Effects of Labrador Tea, Peppermint, and Winter Savory Essential Oils on Fusobacterium nucleatum


Abstract
1. Introduction
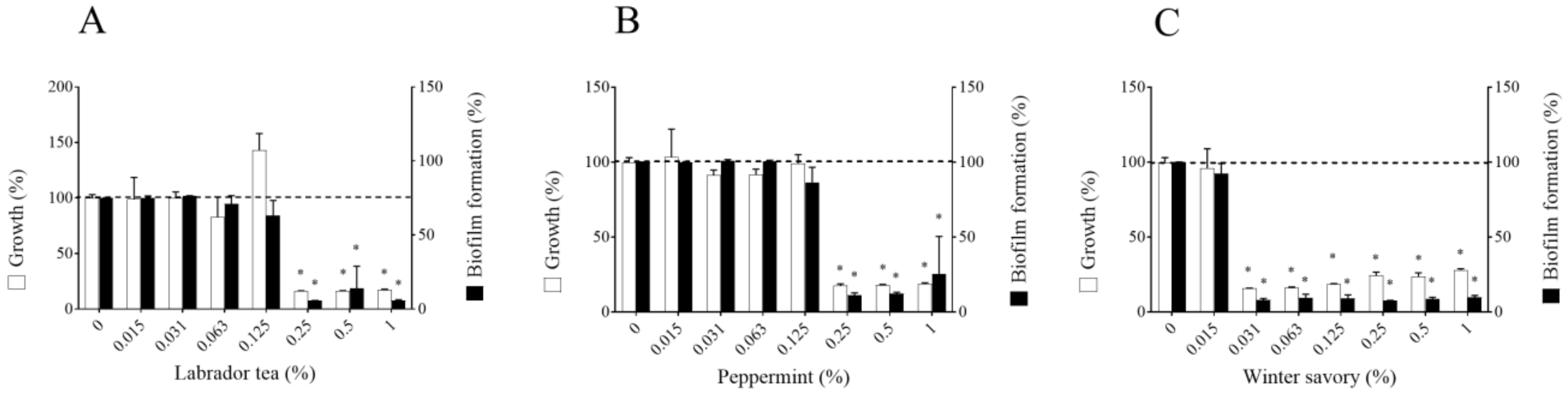
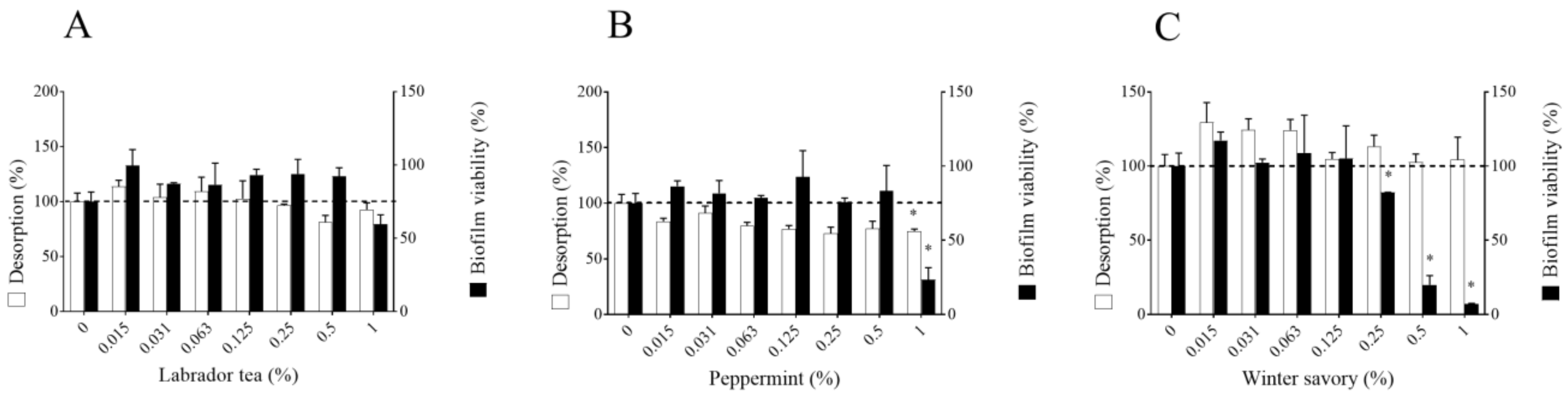
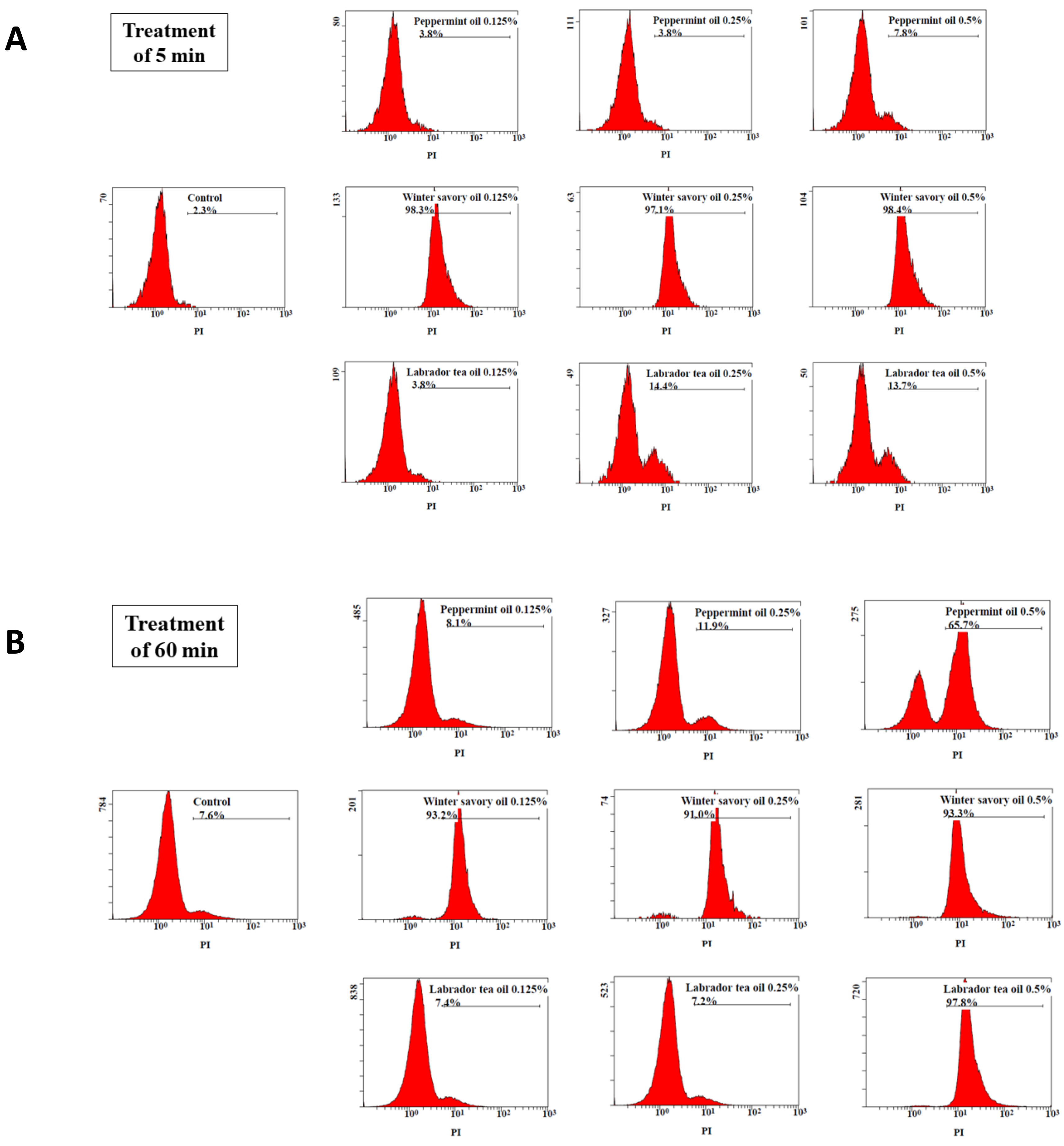
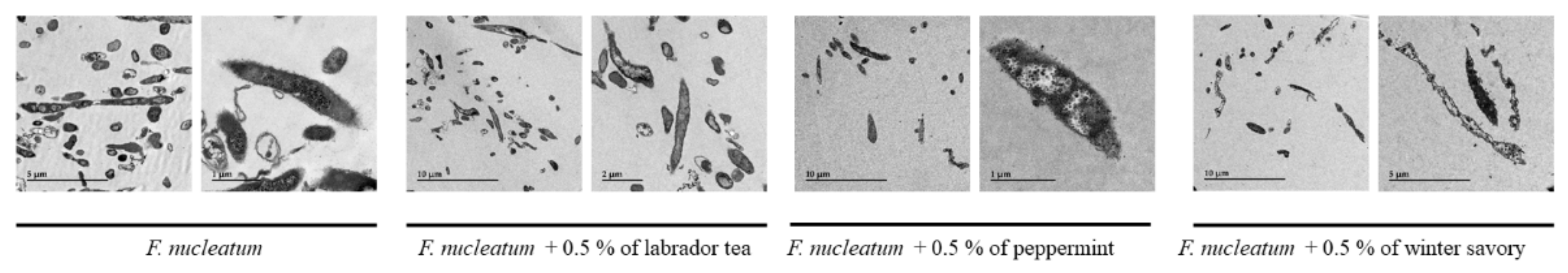
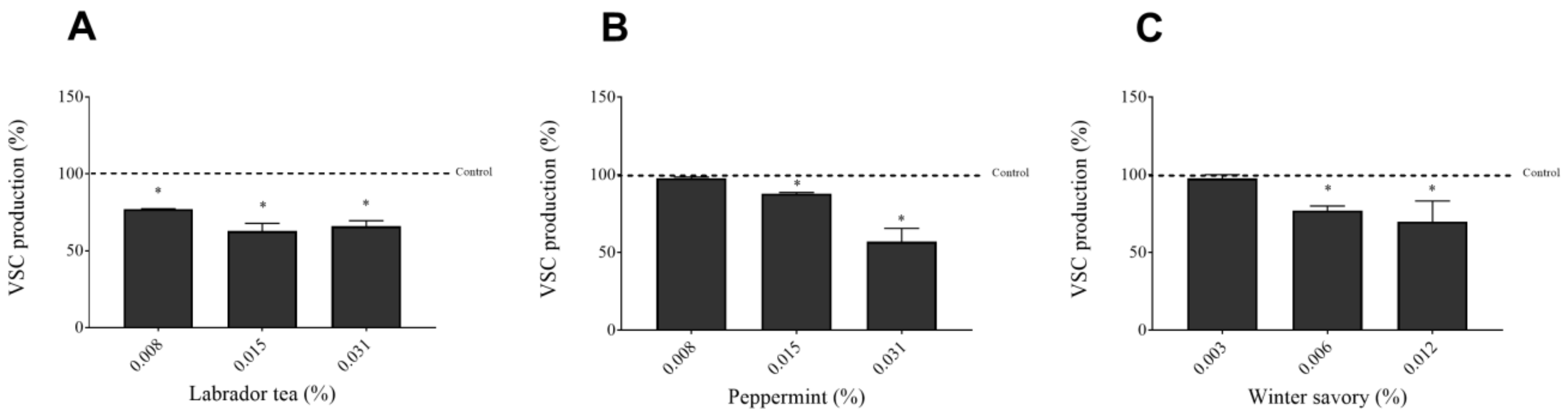
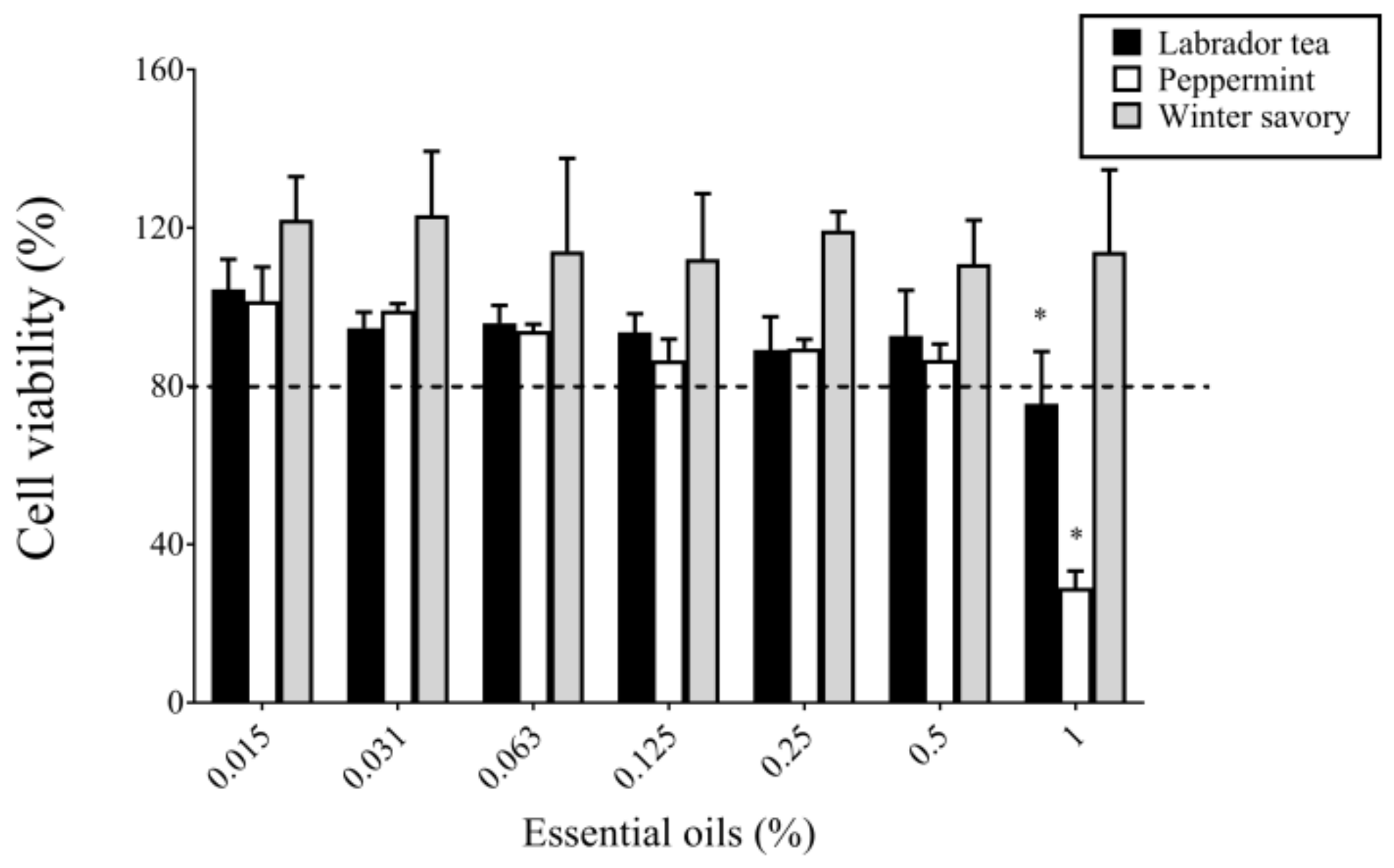
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacteria and Growth Conditions
4.2. Essential Oils
4.3. Determination of Minimum Inhibitory Concentrations and Minimum Bactericidal Concentrations
4.4. Biofilm Formation
4.5. Biofilm Desorption and Killing
4.6. Bacterial Membrane Permeability
4.7. Transmission Electron Microscopy Observation
4.8. VSC Production
4.9. In Vitro Cytotoxicity
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seemann, R.; Conceicao, M.D.; Filippi, A.; Greenman, J.; Lenton, P.; Nachnani, S.; Quirynen, M.; Roldan, S.; Schulze, H.; Sterer, N.; et al. Halitosis management by the general dental practitioner—results of an international consensus workshop. J. Breath Res. 2014, 8, 017101. [Google Scholar] [CrossRef] [PubMed]
- Broek, A.M.V.D.; Feenstra, L.; De Baat, C. A review of the current literature on aetiology and measurement methods of halitosis. J. Dent. 2007, 35, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Bollen, C.M.L.; Beikler, T. Halitosis: The multidisciplinary approach. Int. J. Oral Sci. 2012, 4, 55–63. [Google Scholar] [CrossRef]
- Suzuki, N.; Yoneda, M.; Takeshita, T.; Hirofuji, T.; Hanioka, T. Induction and inhibition of oral malodor. Mol. Oral Microbiol. 2019, 34, 85–96. [Google Scholar] [CrossRef]
- Persson, S.; Edlund, M.-B.; Claesson, R.; Carlsson, J. The formation of hydrogen sulfide and methyl mercaptan by oral bacteria. Oral Microbiol. Immunol. 1990, 5, 195–201. [Google Scholar] [CrossRef]
- Carlsson, J.; Larsen, J.T.; Edlund, M.B. Utilization of glutathione (L-gammaglutamyl-L-cysteinylglycine) by Fusobacterium nucleatum subspecies nucleatum. Oral Microbiol. Immunol. 1994, 9, 297–300. [Google Scholar] [CrossRef]
- Amou, T.; Hinode, D.; Yoshioka, M.; Grenier, D. Relationship between halitosis and periodontal disease-associated oral bacteria. Int. J. Dent. Hyg. 2014, 12, 145–151. [Google Scholar] [CrossRef]
- Loozen, G.; Ozcelik, O.; Boon, N.; De Mol, A.; Schoen, C.; Quirynen, M.; Teughels, W. Inter-bacterial correlations in subgingival biofilms: A large-scale survey. J. Clin. Periodontol. 2013, 41, 1–10. [Google Scholar] [CrossRef]
- Han, Y.W. Fusobacterium nucleatum: A commensal-turned pathogen. Curr. Opin. Microbiol. 2015, 23, 141–147. [Google Scholar] [CrossRef]
- Brennan, C.A.; Garrett, W.S. Fusobacterium nucleatum—Symbiont, opportunist and oncobacterium. Nat. Rev. Genet. 2019, 17, 156–166. [Google Scholar] [CrossRef]
- Nagraj, S.K.; Eachempati, P.; Uma, E.; Singh, V.P.; Ismail, N.M.; Varghese, E. Interventions for managing halitosis. Cochrane Database Syst. Rev. 2019, 12. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.; Camele, I. An Overview of the Biological Effects of Some Mediterranean Essential Oils on Human Health. BioMed Res. Int. 2017, 2017, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Dampc, A.; Luczkiewicz, M. Labrador tea – the aromatic beverage and spice: A review of origin, processing and safety. J. Sci. Food Agric. 2015, 95, 1577–1583. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, G.; Rahman, L.U. Ethnomedicinal, phytochemical and pharmacological updates on Peppermint (Mentha x piperita L.)—A review. Phytother. Res. 2020, 34, 2088–2139. [Google Scholar] [CrossRef] [PubMed]
- Tepe, B.; Cilkiz, M. A pharmacological and phytochemical overview on Satureja. Pharm. Biol. 2015, 54, 375–412. [Google Scholar] [CrossRef]
- Poaty, B.; Lahlah, J.; Porqueres, F.; Bouafif, H. Composition, antimicrobial and antioxidant activities of seven essential oils from the North American boreal forest. World J. Microbiol. Biotechnol. 2015, 31, 907–919. [Google Scholar] [CrossRef]
- Valdivieso-Ugarte, M.; Gomez-Llorente, C.; Plaza-Díaz, J.; Gil, Á. Antimicrobial, Antioxidant, and Immunomodulatory Properties of Essential Oils: A Systematic Review. Nutrients 2019, 11, 2786. [Google Scholar] [CrossRef]
- Lebel, G.; Vaillancourt, K.; Bercier, P.; Grenier, D. Antibacterial activity against porcine respiratory bacterial pathogens and in vitro biocompatibility of essential oils. Arch. Microbiol. 2019, 201, 833–840. [Google Scholar] [CrossRef]
- Vaillancourt, K.; Lebel, G.; Yi, L.; Grenier, D. In vitro antibacterial activity of plant essential oils against Staphylococcus hyicus and Staphylococcus aureus, the causative agents of exudative epidermitis in pigs. Arch. Microbiol. 2018, 200, 1001–1007. [Google Scholar] [CrossRef]
- Šimunović, K.; Bucar, F.; Klančnik, A.; Pompei, F.; Paparella, A.; Možina, S.S. In Vitro Effect of the Common Culinary Herb Winter Savory (Satureja montana) against the Infamous Food Pathogen Campylobacter jejuni. Foods 2020, 9, 537. [Google Scholar] [CrossRef] [PubMed]
- Oussalah, M.; Caillet, S.; Lacroix, M. Mechanism of Action of Spanish Oregano, Chinese Cinnamon, and Savory Essential Oils against Cell Membranes and Walls of Escherichia coli O157:H7 and Listeria monocytogenes. J. Food Prot. 2006, 69, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- O#x2019;Bryan, C.A.; Pendleton, S.J.; Crandall, P.G.; Ricke, S.C. Potential of Plant Essential Oils and Their Components in Animal Agriculture—In vitro Studies on Antibacterial Mode of Action. Front. Veter Sci. 2015, 2, 35. [Google Scholar] [CrossRef]
- Popa, M.; Marutescu, L.G.; Oprea, E.; Bleotu, C.; Kamerzan, C.; Chifiriuc, M.-C.; Pircalabioru, G.G. In Vitro Evaluation of the Antimicrobial and Immunomodulatory Activity of Culinary Herb Essential Oils as Potential Perioceutics. Antibiotics 2020, 9, 428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Y.; Zhu, X.; Cao, P.; Wei, S.; Lu, Y. Antibacterial and antibiofilm activities of eugenol from essential oil of Syzygium aromaticum (L.) Merr. & L. M. Perry (clove) leaf against periodontal pathogen Porphyromonas gingivalis. Microb. Pathog. 2017, 113, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Ito, S.; Kamo, M.; Kezuka, Y.; Tamura, H.; Kunimatsu, K.; Kato, H. Production of hydrogen sulfide by two enzymes associated with biosynthesis of homocysteine and lanthionine in Fusobacterium nucleatum subsp. nucleatum ATCC 25586. Microbiology 2010, 156, 2260–2269. [Google Scholar] [CrossRef]
- Quirynen, M.; Van Eldere, J.; Pauwels, M.; Bollen, C.M.; Van Steenberghe, D. In vitro volatile sulfur compound production of oral bacteria in different culture media. Quintessence Int. 1999, 30, 351–356. [Google Scholar] [PubMed]
- Chen, W.; Kajiya, M.; Giro, G.; Ouhara, K.; Mackler, H.E.; Mawardi, H.; Boisvert, H.; Duncan, M.J.; Sato, K.; Kawai, T. Bacteria-derived hydrogen sulfide promotes IL-8 production from epithelial cells. Biochem. Biophys. Res. Commun. 2010, 391, 645–650. [Google Scholar] [CrossRef]
- Murata, T.; Yaegaki, K.; Qian, W.; Herai, M.; Calenic, B.; Imai, T.; Sato, T.; Tanaka, T.; Kamoda, T.; Ii, H. Hydrogen sulfide induces apoptosis in epithelial cells derived from human gingiva. J. Breath Res. 2008, 2, 017007. [Google Scholar] [CrossRef]
- Zhang, J.-H.; Dong, Z.; Chu, L. Hydrogen sulfide induces apoptosis in human periodontium cells. J. Periodontal Res. 2010, 45, 71–78. [Google Scholar] [CrossRef]
- Yaegaki, K.; Qian, W.; Murata, T.; Imai, T.; Sato, T.; Tanaka, T.; Kamoda, T. Oral malodorous compound causes apoptosis and genomic DNA damage in human gingival fibroblasts. J. Periodontal Res. 2008, 43, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Pitts, G.; Pianotti, R.; Feary, T.; McGuiness, J.; Masurat, T. The in vivo Effects of an Antiseptic Mouthwash on Odor-producing Microorganisms. J. Dent. Res. 1981, 60, 1891–1896. [Google Scholar] [CrossRef] [PubMed]
- Ramji, N.; Ramji, N.; Iyer, R.; Chandrasekaran, S. Phenolic antibacterials from Piper betle in the prevention of halitosis. J. Ethnopharmacol. 2002, 83, 149–152. [Google Scholar] [CrossRef]
- Lebel, G.; Haas, B.; Adam, A.-A.; Veilleux, M.-P.; Ben Lagha, A.; Grenier, D. Effect of cinnamon (Cinnamomum verum) bark essential oil on the halitosis-associated bacterium Solobacterium moorei and in vitro cytotoxicity. Arch. Oral Biol. 2017, 83, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Coronel-Leon, J.; Lopez, A.; Espuny, M.J.; Beltran, M.T.; Molinos-Gomez, A.; Rocabayera, X.; Manresa, A. Assessment of antimicrobial activity of Nα-lauroyl arginate ethylester (LAE®) against Yersenia enterocolitica and Lactobacillus plantarum by flow cytometry and transmission electron microscopy. Food Cont. 2016, 63, 1–10. [Google Scholar] [CrossRef]
- Vanrobaeys, M.; De Herdt, P.; Charlier, G.; Ducatelle, R.; Haesebrouck, F. Ultrastructure of surface components of Streptococcus gallolytics (S. bovis) strains of differing virulence isolated from pigeons. Microbiology 1999, 145, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Gröger, S.; Michel, J.; Meyle, J. Establishment and characterization of immortalized human gingival keratinocyte cell lines. J. Periodontal Res. 2008, 43, 604–614. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Labrador Tea | Peppermint | Winter Savory |
|---|---|---|---|
| β-Bisabolene | 0.94 | - | 0.85 |
| Borneol | - | - | 2.16 |
| Bornyl acetate | 1.89 | - | - |
| Camphene | 1.21 | - | 0.69 |
| Carvacrol | - | - | 43.84 |
| β-Caryophyllene | 0.6 | 2.6 | - |
| 1,8-Cineol | - | 5.31 | 0.56 |
| p-Cymene | 1.63 | - | 14.3 |
| β-Elemene | 1.3 | - | - |
| Eudesma-3,7-dien-2-one | 1 | - | - |
| Germacrene B | 9.76 | - | - |
| Germacrene D | 1.82 | - | - |
| Germacrone | 8.51 | - | - |
| α-Humulene | 2.77 | - | - |
| Ledol | 0.81 | - | - |
| Limonene | 2.34 | 2.52 | 0.5 |
| Linalool | - | - | 1.04 |
| Menthofurane | - | 7.1 | - |
| Menthol | - | 42.3 | |
| Menthone | - | 24.7 | - |
| Iso-Menthone | - | 3.4 | - |
| Myrcene | - | - | 1.11 |
| Myrtenal | 2.17 | - | - |
| Oct-1-en-3-ol | - | - | 0.64 |
| α-Pinene | 3.25 | 0.76 | 1.97 |
| β-Pinene | 3.25 | 1.07 | - |
| trans-Pinocarveol | 1.44 | - | - |
| Pinocarvone | 0.83 | - | - |
| Pulegone | - | 6.7 | - |
| Sabina ketone | 0.5 | - | - |
| Sabinene | 11.93 | - | - |
| Dehydro sabinene ketone + trans-p-mentha-2,8-dien-1-ol | 0.65 | - | - |
| α-Selinene | 8.89 | - | - |
| β-Selinene | 10.95 | - | - |
| Terpinen-4-ol | 1.68 | - | 4.05 |
| α-Terpinene | 0.72 | - | 1.14 |
| γ-Terpinene | 2.58 | - | 12.66 |
| Thuj-3-en-10-al | 0.86 | - | - |
| Thymol | - | - | 6.71 |
| Thymol methyl ether | - | - | 3.09 |
| Different compounds at a concentration < 0.5% | 14.12 | 2.82 | 3.08 |
| Essential Oil | MIC (%; v/v) | MBC (%; v/v) | ||||
|---|---|---|---|---|---|---|
| ATCC 25586 | ATCC 10953 | ATCC 49256 | ATCC 25586 | ATCC 10953 | ATCC 49256 | |
| Labrador tea | 0.25 | 0.25 | 0.25 | 0.5 | 0.5 | 0.5 |
| Peppermint | 0.25 | 0.5 | 0.5 | 0.5 | 0.5 | 1 |
| Winter savory | 0.031 | 0.031 | 0.063 | 0.031 | 0.031 | 0.063 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Lagha, A.; Vaillancourt, K.; Maquera Huacho, P.; Grenier, D. Effects of Labrador Tea, Peppermint, and Winter Savory Essential Oils on Fusobacterium nucleatum. Antibiotics 2020, 9, 794. https://doi.org/10.3390/antibiotics9110794
Ben Lagha A, Vaillancourt K, Maquera Huacho P, Grenier D. Effects of Labrador Tea, Peppermint, and Winter Savory Essential Oils on Fusobacterium nucleatum. Antibiotics. 2020; 9(11):794. https://doi.org/10.3390/antibiotics9110794
Chicago/Turabian StyleBen Lagha, Amel, Katy Vaillancourt, Patricia Maquera Huacho, and Daniel Grenier. 2020. "Effects of Labrador Tea, Peppermint, and Winter Savory Essential Oils on Fusobacterium nucleatum" Antibiotics 9, no. 11: 794. https://doi.org/10.3390/antibiotics9110794
APA StyleBen Lagha, A., Vaillancourt, K., Maquera Huacho, P., & Grenier, D. (2020). Effects of Labrador Tea, Peppermint, and Winter Savory Essential Oils on Fusobacterium nucleatum. Antibiotics, 9(11), 794. https://doi.org/10.3390/antibiotics9110794