
Identification of an RNA Silencing Suppressor Encoded by a Symptomless Fungal Hypovirus, Cryphonectria Hypovirus 4

 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Viral and Fungal Strain
2.2. Fungal Transformation
2.3. Functional Expression in Escherichia coli of the Putative p24 Proteinase Domain of CHV4
2.4. RNA Extraction, RT-PCR and RNA Blotting
2.5. Small RNA Analysis
2.6. Confocal Laser Scanning Microscopy of the Reporter Fungal Strain Infected by Diverse Viruses
3. Results
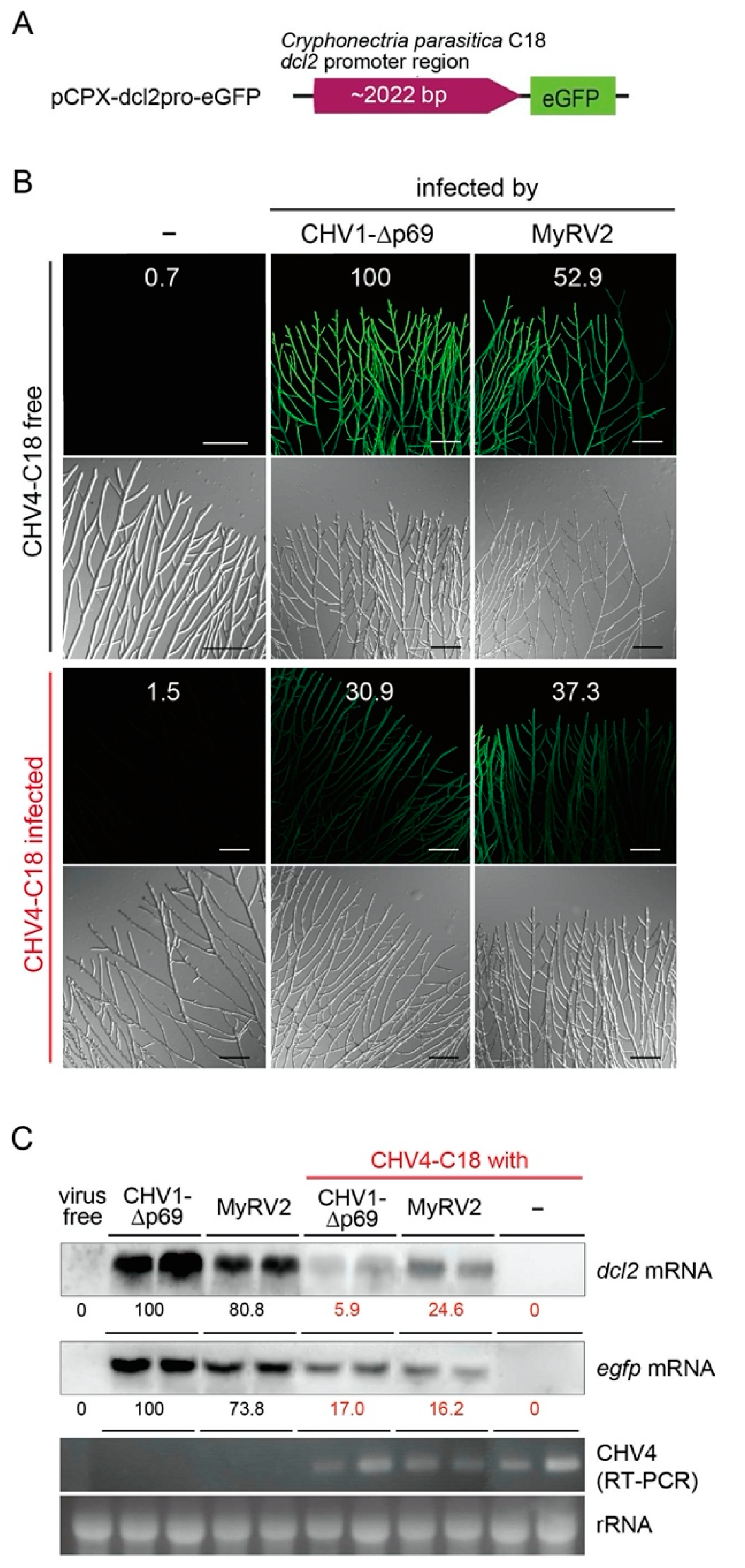
3.1. Cryphonectria Parasitica Hypovirus 4 Suppresses Upregulation of dcl2
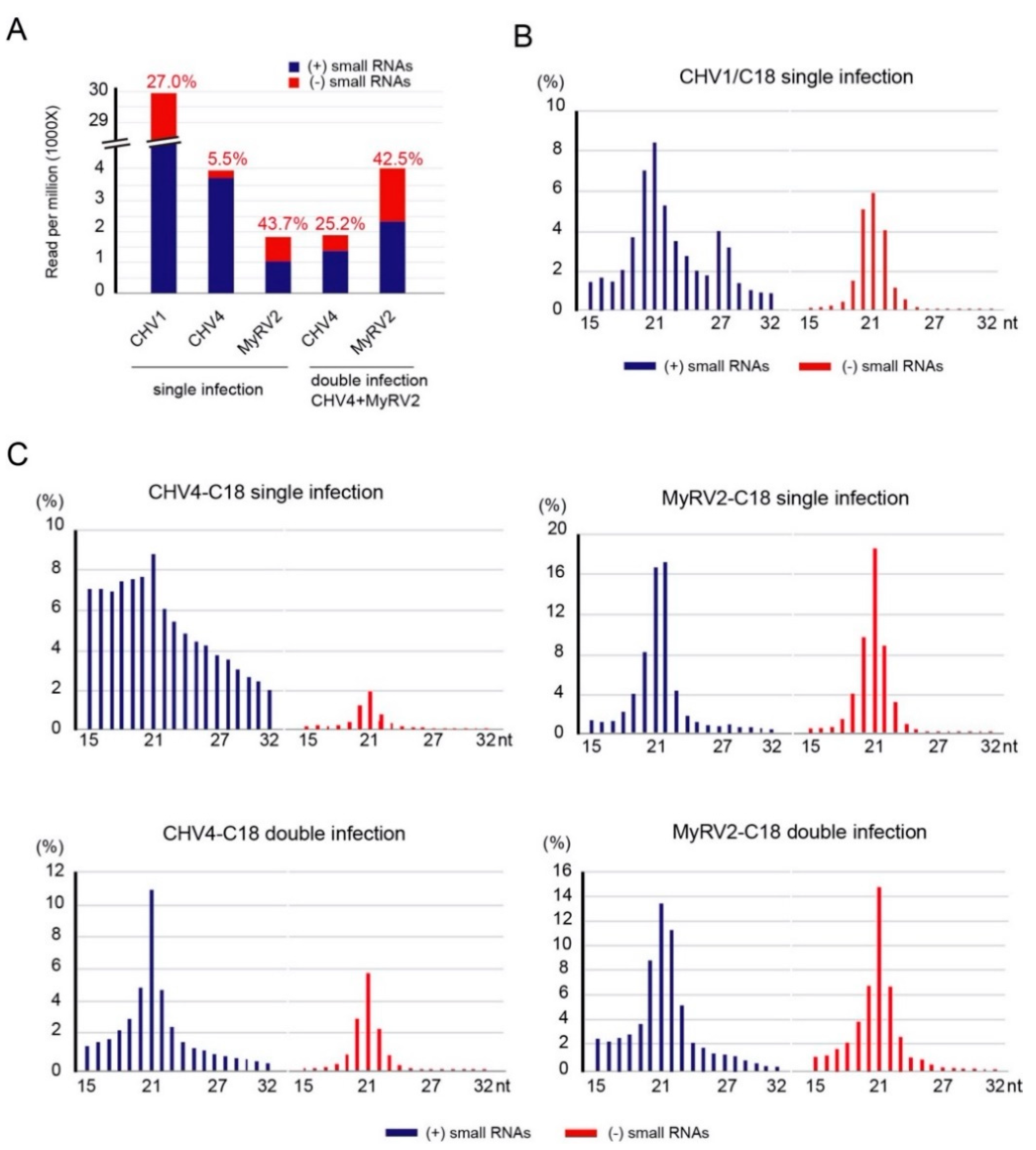
3.2. Co-infection Results in Alterations in Profiles of Virus-Derived Small RNAs
3.3. CHV4 Encodes a Papain-Like Cysteine Protease p24
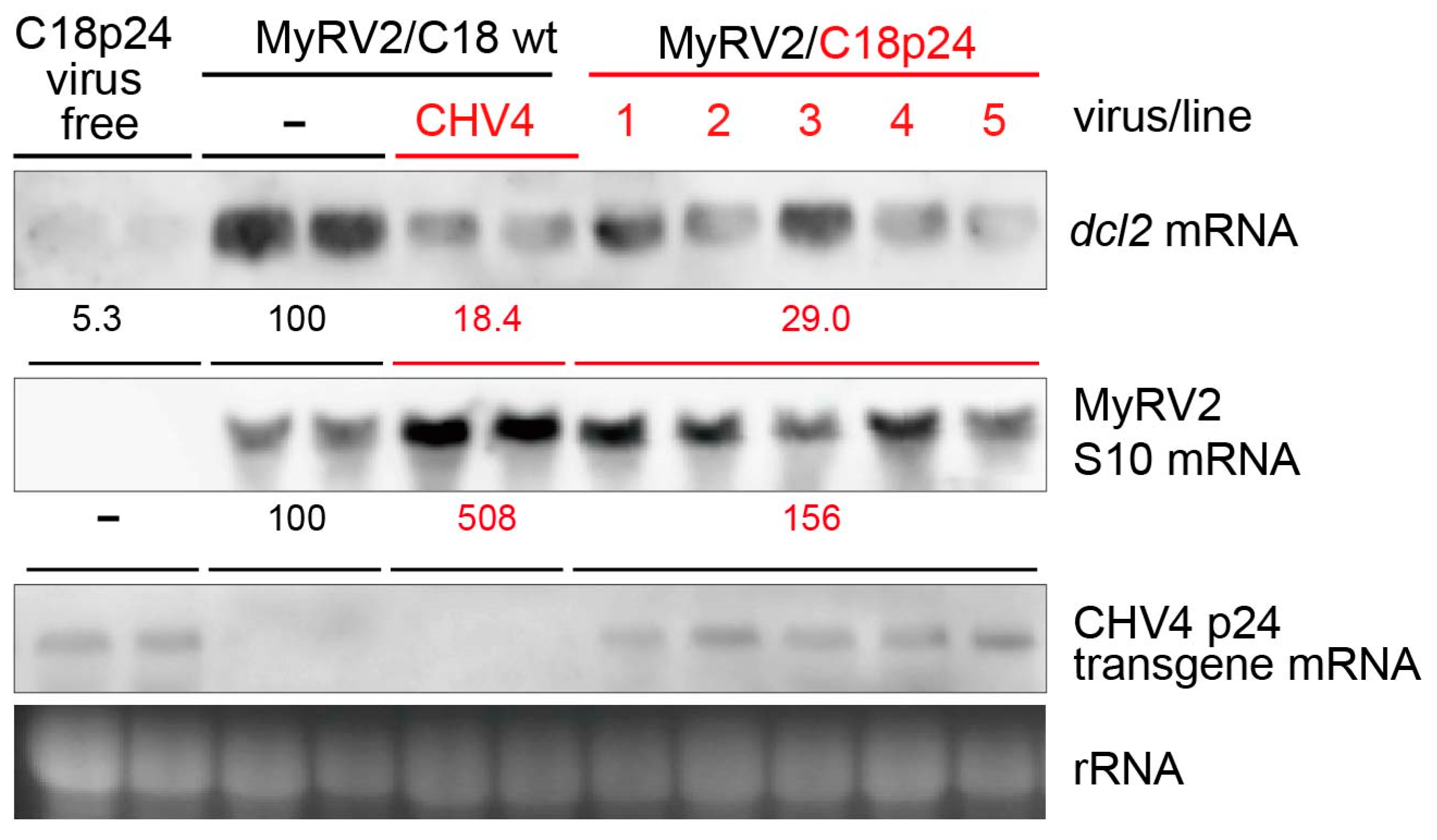
3.4. CHV4-C18 p24 Compromises dcl2 Upregulation and Enhances MyRV2 Accumulation
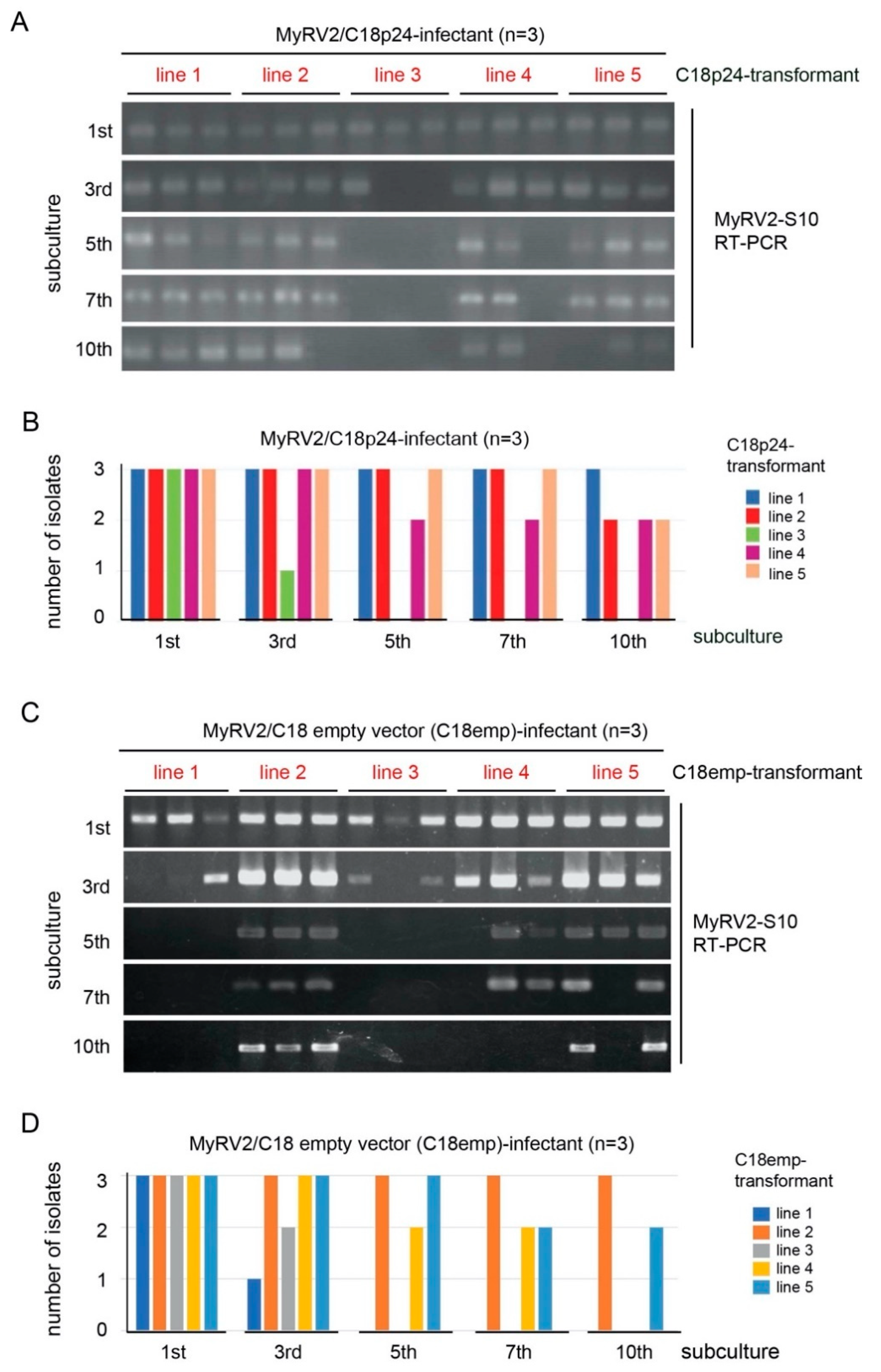
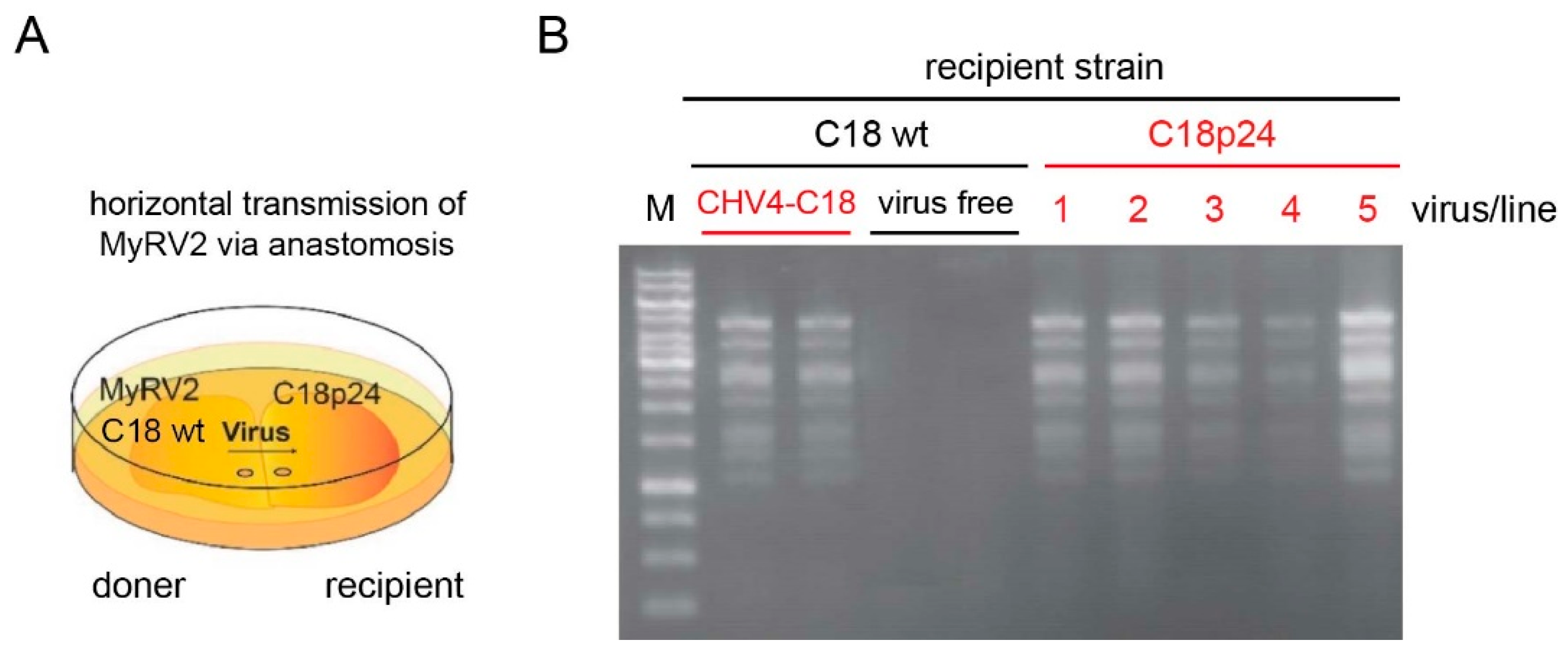
3.5. CHV4-C18 p24 Enhances MyRV2 Stable Infection and Horizontal Transmissibility
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Segers, G.C.; Zhang, X.; Deng, F.; Sun, Q.; Nuss, D.L. Evidence that RNA silencing functions as an antiviral defense mechanism in fungi. Proc. Natl. Acad. Sci. USA 2007, 104, 12902–12906. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Choi, G.H.; Nuss, D.L. A single Argonaute gene is required for induction of RNA silencing antiviral defense and promotes viral RNA recombination. Proc. Natl. Acad. Sci. USA 2009, 106, 17927–17932. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lee, K.M.; Cho, W.K.; Park, J.Y.; Kim, K.H. Differential contribution of RNA interference components in response to distinct Fusarium graminearum virus infections. J. Virol. 2018, 92, e01756-17. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Park, J.Y.; Heo, J.I.; Kim, K.H. The ORF2 protein of Fusarium graminearum virus 1 suppresses the transcription of FgDICER2 and FgAGO1 to limit host antiviral defences. Mol. Plant Pathol. 2020, 21, 230–243. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.; Iritani, A.; Ohkita, S.; Vu, B.V.; Yokoya, K.; Matsubara, A.; Ikeda, K.I.; Suzuki, N.; Nakayashiki, H. A fungal Argonaute interferes with RNA interference. Nucleic Acids Res. 2018, 46, 2495–2508. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; Gilbert, K.B.; Carrington, J.C. Small RNA-based antiviral defense in the phytopathogenic fungus Colletotrichum higginsianum. PLoS Pathog. 2016, 12, e1005640. [Google Scholar] [CrossRef]
- Honda, S.; Eusebio-Cope, A.; Miyashita, S.; Yokoyama, A.; Aulia, A.; Shahi, S.; Kondo, H.; Suzuki, N. Establishment of Neurospora crassa as a model organism for fungal virology. Nat. Commun. 2020, 11, 5627. [Google Scholar] [CrossRef]
- Mochama, P.; Jadhav, P.; Neupane, A.; Marzano, S.Y.L. Mycoviruses as triggers and targets of RNA silencing in white mold fungus Sclerotinia sclerotiorum. Viruses 2018, 10, 214. [Google Scholar] [CrossRef]
- Neupane, A.; Feng, C.; Mochama, P.K.; Saleem, H.; Lee Marzano, S.Y. Roles of argonautes and dicers on Sclerotinia sclerotiorum antiviral RNA silencing. Front. Plant Sci. 2019, 10, 976. [Google Scholar] [CrossRef]
- Lee Marzano, S.Y.; Neupane, A.; Domier, L. Transcriptional and small RNA responses of the white mold fungus Sclerotinia sclerotiorum to infection by a virulence-attenuating hypovirus. Viruses 2018, 10, 713. [Google Scholar] [CrossRef]
- Yaegashi, H.; Shimizu, T.; Ito, T.; Kanematsu, S. Differential Inductions of RNA silencing among encapsidated double-stranded RNA mycoviruses in the white root rot fungus Rosellinia necatrix. J. Virol. 2016, 90, 5677–5692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Segers, G.C.; Sun, Q.; Deng, F.; Nuss, D.L. Characterization of hypovirus-derived small RNAs generated in the chestnut blight fungus by an inducible DCL-2-dependent pathway. J. Virol. 2008, 82, 2613–2619. [Google Scholar] [CrossRef] [PubMed]
- Nuss, D.L. Mycoviruses, RNA silencing, and viral RNA recombination. Adv. Virus Res. 2011, 80, 25–48. [Google Scholar] [PubMed]
- Zhang, D.X.; Spiering, M.J.; Nuss, D.L. Characterizing the roles of Cryphonectria parasitica RNA-dependent RNA polymerase-like genes in antiviral defense, viral recombination and transposon transcript accumulation. PLoS ONE 2014, 9, e108653. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.W. RNA-based antiviral immunity. Nat. Rev. Immunol. 2010, 10, 632–644. [Google Scholar] [CrossRef]
- Voinnet, O. Use, tolerance and avoidance of amplified RNA silencing by plants. Trends Plant Sci. 2008, 13, 317–328. [Google Scholar] [CrossRef]
- Qu, F.; Ye, X.; Morris, T.J. Arabidopsis DRB4, AGO1, AGO7, and RDR6 participate in a DCL4-initiated antiviral RNA silencing pathway negatively regulated by DCL1. Proc. Natl. Acad. Sci. USA 2008, 105, 14732–14737. [Google Scholar] [CrossRef]
- Burgyan, J.; Havelda, Z. Viral suppressors of RNA silencing. Trends Plant Sci. 2011, 16, 265–272. [Google Scholar] [CrossRef]
- Li, F.F.; Wang, A.M. RNA-targeted antiviral immunity: More than just RNA silencing. Trends Microbiol. 2019, 27, 792–805. [Google Scholar] [CrossRef]
- Michaeli, S.; Clavel, M.; Lechner, E.; Viotti, C.; Wu, J.; Dubois, M.; Hacquard, T.; Derrien, B.; Izquierdo, E.; Lecorbeiller, M.; et al. The viral F-box protein P0 induces an ER-derived autophagy degradation pathway for the clearance of membrane-bound AGO1. Proc. Natl. Acad. Sci. USA 2019, 116, 22872–22883. [Google Scholar] [CrossRef]
- Choi, G.H.; Pawlyk, D.M.; Nuss, D.L. The autocatalytic protease p29 encoded by a hypovirulence-associated virus of the chestnut blight fungus resembles the potyvirus-encoded protease HC-Pro. Virology 1991, 183, 747–752. [Google Scholar] [CrossRef]
- Choi, G.H.; Shapira, R.; Nuss, D.L. Cotranslational autoproteolysis involved in gene expression from a double-stranded RNA genetic element associated with hypovirulence of the chestnut blight fungus. Proc. Natl. Acad. Sci. USA 1991, 88, 1167–1171. [Google Scholar] [CrossRef]
- Choi, G.H.; Nuss, D.L. A viral gene confers hypovirulence-associated traits to the chestnut blight fungus. Embo J. 1992, 11, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Craven, M.G.; Pawlyk, D.M.; Choi, G.H.; Nuss, D.L. Papain-like protease p29 as a symptom determinant encoded by a hypovirulence-associated virus of the chestnut blight fungus. J. Virol. 1993, 67, 6513–6521. [Google Scholar] [CrossRef]
- Chiba, S.; Jamal, A.; Suzuki, N. First evidence for internal ribosomal entry sites in diverse fungal virus genomes. MBio 2018, 9, e02350-17. [Google Scholar] [CrossRef] [PubMed]
- Segers, G.C.; van Wezel, R.; Zhang, X.; Hong, Y.; Nuss, D.L. Hypovirus papain-like protease p29 suppresses RNA silencing in the natural fungal host and in a heterologous plant system. Eukaryot. Cell 2006, 5, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Yaegashi, H.; Yoshikawa, N.; Ito, T.; Kanematsu, S. A mycoreovirus suppresses RNA silencing in the white root rot fungus, Rosellinia necatrix. Virology 2013, 444, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Aulia, A.; Andika, I.B.; Kondo, H.; Hillman, B.I.; Suzuki, N. A symptomless hypovirus, CHV4, facilitates stable infection of the chestnut blight fungus by a coinfecting reovirus likely through suppression of antiviral RNA silencing. Virology 2019, 533, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Enebak, S.A.; Hillman, B.I.; Macdonald, W.L. A hypovirulent isolate of Cryphonectria parasitica with multiple, genetically unique dsRNA segments. Mol. Plant-Microbe Interact. 1994, 7, 590–595. [Google Scholar] [CrossRef]
- Enebak, S.A.; Macdonald, W.L.; Hillman, B.I. Effect of dsRNA associated with Isolates of Cryphonectria parasitica from the central Appalachians and their relatedness to other dsRNA from North America and Europe. Phytopathology 1994, 84, 528–534. [Google Scholar] [CrossRef]
- Crouch, J.A.; Dawe, A.; Aerts, A.; Barry, K.; Churchill, A.C.L.; Grimwood, J.; Hillman, B.I.; Milgroom, M.G.; Pangilinan, J.; Smith, M.; et al. Genome sequence of the chestnut blight fungus Cryphonectria parasitica EP155: A fundamental resource for an archetypical invasive plant pathogen. Phytopathology 2020, 110, 1180–1188. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.H.; Nuss, D.L. Hypovirulence of chestnut blight fungus conferred by an infectious viral cDNA. Science 1992, 257, 800–803. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Nuss, D.L. Contribution of protein p40 to hypovirus-mediated modulation of fungal host phenotype and viral RNA accumulation. J. Virol. 2002, 76, 7747–7759. [Google Scholar] [CrossRef] [PubMed]
- Eusebio-Cope, A.; Suzuki, N. Mycoreovirus genome rearrangements associated with RNA silencing deficiency. Nucleic Acids Res. 2015, 43, 3802–3813. [Google Scholar] [CrossRef] [PubMed]
- Andika, I.B.; Jamal, A.; Kondo, H.; Suzuki, N. SAGA complex mediates the transcriptional up-regulation of antiviral RNA silencing. Proc. Natl. Acad. Sci. USA 2017, 114, E3499–E3506. [Google Scholar] [CrossRef]
- Urayama, S.; Katoh, Y.; Fukuhara, T.; Arie, T.; Moriyama, H.; Teraoka, T. Rapid detection of Magnaporthe oryzae chrysovirus 1-A from fungal colonies on agar plates and lesions of rice blast. J. Gen. Plant Pathol. 2015, 81, 97–102. [Google Scholar] [CrossRef]
- Hyodo, K.; Suzuki, N.; Okuno, T. Hijacking a host scaffold protein, RACK1, for replication of a plant RNA virus. New Phytol 2019, 221, 935–945. [Google Scholar] [CrossRef]
- Seguin, J.; Otten, P.; Baerlocher, L.; Farinelli, L.; Pooggin, M.M. MISIS-2: A bioinformatics tool for in-depth analysis of small RNAs and representation of consensus master genome in viral quasispecies. J. Virol. Methods 2016, 233, 37–40. [Google Scholar] [CrossRef]
- Aulia, A.; Tabara, M.; Telengech, P.; Fukuhara, T.; Suzuki, N. Dicer monitoring in a model filamentous fungus host, Cryphonectria parasitica. Curr. Res. Virol. Sci. 2020, 1, 100001. [Google Scholar] [CrossRef]
- Andika, I.B.; Kondo, H.; Suzuki, N. Dicer functions transcriptionally and post-transcriptionally in a multilayer antiviral defense. Proc. Natl. Acad. Sci. USA 2019, 116, 2274–2281. [Google Scholar] [CrossRef]
- Shahi, S.; Eusebio-Cope, A.; Kondo, H.; Hillman, B.I.; Suzuki, N. Investigation of host range of and host defense against a mitochondrially replicating mitovirus. J. Virol. 2019, 93, e01503-18. [Google Scholar] [CrossRef] [PubMed]
- Linder-Basso, D.; Dynek, J.N.; Hillman, B.I. Genome analysis of Cryphonectria hypovirus 4, the most common hypovirus species in North America. Virology 2005, 337, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Ghabrial, S.A.; Kim, K.H.; Pearson, M.; Marzano, S.L.; Yaegashi, H.; Xie, J.; Guo, L.; Kondo, H.; Koloniuk, I.; et al. ICTV vVirus tTaxonomy pProfile: Hypoviridae. J. Gen. Virol. 2018, 99, 615–616. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Chen, B.; Nuss, D.L. Mapping of a hypovirus p29 protease symptom determinant domain with sequence similarity to potyvirus HC-Pro protease. J. Virol. 1999, 73, 9478–9484. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Hillman, B.I. In vitro translational analysis of genomic, defective, and satellite RNAs of Cryphonectria hypovirus 3-GH2. Virology 2001, 281, 117–123. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Suzuki, N.; Maruyama, K.; Moriyama, M.; Nuss, D.L. Hypovirus papain-like protease p29 functions in trans to enhance viral double-stranded RNA accumulation and vertical transmission. J. Virol. 2003, 77, 11697–11707. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yaegashi, H.; Kanematsu, S.; Ito, T. Molecular characterization of a new hypovirus infecting a phytopathogenic fungus, Valsa ceratosperma. Virus Res. 2012, 165, 143–150. [Google Scholar] [CrossRef]
- Wang, S.; Kondo, H.; Liu, L.; Guo, L.; Qiu, D. A novel virus in the family Hypoviridae from the plant pathogenic fungus Fusarium graminarum. Virus Res. 2013, 174, 69–77. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. Introduction: The Clans and Families of Cysteine Peptidases. In Handbook of Proteolytic Enzymes, Rawlings, N.D.; Salvesen, G., Ed.; Elsevier: San Diego, CA, USA, 2013; pp. 1743–1773. [Google Scholar]
- Arjona-Lopez, J.M.; Telengech, P.; Jamal, A.; Hisano, S.; Kondo, H.; Yelin, M.D.; Arjona-Girona, M.I.; Kanematsu, S.; Lopez-Herrera, C.; Suzuki, N. Novel, diverse RNA viruses from Mediterranean isolates of the phytopathogenic fungus, Rosellinia necatrix: Insights into evolutionary biology of fungal viruses. Environ. Microbiol. 2018, 20, 1464–1483. [Google Scholar] [CrossRef]
- Csorba, T.; Kontra, L.; Burgyan, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479C–480C, 85–103. [Google Scholar] [CrossRef]
- Chiba, S.; Suzuki, N. Highly activated RNA silencing via strong induction of dicer by one virus can interfere with the replication of an unrelated virus. Proc. Natl. Acad. Sci. USA 2015, 112, E4911–E4918. [Google Scholar] [CrossRef] [PubMed]
- Peever, T.L.; Liu, Y.C.; Milgroom, M.G. Diversity of hypoviruses and other double-stranded RNAs in Cryphonectria parasitica in North America. Phytopathology 1997, 87, 1026–1033. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Reference or Source |
|---|---|---|
| Fungal | ||
| C18 | Cryphonectria parasitica field strain doubly infected by MyRV2 and CHV4-C18 | [29] |
| C18-VF | Virus-free single conidial isolate of C18 | |
| C18p24 | C18 transformed with the CHV4 p24 coding domain | This study |
| C18/MyRV2 | C18 singly infected by MyRV2 | [28] |
| C18/CHV4 | C18 singly infected by CHV4-C18 | [28] |
| C18/MyRV2+CHV4 | C18 doubly infected by MyRV2 and CHV4 | [28] |
| C18/Δp69 | C18 singly infected by Δp69 | This study |
| C18p24/MyRV2 | C18p24 infected by MyRV2 | This study |
| EP155 | Standard strain of Cryphonectria parasitica (virus-free) | ATCC 38755 |
| C18 Δdcl2 | dcl2 knock-out mutant of C18 (RNA silencing defective, virus-free) | [28] |
| Viral | ||
| MyRV2 | Strain belonging to the species Mycoreovirus 2 within the genus Mycoreovirus | [29] |
| CHV4-C18 | Strain of the species Cryphonectria hypovirus 4 | [28] |
| CHV1-EP713 | Prototype of the family Hypoviridae | [32] |
| CHV1-∆p69 | ORF-A deletion mutant of CHV1-EP713 lacking the p29 and p40 coding domain | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aulia, A.; Hyodo, K.; Hisano, S.; Kondo, H.; Hillman, B.I.; Suzuki, N. Identification of an RNA Silencing Suppressor Encoded by a Symptomless Fungal Hypovirus, Cryphonectria Hypovirus 4. Biology 2021, 10, 100. https://doi.org/10.3390/biology10020100
Aulia A, Hyodo K, Hisano S, Kondo H, Hillman BI, Suzuki N. Identification of an RNA Silencing Suppressor Encoded by a Symptomless Fungal Hypovirus, Cryphonectria Hypovirus 4. Biology. 2021; 10(2):100. https://doi.org/10.3390/biology10020100
Chicago/Turabian StyleAulia, Annisa, Kiwamu Hyodo, Sakae Hisano, Hideki Kondo, Bradley I. Hillman, and Nobuhiro Suzuki. 2021. "Identification of an RNA Silencing Suppressor Encoded by a Symptomless Fungal Hypovirus, Cryphonectria Hypovirus 4" Biology 10, no. 2: 100. https://doi.org/10.3390/biology10020100
APA StyleAulia, A., Hyodo, K., Hisano, S., Kondo, H., Hillman, B. I., & Suzuki, N. (2021). Identification of an RNA Silencing Suppressor Encoded by a Symptomless Fungal Hypovirus, Cryphonectria Hypovirus 4. Biology, 10(2), 100. https://doi.org/10.3390/biology10020100