Antiviral Effects of Lindera obtusiloba Leaf Extract on Murine Norovirus-1 (MNV-1), a Human Norovirus Surrogate, and Potential Application to Model Foods

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
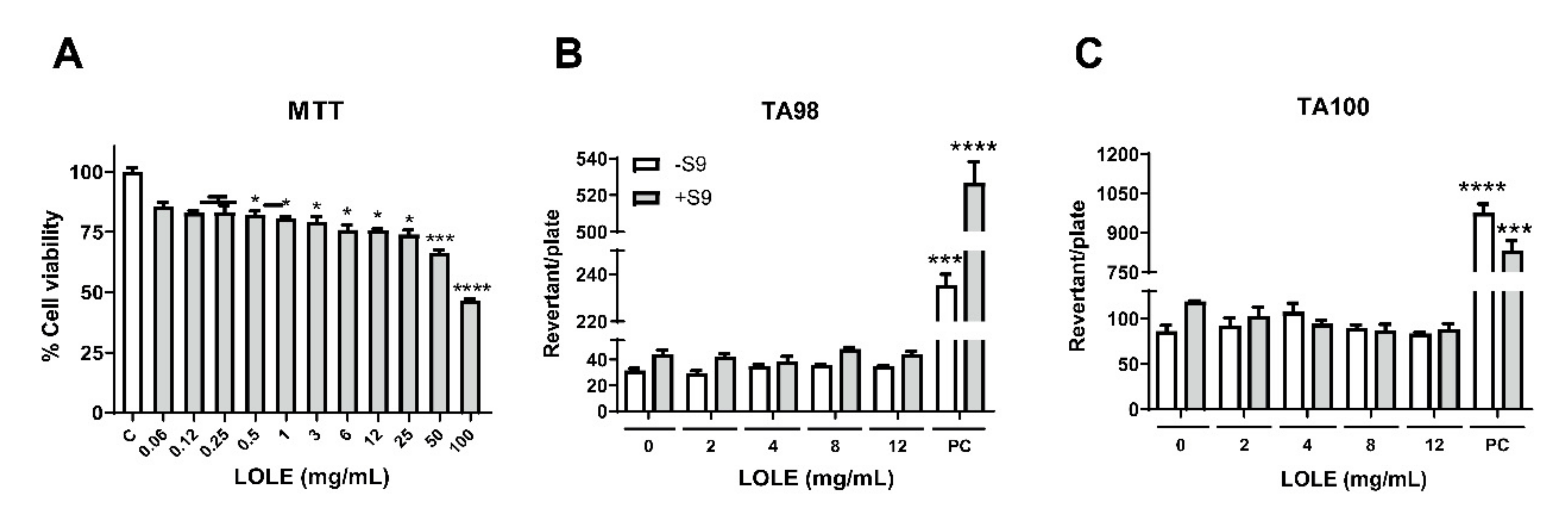
2.1. Evaluation of LOLE Cytotoxicity
2.2. Dose-Response and Incubation Times of LOLE on Neutralization of MNV-1
2.3. Temperature-, Dose-, and Time-Response Relationships between LOLE Exposure and MNV-1 Inactivation
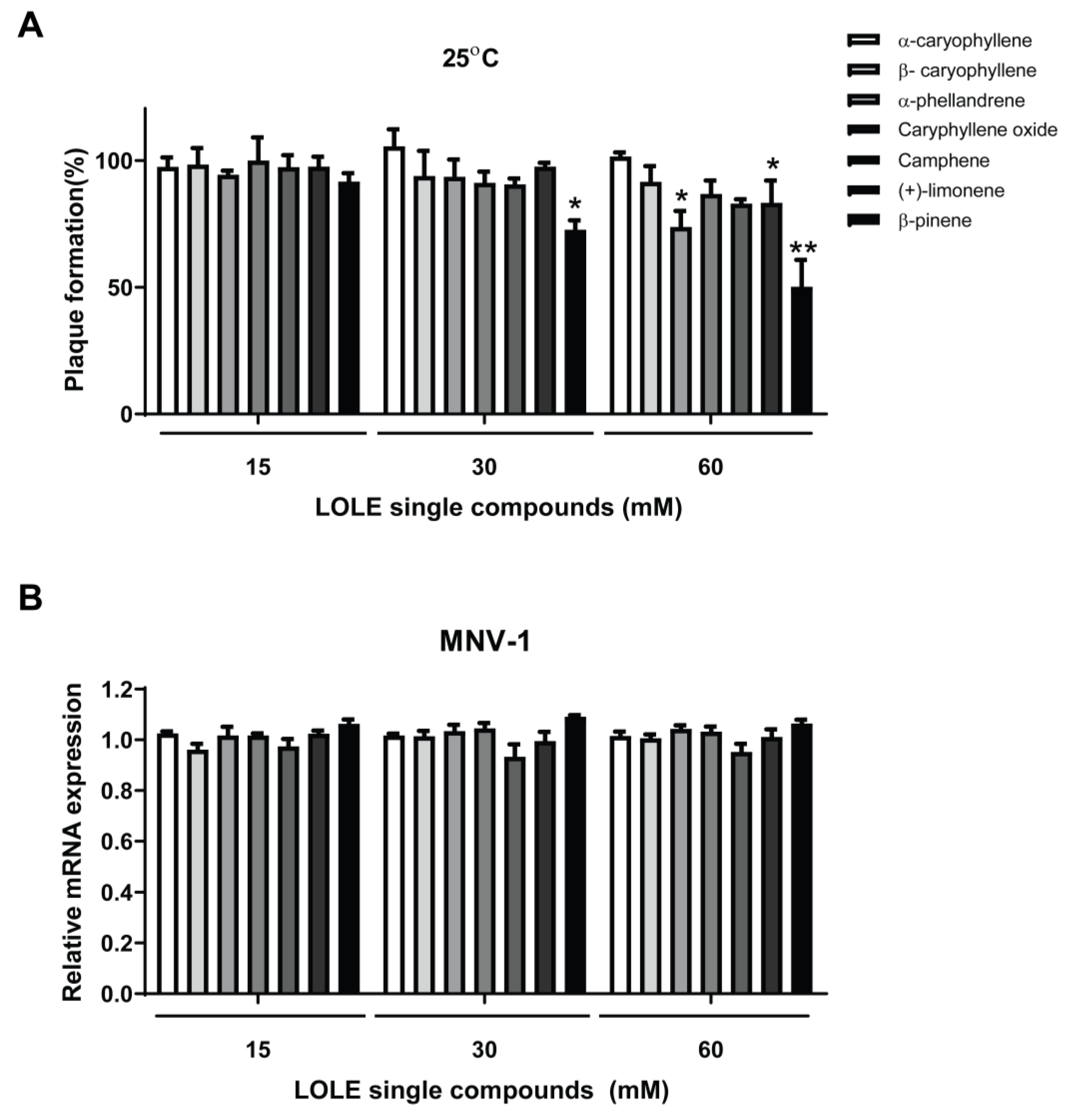
2.4. Anti-Noroviral Activity of LOLE Single Compounds
2.5. LOLE Single Compounds Did not Significantly Reduce MNV-1 Genome Copies
2.6. Inactivation of Norovirus by LOLE in Model Food Systems
3. Discussion
4. Materials and Methods
4.1. Material and Reagents
4.2. 3-[4,5-Dimethylthiazol-2-yl]-2,5-Diphenyltetrazolium Bromide (MTT) Cell Viability Assay
4.3. Mutagenic Ames Test
4.4. Preparation and Titration of MNV-1
4.5. Plaque Reduction Assay
4.6. Plaque Reduction Assay of MNV-1 on Model Foods and Stainless-Steel
4.7. Assessment of MNV-1 Viral Adsorption on Host Cells by Quantitative Polymerase Chain Reaction (qPCR)
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lucero, Y.; Vidal, R. Norovirus vaccines under development. Vaccine 2018, 36, 5435–5441. [Google Scholar] [CrossRef]
- Bok, K.; Green, K.Y. Norovirus Gastroenteritis in Immunocompromised Patients. N. Engl. J. Med. 2013, 368, 971. [Google Scholar] [CrossRef] [PubMed]
- Buzby, J.C.; Roberts, T. The economics of enteric infections: Human foodborne disease costs. Gastroenterology 2009, 136, 1851–1862. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; You, H.J.; Kim, Y.W.; Lee, A.; Ko, G.P.; Lee, S.-J.; Song, M.J. Inactivation of norovirus and surrogates by natural phytochemicals and bioactive substances. Mol. Nutr. Food Res. 2015, 59, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Atmar, R.L.; Bernstein, D.I.; Harro, C.D.; Al-Ibrahim, M.S.; Chen, W.H.; Ferreira, J.; Estes, M.K.; Graham, D.Y.; Opekun, A.R.; Richardson, C. Norovirus vaccine against experimental human Norwalk Virus illness. N. Engl. J. Med. 2011, 365, 2178–2187. [Google Scholar] [CrossRef]
- Malm, M.; Tamminen, K.; Lappalainen, S.; Vesikari, T.; Blazevic, V. Rotavirus recombinant VP6 nanotubes act as an immunomodulator and delivery vehicle for norovirus virus-like particles. J. Immunol. Res. 2016, 2016, 1–13. [Google Scholar] [CrossRef]
- Kapikian, A.Z. The Discovery of the 27-nm Norwalk Virus: An Historic Perspective. J. Infect. Dis. 2000, 181, S295–S302. [Google Scholar] [CrossRef]
- Karst, S.M. Pathogenesis of Noroviruses, Emerging RNA Viruses. Viruses 2010, 2, 748–781. [Google Scholar] [CrossRef]
- Hewitt, J.; Rivera-Aban, M.; Greening, G.E. Evaluation of murine norovirus as a surrogate for human norovirus and hepatitis A virus in heat inactivation studies. J. Appl. Microbiol. 2009, 107, 65–71. [Google Scholar] [CrossRef]
- Cannon, J.L.; Papafragkou, E.; Park, G.W.; Osborne, J.; Jaykus, L.-A.; Vinjé, J. Surrogates for the study of norovirus stability and inactivation in the environment: A comparison of murine norovirus and feline calicivirus. J. Food Prot. 2006, 69, 2761–2765. [Google Scholar] [CrossRef]
- Belliot, G.; Lavaux, A.; Souihel, D.; Agnello, D.; Pothier, P. Use of murine norovirus as a surrogate to evaluate resistance of human norovirus to disinfectants. Appl. Environ. Microbiol. 2008, 74, 3315–3318. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Bae, S.Y.; Oh, M.; Seok, J.H.; Kim, S.; Chung, Y.B.; Gowda, K.G.; Mun, J.Y.; Chung, M.S.; Kim, K.H. Antiviral effects of black raspberry (Rubus coreanus) seed extract and its polyphenolic compounds on norovirus surrogates. Biosci. Biotechnol. Biochem. 2016, 80, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.C.; Kim, Y.W.; Ryu, S.; Lee, A.; Lee, J.-S.; Song, M.J. Suppression of norovirus by natural phytochemicals from Aloe vera and Eriobotryae Folium. Food Control 2017, 73, 1362–1370. [Google Scholar] [CrossRef]
- Su, X.; D’Souza, D.H. Naturally Occurring Flavonoids Against Human Norovirus Surrogates. Food Environ. Virol. 2013, 5, 97–102. [Google Scholar] [CrossRef]
- Barclay, L.; Park, G.; Vega, E.; Hall, A.; Parashar, U.; Vinjé, J.; Lopman, B. Infection control for norovirus. Clin. Microbiol. Infect. 2014, 20, 731–740. [Google Scholar] [CrossRef]
- DiCaprio, E. Recent advances in human norovirus detection and cultivation methods. Curr. Opin. Food Sci. 2017, 14, 93–97. [Google Scholar] [CrossRef]
- Cook, N.; Knight, A.; Richards, G.P. Persistence and elimination of human norovirus in food and on food contact surfaces: A critical review. J. Food Prot. 2016, 79, 1273–1294. [Google Scholar] [CrossRef]
- Tung, G.; Macinga, D.; Arbogast, J.; Jaykus, L.-A. Efficacy of commonly used disinfectants for inactivation of human noroviruses and their surrogates. J. Food Prot. 2013, 76, 1210–1217. [Google Scholar] [CrossRef]
- Barbieri, R.; Coppo, E.; Marchese, A.; Daglia, M.; Sobarzo-Sanchez, E.; Nabavi, S.F.; Nabavi, S.M. Phytochemicals for human disease: An update on plant-derived compounds antibacterial activity. Microbiol. Res. 2017, 196, 44–68. [Google Scholar] [CrossRef]
- Li, D.; Baert, L.; Zhang, D.; Xia, M.; Zhong, W.; Van Coillie, E.; Jiang, X.; Uyttendaele, M. Effect of Grape Seed Extract on Human Norovirus GII.4 and Murine Norovirus 1 in Viral Suspensions, on Stainless Steel Discs, and in Lettuce Wash Water. Appl. Environ. Microbiol. 2012, 78, 7572–7578. [Google Scholar] [CrossRef]
- Kim, S.-H.; Do, J.-S.; Chung, H.-J. Antimicrobial activities of Lindera obtusiloba Blume and Zanthoxylum piperitum DC extracts. Korean J. Food Preserv. 2014, 21, 427–433. [Google Scholar] [CrossRef]
- Freise, C.; Ruehl, M.; Erben, U.; Neumann, U.; Seehofer, D.; Kim, K.Y.; Trowitzsch-Kienast, W.; Stroh, T.; Zeitz, M.; Somasundaram, R. A hepatoprotective Lindera obtusiloba extract suppresses growth and attenuates insulin like growth factor-1 receptor signaling and NF-kappaB activity in human liver cancer cell lines. BMC Complement. Altern. Med. 2011, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Lee, S.-Y.; Kim, B.; Kim, D.K.; Choi, J.-B.; Kim, J.-A.; Lee, H.-J.; Joo, I.S.; Lee, J.S.; Kang, D.H.; et al. Inactivation of Norovirus by Lemongrass Essential Oil Using a Norovirus Surrogate System. J. Food Prot. 2017, 80, 1293–1302. [Google Scholar] [CrossRef] [PubMed]
- Gottardi, D.; Bukvicki, D.; Prasad, S.; Tyagi, A.K. Beneficial Effects of Spices in Food Preservation and Safety. Front. Microbiol. 2016, 7, 1394. [Google Scholar] [CrossRef]
- Del Nobile, M.A.; Lucera, A.; Costa, C.; Conte, A. Food applications of natural antimicrobial compounds. Front. Microbiol. 2012, 3, 287. [Google Scholar]
- Hong, C.O.; SeoMun, Y.; Koo, Y.C.; Nam, M.H.; Lee, H.A.; Kim, J.H.; Yang, S.Y.; Lee, S.H.; No, S.H.; Lee, K.W. Single and 14-day repeated oral toxicity studies of 70% ethanol extract of Lindera obtusiloba blume leaves. J. Korean Soc. Food Sci. Nutr. 2009, 38, 1324–1330. [Google Scholar] [CrossRef]
- Han, A.R.; Kim, S.W.; Chun, S.H.; Nam, M.H.; Hong, C.O.; Kim, B.H.; Kim, T.C.; Lee, K.W. Effect of diet containing whole wheat bread with capsosiphon fulvescens and Lindera obtusiloba ethanol extracts on plasma glucose and lipid levels in rats. Korean J. Food Sci. Technol. 2016, 48, 178–186. [Google Scholar] [CrossRef]
- Gilling, D.H.; Kitajima, M.; Torrey, J.R.; Bright, K.R. Antiviral efficacy and mechanisms of action of oregano essential oil and its primary component carvacrol against murine norovirus. J. Appl. Microbiol. 2014, 116, 1149–1163. [Google Scholar] [CrossRef]
- Elizaquível, P.; Azizkhani, M.; Aznar, R.; Sánchez, G. The effect of essential oils on norovirus surrogates. Food Control 2013, 32, 275–278. [Google Scholar] [CrossRef]
- Ali, N.A.M.; Jantan, I. The essential oils of Lindera pipericarpa. J. Trop. For. Sci. 1993, 6, 124–130. [Google Scholar]
- Kwon, D.-J.; Kim, J.-K.; Bae, Y.-S. Essential oils from leaves and twigs of Lindera obtusiloba. J. Korean Soc. For. Sci. 2007, 96, 65–69. Available online: https://www.koreascience.or.kr/article/JAKO200710103466805.page (accessed on 23 June 2020).
- Rahman, A.; Shanta, Z.S.; Rashid, M.; Parvin, T.; Afrin, S.; Khatun, M.K.; Sattar, M. In vitro antibacterial properties of essential oil and organic extracts of Premna integrifolia Linn. Arab. J. Chem. 2016, 9, S475–S479. [Google Scholar] [CrossRef]
- Joshi, S.; Howell, A.B.; D’Souza, D.H. Blueberry proanthocyanidins against human norovirus surrogates in model foods and under simulated gastric conditions. Food Microbiol. 2017, 63, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Gilling, D.H.; Kitajima, M.; Torrey, J.R.; Bright, K.R. Mechanisms of antiviral action of plant antimicrobials against murine norovirus. Appl. Environ. Microbiol. 2014, 80, 4898–4910. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Lee, H.m.; Kim, S.H.; Eun, J.B.; Ha, J.-H. Recovery of intact human norovirus from cabbage Kimchi stored at 4 °C and 10 °C during fermentation. LWT 2017, 78, 258–264. [Google Scholar] [CrossRef]
- Banach, J.L.; Sampers, I.; Van Haute, S.; der Fels-Klerx, V. Effect of disinfectants on preventing the cross-contamination of pathogens in fresh produce washing water. Int. J. Environ. Res. Public Health 2015, 12, 8658–8677. [Google Scholar] [CrossRef]
- DiCaprio, E.; Purgianto, A.; Ma, Y.; Hughes, J.; Dai, X.; Li, J. Attachment and localization of human norovirus and animal caliciviruses in fresh produce. Int. J. Food Microbiol. 2015, 211, 101–108. [Google Scholar] [CrossRef]
- Lou, F.; Neetoo, H.; Chen, H.; Li, J. Inactivation of a human norovirus surrogate by high-pressure processing: Effectiveness, mechanism, and potential application in the fresh produce industry. Appl. Environ. Microbiol. 2011, 77, 1862–1871. [Google Scholar] [CrossRef]
- Sharps, C.P.; Kotwal, G.; Cannon, J.L. Human norovirus transfer to stainless steel and small fruits during handling. J. Food Prot. 2012, 75, 1437–1446. [Google Scholar] [CrossRef]
- Hwang, S.H.; Choi, S.J.; Hwang, Y.S.; Lim, S.S. Comparison analysis of essential oils composition in difference parts from Lindera obtusiloba BL. according to the season by gas Chromatography-Mass Spectrometry (GC-MS). Korean J. Pharmacogn. 2013, 44, 30–40. Available online: https://www.koreascience.or.kr/article/JAKO201317642692157.page (accessed on 14 July 2020).
- Maron, D.M.; Ames, B.N. Revised methods for the Salmonella mutagenicity test. Mutat. Res. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Ganguli, P.S.; Chen, W.; Yates, M.V. Detection of murine norovirus-1 by using TAT peptide-delivered molecular beacons. Appl. Environ. Microbiol. 2011, 77, 5517–5520. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gonzalez-Hernandez, M.B.; Cunha, J.B.; Wobus, C.E. Plaque assay for murine norovirus. J. Vis. Exp. 2012, e4297. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Breiman, A.; Le Pendu, J.; Uyttendaele, M. Anti-viral Effect of Bifidobacterium adolescentis against Noroviruses. Front. Microbiol. 2016, 7, 864. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solis-Sanchez, D.; Rivera-Piza, A.; Lee, S.; Kim, J.; Kim, B.; Choi, J.B.; Kim, Y.W.; Ko, G.P.; Song, M.J.; Lee, S.-J. Antiviral Effects of Lindera obtusiloba Leaf Extract on Murine Norovirus-1 (MNV-1), a Human Norovirus Surrogate, and Potential Application to Model Foods. Antibiotics 2020, 9, 697. https://doi.org/10.3390/antibiotics9100697
Solis-Sanchez D, Rivera-Piza A, Lee S, Kim J, Kim B, Choi JB, Kim YW, Ko GP, Song MJ, Lee S-J. Antiviral Effects of Lindera obtusiloba Leaf Extract on Murine Norovirus-1 (MNV-1), a Human Norovirus Surrogate, and Potential Application to Model Foods. Antibiotics. 2020; 9(10):697. https://doi.org/10.3390/antibiotics9100697
Chicago/Turabian StyleSolis-Sanchez, Diana, Adriana Rivera-Piza, Soyoung Lee, Jia Kim, Bomi Kim, Joo Bong Choi, Ye Won Kim, Gwang Pyo Ko, Moon Jung Song, and Sung-Joon Lee. 2020. "Antiviral Effects of Lindera obtusiloba Leaf Extract on Murine Norovirus-1 (MNV-1), a Human Norovirus Surrogate, and Potential Application to Model Foods" Antibiotics 9, no. 10: 697. https://doi.org/10.3390/antibiotics9100697
APA StyleSolis-Sanchez, D., Rivera-Piza, A., Lee, S., Kim, J., Kim, B., Choi, J. B., Kim, Y. W., Ko, G. P., Song, M. J., & Lee, S.-J. (2020). Antiviral Effects of Lindera obtusiloba Leaf Extract on Murine Norovirus-1 (MNV-1), a Human Norovirus Surrogate, and Potential Application to Model Foods. Antibiotics, 9(10), 697. https://doi.org/10.3390/antibiotics9100697