Simple Summary
The current study reports fungal diversities in the rhizoplane (RP) and rhizosphere (RS) samples of the geographically isolated Korean bellflower (Campanula takesimana) obtained from its original habitats of the eastern coast of the Korean Peninsula for the first time. The identification of specific taxa in each site may provide a better understanding of the interaction between the soil fungi and Korean bellflower.
Abstract
Fungal communities in the rhizoplane (RP) and rhizosphere (RS) of geographically isolated C. takesimana habitats in different environments such as oceanic (Seodo, the Dokdo Islands), coastline (Sadong, Ulleungdo Island), and inland (Taeha, Ulleungdo Island) regions were analyzed by MiSeq sequencing. In total, 1279 operational taxonomic units (OTUs) were obtained and they were further classified into 185 genera belonging to five phyla. The total number of fungal taxa in the RP samples was lower than those in the RS samples in all the sampled locations, providing an indication of the existence of a certain level of the selective pressures from the host plant. The richness of the RP in the Dokdo Islands was higher than that of Ulleungdo Island, but the richness of the RS in the Dokdo Islands was lower than that of Ulleungdo Island. These results suggest evidence for strong effects of a harsh geo-climate on the RP and RS fungal diversities in the Dokdo Islands. Additionally, a total of 82 fungal genera were identified in all three RP samples and 63 genera (77%) were uniquely found in each of the geographical regions and 43 genera (52.4%) showed high dependency on the C. takesimana vegetation. It was found that the genus Mortierella was the most dominant taxon in all the samples. The geo-ecological isolation of the Korean bellflower may have caused unique formation of the RP and RS fungal communities in the natural habitats.
1. Introduction
There are 84 herbaceous plant species belonging to the family Campanulaceae (bellflower family) that are native to the East Asia and the Korean Peninsula [1]. Many of them, such as Platycodon grandiflorus (Jacq.) A.DC., Codonopsis lanceolata (Siebold and Zucc.) Benth. and Hook.f. ex Trautv., Campanula punctata Lam., and Campanula takesimana, are reported to have a variety of pharmacological effects on respiratory or metabolic diseases [2,3,4,5,6]. Campanula punctata is native to the inland terrain of the Korean Peninsula, while C. takesimana, commonly known as the Korean bellflower, is endemic only to Ulleung Island, located in the East Sea of Korea [1]. Recently, a new C. takesimana community was also found on Seodo (the West Island), one of the Dokdo Islands, which has served as home to many wild halotolerant plants [7,8].
Campanula takesimana shows slight morphological differences compared to C. punctata. Its distinctive light pink or purple petals are the key taxonomic feature of C. takesimana which has selectively survived and evolved only on the volcanic islands of Korea [1,9,10]. Based on the internal databases of the Korean National Institute of Biological Resources (NIBR), the Korean bellflower is defined as a halotolerant, herbaceous, and perennial species that only grows naturally in the maritime terrains of the Korean Peninsula [7]. Its colonies are usually found on cliffs and sloping ground adjacent to the sea and a fully grown colony is 30–100 cm tall [11].
Historically, stems, leaves, and flowers of C. takesimana, except for roots, have long been used in Korea as food ingredients and food additives. Furthermore, pharmacologically active compounds such as chlorogenic acid have also been identified in C. takesimana. It is presumed that its unique antioxidant traits have resulted from a gradual adaptation to the harsh marine environment of these oceanic islands since mycorrhizal symbiosis may alter phenolic acid content in plants and the host plants may establish an efficient way to increase stress tolerance under certain conditions through this mechanism [12,13,14].
It is well documented that geographical isolation causes species differentiation in higher life forms and the bellflower species has been extensively studied as a model organism for geological segregation research since it has a high adaptability to divergent environment [12,15]. It has also been reported that the adaptation and flourishing of higher plants in a barren maritime terrain are, to some extent, driven by the symbiotic relationships between soil fungi and plants [16]. However, there have been no clear answers to how fungal diversity differ between geographically isolated host bellflower species and how fungal communities shift in the course of adaption to given environmental circumstances. Thus, more intensive research on microbial communities is needed to define and eventually secure symbiotic microbial resources of C. takesimana in these remote oceanic islands of Korea [8].
Hence, rhizoplane (RP; where ectomycorrhizas strongly interact with the host plants) and rhizosphere (RS; soil away from areas directly affected by plant roots) fungal communities from the three different geographically isolated natural habitats of the Korean bellflowers were analyzed by culture-independent methods in order to identify fungal taxa associated with C. takesimana and to reveal how geographical segregation of the host affects the compositions of the RS and RP fungal communities. Therefore, the current research focuses on soil fungi analysis around the host plants with use of the recent next-generation sequencing technology so that the largest number of microbes present in the soil could be identified. It should be noted that pyrosequencing may not be a sufficient method to identify all inhabiting fungi since the technique may omit spores, fungi present in low amounts, or some fungi with different internal transcribed spacer (ITS) sequence regions [17].
2. Materials and Methods
The Korean bellflower is capable of surviving under both the extremely harsh conditions of volcanic ocean islands and relatively mild conditions of inland terrains of Korea. Therefore, to identify their natural RP and RS fungal components, the target sampling sites where no human activities are allowed or they are tightly controlled were carefully selected in order to correctly represent such environments. However, it is very difficult to find such places in the mainland of the Korean Peninsula. Hence, a virgin terrain on Ulleung Island that represents the inland geography with very limited human access was selected. Accordingly, the RP and RS samples of the Korean bellflower were collected from 3 geographically segregated sites: (1) the Dokdo Islands (Seodo), a rocky oceanic island with volcanic origin [7,18,19]; (2) the coastline of Ulleungdo Island, which has less harsh conditions than those of Seodo [19]; (3) the central inland area of Ulleungdo Island, which is surrounded by mountain ranges composed of virgin primeval forests (Figure 1 and Table 1) [19,20]. All these areas have been designated as National Reserve by the Cultural Heritage Administration of Korea (CHA) for the purpose of conservation [7]. These sampling sites are geographically isolated and segregated from the mainland of Korea and the Japanese archipelago, respectively [7]. It is generally assumed that no ecological or genetic exchanges by higher animals that can travel long distances have occurred. The primary habitats of the Korean bellflower are cliffs and slopes [11] where human access is almost impossible. Although the origin of the Korean bellflower still remains enigmatic, it is generally believed that C. takesimana is a native species to the Dokdo Islands and Ulleungdo Island.
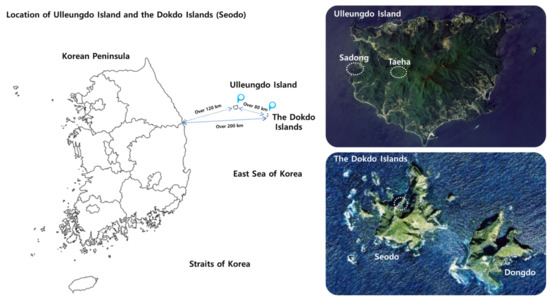
Figure 1.
Geographically segregated natural habitats of the Korean bellflowers. The map images were downloaded from the Cultural Heritage Administration of Korea (CHA) Portal Site (http://www.heritage.go.kr/heri/idx/index.do, accessed on 21 October 2020) and modified for this study.

Table 1.
Sampling site information.
2.1. Sampling of the Host Plant
The RP is the surface of the plant roots where soil microorganisms establish a mutualistic association with the host plant [21], while the RS represents the area of soil around plant roots where highly active interactions among soil, host plant, and microbiome takes place [22]. From each site, about 15–30 fully grown adult plants (15 from Seodo, 30 from Sadong, and 30 from Taeha, respectively) were taken and the topsoil around the roots were sampled in the center of their vegetation for the RP and RS, respectively. The roots of C. takesimana were agitated in sterile distilled water 3 times and the remaining parts were gathered as the RP samples. The RS soil core was taken to the depth of 30 cm with a diameter of 2 mm from each sampling site. Since soil loss due to steep slopes is often observed in Seodo [7], C. takesimana habitats with no signs of soil loss or soil movement were carefully selected for sampling. All the samplings were carried out from 25 to 28 April 2017.
2.2. DNA Extraction and Polymerase Chain Reaction Amplification
All samples were transported to the laboratory at the NIBR for DNA extraction. The soil samples were pooled to obtain a 1 kg composite sample per site. Then, 0.5 g of each sample was used for DNA extraction with a PowerSoil DNA isolation kit (Mo Bio Laboratories, Carlsbad, CA, USA) according to the manufacturer’s protocol. The fungal ITS1 was amplified using the ITS1F (5′-CTT GGT CAT TTA GAG GAA GTA A-3′) and ITS2 (5′-GCT GCG TTC TTC ATC GAT GC-3′) pair of primers [23,24]. The resulting ITS1 amplicons were sequenced at Macrogen (Seoul, Korea) using the paired-end (2 × 300 nt) Illumina MiSeq sequencing system (Illumina, San Diego, CA, USA).
2.3. Sequence Processing and Data Analyses
The paired-end sequences were assembled using PANDAseq software [25]. After assembly, all the sequence data were processed using the Mothur pipeline [26]. For fungal community analysis, the flanking gene fragments were removed from the ITS1 region using ITSx 1.0.9 software [27]. Putative chimeric sequences were detected and removed by the chimera.uchime algorithm available within Mothur [28] in de novo mode. The taxonomic classification was performed using Mothur’s version of the naïve Bayesian classifier, using the UNITE database for fungi [29]. QIIME implementation of UCLUST [30,31] was used to assign the operational taxonomic units (OTUs), defined with a limit threshold of 97% sequence similarity for fungi. All singleton OTUs were removed from the datasets prior to analysis. All samples were standardized by random subsampling using the sub.sample command (http://www.Mothur.org/wiki/Sub.sample, accessed on 7 March 2020) in Mothur. Chao’s richness, Shannon index, Simpson’s diversity index, and rarefaction values [32] were also estimated by using QIIME [33,34].
3. Results and Discussion
3.1. Illumina MiSeq Sequencing Results
Firstly, the OTU values and fungal genera identified in the RS samples were higher than those of the RP samples in all the geographical regions (Table 2). Secondly, the RP/RS ratio values of the OTU and genus in the coastline and inland regions of Ulleungdo Island were lower than those of the Dokdo Islands. Lastly, the numbers of the OTUs and fungal genera detected in the RP of C. takesimana from the Dokdo Islands were the highest among all the RP samples.
Table 2.
Sampling site information comparison of Illumina MiSeq sequencing results.
C. takesimana is resistant to high salt content that constantly flows from the marine environment. However, it is not capable of regulating sodium ions entering their xylem stream unlike other halophytes [35]. On the other hand, the Korean bellflower has certain physiological advantages since they can switch from C3 to C4 or crassulacean acid metabolism photosynthesis under unfavorable conditions such as drought stress [36,37]. Additionally, Campanula species, including C. takesimana, are articulated laticifers which produce multiple secretions and large amounts of organic matter into their root layers. This property may promote mycorrhizal establishment or functionality of salt-resistant heterotrophic fungal species in the RP and/or RS of C. takesimana.
3.2. Variation of the Fungal Phyla Associated with Geographically Isolated Hosts
As shown in Figure 2, the most dominant fungal phylum across all the Dokdo Islands samples was Ascomycota (RP: 43.4% and RS: 27.7%). Ascomycota was also the most dominant phylum in Sadong, the coastline habitat of C. takesimana of Ulleungdo Island (RP: 17.8% and RS: 9.4%). Although there was a tendency of decline in the dominance of Ascomycota with the RS samples, it still remained the most dominant taxon in the Dokdo Islands and Sadong samples. On the other hand, the most abundant phylum in Taeha, the inland area of Ulleungdo Island, was Basidiomycota in the RP (31.1%), while Ascomycota was the second most dominant phylum (16.5%). The second most dominant phylum in the Dokdo Islands was Basidiomycota, whereas the dominance was lowered in the RS to 19.7%. Mucoromycota was the second most dominant phylum in Sadong (RP: 4.8% and RS: 3.1%) and Taeha (RP: 22.4% and RS: 23.8%), respectively.
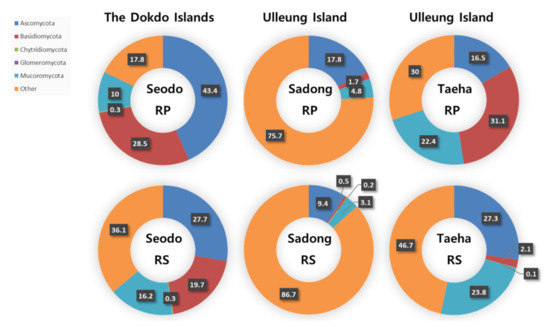
Figure 2.
Fungal phyla distribution in the rhizoplane (RP) and rhizosphere (RS) samples (unit: %).
In summary, Ascomycota was predominantly found in most of these segregated regions. However, the fungal phylum distribution in the RS varied considerably between the three sites since the RS is more susceptible than the RP to environmental influences. Additionally, the dominance of Ascomycota decreased or increased as the distance from the RP increased. In conclusion, no clear patterns were observed in the fungal phyla distribution of the soil samples.
3.3. Variation of Fungal Diverstity
The diversity of fungi in the RP and RS of the host plant was analyzed using the Shannon index, Simpson’s diversity index, and Chao’s richness index. The Shannon index represents the evenness of species distribution and the Simpson’s index indicates the dominance of specific species in a particular environment, respectively [33]. Chao1 richness represents an estimate of the total number of species in the samples [34]. In this study, the RP and RS analysis results from each segregated region did not show any specific patterns associated with the geographical features (Figure 3). Additionally, there were no clear patterns in the dominance of certain fungal taxa in the samples. However, the species abundance showed the distinctive patterns with much higher values in the RS than in the RP in all three geographically segregated regions. The decrease in the fungal taxon abundance in the RP may suggest certain levels of selective pressures from the host [38]. Even though the clear species abundance patterns were shown in all the regions, the highest species abundance was observed in the natural habitat of C. takesimana on the Dokdo Islands. This could be interpreted as more diverse fungal communities being formed under more selective environmental pressure. It is also presumed that a higher fungal diversity in the RP compared to the RS might be the result of seed-associated endophytes, spread with the Korean bellflower seeds.
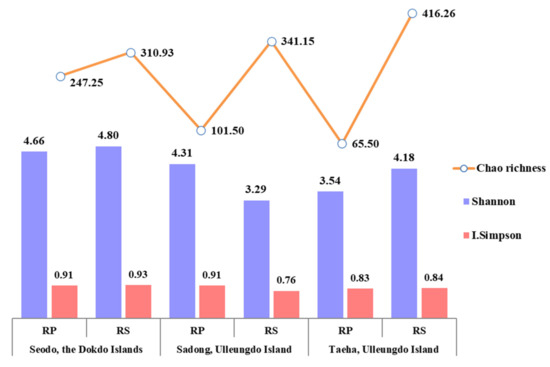
Figure 3.
Fungal diversity variation in the RP and RS samples.
3.4. Variation of Fungal Genera
The main purpose of this study was to identify fungal diversity in the naturally occurring habitat of the Korean bellflower. To achieve this goal, Illumina MiSeq sequencing (Table 3 and Figure 4) and diversity data (Figure 3) analyses were used to determine the fungal taxon distribution of each geographically separated oceanic region (Figure 5). In total, 1279 OTUs at a 97% similarity level were obtained. In Seodo, the Dokdo Islands, 140/341 (RP/RS) OTUs were confirmed, in Sadong, coastline of Ulleungdo Island, 154/389 (RP/RS) OTUs were identified, and in Taeha, inland of Ulleungdo Island, 212/478 (RP/RS) OTUs were found, respectively (Figure 4).
Table 3.
Variation of the major fungal genera.
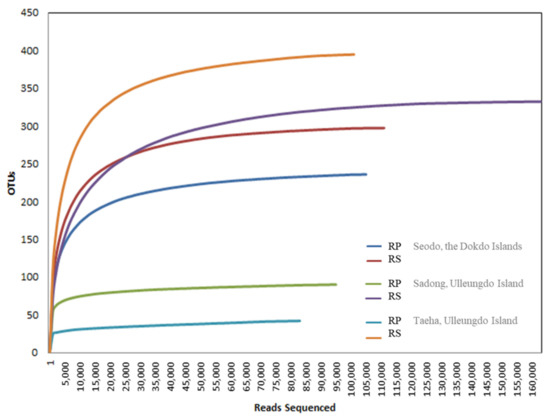
Figure 4.
Illumina MiSeq sequencing rarefaction curve for operational taxonomic units (OTUs) from each site. OTUs were clustered at 3% dissimilarity using CD-HIT. The microbial community in the RP and the RS of Seodo, the Dokdo Islands (OTUs, 140/341; sequencing reads, 84,267/94,745), Sadong, coastline of Ulleungdo Island (OTUs, 154/389; sequencing reads, 95,395/117,939), and Taeha, inland of Ulleungdo Island (OTUs, 212/478; sequencing reads, 86,299/89,864), are shown.
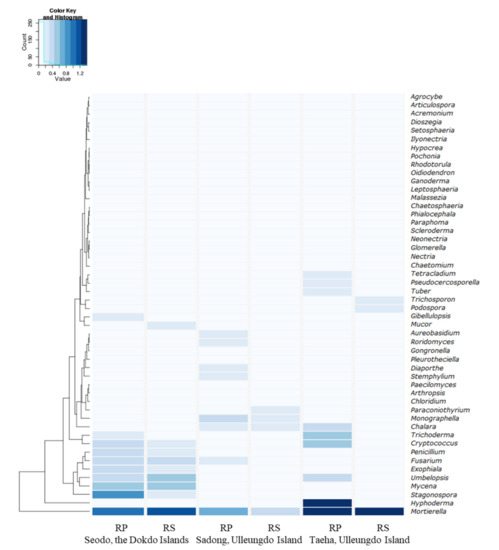
Figure 5.
Heatmap of the 50 major fungal genera, including Mortierella species, distributed around host plants in each geographically segregated environment with a hierarchical clustering.
The most dominant fungal genera in the RPs of the Dokdo Islands samples were Mortierella (7.78%), Fusarium (1.23%), and Penicillium (1.06%). The top three genera identified in the RPs of the Sadong samples were Mortierella (3.54%), Fusarium (0.54%), and Trichoderma (0.38%), while Mortierella (21.26%) and Trichoderma (2.21%) were predominant in the RPs of the Taeha samples (Table 3).
The genus Mortierella is known as a saprophyte that derives nutrients from decaying organic matters such as feces, trees, and insects [39,40]. In addition, Penicillium and Trichoderma are saprophytes [41], but they also play imperative roles in plant growth and nutrient absorption as well as resistance to extreme conditions, pathogens, and grazing by herbivores [42,43,44].
Regarding the difference between the RP and RS in the genus dominance, the most dominant genus, Mortierella, showed decreased percentages in the RPs of the Seodo and Taeha samples compared to the RSs, but the opposite result was observed in the Sadong samples. Therefore, there were no specific patterns in the dominance of Mortierella and other fungal genera including Fusarium, Penicillium, and Trichoderma (Table 3 and Figure 5).
3.5. Host Dependency and Selective Pressure
The present study also aimed to compare and analyze the RP and RS fungal communities in the contrasting habitats of the Korean bellflower. It was confirmed that the C. takesimana RP exerted some selective pressures on their fungal clusters. Therefore, a comparative analysis of unique fungal taxon composition in the RP of C. takesimana from the three geographically separated regions may provide a better understanding of this phenomenon. A total of 82 fungal genera was identified in the three RP samples. There were 58 genera found in Seodo, 34 genera in Sadong, and 15 genera in Taeha, respectively. Among the 82 genera, 63 genera (77%) were uniquely found in each of the geographical region (43 genera in Seodo, 18 genera in Sadong, and 2 genera in Taeha, respectively) (Figure 6).
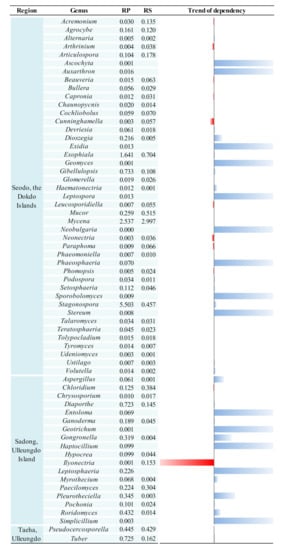
Figure 6.
Unique fungal genera distributed in the RP and RS in each geographically isolated region and their host dependency. Blue bars indicate host dependency and red bars indicate increased dominance ratio as physical distance is away from the RP towards to the RS. Each number indicates the relative percentage of each genus and the trend of dependency of each genus was calculated by the following equation: (RS − RP)/RP.
Meanwhile, the host dependency of each of the uniquely existing RP fungal genera could be identified by comparing the dominance variation between the RS and RP. If a particular taxon was detected only in the RP, but not in the RS, it was assumed that the taxon is dependent on the plant. Among the total unique genera, 43 genera (52.4%) showed a dependence on C. takesimana and it was presumed that a close interaction was established between these fungal taxa and the host plant. There were 27 RP-specific genera with high dependency in Seodo, 14 genera in Sadong, and 2 genera in Taeha, respectively. The fungal genera showing dependencies to the host plant’s RP are visualized in Figure 6. Especially, Ascochyta, Auxarthron, Exidia, Geomyces, Leptospora, Neobulgaria, Phaeosphaeria, Sporobolomyces, and Stereum were found only in the RP of Seodo, but they were not observed in the RS of Seodo. In addition, Entoloma, Geotrichum, Haptocillium, Leptosphaeria, and Simplicillium were found only in the RP of Sadong, but they were not identified in the RS of Sadong. These genera exhibited strong dependencies on the host plant. Therefore, it could be concluded that the ecological isolation of the Korean bellflower may have caused the formation of the unique fungal taxon that have strong interactions with C. takesimana.
The distribution of RS fungi was analyzed and, overall, 172 RS fungal genera were identified—71 genera in Seodo, 86 genera in Taeha, and 101 genera in Sadong were observed, respectively. The uniquely found genera in each site (30 genera from Seodo, 31 genera from Sadong, and 44 genera from Taeha, respectively) comprised 61% of all the identified RS fungal genera. The RS fungal genera that were not identified in the RP (13 genera in Seodo, 27 genera in Sadong, and 41 genera in Taeha, respectively) appear to be independent of the host plant.
In particular, the genus Ilyonectria (Figure 6), which was reported as a plant pathogen or an opportunistic pathogen [45,46,47,48], was significantly suppressed in the RP and its dominance increased steeply in the RS of Sadong (Figure 6). It was also reported that this genus participates in a combination of endophytic and saprotrophic activities of plants [49]. However, no conclusion can be drawn whether it was inhibited by the presence of C. takesimana’s roots since this genus was only identified in Sadong on Ulleungdo Island in this study.
The comparative analysis results showed that the genus Stemphylium was identified only in the RPs of Seodo and Sadong. It has been reported that Stemphylium exhibits pathogenicity to the family Campanulaceae, including Platycodon grandiflorus [50], which is a widely used alternative medicine in various regions, especially in Asia. The members of this genus include S. alfalfae, S. bolickii, S. cannabinum, S. globuliferum, S. lycopersici, S. sarciniforme, S. solani, and S. vesicarium [51] and they are not considered as plant symbionts. Hyphoderma was found in the RPs of Sadong and Taeha on Ulleungdo Island, but this genus was not present in the RSs of the same sampling sites. No conclusion can be made from this result as Hyphoderma is a ubiquitous corticioid homobasidiomycetes genus that have been commonly reported worldwide. The fungal genera that existed only in the RP of Seodo include Ascochyta, Auxarthron, Exidia, Geomyces, Leptospora, Neobulgaria, and Stereum. Even though their dominance was below 0.3% in the RP (Table S1), it seems that these fungal genera within the range of the selective pressures from C. takesimana vegetation are more competitive in this environment than other fungi. Some of the fungal genera only found in the RP are generally known for their pathogenicity to a wide range of plants. Information on the interdependency trends between the host species and microbes was obtained by comparing fungal genera in the RP and RS in this study. However, such dependencies cannot always be interpreted as symbiotic relationships [52] because certain pathogens usually have strong host (species) dependencies according to traditional pathobiology or plant pathophysiology [53,54]. Ascochyta is a genus of ascomycete fungi that contains several species that are pathogenic to crop species [55], but the interactions between Auxarthron species and plants are still unknown. Exidia is a genus of endogenous fungi that produce antioxidants [56] and no pathogenicity has yet been reported for this taxon. Geomyces is a genus of filamentous fungi belonging to the Myxotrichaceae family. Members of this genus are widely distributed, especially in northern temperate regions [57], and they are also known as psychrotolerant fungi associated with Arctic permafrost soils [58]. While the genus Neobulgaria usually grows on fallen dead beech trees, the genus Stereum is found on all kinds of deadwood, hardwood, or dead leaves [59] and they are sometimes observed on living tree leaves. Leptospora is a genus of fungi within the class Dothideomycetes and it is known as a nonpathogenic fungus [60]. As mentioned earlier, soil fungal diversity in the RP are primarily determined by the selective pressure derived from host plant species. Hence, both higher richness and species diversity may play vital roles in enhancing the stability of the corresponding soil fungal communities.
4. Conclusions
In this study, four fungal genera including Fusarium, Mortierella, Penicillium, and Trichoderma were commonly present in all the geographically segregated regions and they were distributed both in the RS and RP samples (Table 3). The genus Mortierella dominated the fungal communities in both the RP and RS and it showed the highest dominance among all the genera identified in this research. Furthermore, its dominance increased as the distance increased from the RPs towards the RSs in Seodo and Taeha. However, the opposite result was observed in the Sadong samples. The cause of this decrease in the fungal composition in the inland region of Ulleungdo Island is not clear. It was assumed that a kind of competition with other fungal genera rather than interaction with the host plant may have caused this since the RS is more susceptible to a variety of biotic and abiotic stresses. The genus Mortierella is usually nonpathogenic for plants, animals, and humans [61]. M. wolfii is the only Mortierella species that is pathogenic to humans and other animals and it is commonly isolated from soil, rotten silage, and similar substrates, often causing bovine abortion, pneumonia, and systemic mycosis [62]. However, given the remote conditions of the major habitats of the Korean bellflower, the human activity-induced pathogenicity of Mortierella with the host species is neglectable. Several Basidiomycota fungal genera have been reported to help their host plant species by taking up soil minerals from land via forming mycorrhizae at the roots of vascular plants under unfavorable conditions. Mortierella, in turn, obtains materials for survival synthesized by its host plants through photosynthesis [63,64]. Several Mortierella species have shown that they are able to confer enhanced tolerance to rot disease in Crocus corms [65] and increased resistance to salt stress in halophytic plants [66], respectively. It has also been reported that the Mortierella species promote their host plants’ growth by producing phyto-hormones [61]. Mortierella also exists as an ectrophic mycorrhiza in the roots of the bellflower, having a positive effect on the growth of the host species. The comparative analysis results indicated that there was no specific pattern of the variation of dominance depending on the distance from the host plant’s RP, but our study was able to show the distribution of Mortierella as the most dominant fungal genus present in the RP and RS of C. takesimana. It would be interesting for further research to investigate the ecological roles of Mortierella and other dominant fungi with respect to the plant’s growth and function and its adaptation to the harsh environments by means of transcriptomic analysis.
To our knowledge, this is the first report to investigate the fungal diversities in RP and RS samples of C. takesimana obtained from its natural habitats. The current results obtained by the analysis of both host dependency and selective pressure may enhance our understanding of C. takesimana’s ability to withstand the harsh environments of the Dokdo Islands and Ulleung Island since the identification of specific taxa in each site can provide a brief overview of the presence of the dominant RP and RS fungi in C. takesimana and this information would help us to narrow down future research on how these fungi function with the host plant under these extreme conditions. Lastly, it was reported that microbial communities may vary across seasons more than in response to long-term climate changes [67]. Hence, it would also be interesting to investigate the fungal diversities in the RP and RS of the Korean bellflower on a regular basis at specific times of the year.
Supplementary Materials
The following are available online at https://www.mdpi.com/2079-7737/10/2/138/s1, Table S1: Fungal genus distribution, Table S2: Soil analysis results.
Author Contributions
Y.-H.Y., J.W.H., and B.-H.L. contributed to the conception and design of the study; J.M.P. organized the database, performed the statistical analysis, and wrote the first draft of the manuscript; J.M.P. wrote sections of the manuscript. J.M.P., B.K., and Y.-C.C. carried out Illumina MiSeq sequencing, data submission, and the assembly and annotations. J.M.P. and B.-H.L. generated and analyzed the sequencing data. J.M.P. and B.K. carried out the genomic analysis and illustrations. J.W.H. critically reviewed and summarized all the literature and the manuscript. Y.-H.Y. supervised the project and critically evaluated the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the National Institute of Biological Resources (NIBR), funded by the Ministry of Environment (MOE) of the Republic of Korea (NIBR202030102, NIBR202131102).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the raw sequences obtained from this study were deposited at the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SAR) under the project accession number PRJNA616069.
Acknowledgments
The authors would like to thank the Ulleung County Office for their support in the field survey.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Wiersema, J.H. GRIN Taxonomy. US National Plant Germplasm System. Checklist Dataset. 2019. Available online: https://doi.org/10.15468/ao14pp (accessed on 4 April 2020).
- Brandt, K.; Kondo, T.; Aoki, H.; Goto, T. Structure and biosynthesis of anthocyanins in flowers of Campanula. Phytochemistry 1993, 33, 209–212. [Google Scholar] [CrossRef]
- Stuchlíková, L.; Jirásko, R.; Skálová, L.; Pavlík, F.; Szotáková, B.; Holčapek, M.; Vaněk, T.; Podlipná, R. Metabolic pathways of benzimidazole anthelmintics in harebell (Campanula rotundifolia). Chemosphere 2016, 157, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Folquitto, D.G.; Swiech, J.N.D.; Pereira, C.B.; Bobek, V.B.; Halila Possagno, G.C.; Farago, P.V.; Miguel, M.D.; Duarte, J.L.; Miguel, O.G. Biological activity, phytochemistry and traditional uses of genus Lobelia (Campanulaceae): A systematic review. Fitoterapia 2019, 134, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.Y.; Bo, A.; Yang, M.; Xu, J.F.; Jiang, L.L.; Zhou, B.C.; Li, M.H. The pharmacological effects and health benefits of Platycodon grandiflorus-A medicine food homology species. Foods 2020, 9, 142. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.; Wang, P.; Wang, X.; Zhou, X.; Hu, X.; Chen, F. Dietary Platycodon grandiflorus attenuates hepatic insulin resistance and oxidative stress in high-fat-diet induced non-alcoholic fatty liver disease. Nutrients 2020, 12, 480. [Google Scholar] [CrossRef] [PubMed]
- Cultural Heritage Administration (CHA) of Korea. Natural Heritage of Korea, Dokdo. Available online: http://116.67.83.213/NEW_PDF/EM036763_01.pdf (accessed on 17 October 2020).
- Park, J.M.; Hong, J.W.; Son, J.S.; Hwang, Y.J.; Cho, H.M.; You, Y.H.; Ghim, S.Y. A strategy for securing unique microbial resources–focusing on Dokdo islands-derived microbial resources. Isr. J. Ecol. Evol. 2018, 64, 1–15. [Google Scholar] [CrossRef]
- Park, K.R.; Jung, H.J. Isozyme and morphological variation in Campanula punctata and C. takesimana (Campanulaceae). Korean J. Plant Taxon. 2000, 30, 1–16. [Google Scholar] [CrossRef]
- Shulkina, T.V.; Gaskin, J.F.; Eddie, W.M.M. Morphological studies toward an improved classification of Campanulaceae s. str. Ann. MO. Bot. Gard. 2003, 90, 576–591. [Google Scholar] [CrossRef]
- Ahn, Y.H.; Jo, D.K.; Kim, K.S.; Lee, J.H.; Shin, C.H.; Lee, S.J.; Kang, K.H. Ecological characteristics of native Campanula takesimana habitats in Ulleung-do Island. J. Plant Environ. 2005, 1, 19–24. [Google Scholar]
- Thorpe, R.S.; Surget-Groba, Y.; Johansson, H. Genetic tests for ecological and allopatric speciation in anoles on an island archipelago. PLoS Genet. 2010, 6, e1000929. [Google Scholar] [CrossRef] [PubMed]
- Aseel, D.G.; Rashad, Y.M.; Hammad, S.M. Arbuscular mycorrhizal fungi trigger transcriptional expression of flavonoid and chlorogenic acid biosynthetic pathways genes in tomato against Tomato Mosaic Virus. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Haruma, T.; Yamaji, K.; Ogawa, K.; Masuya, H.; Sekine, Y.; Kozai, N. Root-endophytic Chaetomium cupreum chemically enhances aluminium tolerance in Miscanthus sinensis via increasing the aluminium detoxicants, chlorogenic acid and oosporein. PLoS ONE 2019, 14, e0212644. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, C.B.; Ma, X.G.; Liang, Q.L.; He, X.J. Phylogeographic analysis of a temperate-deciduous forest restricted plant (Bupleurum longiradiatum Turcz.) reveals two refuge areas in China with subsequent refugial isolation promoting speciation. Mol. Phylogenet. Evol. 2013, 68, 628–643. [Google Scholar] [CrossRef] [PubMed]
- Baez-Rogelio, A.; Morales-García, Y.E.; Quintero-Hernández, V.; Muñoz-Rojas, J. Next generation of microbial inoculants for agriculture and bioremediation. Microb. Biotechnol. 2017, 10, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Perlińska-Lenart, U.; Piłsyk, S.; Gryz, E.; Turło, J.; Hilszczańska, D.; Kruszewska, J.S. Identification of bacteria and fungi inhabiting fruiting bodies of Burgundy truffle (Tuber aestivum Vittad.). Arch. Microbiol. 2020, 202, 2727–2738. [Google Scholar] [CrossRef] [PubMed]
- National Geographic Information Institute (NGII) of Korea. Geography of Dokdo. 2015. Available online: https://map.ngii.go.kr/ms/pblictn/dokdoGrphHist.do (accessed on 17 October 2020).
- You, Y.H.; Park, J.M.; Park, J.H.; Kim, J.G. Specific rhizobacterial resources: Characterization and comparative analysis from contrasting coastal environments of Korea. J. Basic Microbiol. 2016, 56, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.S.; Watts, P. The Plant Geography of Korea with an Emphasis on the Alpine Zones; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Vega, N.W.O. A review on beneficial effects of rhizosphere bacterial on soil nutrient availability and plant nutrient uptake. Rev. Environ. Contam. Toxicol. 2007, 223, 33–52. [Google Scholar]
- Flores-Vargas, R.D.; O’Hara, G.W. Isolation and characterization of rhizosphere bacterial with potential for biological control of weed in vineyards. J. Appl. Microbiol. 2006, 100, 946–954. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sánchez-García, M.; Ebersberger, I.; de Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Hoiland, K.; Kjoller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi-recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Huttley, G.A. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Heck, K.L.; van Belle, G.; Simberloff, D. Explicit calculation of the rarefaction diversity measurement and the determination of sufficient sample size. Ecology 1975, 56, 1459–1461. [Google Scholar] [CrossRef]
- Lambshead, P.J.D.; Platt, H.M.; Shaw, K.M. The detection of differences among assemblages of marine benthic species based on an assessment of dominance and diversity. J. Nat. Hist. 1983, 17, 859–874. [Google Scholar] [CrossRef]
- Chao, A.; Shen, T.J. Program SPADE (Species Prediction and Diversity Estimation) Program and User’s Guide; National Tsing Hua University: Hsinchu, Taiwan, 2010; Available online: http://chao.stat.nthu.edu.tw/wordpress/ (accessed on 17 October 2020).
- Chapman, V.J. Salt Marshes and Salt Deserts of the World. In Ecology of Halophytes; Reimold, R.J., Queen, W.H., Eds.; Academic Press: New York, NY, USA, 1974; pp. 3–19. [Google Scholar]
- Von Caemmerer, S.; Edwards, G.E.; Koteyeva, N.; Cousins, A.B. Single-cell C4 photosynthesis in aquatic and terrestrial plants: A gas exchange perspective. Aquat. Bot. 2014, 118, 71–80. [Google Scholar] [CrossRef]
- Sage, R.F. A portrait of the C4 photosynthetic family on the 50th anniversary of its discovery: Species number, evolutionary lineages, and Hall of Fame. J. Exp. Bot. 2017, 68, e11–e28. [Google Scholar] [CrossRef]
- Barton, L.L.; Northup, D.E. Microbial Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Deacon, J.W. Fungal Biology, 4th ed.; Wiley-Blackwell: Malden, MA, USA, 2005. [Google Scholar]
- Webster, J.; Weber, R.W.S. Introduction to Fungi; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Min, Y.J.; Park, M.S.; Fong, J.J.; Quan, Y.; Jung, S.; Lim, Y.W. Diversity and saline resistance of endophytic fungi associated with Pinus thunbergii in coastal shelterbelts of Korea. J. Microbiol. Biotechnol. 2014, 24, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F.; Kubota, M.; Koyama, H.; Hyakumachi, M. The plant growth promoting fungus Penicillium simplicissimum GP17-2 induces resistance in Arabidopsis thaliana by activation of multiple defense signals. Plant Cell Physiol. 2007, 48, 1724–1736. [Google Scholar] [CrossRef]
- Khan, S.A.; Hamayun, M.; Yoon, H.; Kim, H.Y.; Suh, S.J.; Hwang, S.K.; Kim, J.M.; Lee, I.J.; Choo, Y.S.; Yoon, U.H.; et al. Plant growth promotion and Penicillium citrinum. BMC Microbiol. 2008, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Del Pilar Martínez-Diz, M.; Andrés-Sodupe, M.; Berbegal, M.; Bujanda, R.; Díaz-Losada, E.; Gramaje, D. Droplet digital PCR technology for detection of Ilyonectria liriodendri from grapevine environmental samples. Plant Dis. 2020, 104, 1144–1150. [Google Scholar] [CrossRef]
- Farh, M.E.; Kim, Y.J.; Singh, P.; Yang, D.C. Cross interaction between Ilyonectria mors-panacis isolates infecting Korean ginseng and ginseng saponins in correlation with their pathogenicity. Phytopathology 2017, 107, 561–569. [Google Scholar] [CrossRef]
- Farh, M.E.; Kim, Y.J.; Abbai, R.; Singh, P.; Jung, K.H.; Kim, Y.J.; Yang, D.C. Pathogenesis strategies and regulation of ginsenosides by two species of Ilyonectria in Panax ginseng: Power of speciation. J. Ginseng Res. 2020, 44, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Chen, M.; Ma, Y.; Du, Z.; Yuan, N.; Li, Y.; Xiao, J.; Zhang, Y. Whole-genome and time-course dual RNA-Seq analyses reveal chronic pathogenicity-related gene dynamics in the ginseng rusty root rot pathogen Ilyonectria robusta. Sci. Rep. 2020, 10, 1586. [Google Scholar] [CrossRef]
- Liao, H.L.; Bonito, G.; Rojas, J.A.; Hameed, K.; Wu, S.; Schadt, C.W.; Labbé, J.; Tuskan, G.A.; Martin, F.; Grigoriev, I.V.; et al. Fungal endophytes of Populus trichocarpa alter host phenotype, gene expression, and rhizobiome composition. Mol. Plant Microbe Interact. 2019, 32, 853–864. [Google Scholar] [CrossRef]
- Deng, J.X.; Paul, N.C.; Li, M.J.; Cho, H.S.; Lee, H.B.; Yu, S.H. Stemphylium platycodontis sp. nov., isolated from Platycodon grandiflorus in Korea. Mycol. Prog. 2014, 13, 477–482. [Google Scholar] [CrossRef]
- Olsen, K.J.K.; Rossman, A.; Andersen, B. Metabolite production by species of Stemphylium. Fungal Biol. 2018, 122, 172–181. [Google Scholar] [CrossRef]
- Johnson, N.C.; Graham, J.H.; Smith, F.A. Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytol. 1997, 135, 575–585. [Google Scholar] [CrossRef]
- Bäumler, A.; Fang, F.C. Host specificity of bacterial pathogens. Cold Spring Harb. Perspect. Med. 2013, 3, a010041. [Google Scholar]
- Morris, C.E.; Moury, B. Revisiting the concept of host range of plant pathogens. Annu. Rev. Phytopathol. 2019, 57, 63–90. [Google Scholar] [CrossRef]
- Kutcher, H.R.; Johnston, A.M.; Bailey, K.L.; Malhi, S.S. Managing crop losses from plant diseases with foliar fungicides, rotation and tillage on a Black Chernozem in Saskatchewan, Canada. Field Crops Res. 2011, 124, 205–212. [Google Scholar] [CrossRef]
- Atiphasaworn, P.; Monggoot, S.; Gentekaki, E.; Brooks, S.; Pripdeevech, P. Antibacterial and antioxidant constituents of extracts of endophytic fungi isolated from Ocimum basilicum var. thyrsiflora leaves. Curr. Microbiol. 2017, 74, 1185–1193. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; David, J.C.; Stalpers, J.A. Ainsworth and Bisby’s Dictionary of the Fungi, 9th ed.; CABI Publishing: Hague, The Netherlands, 2001. [Google Scholar]
- Blehert, D.S.; Hicks, A.C.; Behr, M.; Meteyer, C.U.; Berlowski-Zier, B.M.; Buckles, E.L.; Coleman, J.T.H.; Darling, S.R.; Gargas, A.; Niver, R.; et al. Bat white-nose syndrome: An emerging fungal pathogen? Science 2009, 323, 227. [Google Scholar] [CrossRef] [PubMed]
- Kryvomaz, T.I.; Michaud, A.; Minter, D.W. IMI Descriptions of Fungi and Bacteria; CABI Publication: Haue, The Netherlands, 2019. [Google Scholar]
- Li, C.J.; Gao, J.H.; Nan, Z.B. Interactions of Neotyphodium gansuense, Achnatherum inebrians, and plant-pathogenic fungi. Mycol. Res. 2007, 111, 1220–1227. [Google Scholar] [CrossRef]
- Ozimek, E.; Jaroszuk-Ściseł, J.; Bohacz, J.; Korniłłowicz-Kowalska, T.; Tyśkiewicz, R.; Słomka, A.; Nowak, A.; Hanaka, A. Synthesis of indoleacetic acid, gibberellic acid and ACC-deaminase by Mortierella strains promote winter wheat seedlings growth under different conditions. Int. J. Mol. Sci. 2018, 19, 3218. [Google Scholar] [CrossRef]
- Davies, J.L.; Wobeser, G.A. Systemic infection with Mortierella wolfii following abortion in a cow. Can. Vet. J. 2010, 51, 1391. [Google Scholar]
- Zhang, H.; Wu, X.; Li, G.; Qin, P. Interactions between arbuscular mycorrhizal fungi and phosphate-solubilizing fungus (Mortierella sp.) and their effects on Kostelelzkya virginica growth and enzyme activities of rhizosphere and bulk soils at different salinities. Biol. Fertil. Soils 2011, 47, 543. [Google Scholar] [CrossRef]
- Toju, H.; Sato, H. Root-associated fungi shared between arbuscular mycorrhizal and ectomycorrhizal conifers in a temperate forest. Front. Microbiol. 2018, 9, 433. [Google Scholar] [CrossRef] [PubMed]
- Wani, Z.A.; Kumar, A.; Sultan, P.; Bindu, K.; Riyaz-Ul-Hassan, S.; Ashraf, N. Mortierella alpina CS10E4, an oleaginous fungal endophyte of Crocus sativus L. enhances apocarotenoid biosynthesis and stress tolerance in the host plant. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.S.; Qin, F.F.; Qin, P.; Pan, S.M. Evidence that arbuscular mycorrhizal and phosphate-solubilizing fungi alleviate NaCl stress in the halophyte Kosteletzkya virginica: Nutrient uptake and ion distribution within root tissues. Mycorrhiza 2014, 24, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Yuste, J.C.; Fernandez-Gonzalez, A.J.; Fernandez-Lopez, M.; Ogaya, R.; Penuelas, J.; Sardans, J.; Lloret, F. Strong functional stability of soil microbial communities under semiarid Mediterranean conditions and subjected to long-term shifts in baseline precipitation. Soil Biol. Biochem. 2014, 69, 223–233. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).