Soil Oxidation-Reduction in Wetlands and Its Impact on Plant Functioning

Abstract
:1. Introduction
2. Soil Oxidation-Reduction
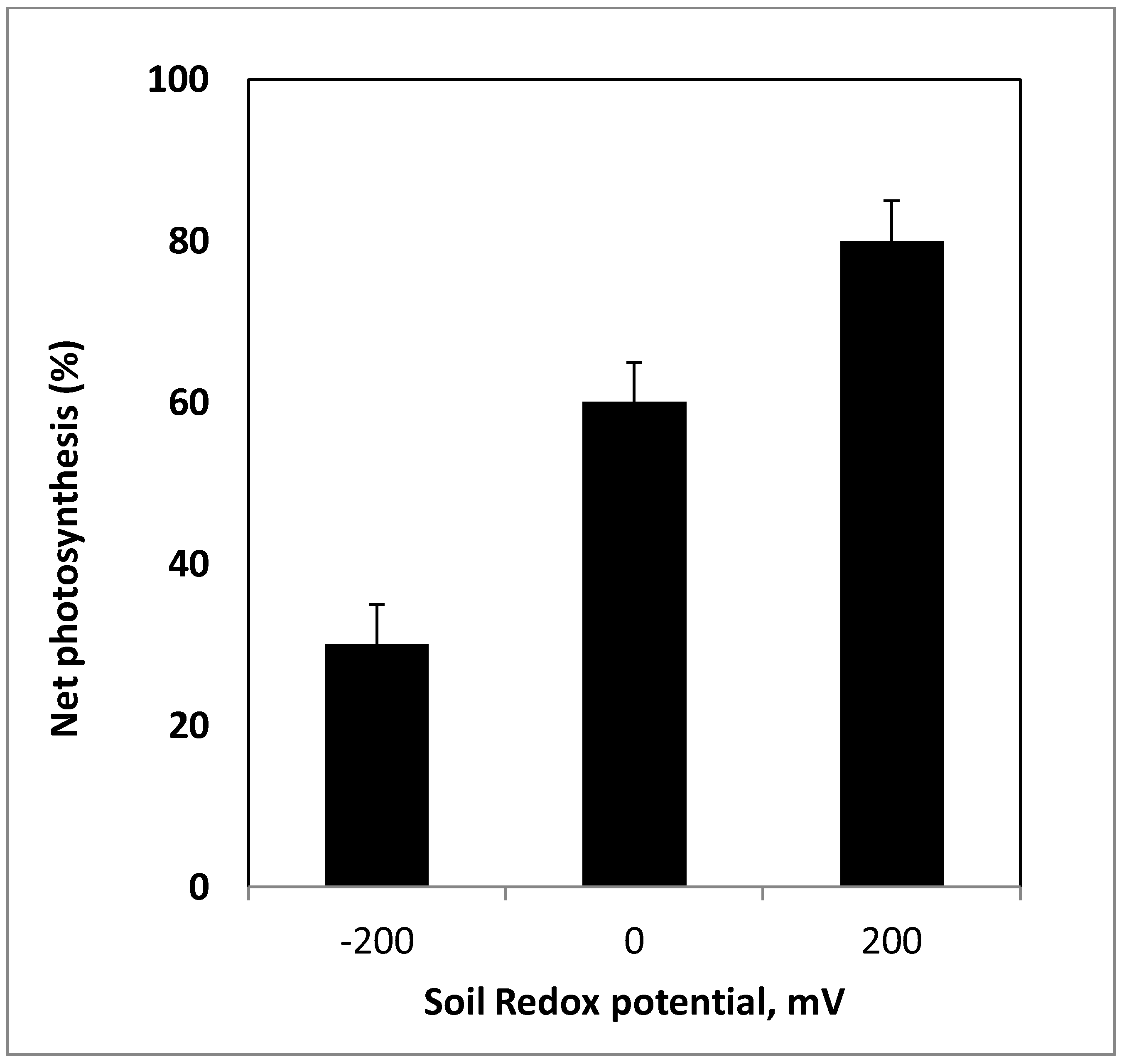
3. Quantifying Soil Redox Potential Conditions
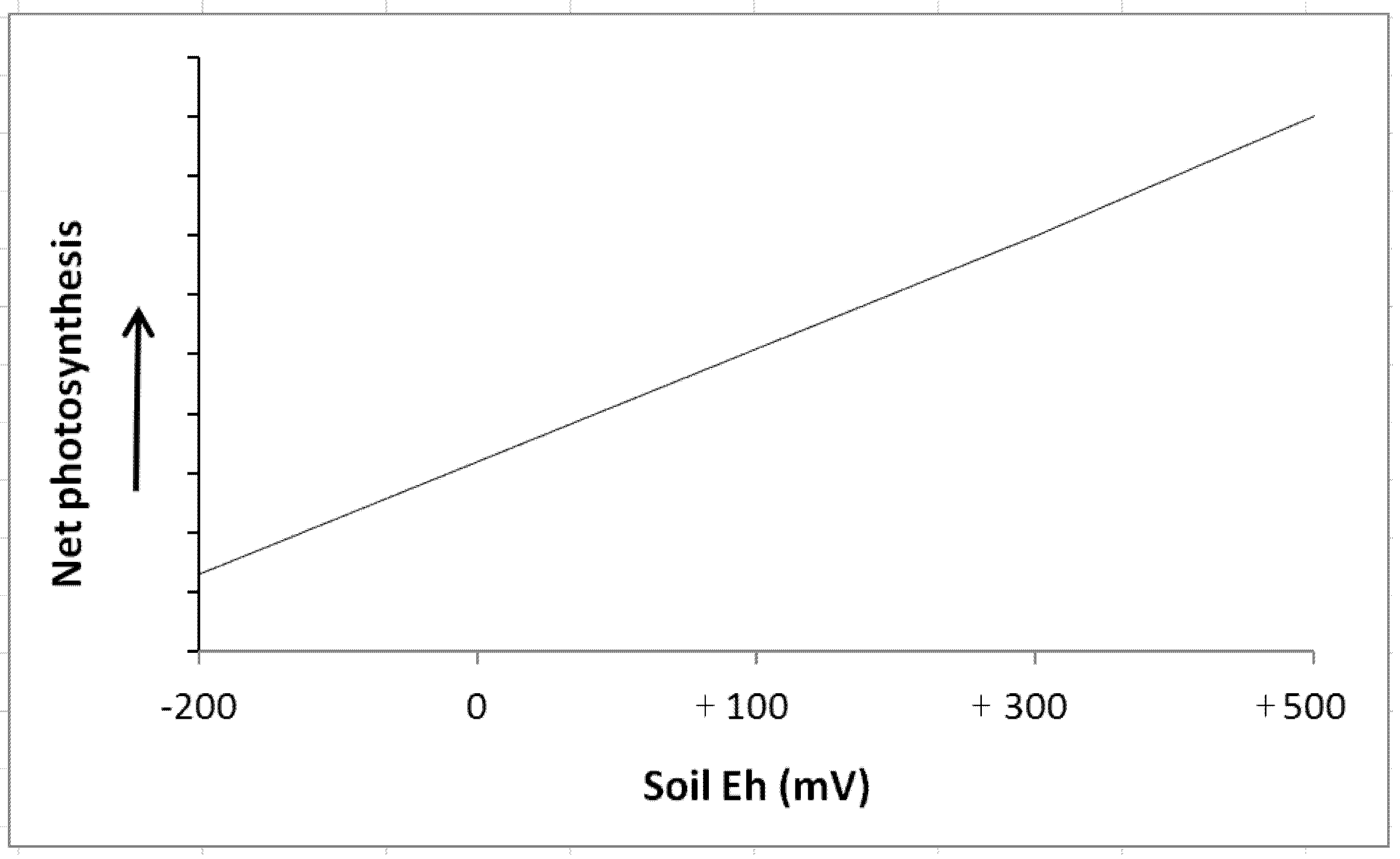
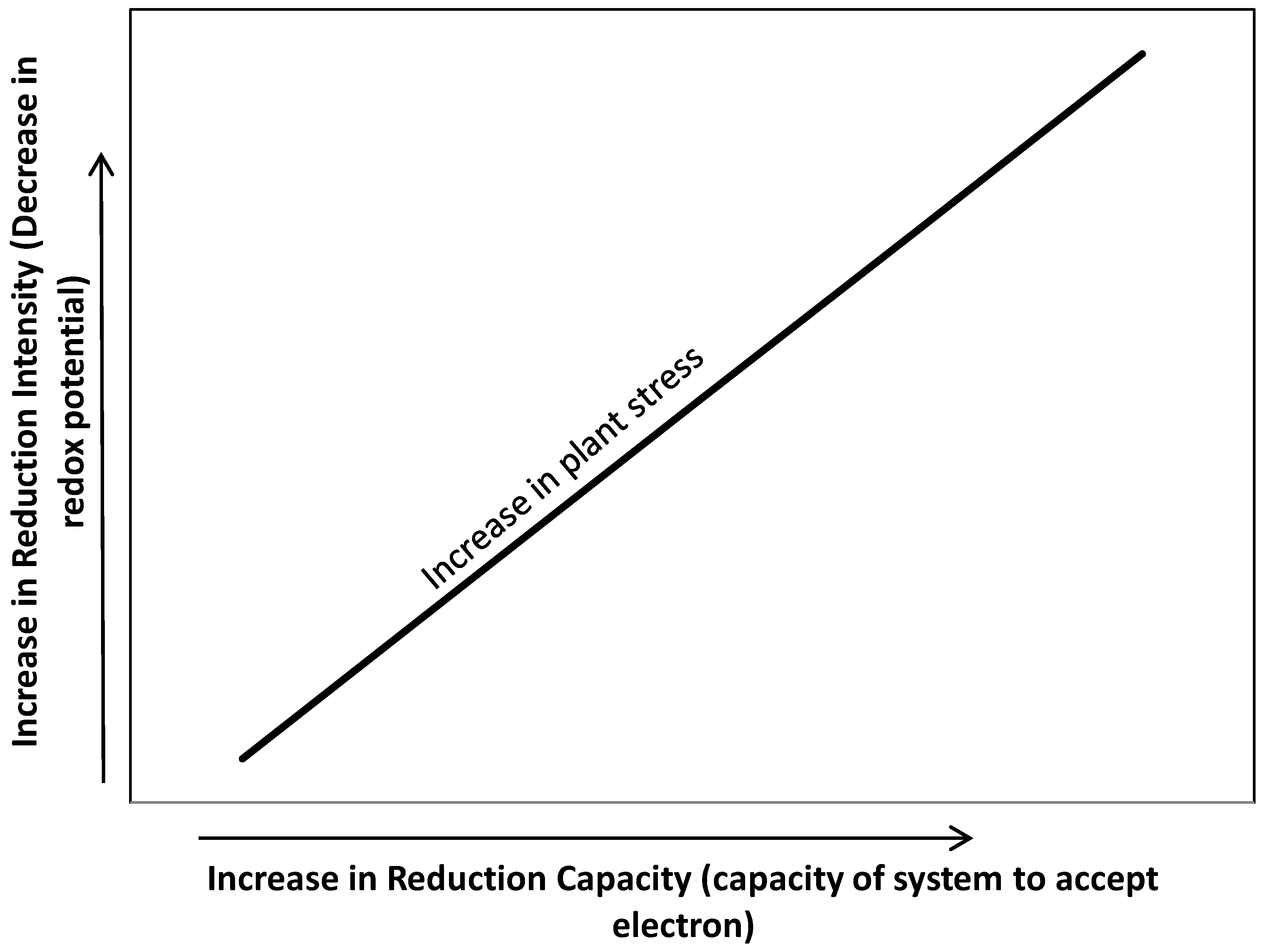
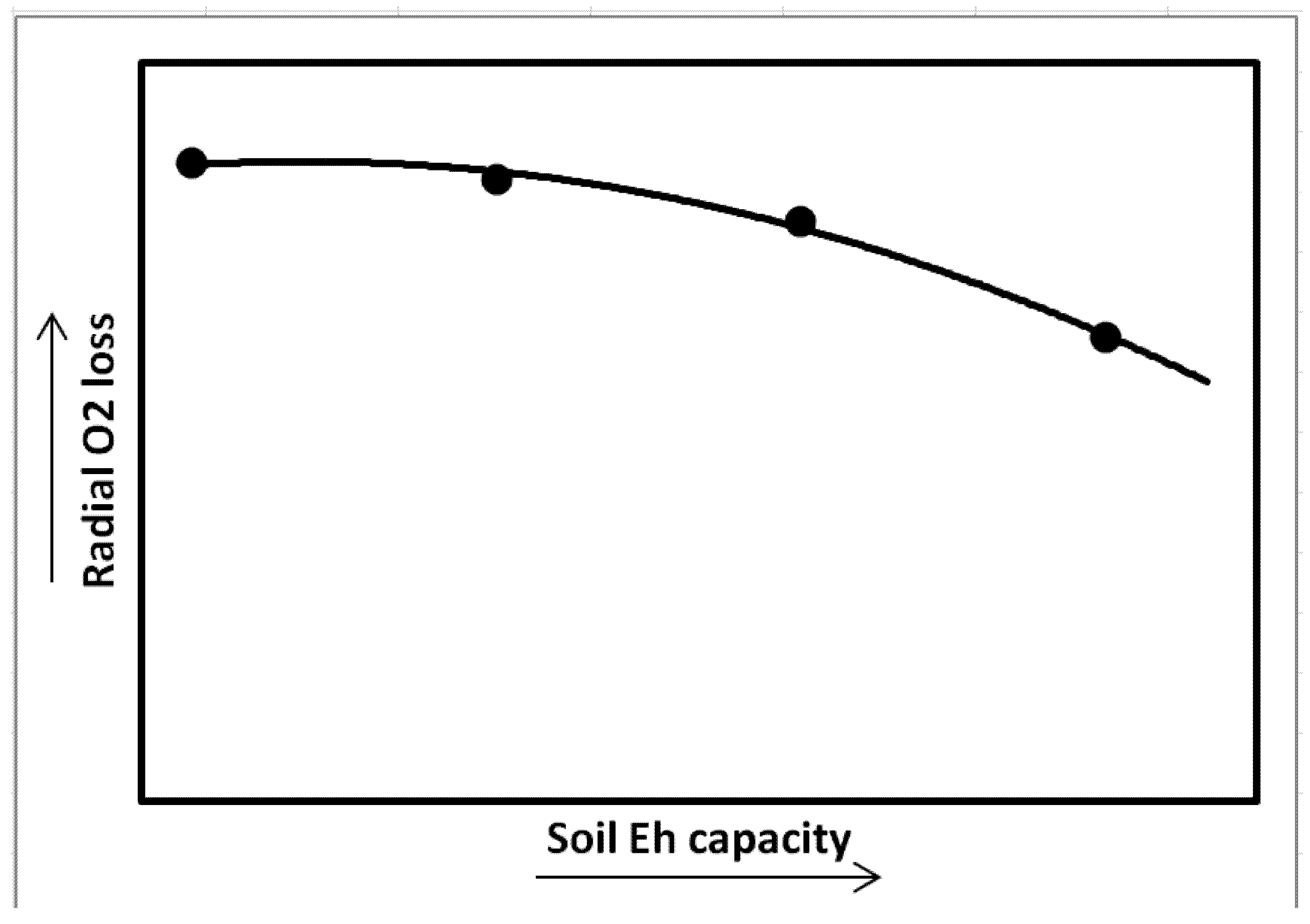
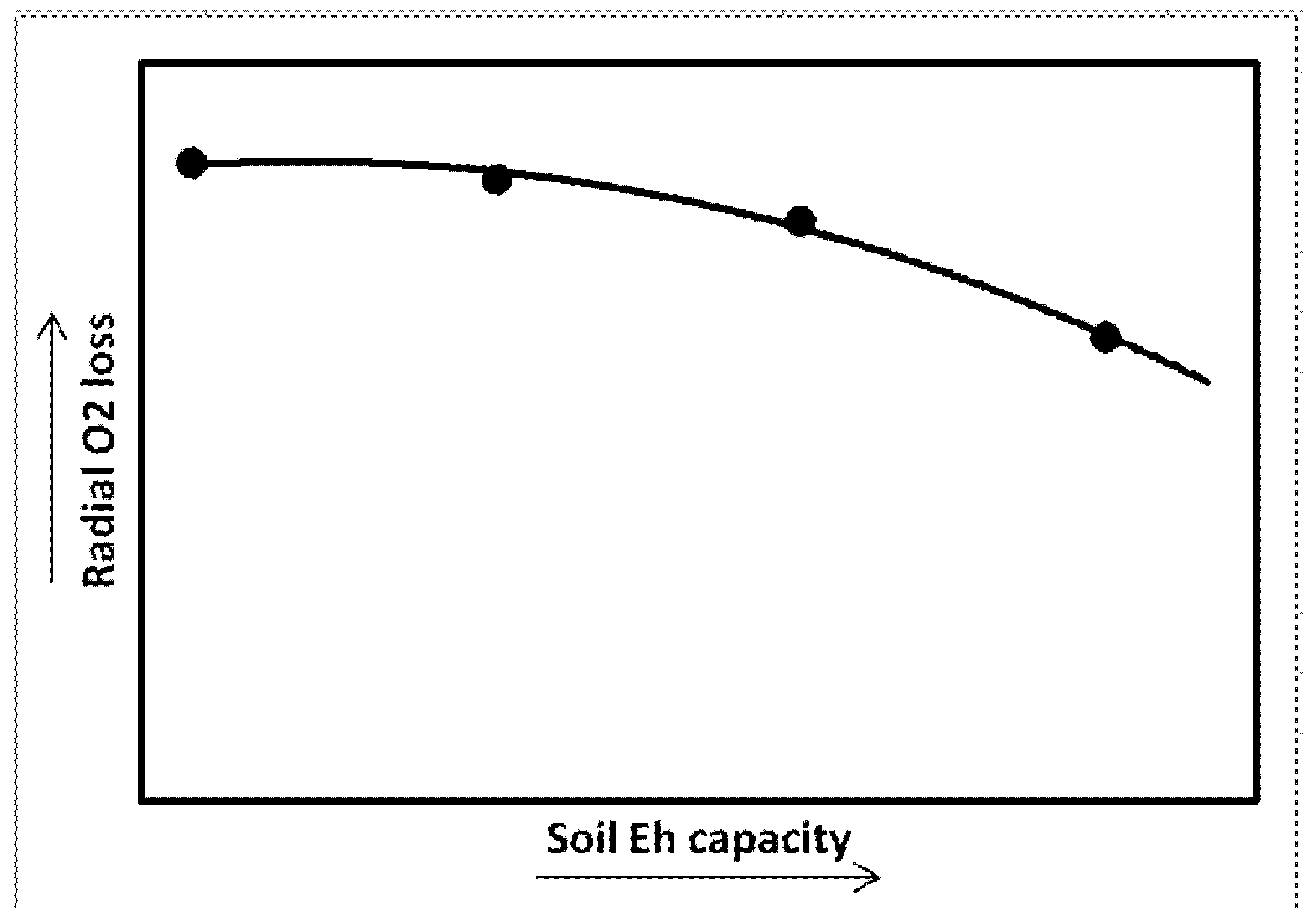
4. Intensity and Capacity of Soil Reduction
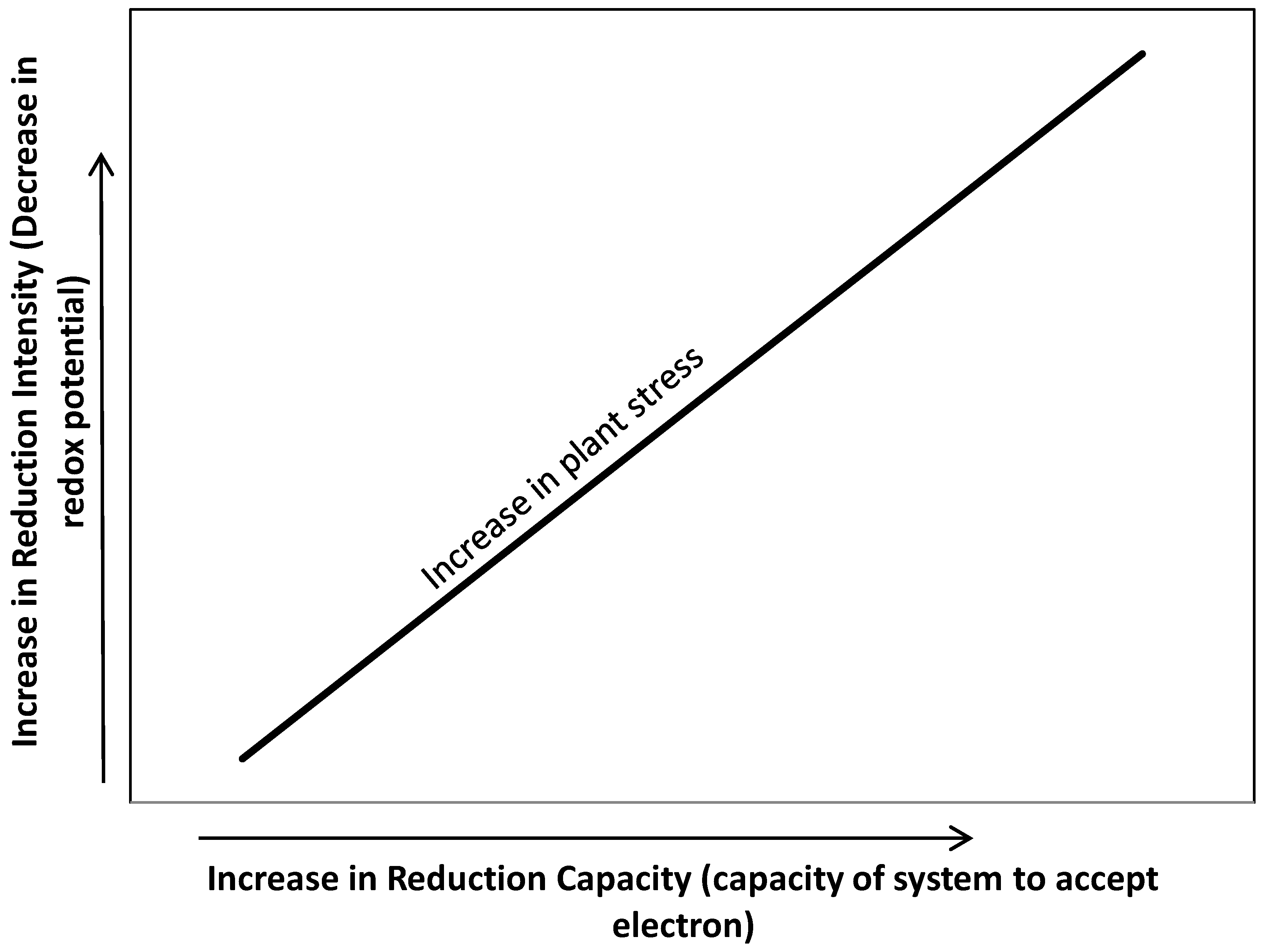
5. Soil Reduction and Wetland Plant Functioning
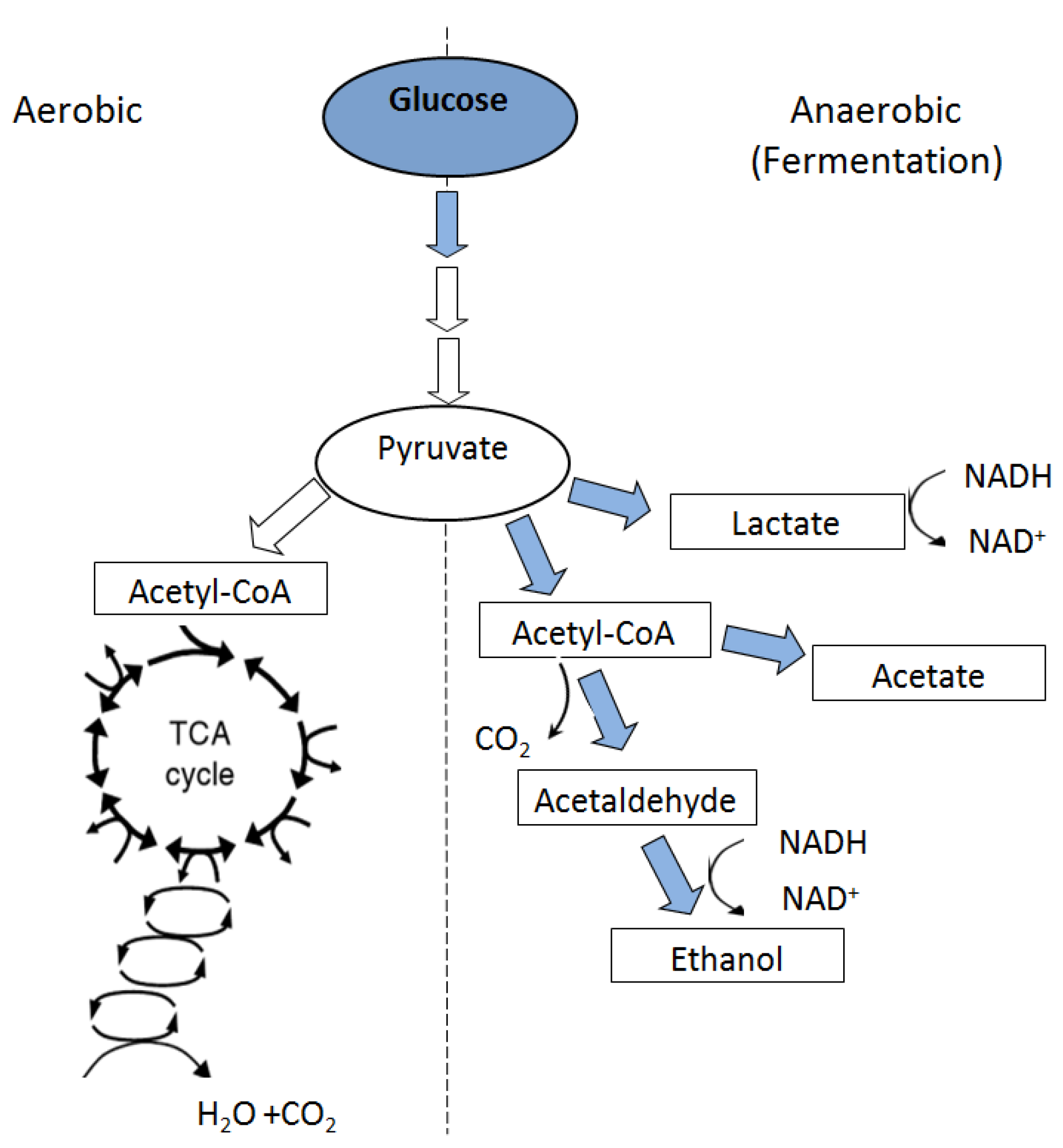
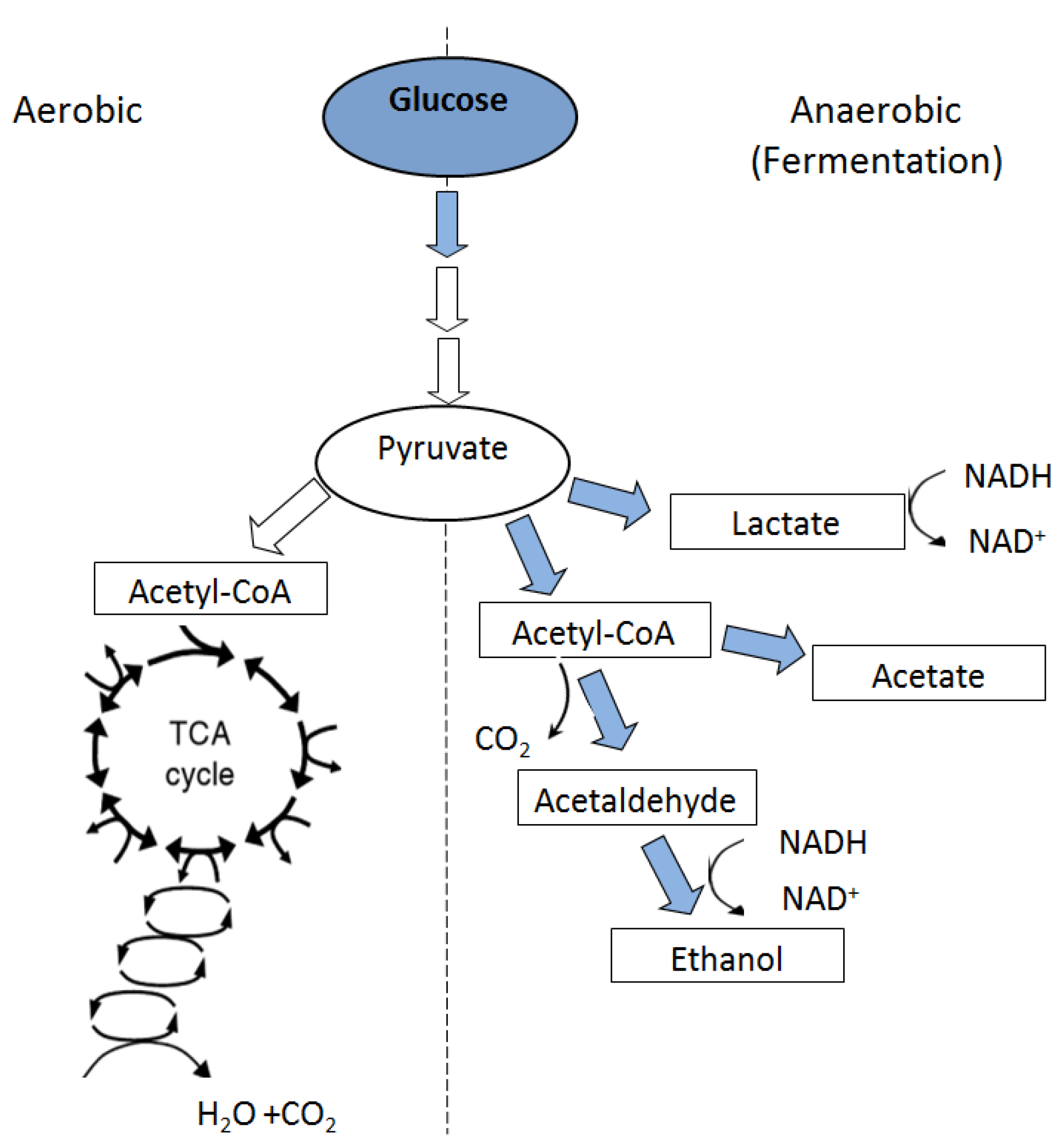
6. Metabolic Responses to Flooding
7. The Internal Oxygen Transport System
8. Plant Nutrition
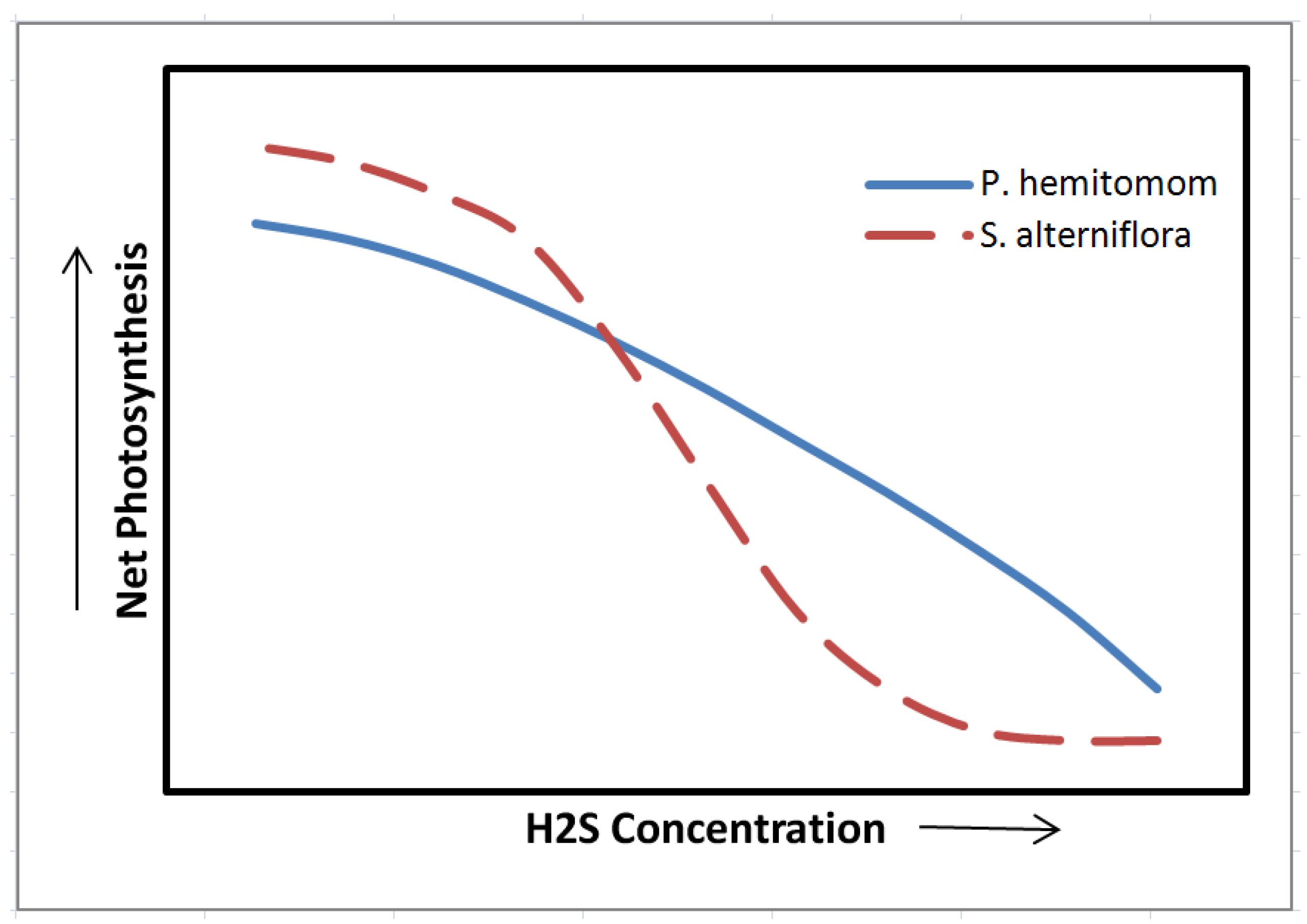
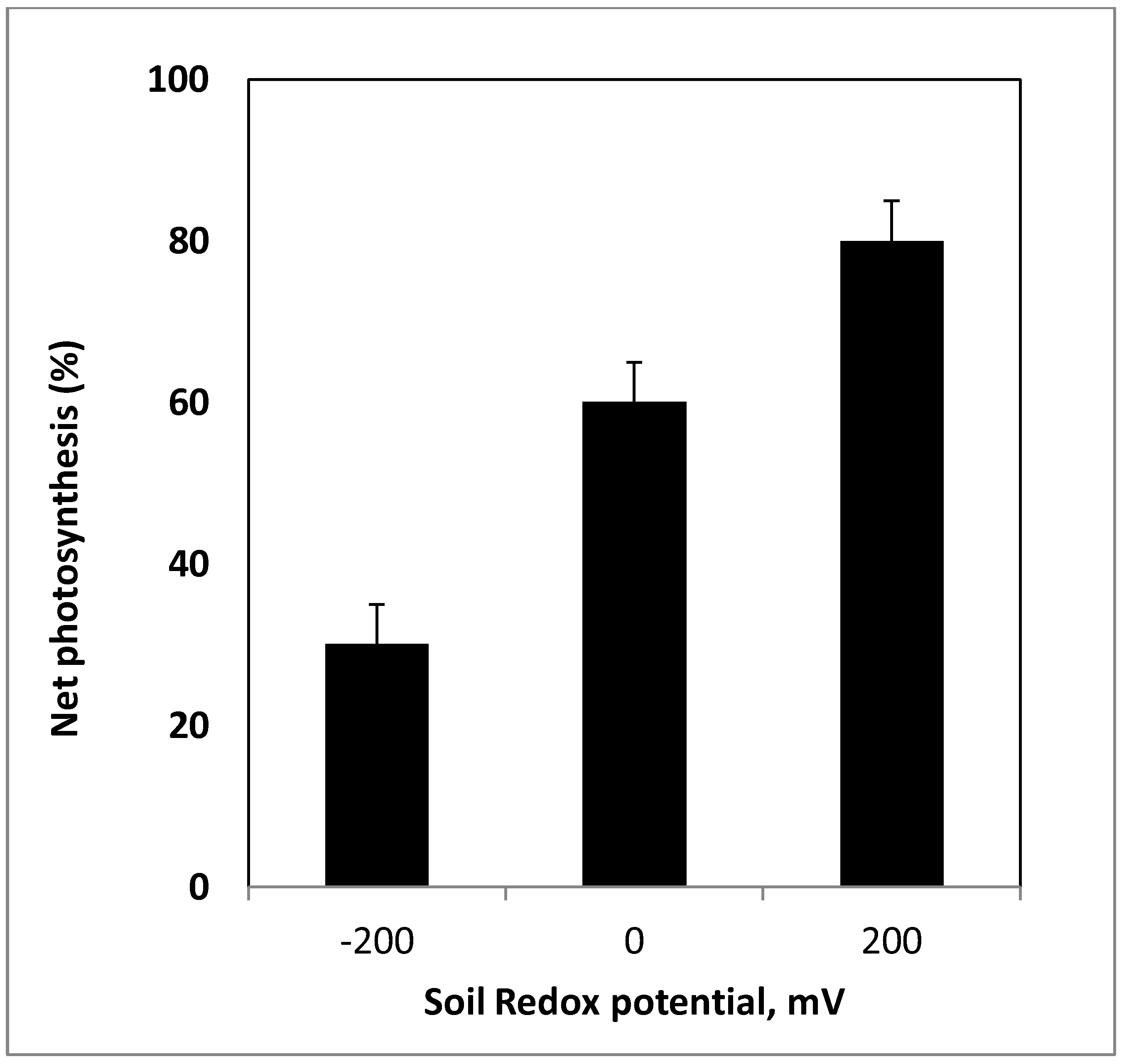
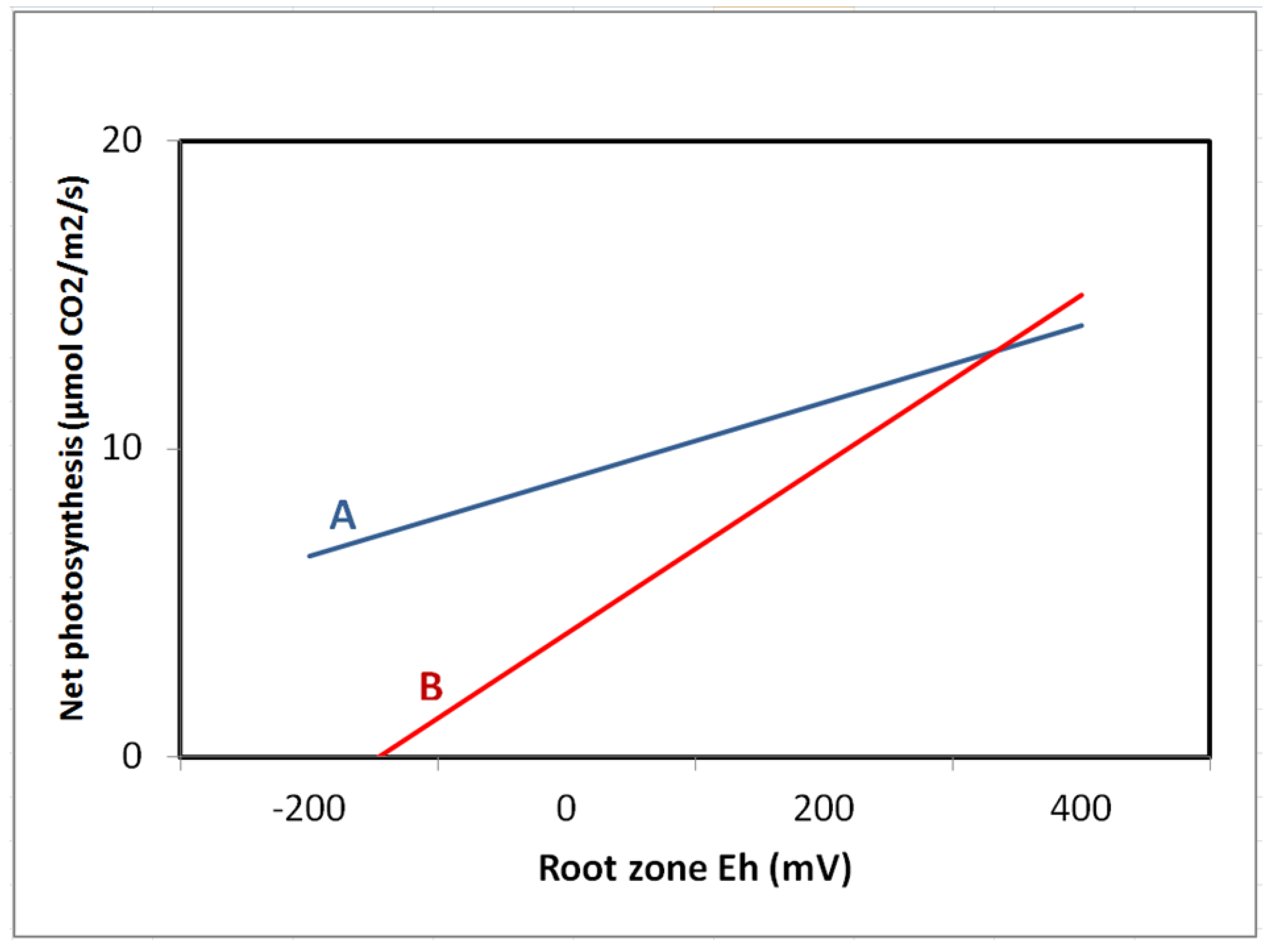
9. Plant Water Relations and Gas Exchange

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Soil Eh (mV) | Photosyn. Response (%) | Reference | |
|---|---|---|---|---|
| Marsh plants | ||||
| Typha domingensis | +200 | 18 | [157] | |
| 0 | 49 | [157] | ||
| −200 | 75 | [157] | ||
| Cladium Jamaicense | +200 | 24 | [157] | |
| 0 | 46 | [157] | ||
| −200 | 100 | [157] | ||
| Spartina alterniflora | <−200 | 15–21 | [131] | |
| Spartina patens | +230 | 7 | [91] | |
| −110 | 18 | [91] | ||
| Woody speccies | ||||
| Quercus lyrata | +340 | 54 | [93] | |
| +175 | 66 | [93] | ||
| Ligustrum sinese | +200/−200 | 86 | [132] | |
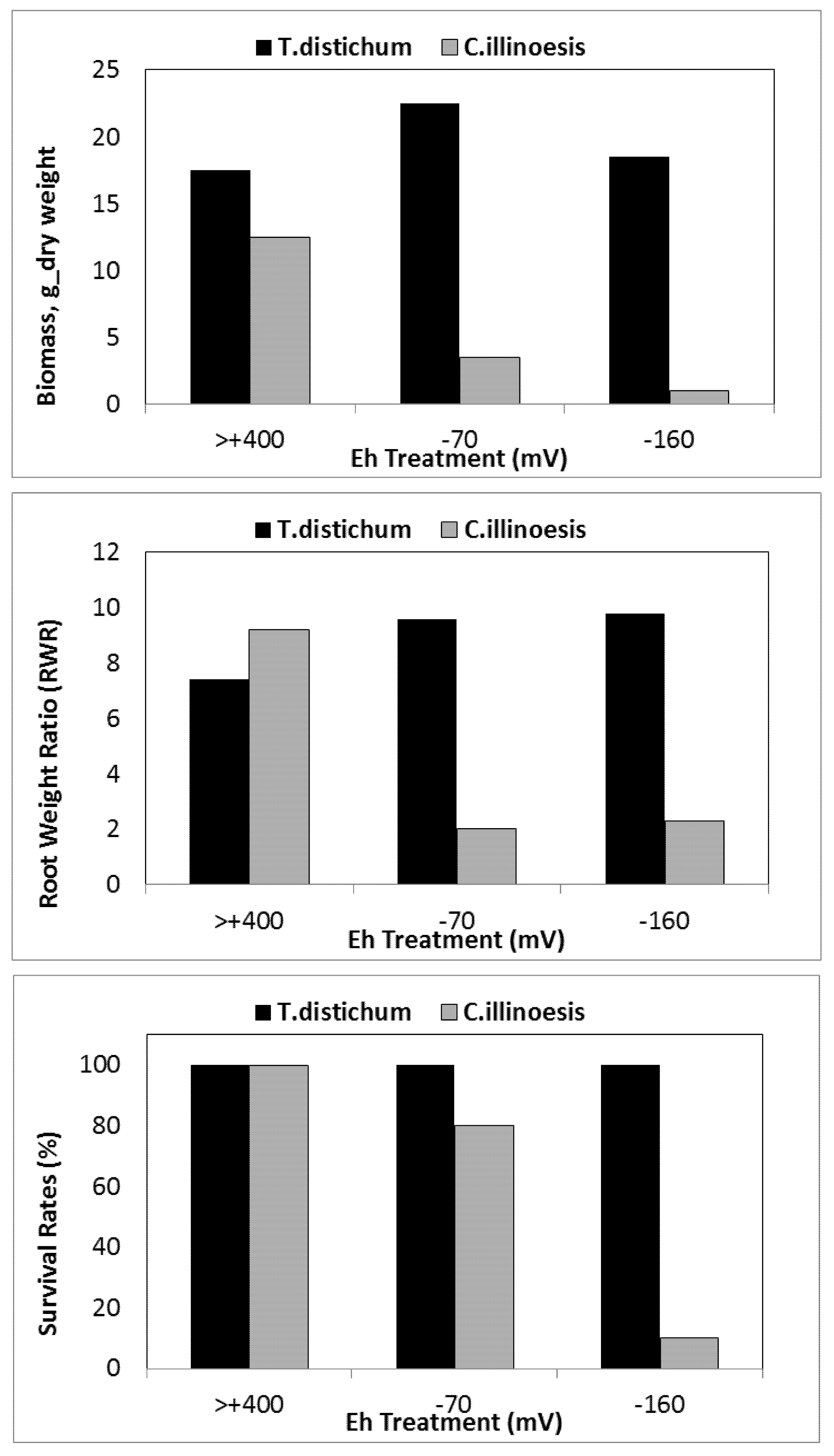
| Taxodium distichum | −70 | 5 | [158] | |
| −160 | 22 | [158] | ||
| Taxodium distichum | +350/+175 | 5 | [159] | |
| Taxodium distichum | −40/−70 | 36 | [128] | |
| +18/+172 | Decreased/recovered | [160] | ||
| Salix nigra | −130 | 53 | [158] | |
| +50 to −80 | 5 | [86] | ||
| Quercus nuttallii | +100/−220 | 35–68 | [88] | |
| Quercus nuttallii | +350/+175 | 21 | [159] | |
| Quercus falcata | +100/−220 | 65–87 | [88] | |
| Quercus michauxii | +350/+175 | 58 | [159] | |
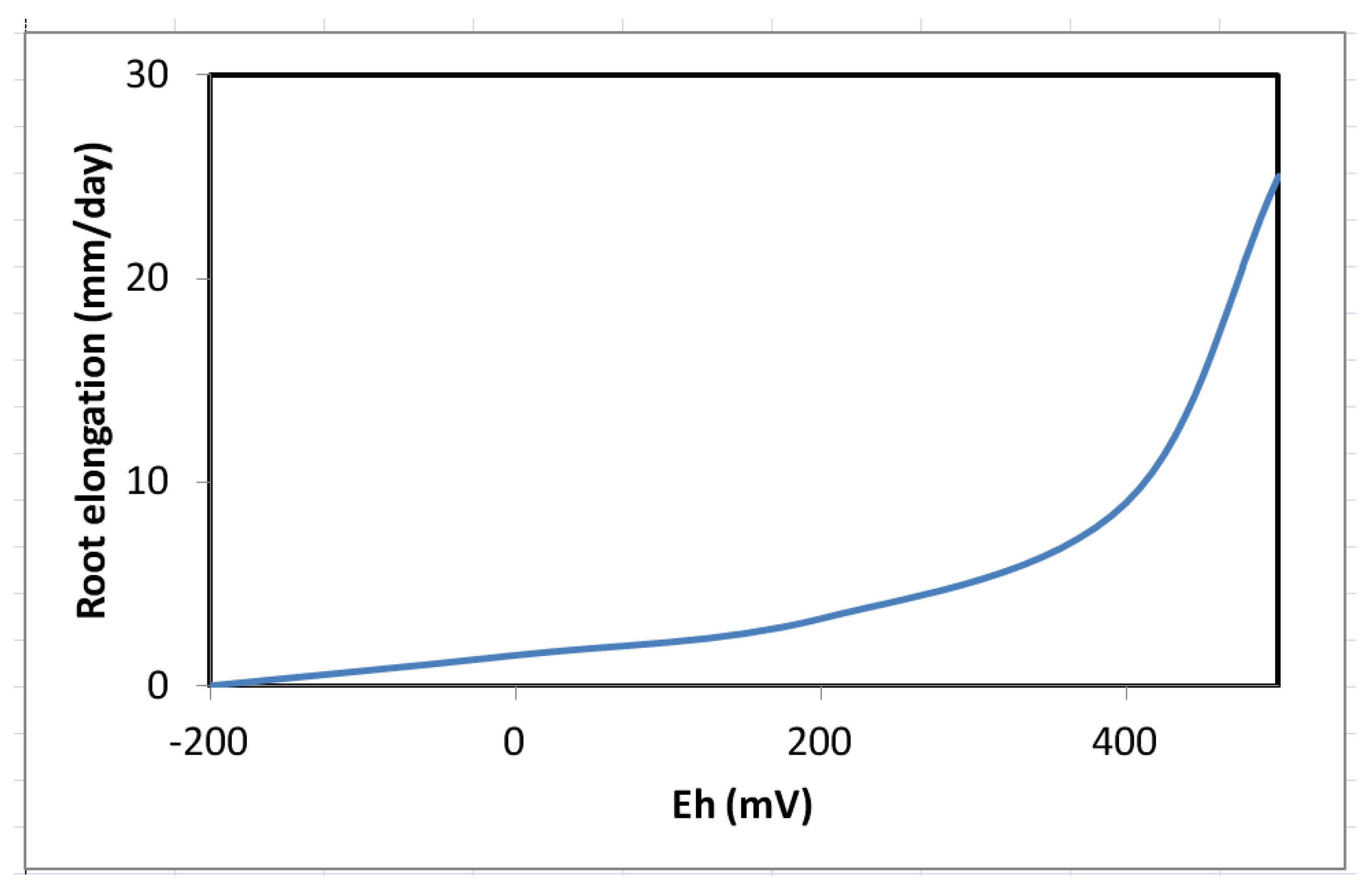
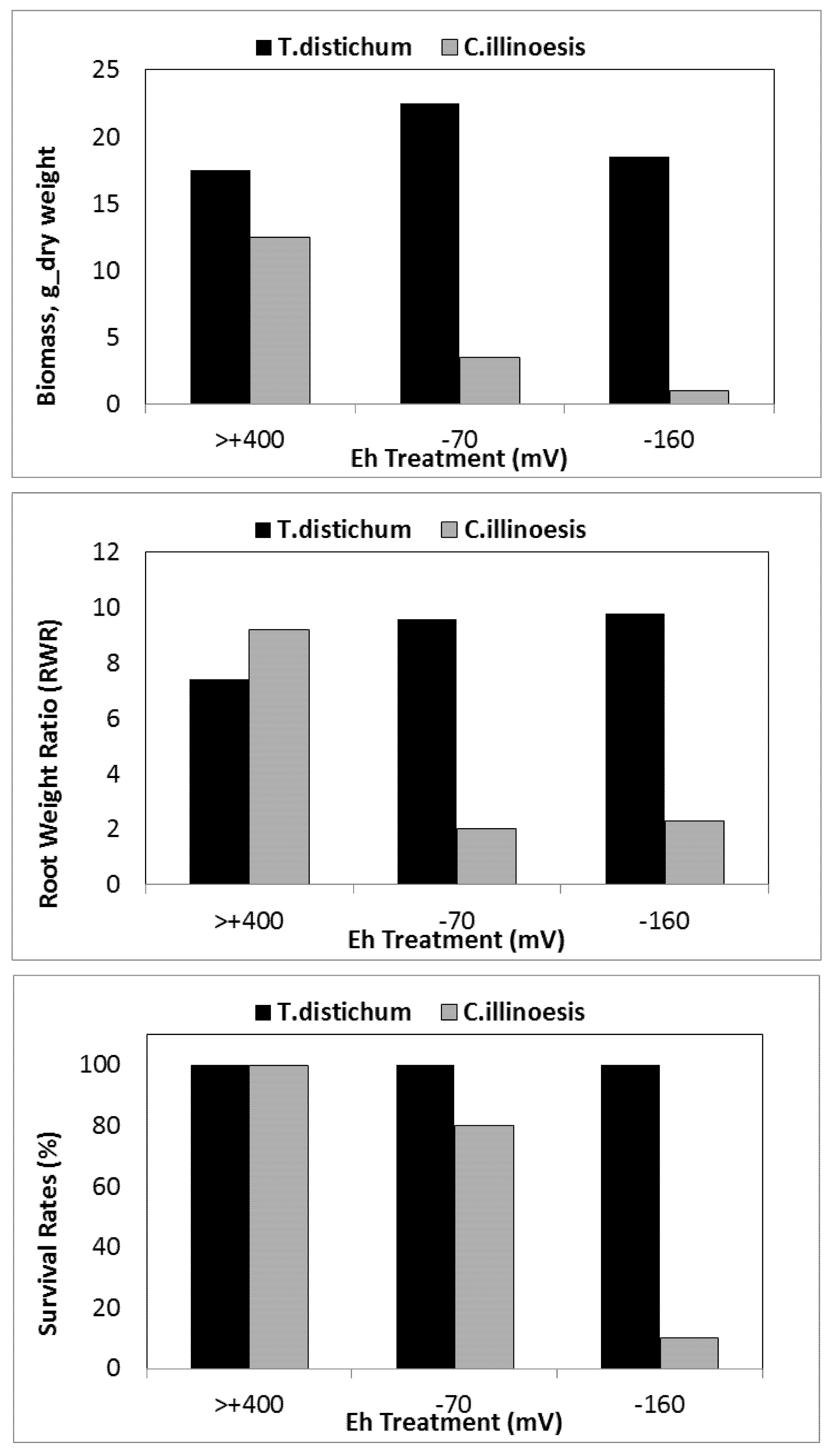
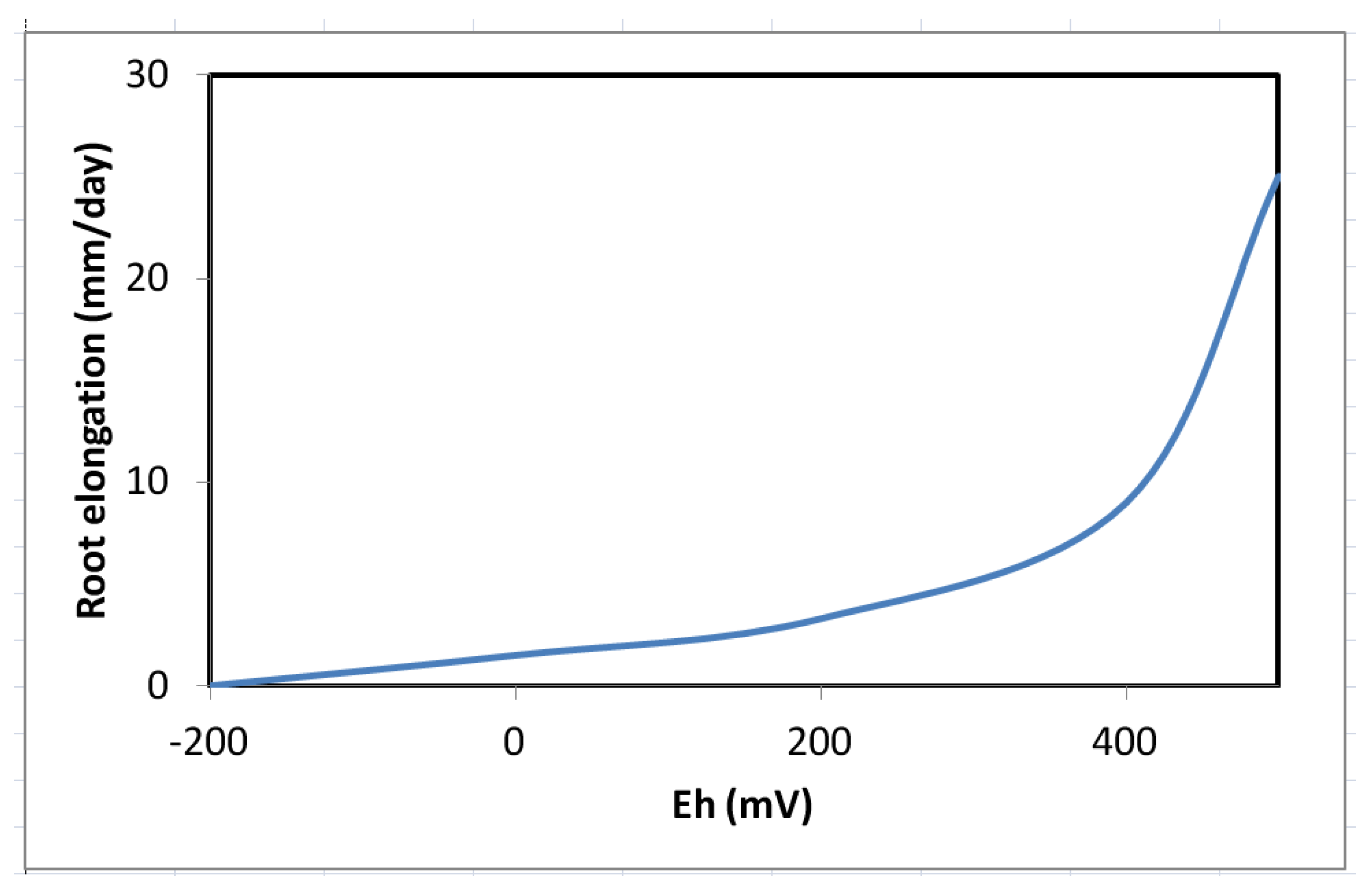
10. Growth and Biomass Production
11. Concluding Remarks
References
- Gambrell, R.P.; Patrick, W.H. Chemical and Microbiological Properties of Anaerobic Soils nad Sediments. In Plant Life in Anaerobic Environments; Hook, D.D., Crawford, R.M.M., Eds.; Ann Arbor Science: Ann Arbor, MI, USA, 1978; pp. 375–423. [Google Scholar]
- Ponnamperuma, F.N. Effects of Flooding on Soils. In Flooding and Plant Growth; Kozlowski, T.T., Ed.; Academic Press Inc.: Orlando, FL, USA, 1984; pp. 1–44. [Google Scholar]
- Gambrell, R.P.; DeLaune, R.D.; Patrick, W.H. Redox Processes in Soils Following Oxygen Depletion. In Plant Life Under Oxygen Deprivation: Ecology, Physiology, and Biochemistry; Jackson, M.B., Davies, D.D., Lambers, H., Eds.; SPB Academic Publishing BV: The Hague, The Netherlands, 1991; pp. 101–117. [Google Scholar]
- Jackson, M.B.; Drew, M.C. Effects of Flooding on Growth and Metabolism of Herbaceous Plants. In Flooding and Plant Growth; Kozlowski, T.T., Ed.; Academic Press, Inc: Waltham, MA, USA, 1984; pp. 47–128. [Google Scholar]
- Greenway, H.; Armstrong, W.; Colmer, T.D. Coditions leading to high CO2 (>5 kPa) in waterlogged flooded soils and possible effects on root growth and metabolism. Ann. Bot. 2006, 98, 9–32. [Google Scholar]
- Ponnamperuma, F.N. The chemistry of submerged soil. Adv. Agron. 1972, 24, 29–96. [Google Scholar]
- DeLaune, R.D.; Pezeshki, S.R.; Pardue, J.H. An oxidation-reduction buffer for evaluating physiological response of plants to root oxygen stress. Environ. Exp. Bot. 1990, 30, 243–247. [Google Scholar] [CrossRef]
- Patrick, W.H.; DeLaune, R.D. Chemical and biological redox systems affecting nutrient availability in the coastal wetlands. Geosci. Man 1977, XVIII, 131–137. [Google Scholar]
- Reddy, K.R.; DeLaune, R.D. Biogeochemistry of Wetalnds: Science and Applications; CRC Press: Boca Raton, FL, USA, 2008; p. 774. [Google Scholar]
- Pearsall, W.H.; Mortimer, C.H. Oxidation-reduction potentials in waterlogged soils, natural waters and muds. J. Ecol. 1939, 27, 483–501. [Google Scholar] [CrossRef]
- Mortimer, C.H. The exchange of dissolved substances between mud and water in lakes. J. Ecol. 1941, 29, 280–329. [Google Scholar]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 4th ed; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2007; p. 582. [Google Scholar]
- Kludze, H.K.; DeLaune, R.D. Straw application effects on Methane and oxygen exchange and growth in rice. Soil Sci. Soc. Am. J. 1995, 59, 824–830. [Google Scholar]
- Kludze, H.K.; DeLaune, R.D. Gaseous exchange and wetland plant response to soil redox intensity and capacity. Soil Sci. Soc. Am. J. 1995, 59, 939–945. [Google Scholar]
- Kludze, H.K. Gaseous exchange and wetland plant response to soil redox conditions. Ph.D. Dissertation, Louisiana State University, Baton Rouge, LA, USA, 1994. [Google Scholar]
- Kludze, H.K.; DeLaune, R.D.; Patrick, W.H. Aerenchyma formation and methane and oxygen exchange in rice. Soil. Sci. Soc. Am. J. 1993, 51, 386–391. [Google Scholar]
- Kludze, H.K.; Pezeshki, S.R.; DeLaune, R.D. Evaluation of root oxygenation and growth in baldcypress in response to short-term soil hypoxia. Can. J. For. Res. 1994, 24, 804–809. [Google Scholar] [CrossRef]
- Sorrell, B.K.; Brix, H.; Orr, P.T. Oxygen exchange by entire root system of Cyperus involucratus and Eleocharis sphacelata. J. Aquat. Plant Manage. 1993, 31, 24–28. [Google Scholar]
- Armstrong, W.; Brandle, R.; Jackson, M.B. Mechanisms of flood tolerance in plants. Acta Bot. Neerl. 1994, 43, 307–358. [Google Scholar]
- Armstrong, J.; Armstrong, W.; Beckett, P.M.; Halder, J.E.; Lythe, S.; Holt, R.; Sinclair, A. Pathways of aeration and the mechanisms and beneficial effects of humidity- and venturi-induced convections in Phragmites australis. Aquat. Bot. 1996, 54, 177–197. [Google Scholar] [CrossRef]
- Armstrong, J.; Armstrong, W.; van Der Putten, W.H. Phragmites die-back: Bud and root death, blockage within the aeration and vascular systems and the possible role of phytotoxins. New Phytol. 1996, 133, 399–414. [Google Scholar]
- Pezeshki, S.R. Plant Responses to Flooding. In Plant-Environment Interactions; Wilkinson, R.E., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1994; pp. 289–321. [Google Scholar]
- Pezeshki, S.R. Wetland plant responses to flooding. Environ. Exp. Bot. 2001, 46, 299–312. [Google Scholar] [CrossRef]
- Sorrell, B.K. Effect of external oxygen demand on radia oxygen loss by Juncus roots in titanium citrate solutions. PlantCell Environ. 1999, 22, 1587–1593. [Google Scholar]
- Van Wijck, C.; de Groot, C.J.; Grillas, P. The effect of anaerobic sediment on the growth of Potamogeton pectinatus: The role of organic matter, sulphide, and ferrous iron. Aquat. Bot. 1992, 44, 31–49. [Google Scholar] [CrossRef]
- Laskov, C.; Horn, O.; Hupfer, M. Environmental factors regulating the radial oxygen loss from roots of Myriophyllum spicatum and Potamogeton crispus. Aquat. Bot. 2006, 84, 333–340. [Google Scholar]
- Armstrong, W.; Wright, E.J.; Lythe, S.; Gaynard, T.J. Plant zonation and the effects of the spring-neap tidal cycle on soil aeration in a humber salt marsh. J. Ecol. 1985, 73, 323–339. [Google Scholar] [CrossRef]
- Pennington, M.R.; Walters, M.B. The response of vegetation zonation and soil redox potential in created wetlands. For. Ecol. Manag. 2006, 233, 1–10. [Google Scholar] [CrossRef]
- Davy, A.J.; Brown, M.J.H.; Mossman, H.L.; Grant, A. Colonization of a newly developing saltmarsh: Disentangling independent effects of elevation and redox potential on halophytes. J. Ecol. 2001, 99, 1350–1357. [Google Scholar]
- Justin, S.H.F.W.; Armstrong, W. The anatomical characteristics of roots and plant response to soil flooding. New Phytol. 1987, 106, 465–495. [Google Scholar]
- Pedersen, O.; Sand-Jensen, K. Adaptation of submerged Lobelia dortmanna to aerial life form: Morphology, carbon sources, and oxygen dynamics. Oikos 1992, 65, 89–96. [Google Scholar] [CrossRef]
- Pedersen, O.; Sand-Jensen, K.; Revsbech, N.P. Diel pulses of O2 and CO2 in sandy lake sediments inhabited by Lobelia dortmanna. Ecology 1995, 76, 1536–1545. [Google Scholar]
- Drew, M.C. Sensing soil oxygen. Plant Cell Environ. 1990, 13, 681–693. [Google Scholar] [CrossRef]
- Drew, M.C. Oxygen deficiency and root metabolism: Injury and acclimation under hypoxia and anoxia. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 223–250. [Google Scholar]
- Hook, D.D.; Crawford, R.M.M. Plant Life in Anaerobic Environments; Ann Arbor Science: Woburn, MA, USA, 1978; p. 564. [Google Scholar]
- Vartapetian, B.B.; Jackson, M.B. Plant adaptations to anaerobic stress. Ann. Bot. 1997, 79, 3–20. [Google Scholar] [CrossRef]
- Armstrong, W. The relationship between oxidation-reduction potentials and oxygen diffusion levels in some waterlogged organic soils. J. Soil Sci. 1967, 18, 27–34. [Google Scholar]
- Turner, F.T.; Patrick, W.H. Chemical changes in waterlogged soils as a result of oxygen depletion. Trans. 9th Int. Congr. Soil Sci. 1968, 4, 53–56. [Google Scholar]
- Armstrong, W.; Armstrong, J.; Beckett, P.M. Pressurized aeration in wetland macrophytes: Some theoretical aspects of humidity-induced convection and thermal transpiration. Folia Geobot. Phytotax. 1996, 31, 25–36. [Google Scholar] [CrossRef]
- Greenway, H.; Gibbs, J. Mechanisms of anoxia tolerance in plants: II> Energy requirements for maintenance and energy distribution to essential processes. Funct. Plant Biol. 2003, 30, 999–1036. [Google Scholar] [CrossRef]
- Voesenek, L.A.C.J.; Colmer, T.D.; Pierik, R.; Millenaar, F.F.; Peeters, A.J.M. How plants cope with submergence. New Phytol. 2006, 170, 213–226. [Google Scholar]
- DeLaune, R.D.; Smith, C.J.; Patrick, W.H. Nitrogen losses from a Louisiana Gulf Coast salt marsh. Est. Coast. Shelf Sci. 1983, 17, 133–141. [Google Scholar] [CrossRef]
- Armstrong, J.; Drew, M.C. Root Growth and Metabolism under Oxygen Deficiency. In Plant Roots: The Hidden Half; Waisel, Y., Eshel, A., Kafkaf, U., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 729–761. [Google Scholar]
- DeLaune, R.D.; Patrick, W.H.; Buresh, R.J. Sedimentation rates determined by Cs-137 dating in a rapidly accreting salt marsh. Nature 1978, 275, 532–533. [Google Scholar]
- Drew, M.C.; Lynch, J.M. Soil anaerobiosis, micro-organisms and root functions. Ann. Rev. Phytopathol. 1980, 18, 37–66. [Google Scholar] [CrossRef]
- King, G.M.; Klug, M.J.; Wiegert, R.G.; Chalmers, A.G. Relation of soil water movement and sulfide concentration to Spartina alterniflora production in a Georgia satlmarsh. Science 1982, 218, 61–63. [Google Scholar]
- Ingold, A.; Havill, D.C. The influence of sulfide on the distribution of higher plants in salt marshes. J. Ecol. 1984, 72, 1043–1054. [Google Scholar] [CrossRef]
- Havill, D.C.; Ingold, A.; Pearson, J. Sulfide tolerance in coastal halophytes. Vegetatio 1985, 62, 279–285. [Google Scholar] [CrossRef]
- Tanaka, A.; Mulleriyawa, R.P.; Yasu, T. Possibility of hydrogen sulfide induced iron toxicity of the rice plant. Soil Sci. Plant Nutr. 1968, 4, 1–6. [Google Scholar]
- Allam, A.I.; Hollis, J.P. Sulfide inhibition of oxidases in rice roots. Phytopathology 1972, 62, 634–639. [Google Scholar] [CrossRef]
- Rao, D.N.; Mikkelsen, D.S. Effects of acidic, propionic, and butyric acids on rice seedling growth and nutrition. Plant Soil 1977, 47, 323–334. [Google Scholar] [CrossRef]
- Armstrong, J.; Armstrong, W. Phragmites dieback: Toxic effects of propionic, butyric and caproic acids in relation to pH. New Phytol. 1999, 142, 201–217. [Google Scholar] [CrossRef]
- Atwell, B.J.; Greenway, H. The relationship between growth and oxygen uptake in hypoxic rice seedlings. J. Exp. Bot. 1987, 38, 454–465. [Google Scholar]
- Armstrong, W. Aeration in higher plants. Adv. Bot. Res. 1979, 7, 225–332. [Google Scholar] [CrossRef]
- Drew, M.C. Soil aeration and plant root metabolism. Soil Sci. 1992, 154, 259–268. [Google Scholar] [CrossRef]
- Hopkins, W.G.; Huner, N.P. Introduction to Plant Physiology, 4th ed; John Wiley & Sons, Inc.: New York, NY, USA, 2009; p. 503. [Google Scholar]
- Keeley, J.E. Population differentation along a flood frequency gradient: Physiological adaptation to flooding in Nyssa sylvatica. Ecol. Monogr. 1979, 49, 89–108. [Google Scholar] [CrossRef]
- Mendelssohn, I.A.; McKee, K.L.; Patrick, W.H. Oxygen deficiency in Spartina alterniflora roots: Metabolic adaptation to anoxia. Science 1981, 214, 439–441. [Google Scholar]
- Crawford, R.M.M. Oxygen availability as an ecological limit to plant distribution. Adv. Ecol. Res. 1992, 23, 93–285. [Google Scholar] [CrossRef]
- Kennedy, R.A.; Rumpho, M.E.; Fox, T.C. Aerobic metabolism in plants. Plant Phys. 1992, 100, 1–6. [Google Scholar] [CrossRef]
- Kogawara, S.; Yamanoshita, T.; Norisada, M.; Masumori, M.; Kojima, K. Photosynthesis and photoassimilate transport during root hypoxia in Melaleuca cajuputi, a flood-tolerant species, and in Eucalyptus camadulensis, a moderately floo-tolerant species. Tree Physiol. 2006, 26, 1413–1423. [Google Scholar] [CrossRef]
- Roberts, J.K.M.; Andrade, F.H.; Anderson, J.C. Further evidence that cytoplasmic acidosis is a determinant of flooding intolerance in plants. Plant Physiol. 1985, 77, 492–494. [Google Scholar]
- Barmore, C.R. Effect of ethylene on chlorophyllase activity and chlorophyll content in calamond in rind tissue. HortScience 1975, 10, 595–596. [Google Scholar]
- Berry, J.; Bjorkman, O. Photosynthetic and adaptation to temperature in higher plants. Annu. Rev. Plant Physiol. 1980, 31, 491–543. [Google Scholar] [CrossRef]
- Majumdar, S.; Ghosh, S.; Glick, B.R.; Dumbroff, E.B. Activities of chlorophyllase, phosphoenolpyruvate carboxylase and ribulose-1, 5-bisphosphate carboxylase in the primary leavaes of soybean during senescence and drought. Physiol. Plant 1991, 81, 473–480. [Google Scholar] [CrossRef]
- Pezeshki, S.R. Response of baldcypress seedlings to hypoxia: Leaf protein content, ribulose-1, 5-bisphosphate carboxylase/oxygenase activity and photosynthesis. Photosynthetica 1994, 30, 59–68. [Google Scholar]
- Li, S.; Pezeshki, S.R.; Goodwin, S.; Shields, F.D. Physiological responses of black willow (Salix nigra) cuttings to a range of soil moisture regimes. Photosynthetica 2004, 42, 585–590. [Google Scholar] [CrossRef]
- Saglio, P.H.; Drew, M.C.; Pradet, A. Metabolic acclimation to anoxia induced by low (2–4 KPa partial pressure) oxygen pretreatment (hypoxia) in root tips of Zea mays. Plant Physiol. 1988, 86, 61–66. [Google Scholar] [CrossRef]
- Bertani, A.; Brambilla, I.; Menegus, F. Effect of anaerobiosis on rice seedlings: Growth, metabolic rate and fate of fermentation products. J. Exp. Bot. 1980, 31, 325–331. [Google Scholar]
- MacDonald, R.C.; Kimmerer, T.W. Metabolism of transpired ethanol by eastern cottonwood (Populus deltoides). Plant Physiol. 1993, 102, 173–179. [Google Scholar]
- Jackson, M.B. Long-distance signaling from roots to shoots assessed: The flooding story. J. Exp. Bot. 2002, 53, 175–181. [Google Scholar] [CrossRef]
- Dat, J.F.; Capelli, N.; Folzer, H.; Bourgeade, P.; Badot, P.-M. Sensing and signaling during plant flooding. Plant Physiol. Biochem. 2004, 42, 273–282. [Google Scholar] [CrossRef]
- Hattori, Y.; Nagai, K.; Furukawa, S.; Song, X.; Kawano, R.; Sakakibara, H.; Wu, J.; Matsumoto, T.; Yoshimura, A.; Kitano, H.; et al. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature 2009, 460, 1026–1030. [Google Scholar]
- McKee, K.L. Growth and physiological responses of neotropical mangrove seedlings to root zone hypoxia. Tree Physiol. 1996, 16, 883–889. [Google Scholar] [CrossRef]
- Youssef, T.; Saenger, P. Anatomical adaptive strategies to flooding and rhizophere oxidation in mangrove seedlings. Aust. J. Bot. 1996, 44, 297–313. [Google Scholar] [CrossRef]
- Youssef, T.; Saenger, P. Photosynthetic gas exchange and accumulation of phytotoxins in mangrove seedlings in response to soil physico-chemical characteristics associated with waterlogging. Tree Physiol. 1998, 18, 317–324. [Google Scholar] [CrossRef]
- Teal, J.M.; Kanwisher, J.W. Gas transport in the marsh grass Spartina alterniflora. J. Exp. Bot. 1966, 17, 355–361. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Responses of woody plants to flooding and salinity. Tree Physiol. Monogr. 1997, 1, 1–29. [Google Scholar]
- Armstrong, W.; Justin, S.H.F.W.; Beckett, P.M.; Lythe, S. Root adaptation to soil waterlogging. Aquat. Bot. 1991, 39, 57–73. [Google Scholar] [CrossRef]
- Perata, P.; Alpi, A. Plant responses to anaerobiosis. Plant Sci. 1993, 93, 1–17. [Google Scholar] [CrossRef]
- Iwanaga, F.; Yamamoto, F. Growth, morphology and photosynthetic activity in flooded Alnus japonica seedlings. J. For. Res. 2007, 12, 243–246. [Google Scholar] [CrossRef]
- Iwanaga, F.; Yamamoto, F. Effects of flooding depth on growth, morphology and photosynthesis in Alnus japonica species. New For. 2008, 35, 1–14. [Google Scholar] [CrossRef]
- Dacey, J.W.H. Pressurized ventilation in the yellow water lily. Ecology 1981, 62, 1137–1147. [Google Scholar] [CrossRef]
- Armstrong, W.; Beckett, P.M. Internal aeration and the development of stelar anoxia in submerged roots: A multi-shelled mathematical model combining axial diffusion of oxygen in the cortex with radial losses to the stele, the wall layers, and the rhizosphere. New Phytol. 1987, 105, 221–245. [Google Scholar] [CrossRef]
- Luxmoore, R.J.; Sojka, R.E.; Stolzy, L.H. Root porosity and growth responses of wheat to aeration and light intensity. Soil Sci. 1972, 113, 354–357. [Google Scholar]
- Li, S.; Pezeshki, S.R.; Shields, F.D. Partial flooding enhances aeration in adventitious roots of black willow (Salix nigra) cuttings. J. Plant Physiol. 2006, 163, 619–628. [Google Scholar] [CrossRef]
- Kludze, H.K.; DeLaune, R.D. Methane emission and growth of Spartina patens in response to soil redox intensity. Soil Sci. Soc. Am. J. 1994, 58, 1838–1845. [Google Scholar]
- Pezeshki, S.R.; Anderson, P.A. Responses of three bottomland woody species with different flood-tolerance capabilities to various flooding regimes. Wetl. Ecol. Manag. 1997, 4, 245–256. [Google Scholar]
- Brix, H.; Sorrell, B.K. Oxygen stress in wetland plants: Comparison of de-oxygenated and reducing root environments. Funct. Ecol. 1996, 10, 521–526. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; Matthews, S.W.; DeLaune, R.D. Root cortex structure and metabolic response of Spartina patens to soil redox conditions. Environ. Exp. Bot. 1991, 31, 91–97. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; Pardue, J.H.; DeLaune, R.D. The influence of oxygen deficiency and redox potential on alcohol dehydrogenase activity, root porosity, ethylene production and photosynthesis in Spartina patens. Environ. Exp. Bot. 1993, 33, 565–573. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; DeLaune, R.D.; Anderson, P.H. Effect of flooding on elemental uptake and biomass allocation in seedlings of three bottomland tree species. J. Plant Nutr. 1999, 22, 1481–1494. [Google Scholar] [CrossRef]
- DeLaune, R.D.; Pezeshki, S.R.; Lindau, C.W. Influence of soil redox potential on nitrogen uptake and growth of wetland oak seedlings. J. Plant Nutr. 1998, 21, 757–768. [Google Scholar] [CrossRef]
- DeLaune, R.D.; Jugsujinda, A.; Reddy, K.R. Effect of root oxygen stress on phosphorus uptake by cattail. J. Plant Nutr. 1999, 22, 459–466. [Google Scholar] [CrossRef]
- Else, M.; Coupland, A.D.; Dutton, L.; Jackson, M.B. Decreased root hydraulic conductivity reduces leaf water potential, initiates stomatal closure, and slows leaf expansion in flooded plantsof castor oil (Ricinus communis) despite diminished delivery of ABA from roots to shoots in xylem sap. Physiol. Plant 2001, 111, 46–54. [Google Scholar]
- Jackson, M.B.; Saker, L.R.; Crisp, C.M.; Else, M.A.; Janowiak, F. Ionic and pH signaling from roots to shoots of flooded tomato plants in relation to stomatal closure. Plant Soil 2003, 253, 103–113. [Google Scholar] [CrossRef]
- Gibbs, J.; Turner, D.W.; Armstrong, W.; Darwent, M.J.; Greenway, H. Response to oxygen deficiency in primary maize roots. I. Development of oxygen deficiency in the stele reduces radial solute transport to the xylem. Aust. J. Plant Physiol. 1998, 25, 745–758. [Google Scholar] [CrossRef]
- Hook, D.D.; DeBell, D.S.; McKee, W.H.; Askew, J.L. Responses of loblolly pine (mesophyte) and swamp tupelo (hydrophyte) seedlings to soil flooding and phosphorous. Plant Soil 1983, 71, 387–394. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; Pan, S.Z.; DeLaune, R.D.; Patrick, W.H. Sulfide-induced toxicity: Inhibition of carbon assimilation in Spartina alterniflora. Photosynthetica 1988, 22, 437–442. [Google Scholar]
- Pavanasasivam, V.; Axley, J.H. Influence of flooding on the availability of soil zinc. Commun. Soil Sci. Plant Anal. 1980, 11, 163–174. [Google Scholar]
- Good, B.G.; Patrick, W.H. Gas composition and respiration of water oak (Quercus nigra) and green ash (Fraxinus pennsylvanica) roots after prolonged flooding. Plant Soil 1987, 97, 419–427. [Google Scholar] [CrossRef]
- McKevlin, M.R.; Hook, D.D.; McKee, W.H.; Wallace, S.U.; Woodruff, J.R. Loblolly pine seedling root anatomy and iron accumulation as affected by soil waterlogging. Can. J. For. Res. 1987, 17, 1257–1264. [Google Scholar] [CrossRef]
- Gries, C.; Kappen, L.; Losch, R. Mechanism of flood tolerance in reed (Phragmites australis). New Phytol. 1990, 114, 589–593. [Google Scholar] [CrossRef]
- Armstrong, W. Waterlogged Soils. In Environment and Plant Ecology; Etherington, J.R., Ed.; Wiley: New York, NY, USA, 1975; p. 181. [Google Scholar]
- Carlson, P.R.; Forresst, J. Uptake of dissolved sulfide by Spartina alterniflora: Evidence from natural sulfur isotope abundance ratios. Science 1982, 216, 633–635. [Google Scholar]
- Pearson, J.; Havill, D.C. The effect of hypoxia and sulfide on culture grown wetland an non-wetland plants. J. Exp. Bot. 1988, 39, 363–370. [Google Scholar] [CrossRef]
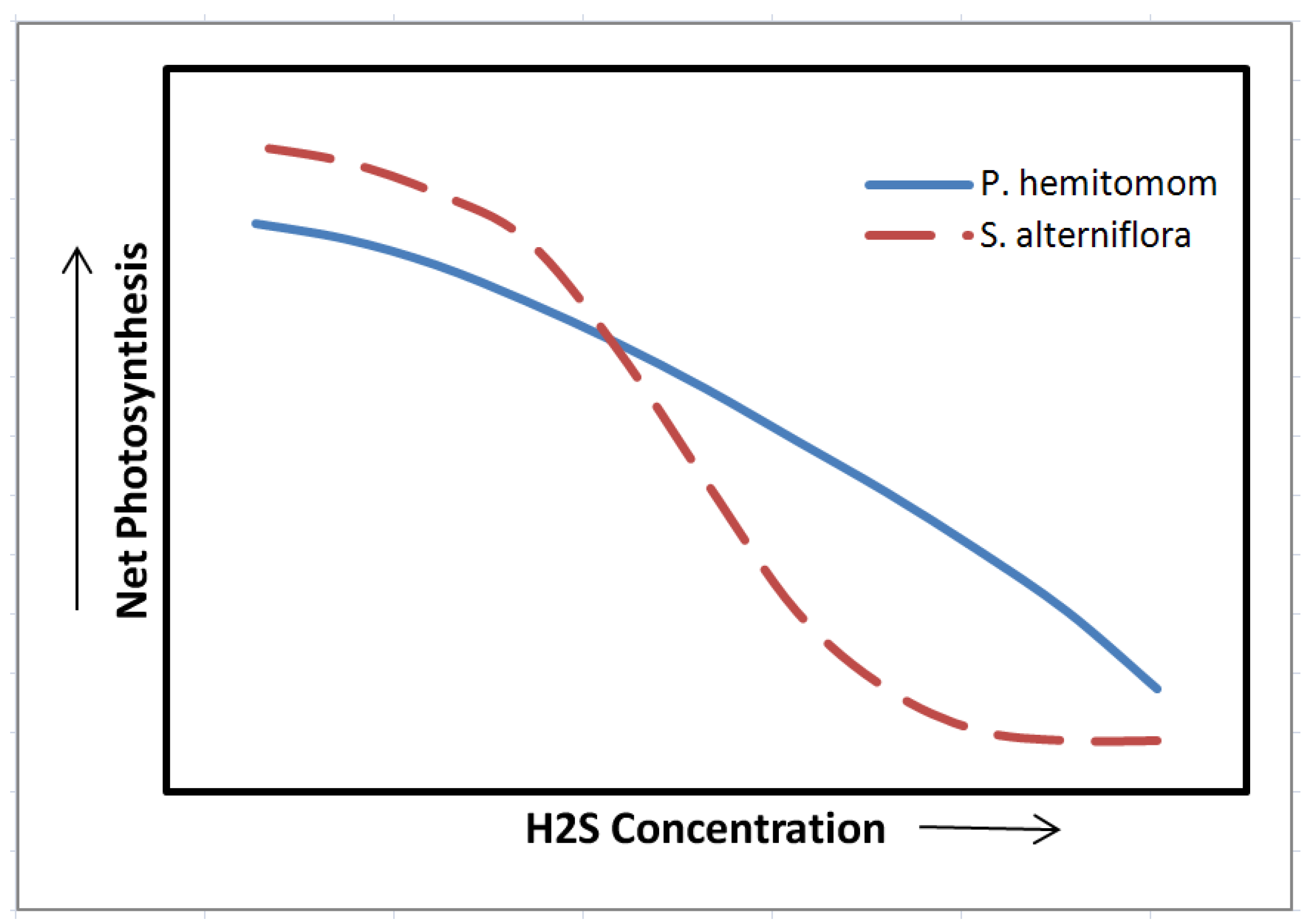
- Pezeshki, S.R.; DeLaune, R.D.; Pan, S.Z. Relationship of soil hydrogen sulfide level to net carbon assimilation of Panicum hemitomon and Spartina patens. Vegetatio 1991, 95, 159–166. [Google Scholar]
- Shimazaki, K.; Sugahara, K. Inhibition site of the electron transport system in lettuce chloroplasts by fumigation of leaves with SO2. Plant Cell Physiol. 1980, 21, 125–135. [Google Scholar]
- Wellburn, A.R.; Higginson, C.; Robinson, D.; Walmsley, C. Biochemical explanations of more than additive inhibitory effects of low atmospheric levels of sulphur dioxide plus nitrogen dioxide upon plants. New Phystol. 1981, 88, 223–237. [Google Scholar] [CrossRef]
- Garsed, S.G. The use of sulfite solution for studying the effects of SO2 on higher plants. Environ. Pollut. Ser. 1981, 24, 883–886. [Google Scholar]
- Khan, A.A.; Malhotra, S.S. Ribulose bisphosphate carboxylase and glycollate oxidase from jack pine: Effects of sulphur dioxide fumigation. Phytochemistry 1982, 21, 2607–2612. [Google Scholar]
- Dropff, M.J. Physiological effects of sulfide dioxide: 1. The effect of SO2 on photosynthesis and stomatal regulation of Vicia faba L. Plant Cell Environ. 1987, 10, 753–760. [Google Scholar]
- Furtig, K.; Ruegsegger, A.; Brunhold, C.; Brandle, R. Sulfide utilization and injuries in hypoxic roots and rhizomes of Common Reed (Phragmites australis). Folia Geobot. Phytotax. 1996, 31, 143–151. [Google Scholar] [CrossRef]
- DeLaune, R.D.; Smith, C.J.; Patrick, W.H. Relationship of marsh elevation, redox potential and sulfide to Spartina alterniflora productivity. Soil Sci. Soc. Am. J. 1983, 47, 930–935. [Google Scholar] [CrossRef]
- Everard, J.D.; Drew, M.C. Mechanisms controlling changes in water movement through the roots of Helianthus annuus L. during continuous exposure to oxygen deficiency. J. Exp. Bot. 1989, 40, 95–103. [Google Scholar] [CrossRef]
- Else, M.A.; Janowiak, F.; Atkinson, C.J.; Jackson, M.B. Root signals and stomatal closure in relation to photosynthesis, chlorophyll a fluorescence and adventitious rooting of flooded tomato plants. Ann. Bot. 2009, 103, 313–323. [Google Scholar]
- Pociecha, E.; Koscielniak, J.; Filek, W. Effects of root flooding and stage of development on the growth and photosynthesis of field bean (Vicia faba L. minor). Acta Physiol. Plant. 2008, 30, 529–535. [Google Scholar]
- Naumann, J.C.; Young, D.R.; Anderson, J.E. Leaf chlorophyll fluorescence and physiological response to freshwater and saltwater flooding in the evergreen shrub, Myrica cerifera. Environ. Exp. Bot. 2008, 63, 402–409. [Google Scholar]
- Hiron, R.W.P.; Wright, S.T.C. The role of endogenous abscisic acid in the response of plants to stress. J. Exp. Bot. 1973, 24, 769–781. [Google Scholar]
- Pereira, J.S.; Kozlowski, T.T. Variation among woody angiosperms in response to flooding. Physiol. Plant. 1977, 41, 184–192. [Google Scholar]
- Sena Gomes, A.R.; Kozlowski, T.T. Responses of Melaleuca quinquenervia seedlings to flooding. Physiol. Plant 1980, 49, 373–377. [Google Scholar] [CrossRef]
- Tang, Z.C.; Kozlowski, T.T. Some physiological and morphological responses of Quercus macrocarpa seedlings to flooding. Can. J. For. Res. 1982, 12, 196–202. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; Chambers, J.L. Stomatal and photosynthetic response of sweetgum (Liquidambar styraciflua L.) to flooding. Can. J. For. Res. 1985, 15, 371–375. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; Chambers, J.L. Response of cherrybark oak (Quercus falcata var. pagodaefolia) seedlings to short-term flooding. For. Sci. 1985, 31, 760–771. [Google Scholar]
- Pezeshki, S.R.; DeLaune, R.D.; Patrick, W.H. Differential Response of selected mangroves to soil flooding and salinity: Gas exchange and biomass partitioning. Can. J. For. Res. 1990, 20, 869–874. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; DeLaune, R.D.; Meeder, J.F. Carbon assimilation and biomass partitioning in Avicennia germinans and Rhizophora mangle seedlings in response to soil redox conditions. Environ. Exp. Bot. 1997, 37, 161–171. [Google Scholar]
- Pezeshki, S.R. Differences in patterns of photosynthetic responses to hypoxia in flood-tolerant and flood-sensitive tree species. Phtosynthetica 1993, 28, 423–430. [Google Scholar]
- Krauss, K.W.; Doyle, T.W.; Howard, R.J. Is there evidence of adaptation to tidal flooding in saplings of baldcypress subjected to different salinity regimes? Environ. Exp. Bot. 2009, 67, 118–126. [Google Scholar] [CrossRef]
- Gravatt, D.A.; Kirby, C.J. Patterns of photosynthesis and starch allocation in seedlings of four bottomland hardwood tree species subjected to flooding. Tree Physiol. 1998, 18, 411–417. [Google Scholar] [CrossRef]
- Mielke, M.S.; Shaffer, B. Leaf gas exchange, chlorophyll fluorescence and pigment indexes of Eugenia uniflora L. in response to changes in light intensity and soil flooding. Tree Physiol. 2010, 30, 45–55. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; DeLaune, R.D.; Patrick, W.H. Effect of fluctuating rhizosphere redox potential on carbon assimilation of Spartina alterniflora. Oecologia 1989, 80, 132–135. [Google Scholar]
- Brown, C.E.; Pezeshki, S.R. A study on waterlogging as a potential silvicultural tool to control Ligustrum sinense populations in western Tennessee. Wetlands 2000, 20, 429–437. [Google Scholar]
- Taylor, G.E.; Gunderson, C.A. Physiological site of ethylene effects on carbon dioxide assimilation in Glycine max. Plant Physiol. 1988, 86, 85–92. [Google Scholar] [CrossRef]
- Wample, R.L.; Thornton, R.K. Differences in the response of sunflower (Helanthus annuus) subjected to flooding and drought stress. Physiol. Plant 1984, 61, 611–616. [Google Scholar]
- Pezeshki, S.R.; Santos, M.I. Relationship among rhizosphere oxygen deficiency, root restriction, photosynthesis and growth in baldcypress (Taxodium distichum) seedlings. Photosynthetica 1998, 35, 381–390. [Google Scholar] [CrossRef]
- Ahmed, S.; Nawata, E.; Sakuratania, T. Changes of endogenous ABA and ACC and their correlation to photosynthesis and water relations in mugbean (Vigna radiate) during waterlogging. Environ. Exp. Bot. 2006, 57, 278–284. [Google Scholar]
- McKevlin, M.R.; Hook, D.D.; McKee, W.H. Growth and nutrient use efficiency of water tupelo seedlings in flooded and well drained soil. Tree Physiol. 1995, 15, 753–758. [Google Scholar]
- Bowes, G. Growth at elevated CO2: Photosynthetic responses mediated through rubisco. Plant Cell Environ. 1991, 14, 795–806. [Google Scholar] [CrossRef]
- Vu, J.C.V.; Yelenosky, G. Photosynthetic responses of rough lemon and sour orange to soil flooding, chilling and short-term temperature fluctuation during growth. Environ. Exp. Bot. 1992, 32, 471–477. [Google Scholar] [CrossRef]
- Liao, C.T.; Lin, C.H. Effect of flooding stress on photosynthetic activities of Momordica charantia. Plant Physiol. Biochem. 1994, 32, 479–485. [Google Scholar]
- Dann, M.S.; Pell, E.J. Decline of activity and quantity of ribulose bisphosphate carboxylase/owygenase and net photosynthesis in ozone-treated potato foliage. Plant Physiol. 1989, 91, 427–432. [Google Scholar] [CrossRef]
- Lauer, M.J.; Pallardy, S.G.; Belvins, D.C.; Randall, D.D. Whole leaf carbon exchange characteristics of phosphate deficient soybeans (Glycine max L.). Plant Physiol. 1989, 91, 848–854. [Google Scholar] [CrossRef]
- Li, S.; Goodwin, S.; Pezeshki, S.R. Photosynthetic gene expression in black willow under various soil moisture regimes. Biol. Plant 2007, 51, 593–596. [Google Scholar] [CrossRef]
- Sij, J.W.; Swanson, C.A. Effect of petiole anoxia on phloem transport in squash. Plant Physiol. 1973, 51, 368–371. [Google Scholar] [CrossRef]
- Qureshi, F.A.; Spanner, D.C. The effect of nitrogen on the movement of tracers down the stolon of Saxifraga sarmentosa with some observations on the influence of light. Planta 1973, 110, 131–144. [Google Scholar] [CrossRef]
- Vartapetian, B.B. Flood-Sensitive Plants under Primary and Secondary Anoxia: Ultrastructural and Metabolic Responses. In Plant Life Under Oxygen Deprivation; Jackson, M.B., Davies, D.D., Lambers, H., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1991; pp. 201–216. [Google Scholar]
- Kennedy, R.A.; Fox, T.C.; Everard, J.D.; Rumpho, M.E. Biochemical Adaptations to Anoxia: Potential Role of Mitochondrial Metabolism to Flood Tolerance in Echinochloa Phyllopogon (Barnyard Grass). In Plant Life Under Oxygen Deprivation; Jackson, M.B., Davies, D.D., Lambers, H., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1991; pp. 217–227. [Google Scholar]
- Yamamoto, F.; Kozlowski, T.T. Effect of flooding, tilting of stem, and ethrel application on growth, stem anatomy, and ethylene production of Acer platanoides seedlings. Scand. J. For. Res. 1987, 2, 141–156. [Google Scholar] [CrossRef]
- Yamamoto, F.; Sakata, T.; Terazawa, K. Growth, morphology, stem anatomy and ethylene production in flooded Alnus japonica seedlings. IAWA J. 1995, 16, 47–59. [Google Scholar]
- Yamamoto, F.; Sakata, T.; Terazawa, K. Physiological, anatomical and morphological responses of Fraxinus mandshurica seedlings to flooding. Tree Physiol. 1995, 15, 713–719. [Google Scholar]
- Webb, T.; Armstrong, W. The effects of anoxia and carbohydrates on the growth and viability of rice, pea and pumpkin roots. J. Exp. Bot. 1983, 34, 579–603. [Google Scholar] [CrossRef]
- Pezeshki, S.R. Root responses of flood-tolerant and flood-sensitive tree species to soil redox conditions. Trees 1991, 5, 180–186. [Google Scholar]
- Pezeshki, S.R.; DeLaune, R.D. Influence of sediment oxidation-reduction potential on root elongation in Spartina patens. Acta Oecologia 1990, 11, 377–383. [Google Scholar]
- Bertani, A.; Brambilla, I. Effect of decreasing oxygen concentration on wheat roots: Growth and induction of anaerobic metabolism. Z. Pflanzenphysiol. 1982, 108, 283–288. [Google Scholar]
- Lissner, J.; Mendelssohn, I.A.; Anastasiou, C.J. A method for cultivating plants under controlled redox intensities in hydroponics. Aquat. Bot. 2003, 76, 93–108. [Google Scholar] [CrossRef]
- Will, R.E.; Seiler, J.R.; Feret, P.P.; Aust, W.M. Effects of rhizosphere inundation on the growth and physiology of wet and dry -site Acer rubrum (red maple) populations. Am. Midl. Nat. 1995, 134, 127–139. [Google Scholar] [CrossRef]
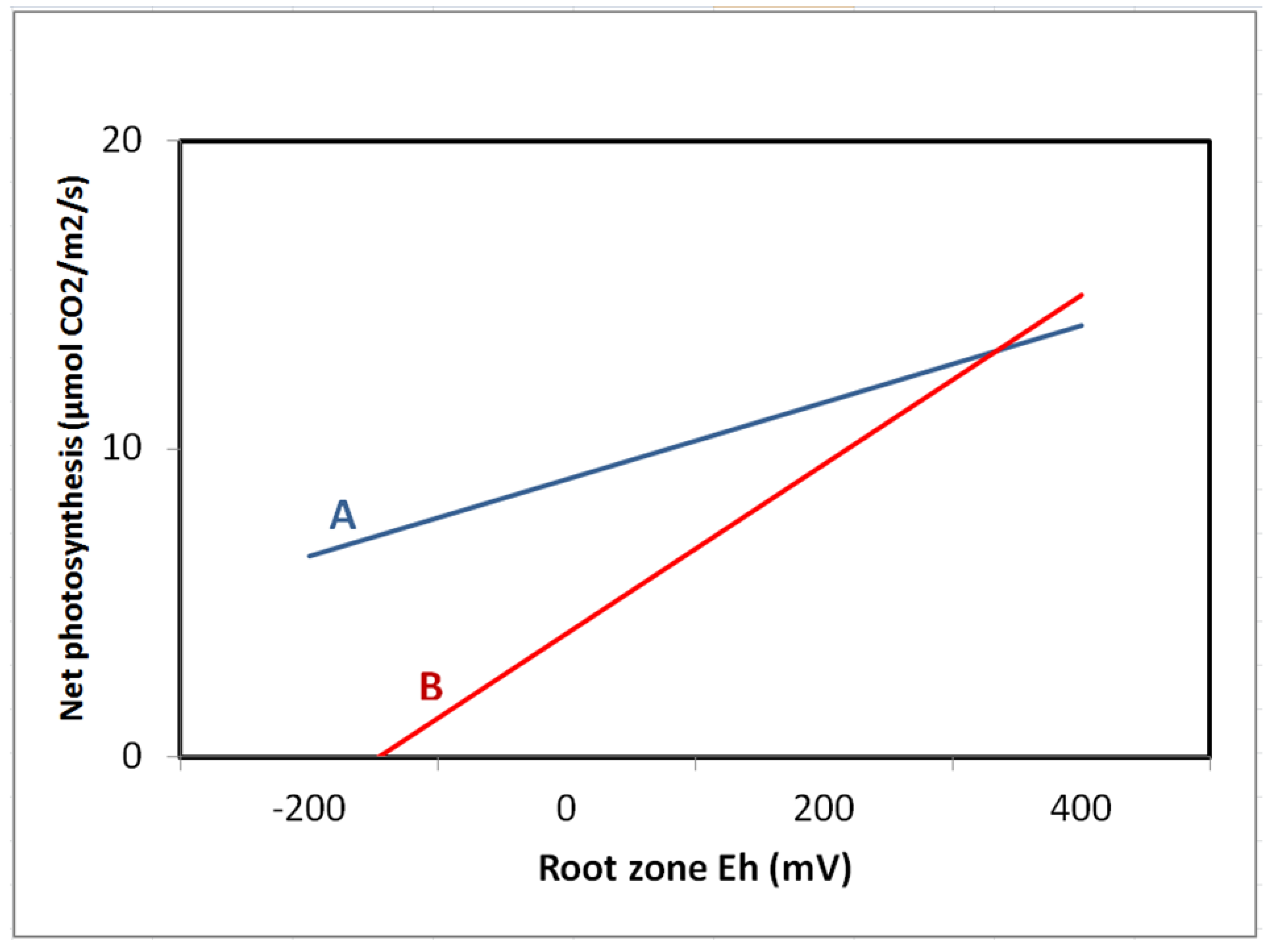
- Pezeshki, S.R.; DeLaune, R.D.; Kludze, H.K.; Choi, H.S. A comparative study of gas exchange characteristics of cattail (Typha domingensis) and sawgrass (Cladium jamaicense) to soil redox conditions. Aquat. Bot. 1996, 54, 25–35. [Google Scholar]
- Pezeshki, S.R.; DeLaune, R.D. Responses of seedlings of selected woody species to soil oxidation-reduction conditions. Environ. Exp. Bot. 1998, 40, 123–133. [Google Scholar] [CrossRef]
- Anderson, P.H.; Pezeshki, S.R. The effects of intermittent flooding on seedlings of three forest species. Photosynthetica 1999, 37, 543–552. [Google Scholar]
- Elcan, J.M.; Pezeshki, S.R. Effects of flooding on susceptibility of Taxodium distichum seedlings to drought. Photosynthetica 2002, 40, 177–182. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pezeshki, S.R.; DeLaune, R.D. Soil Oxidation-Reduction in Wetlands and Its Impact on Plant Functioning. Biology 2012, 1, 196-221. https://doi.org/10.3390/biology1020196
Pezeshki SR, DeLaune RD. Soil Oxidation-Reduction in Wetlands and Its Impact on Plant Functioning. Biology. 2012; 1(2):196-221. https://doi.org/10.3390/biology1020196
Chicago/Turabian StylePezeshki, S. R., and R. D. DeLaune. 2012. "Soil Oxidation-Reduction in Wetlands and Its Impact on Plant Functioning" Biology 1, no. 2: 196-221. https://doi.org/10.3390/biology1020196
APA StylePezeshki, S. R., & DeLaune, R. D. (2012). Soil Oxidation-Reduction in Wetlands and Its Impact on Plant Functioning. Biology, 1(2), 196-221. https://doi.org/10.3390/biology1020196