Antibiotic Resistance of Gram-Negative Bacteria from Wild Captured Loggerhead Sea Turtles


 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Loggerhead Turtles’ Size and Health Status
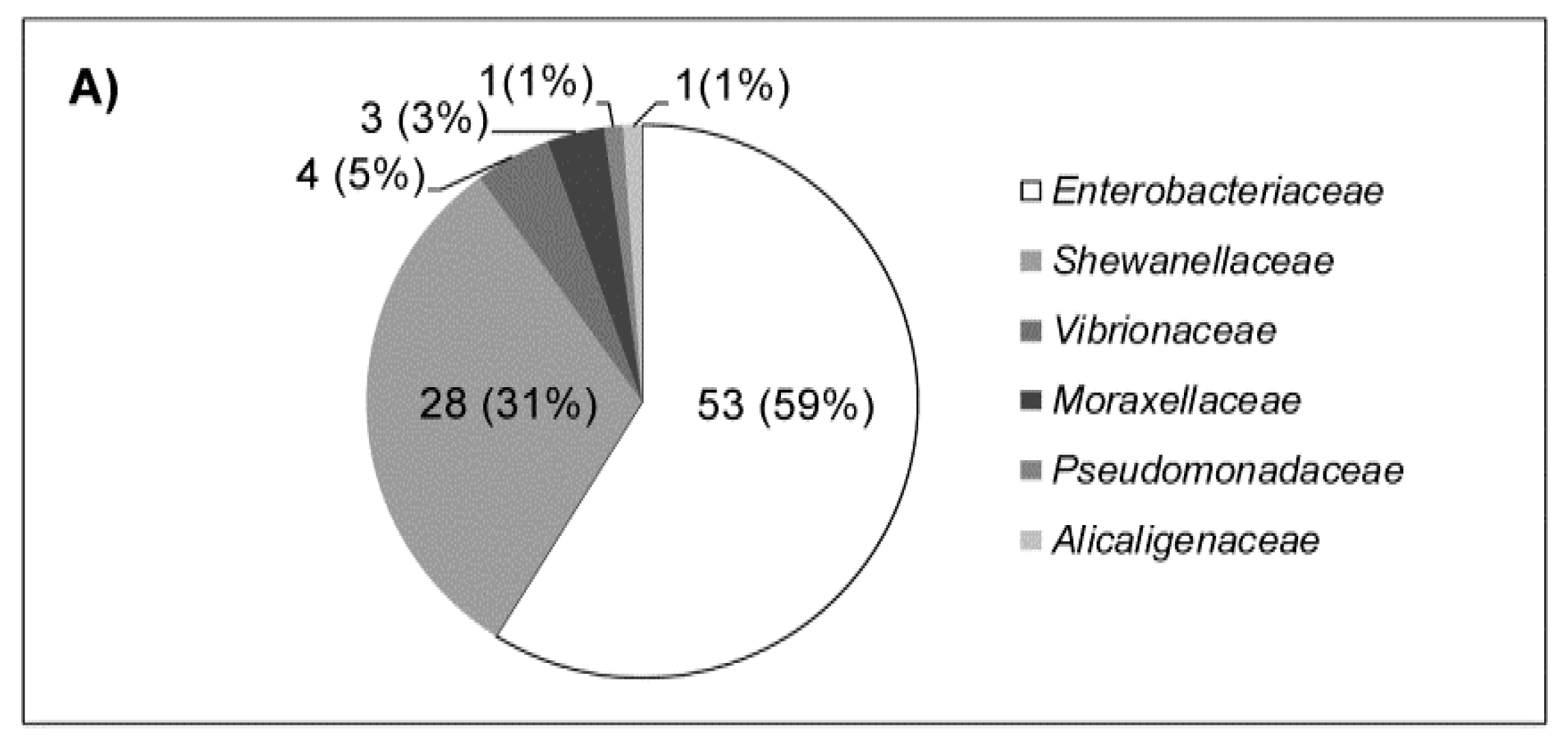
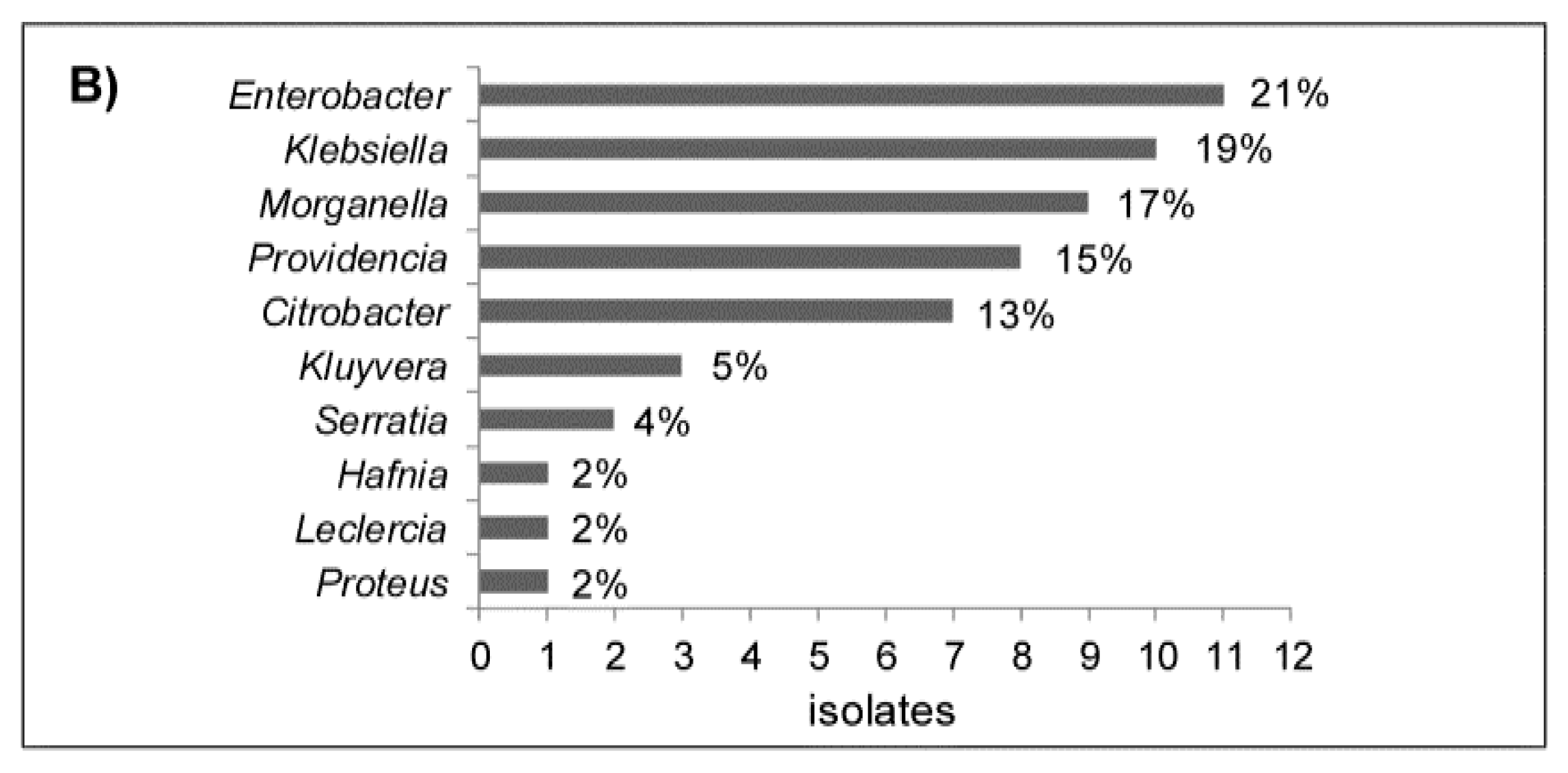
2.2. Cloaca Associated Bacteria
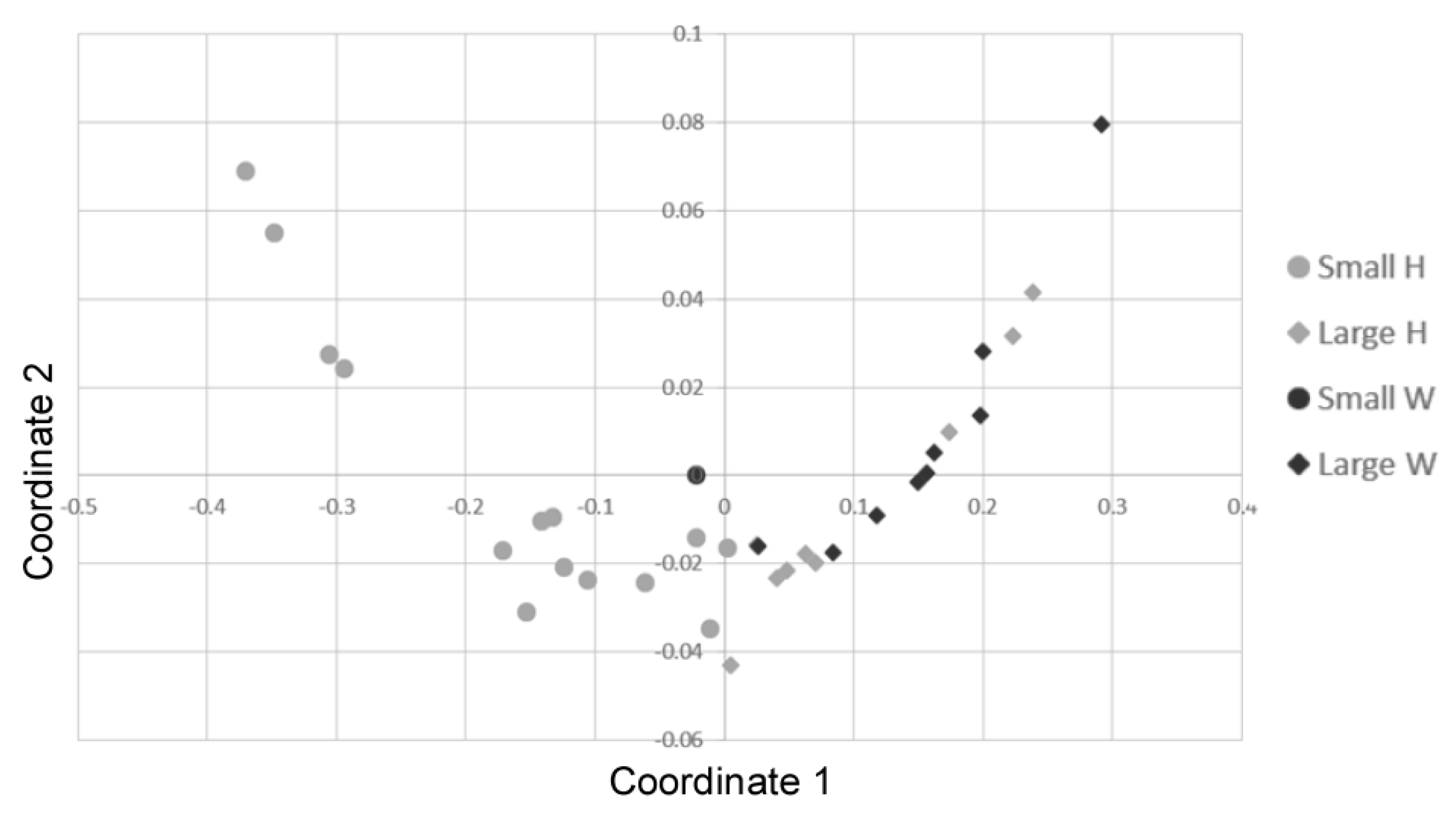
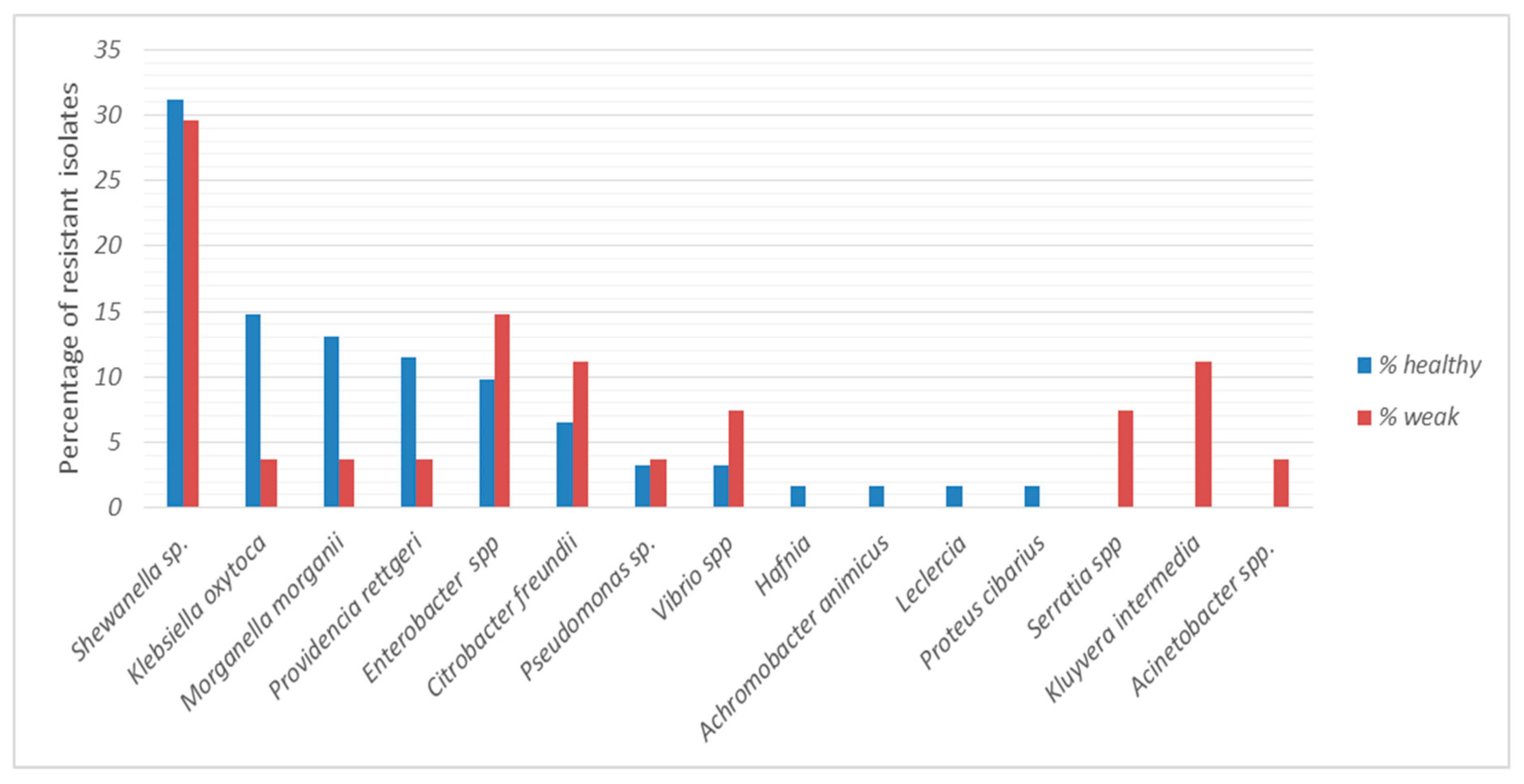
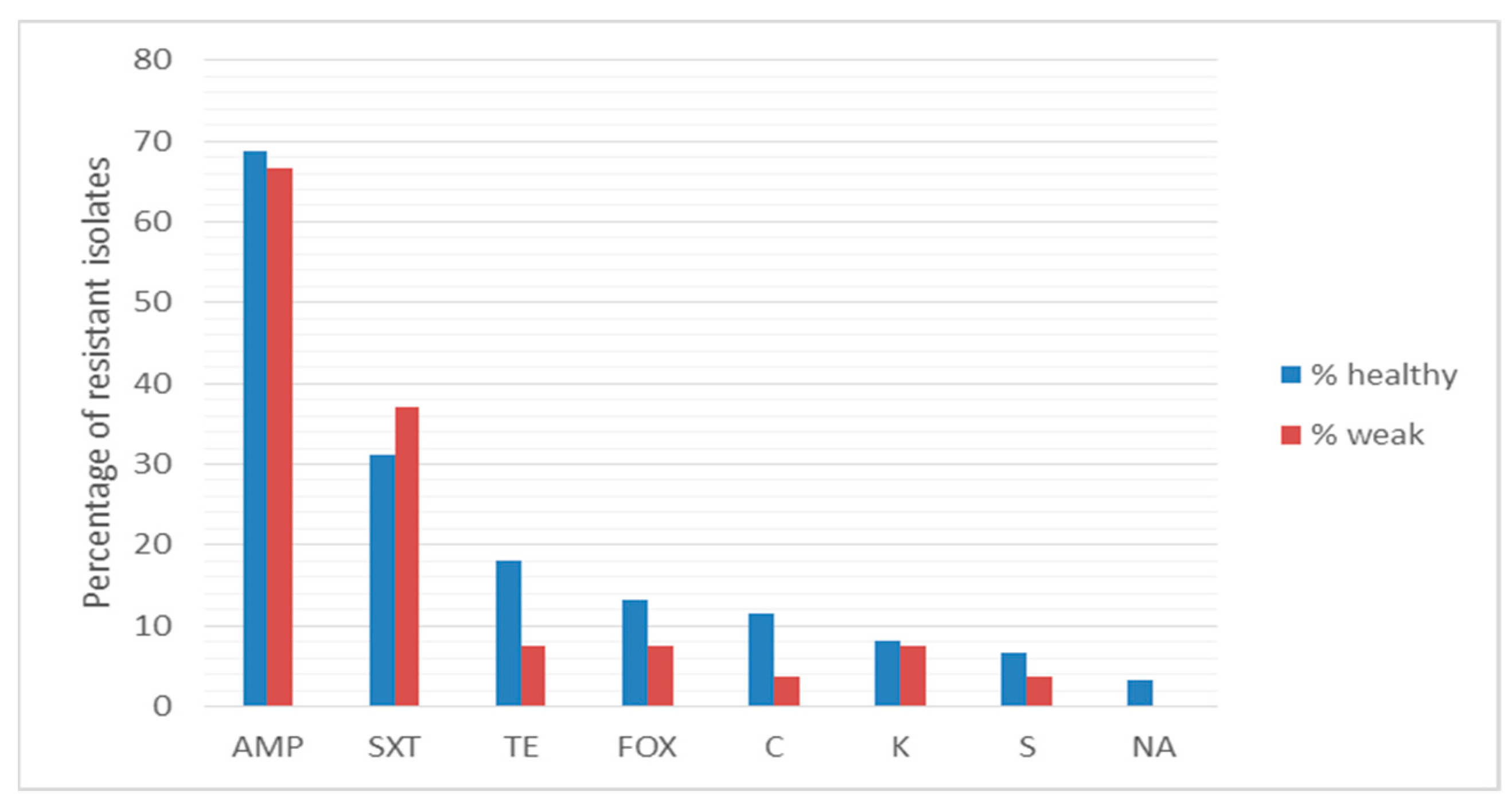
2.3. Antibiotic Resistance
3. Discussion
4. Materials and Methods
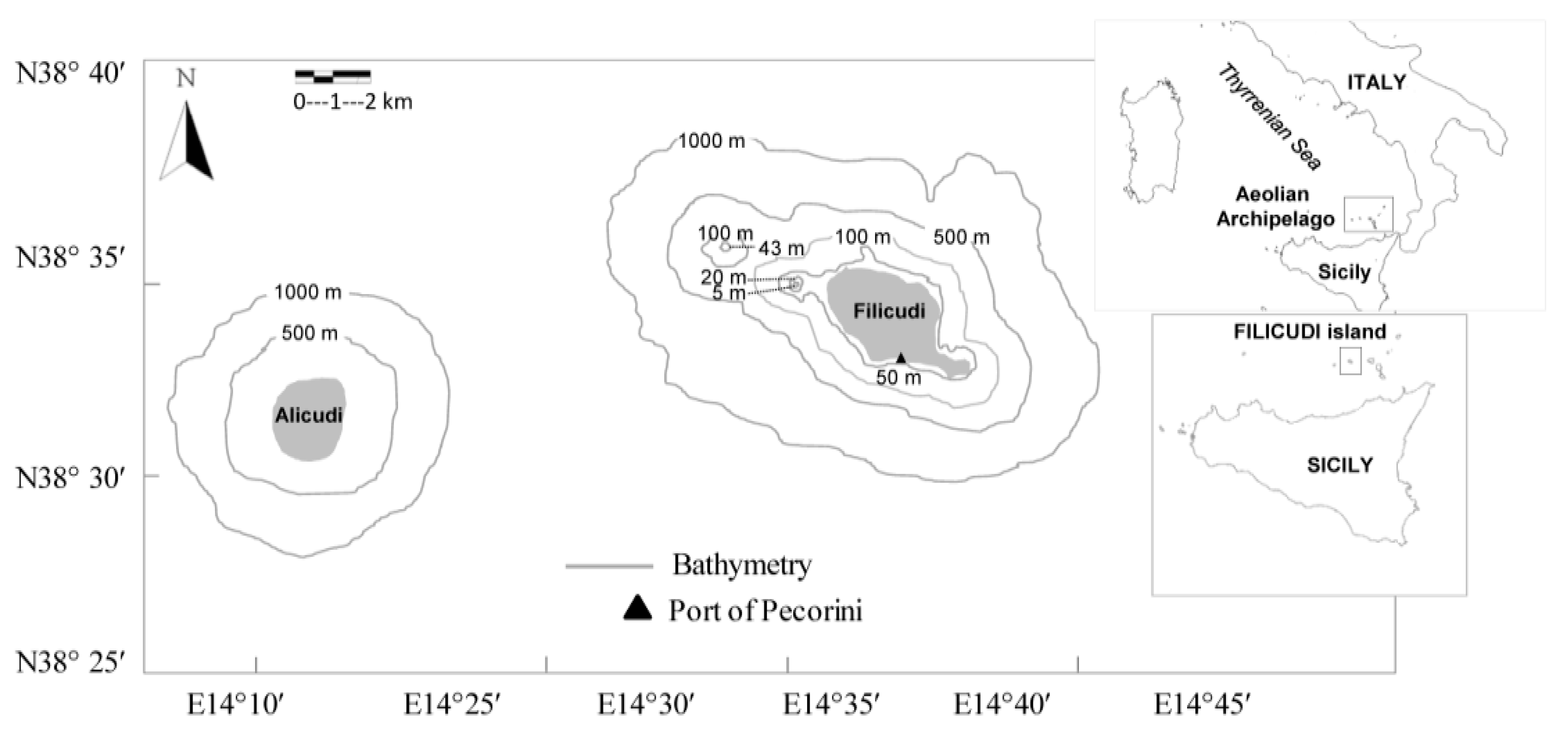
4.1. Sample Collection
4.2. Cloaca Associated Bacteria Analysis
4.3. Antibiotic Susceptibility Test
4.4. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vitale, M.; Gaglio, S.; Galluzzo, P.; Cascone, G.; Piraino, C.; Lo Presti, V.D.; Alduina, R. Antibiotic resistance profiling, analysis of virulence aspects Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic pollution in the environment: From microbial ecology to public policy. Microorganisms 2019, 7, 180. [Google Scholar]
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4, 481–511. [Google Scholar] [CrossRef]
- Carroll, D.; Wang, J.; Fanning, S.; McMahon, B.J. Antimicrobial resistance in wildlife: Implications for public health. Zoonoses Public Health 2015, 62, 534–542. [Google Scholar] [CrossRef]
- Hatosy, S.M.; Martiny, A.C. The ocean as a global reservoir of antibiotic resistance genes. Appl. Environ. Microbiol. 2015, 81, 7593–7599. [Google Scholar] [CrossRef]
- Al-Bahry, S.N.; Mahmoud, I.Y.; Al-Zadjali, M.; Elshafie, A.; Al-Harthy, A.; Al-Alawi, W. Antibiotic resistant bacteria as bio-indicator of polluted effluent in the green turtles, Chelonia mydas in Oman. Mar. Environ. Res. 2011, 71, 139–144. [Google Scholar] [CrossRef]
- Al-Bahry, S.; Mahmoud, I.; Elshafie, A.; Al-Harthy, A.; Al-Ghafri, S.; Al-Amri, I.; Alkindi, A. Bacterial flora and antibiotic resistance from eggs of green turtles Chelonia mydas: An indication of polluted effluents. Mar. Pollut. Bull. 2009, 58, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Barros, J.; Igrejas, G.; Andrade, M.; Radhouani, H.; Lopez, M.; Torres, C.; Poeta, P. Gilthead seabream (Sparus aurata) carrying antibiotic resistant enterococci. A potential bioindicator of marine contamination? Mar. Pollut. Bull. 2011, 62, 1245–1248. [Google Scholar] [CrossRef] [PubMed]
- Prichula, J.; Pereira, R.I.; Wachholz, G.R.; Cardoso, L.A.; Tolfo, N.C.; Santestevan, N.A.; Medeiros, A.W.; Tavares, M.; Frazzon, J.; d’Azevedo, P.A.; et al. Resistance to antimicrobial agents among enterococci isolated from fecal samples of wild marine species in the southern coast of Brazil. Mar. Pollut. Bull. 2016, 105, 51–57. [Google Scholar] [CrossRef]
- Foti, M.; Giacopello, C.; Bottari, T.; Fisichella, V.; Rinaldo, D.; Mammina, C. Antibiotic resistance of gram negatives isolates from loggerhead sea turtles (Caretta caretta) in the central Mediterranean Sea. Mar. Pollut. Bull. 2009, 58, 1363–1366. [Google Scholar] [CrossRef] [PubMed]
- Pace, A.; Dipineto, L.; Fioretti, A.; Hochscheid, S. Loggerhead sea turtles as sentinels in the western mediterranean: Antibiotic resistance and environment-related modifications of gram-negative bacteria. Mar. Pollut. Bull. 2019, 149, 110575. [Google Scholar] [CrossRef]
- Thaller, S. Introduction to war-related injuries. J. Craniofacial Surg. 2010, 21, 952–953. [Google Scholar] [CrossRef]
- Thaller, M.C.; Migliore, L.; Marquez, C.; Tapia, W.; Cedeno, V.; Rossolini, G.M.; Gentile, G. Tracking acquired antibiotic resistance in commensal bacteria of galapagos land iguanas: No man, no resistance. PLoS ONE 2010, 5, e8989. [Google Scholar] [CrossRef]
- Alduina, R.; Gambino, D.; Presentato, A.; Gentile, A.; Sucato, A.; Savoca, D.; Filippello, S.; Visconti, G.; Caracappa, G.; Vicari, D.; et al. Is Caretta caretta a carrier of antibiotic resistance in the Mediterranean Sea? Antibiot. -Basel 2020, 9, 116. [Google Scholar] [CrossRef] [PubMed]
- Broderick, A.C.; Coyne, M.S.; Fuller, W.J.; Glen, F.; Godley, B.J. Fidelity and over-wintering of sea turtles. Proc. R. Soc. B Biol. Sci. 2007, 274, 1533–1538. [Google Scholar] [CrossRef] [PubMed]
- Casale, P.; Abbate, G.; Freggi, D.; Conte, N.; Oliverio, M.; Argano, R. Foraging ecology of loggerhead sea turtles Caretta caretta in the central mediterranean sea: Evidence for a relaxed life history model. Mar. Ecol. Prog. Ser. 2008, 372, 265–276. [Google Scholar] [CrossRef]
- Oros, J.; Torrent, A.; Calabuig, P.; Deniz, S. Diseases and causes of mortality among sea turtles stranded in the canary islands, spain (1998-2001). Dis. Aquat. Org. 2005, 63, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Bjorndal, K.A. Foraging ecology and nutrition of sea turtles. Biol. Sea Turt. 1997, 1, 32. [Google Scholar]
- Tomas, J.; Guitart, R.; Mateo, R.; Raga, J.A. Marine debris ingestion in loggerhead sea turtles, caretta caretta, from the western mediterranean. Mar. Pollut. Bull. 2002, 44, 211–216. [Google Scholar] [CrossRef]
- Casale, P.; Cattarino, L.; Freggi, D.; Rocco, M.; Argano, R. Incidental catch of marine turtles by italian trawlers and longliners in the central Mediterranean. Aquat. Conserv. 2007, 17, 686–701. [Google Scholar] [CrossRef]
- Casale, P. Sea turtle by-catch in the mediterranean. Fish Fish. 2011, 12, 299–316. [Google Scholar] [CrossRef]
- Campani, T.; Baini, M.; Giannetti, M.; Cancelli, F.; Mancusi, C.; Serena, F.; Marsili, L.; Casini, S.; Fossi, M.C. Presence of plastic debris in loggerhead turtle stranded along the Tuscany coasts of the pelagos sanctuary for Mediterranean marine mammals (italy). Mar. Pollut. Bull. 2013, 74, 225–230. [Google Scholar] [CrossRef]
- Caracappa, S.; Persichetti, M.F.; Piazza, A.; Caracappa, G.; Gentile, A.; Marineo, S.; Crucitti, D.; Arculeo, M. Incidental catch of loggerhead sea turtles (Caretta caretta) along the Sicilian coasts by longline fishery. PeerJ 2018, 6, e5392. [Google Scholar] [CrossRef]
- Caracappa, S.; Pisciotta, A.; Persichetti, M.F.; Caracappa, G.; Alduina, R.; Arculeo, M. Nonmodal scutes patterns in the loggerhead sea turtle (Caretta caretta): A possible epigenetic effect? Can. J. Zool. 2016, 94, 379–383. [Google Scholar] [CrossRef]
- Favalli, M.; Karatson, D.; Mazzuoli, R.; Pareschi, M.T.; Ventura, G. Volcanic geomorphology and tectonics of the Aeolian archipelago (Southern Italy) based on integrated dem data. B Volcanol. 2005, 68, 157–170. [Google Scholar] [CrossRef]
- Blasi, M.F.; Roscioni, F.; Mattei, D. Interaction of loggerhead turtles (Caretta caretta) with traditional fish aggregating devices (fads) in the Mediterranean Sea. Herpetol. Conserv. Biol. 2016, 11, 386–401. [Google Scholar]
- Blasi, M.F.; Mattei, D. Seasonal encounter rate, life stages and main threats to the loggerhead sea turtle (Caretta caretta) in the Aeolian archipelago (Southern Thyrrenian Sea). Aquat. Conserv. 2017, 27, 617–630. [Google Scholar] [CrossRef]
- Fichi, G.; Cardeti, G.; Cersini, A.; Mancusi, C.; Guarducci, M.; Di Guardo, G.; Terracciano, G. Bacterial and viral pathogens detected in sea turtles stranded along the coast of Tuscany, Italy. Vet. Microbiol. 2016, 185, 56–61. [Google Scholar] [CrossRef]
- Pace, A.; Rinaldi, L.; Ianniello, D.; Borrelli, L.; Cringoli, G.; Fioretti, A.; Hochscheid, S.; Dipineto, L. Gastrointestinal investigation of parasites and enterobacteriaceae in loggerhead sea turtles from Italian coasts. BMC Vet. Res. 2019, 15, 370. [Google Scholar] [CrossRef] [PubMed]
- Gambino, D.; Persichetti, M.F.; Gentile, A.; Arculeo, M.; Visconti, G.; Curro, V.; Caracappa, G.; Crucitti, D.; Piazza, A.; Mancianti, F.; et al. First data on microflora of loggerhead sea turtle (Caretta caretta) nests from the coastlines of Sicily. Biol. Open 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Abdelrhman, K.F.A.; Bacci, G.; Mancusi, C.; Mengoni, A.; Serena, F.; Ugolini, A. A first insight into the gut microbiota of the sea turtle Caretta caretta. Front. Microbiol. 2016, 7, 1060. [Google Scholar] [CrossRef] [PubMed]
- Biagi, E.; D’Amico, F.; Soverini, M.; Angelini, V.; Barone, M.; Turroni, S.; Rampelli, S.; Pari, S.; Brigidi, P.; Candela, M. Faecal bacterial communities from mediterranean loggerhead sea turtles (Caretta caretta). Environ. Microbiol. Rep. 2019, 11, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Arizza, V.; Vecchioni, L.; Caracappa, S.; Sciurba, G.; Berlinghieri, F.; Gentile, A.; Persichetti, M.F.; Arculeo, M.; Alduina, R. New insights into the gut microbiome in loggerhead sea turtles Caretta caretta stranded on the mediterranean coast. PLoS ONE 2019, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Bolten, A.B. Techniques for measuring sea turtles. In Research and Management Techniques for the Conservation of Sea Turtles; IUCN/SSC Marine Turtle Specialist Group Publication No. 4; Eckert, K.L., Bjorndal, K.A., Abreu-Grobois, F.A., Donnelly, M., Eds.; IUCN/SSC Marine Turtle Specialist Group: Washington, DC, USA, 1999; pp. 120–125. [Google Scholar]
- Hyun, D.W.; Jung, M.J.; Kim, M.S.; Shin, N.R.; Kim, P.S.; Whon, T.W.; Bae, J.W. Proteus cibarius sp nov., a swarming bacterium from jeotgal, a traditional korean fermented seafood, and emended description of the genus Proteus. Int. J. Syst. Evol. Micr. 2016, 66, 2158–2164. [Google Scholar] [CrossRef] [PubMed]
- Migliore, L.; Alessi, E.; Busani, L.; Caprioli, A. Effects of the use of Flumequine in aquaculture: microbial resistance and sediment contamination. Fresenius Environ. Bull. 2002, 11, 557–561. [Google Scholar]
- Di Ianni, F.; Dodi, P.L.; Cabassi, C.S.; Pelizzone, I.; Sala, A.; Cavirani, S.; Parmigiani, E.; Quintavalla, F.; Taddei, S. Conjunctival flora of clinically normal and diseased turtles and tortoises. Bmc Vet. Res. 2015, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- George, R.H. Health problems and diseases of sea turtles. In The Biology of Sea Turtles; CRC Press: Boca Raton, FL, USA, 2017; Volume 1, pp. 363–385. [Google Scholar]
- Vega-Manriquez, D.X.; Davila-Arrellano, R.P.; Eslava-Campos, C.A.; Jimenez, E.S.; Negrete-Philippe, A.C.; Raigoza-Figueras, R.; Munoz-Teneria, F.A. Identification of bacteria present in ulcerative stomatitis lesions of captive sea turtles Chelonia mydas. Vet. Res. Commun. 2018, 42, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.; Hernandez, G.; Caballero, M.; Garcia, F. Aerobic bacterial flora of nesting green turtles (Chelonia mydas) from Tortuguero national park, Costa Rica. J. Zoo Wildl. Med. 2006, 37, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Ferronato, B.O.; Genoy-Puerto, A.; Pina, C.I.; Souza, F.L.; Verdade, L.M.; Matushima, E.R. Notes on the hematology of free-living phrynops geoffroanus (testudines: Chelidae) in polluted rivers of southeastern Brazil. Zool. -Curitiba 2009, 26, 795–798. [Google Scholar] [CrossRef]
- dos Santos, G.S.; Solidônio, E.G.; Costa, M.C.V.V.; Melo, R.O.A.; de Souza, I.F.A.C.; Silva, G.R.; Sena, K.X.F.R. Study of the enterobacteriaceae group CESP (Citrobacter, Enterobacter, Serratia, Providencia, Morganella and Hafnia): A review. In The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs; Méndez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2015; pp. 794–805. [Google Scholar]
- Wallace, C.C.; Yund, P.O.; Ford, T.E.; Matassa, K.A.; Bass, A.L. Increase in antimicrobial resistance in bacteria isolated from stranded marine mammals of the northwest atlantic. EcoHealth 2013, 10, 201–210. [Google Scholar] [CrossRef]
- Aguirre, A.A.; Balazs, G.H.; Zimmerman, B.; Spraker, T.R. Evaluation of hawaiian green turtles (Chelonia-mydas) for potential pathogens associated with fibropapillomas. J. Wildl. Dis. 1994, 30, 8–15. [Google Scholar] [CrossRef]
- Al-Bahry, S.N.; Al-Zadjali, M.A.; Mahmoud, I.Y.; Elshafie, A.E. Biomonitoring marine habitats in reference to antibiotic resistant bacteria and ampicillin resistance determinants from oviductal fluid of the nesting green sea turtle, Chelonia-mydas. Chemosphere 2012, 87, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Oros, J.; Calabuig, P.; Deniz, S. Digestive pathology of sea turtles stranded in the canary islands between 1993 and 2001. Vet. Rec. 2004, 155, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Pazdzior, E.; Pekala-Safinska, A.; Wasyl, D. Genotypic diversity among Shewanella spp. collected from freshwater fish. J. Fish Dis. 2019, 42, 677–684. [Google Scholar] [CrossRef]
- Goldschmidt-Clermont, E.; Wahli, T.; Frey, J.; Burr, S.E. Identification of bacteria from the normal flora of perch, Perca fluviatilis l., and evaluation of their inhibitory potential towards Aeromonas species. J. Fish Dis. 2008, 31, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Onishchenko, O.M.; Kiprianova, E.A. Shewanella genus bacteria isolated from the black sea water and molluscs. Mikrobiolohichnyi Zhurnal 2006, 68, 12–21. [Google Scholar] [PubMed]
- Blasi, M.F.; Tomassini, L.; Gelippi, M.; Careddu, G.; Insacco, G.; Polunin, N.V.C. Assessing resource use patterns of Mediterranean loggerhead sea turtles Caretta caretta (Linnaeus, 1758) through stable isotope analysis. Eur. Zool. J. 2018, 85, 72–88. [Google Scholar] [CrossRef]
- Zbinden, J.A.; Bearhop, S.; Bradshaw, P.; Gill, B.; Margaritoulis, D.; Newton, J.; Godley, B.J. Migratory dichotomy and associated phenotypic variation in marine turtles revealed by satellite tracking and stable isotope analysis. Mar. Ecol. Prog. Ser. 2011, 421, 291–302. [Google Scholar] [CrossRef]
- Casale, P.; Mazaris, A.D.; Freggi, D.; Vallini, C.; Argano, R. Growth rates and age at adult size of loggerhead sea turtles (Caretta caretta) in the Mediterranean sea, estimated through capture-mark-recapture records. Sci. Mar. 2009, 73, 589–595. [Google Scholar] [CrossRef]
- Bartoloni, A.; Benedetti, M.; Pallecchi, L.; Larsson, M.; Mantella, A.; Strohmeyer, M.; Bartalesi, F.; Fernandez, C.; Guzman, E.; Vallejos, Y.; et al. Evaluation of a rapid screening method for detection of antimicrobial resistance in the commensal microbiota of the gut. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 119–125. [Google Scholar] [CrossRef]
- Evangelisti, M.; D’Amelia, D.; Di Lallo, G.; Thaller, M.C.; Migliore, L. The relationship between salinity and bacterioplankton in three relic coastal ponds (Macchiatonda Wetland, Italy). J. Water Res. Prot. 2013, 5, 859–866. [Google Scholar] [CrossRef][Green Version]
- Schwieger, F.; Tebbe, C.C. A new approach to utilize PCR-single-strand-conformation polymorphism for 16S rRNA gene-based microbial community analysis. Appl. Environ. Microbiol. 1998, 64, 4870–4876. [Google Scholar] [CrossRef] [PubMed]
- Schmalenberger, A.; Schwieger, F.; Tebbe, C.C. Effect of primers hybridizing to different evolutionarily conserved regions of the small-subunit rRNA gene in PCR-based microbial community analyses and genetic profiling. Appl. Environ. Microbiol. 2000, 67, 3557–3563. [Google Scholar] [CrossRef] [PubMed]
- Vitale, M.; Gaglio, S.; Galluzzo, P.; Cascone, G.; Piraino, C.; Lo Presti, V.D.; Alduina, R. Antibiotic resistance profiling, analysis of virulence aspects and molecular genotyping of staphylococcus aureus isolated in sicily, Italy. Foodborne Pathog. Dis. 2018, 15, 177–185. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blasi, M.F.; Migliore, L.; Mattei, D.; Rotini, A.; Thaller, M.C.; Alduina, R. Antibiotic Resistance of Gram-Negative Bacteria from Wild Captured Loggerhead Sea Turtles. Antibiotics 2020, 9, 162. https://doi.org/10.3390/antibiotics9040162
Blasi MF, Migliore L, Mattei D, Rotini A, Thaller MC, Alduina R. Antibiotic Resistance of Gram-Negative Bacteria from Wild Captured Loggerhead Sea Turtles. Antibiotics. 2020; 9(4):162. https://doi.org/10.3390/antibiotics9040162
Chicago/Turabian StyleBlasi, Monica Francesca, Luciana Migliore, Daniela Mattei, Alice Rotini, Maria Cristina Thaller, and Rosa Alduina. 2020. "Antibiotic Resistance of Gram-Negative Bacteria from Wild Captured Loggerhead Sea Turtles" Antibiotics 9, no. 4: 162. https://doi.org/10.3390/antibiotics9040162
APA StyleBlasi, M. F., Migliore, L., Mattei, D., Rotini, A., Thaller, M. C., & Alduina, R. (2020). Antibiotic Resistance of Gram-Negative Bacteria from Wild Captured Loggerhead Sea Turtles. Antibiotics, 9(4), 162. https://doi.org/10.3390/antibiotics9040162