, G.; Dick, H.B.; Joachim, S.C. From Ganglion Cell to Photoreceptor Layer: Timeline of Deterioration in a Rat Ischemia/Reperfusion Model. Front. Cell. Neurosci. 2019, 13, 174. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Secondary Antibodies | |||||||
|---|---|---|---|---|---|---|---|---|
| Antibody | Source | Company | Research Resource Identification Number | Dilution | Antibody | Company | Research Resource Identification Number | Dilution |
| Anti-Bax | Rabbit | Abcam | ab7977 | 1:100 | Donkey anti-rabbit Alexa Fluor 488 | Jackson Immuno Research | 711-547-003 | 1:500 |
| Anti-calretinin | Goat | Merck Millipore | AB1550 | 1:2000 | Donkey anti-goat Alexa Flour 488 | Dianova | 705-545-147 | 1:500 |
| Anti-cleaved caspase 3 | Rabbit | Sigma-Aldrich | C8487 | 1:300 | Donkey anti-rabbit Alexa Fluor 555 | Invitrogen | A31572 | 1:500 |
| Anti-HIF-1α | Mouse | BD Biosciences | 610959 | 1:100 | Donkey anti-mouse Alexa Fluor 488 | Abcam | A21202 | 1:500 |
| Anti-LC3B | Rabbit | Cell Signaling | 3868 | 1:300 | Donkey anti-rabbit Alexa Fluor 488 | Jackson Immuno Research | 711-547-003 | 1:500 |
| Anti-neuro-filament H | Chicken | Synaptic Systems | 171106 | 1:300 | Donkey anti-chicken Cy3 | Millipore | AP194C | 1:500 |
| Anti-PKCα | Mouse | Santa Cruz | sc-8393 | 1:500 | Donkey anti-mouse Alexa Fluor 555 | Abcam | ab150106 | 1:500 |
| Anti-RBPMS | Rabbit | Merck Millipore | ABN1362 | 1:200 | Donkey anti-rabbit Alexa Fluor 555 | Invitrogen | A31572 | 1:500 |
| Anti-Tuj1 | Mouse | Covance | MMS-435P-100 | 1:300 | Donkey anti-mouse Alexa Flour 488 | Abcam | A21202 | 1:500 |
| Gene | Oligonucleotide Sequence (3′-5′) |
|---|---|
| ACTB for | CACGCCATCCTGCGTCTGGA |
| ACTB rev | AGCACCGTGTTGGCGTAGAG |
| BAX for | AAGCGCATTGGAGATGAACT |
| BAX rev | AAAGTAGAAAAGCGCGACCA |
| BCL-2 for | AATTACCATCGGCGTAGTGC |
| BCL-2 rev | CGTTTCAGCCACCGTAAAAT |
| BECLIN for | AGGAGCTGCCGTTGTACTGT |
| BECLIN rev | CACTGCCTCCTGTGTCTTCA |
| CASPASE 8 for | GCCCAGATCTCTGCCTACAG |
| CASPASE 8 rev | CAGGGCCTTGTTGATTTGTT |
| HIF-1α for | GTAATGCTCCCCTCATCCAA |
| HIF-1α rev | TGGGGCATGGTAAAAGAAAG |
| P53 for | CCTCACCATCATCACACTGG |
| P53 rev | GGCTTCTTCTTTTGCACTGG |
| P62 rev | ATGGGTCCAGTCATCGTCTC |
| P62 for | TCCAGCACAGAGGACAAGTG |
| RPL4 for | CAAGAGTAACTACAACCTTC |
| RPL4 rev | GAACTCTACGATGAATCTTC |
| TUBB3 for | CAGATGTTCGATGCCAAGAA |
| TUBB3 rev | GGGATCCACTCCACGAAGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mueller-Buehl, A.M.; Tsai, T.; Hurst, J.; Theiss, C.; Peters, L.; Hofmann, L.; Herms, F.; Kuehn, S.; Schnichels, S.; Joachim, S.C. Reduced Retinal Degeneration in an Oxidative Stress Organ Culture Model through an iNOS-Inhibitor. Biology 2021, 10, 383. https://doi.org/10.3390/biology10050383
Mueller-Buehl AM, Tsai T, Hurst J, Theiss C, Peters L, Hofmann L, Herms F, Kuehn S, Schnichels S, Joachim SC. Reduced Retinal Degeneration in an Oxidative Stress Organ Culture Model through an iNOS-Inhibitor. Biology. 2021; 10(5):383. https://doi.org/10.3390/biology10050383
Chicago/Turabian StyleMueller-Buehl, Ana M., Teresa Tsai, José Hurst, Carsten Theiss, Laura Peters, Lisa Hofmann, Fenja Herms, Sandra Kuehn, Sven Schnichels, and Stephanie C. Joachim. 2021. "Reduced Retinal Degeneration in an Oxidative Stress Organ Culture Model through an iNOS-Inhibitor" Biology 10, no. 5: 383. https://doi.org/10.3390/biology10050383
APA StyleMueller-Buehl, A. M., Tsai, T., Hurst, J., Theiss, C., Peters, L., Hofmann, L., Herms, F., Kuehn, S., Schnichels, S., & Joachim, S. C. (2021). Reduced Retinal Degeneration in an Oxidative Stress Organ Culture Model through an iNOS-Inhibitor. Biology, 10(5), 383. https://doi.org/10.3390/biology10050383