Susceptibility of Campylobacter Strains to Selected Natural Products and Frontline Antibiotics

 ,
,  , ,
, ,  ,
,
Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Sampling Site
4.2. Sample Collection, Campylobacter Isolation, and Antimicrobial Testing
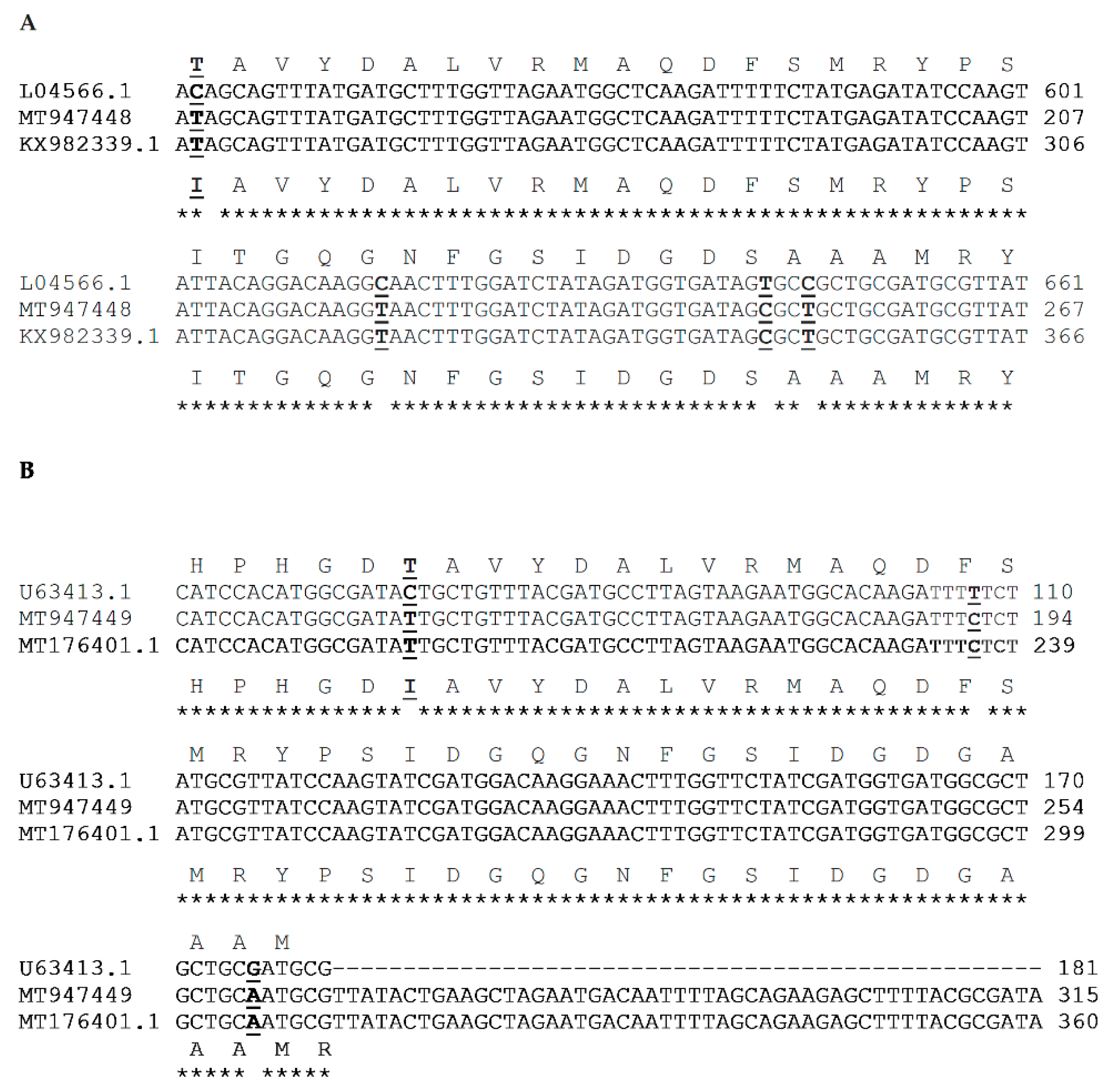
4.3. DNA Extraction, PCR, and Sequencing
4.4. Natural Products and Antibiotics
4.4.1. Plant Extracts, EOs, Pure Phytochemicals, and Conventional Antimicrobials
4.4.2. Determination of MIC and MBC
4.4.3. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scallan, E.; Hoekstra, R.M.; Angulo, F.; Tauxe, R.; Widdwson, M.-A.; Roy, S.; Jones, J.; Griffin, P. Foodborne illness acquired in the United States—major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Dufour, V.; Alazzam, B.; Ermel, G.; Thepaut, M.; Rossero, A.; Tresse, O.; Baysse, C. Antimicrobial Activities of Isothiocyanates Against Campylobacter jejuni Isolates. Front. Cell. Infect. Microbiol. 2012, 2, 53. [Google Scholar] [CrossRef] [PubMed]
- Srey, S.; Jahid, I.K.; Ha, S.-D. Biofilm formation in food industries: A food safety concern. Food Control. 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Klančnik, A.; Možina, S.S.; Zhang, Q. Anti-Campylobacter Activities and Resistance Mechanisms of Natural Phenolic Compounds in Campylobacter. PLoS ONE 2012, 7, e51800. [Google Scholar] [CrossRef] [PubMed]
- Osbjer, K.; Tano, E.; Chhayheng, L.; Mac-Kwashie, A.O.; Fernström, L.; Ellström, P.; Sokerya, S.; Sokheng, C.; Mom, V.; Chheng, K.; et al. Detection of Campylobacter in human and animal field samples in Cambodia. APMIS 2016, 124, 508–515. [Google Scholar] [CrossRef]
- Lynch, C.T.; Lynch, H.; Burke, S.; Hawkins, K.; Buttimer, C.; Mc Carthy, C.; Egan, J.; Whyte, P.; Bolton, D.J.; Coffey, A.; et al. Antimicrobial Resistance Determinants Circulating among Thermophilic Campylobacter Isolates Recovered from Broilers in Ireland Over a One-Year Period. Antibiotics 2020, 9, 308. [Google Scholar] [CrossRef]
- Koolman, L.; Whyte, P.; Burgess, C.; Bolton, D. Distribution of Virulence-Associated Genes in a Selection of Campylobacter Isolates. Foodborne Pathog. Dis. 2015, 12, 424–432. [Google Scholar] [CrossRef]
- Reddy, S.; Zishiri, O.T. Detection and prevalence of antimicrobial resistance genes in Campylobacter spp. isolated from chickens and humans. Onderstepoort J. Vet. Res. 2017, 84, 1–6. [Google Scholar] [CrossRef]
- Elisha, I.L.; Botha, F.S.; McGaw, L.J.; Eloff, J.N. The antibacterial activity of extracts of nine plant species with good activity against Escherichia coli against five other bacteria and cytotoxicity of extracts. BMC Complement. Altern. Med. 2017, 17, 1–10. [Google Scholar] [CrossRef]
- Wallace, R.L.; Bulach, D.; Jennison, A.V.; Valcanis, M.; McLure, A.; Smith, J.J.; Graham, T.; Saputra, T.; Firestone, S.; Symes, S.; et al. Molecular characterization of Campylobacter spp. recovered from beef, chicken, lamb and pork products at retail in Australia. PLoS ONE 2020, 15, e0236889. [Google Scholar] [CrossRef]
- Možina, S.S.; Kurinčič, M.; Klančnik, A.; Mavri, A. Campylobacter and its multi-resistance in the food chain. Trends Food Sci. Technol. 2011, 22, 91–98. [Google Scholar] [CrossRef]
- Frasao, B.D.S.; Medeiros, V.; Barbosa, A.V.; De Aguiar, W.S.; Dos Santos, F.F.; Abreu, D.L.D.C.; Clementino, M.M.; De Aquino, M.H.C. Detection of fluoroquinolone resistance by mutation in gyrA gene of Campylobacter spp. isolates from broiler and laying (Gallus gallus domesticus) hens, from Rio de Janeiro State, Brazil. Ciência Rural 2015, 45, 2013–2018. [Google Scholar] [CrossRef]
- Dai, L.; Sahin, O.; Grover, M.; Zhang, Q. New and alternative strategies for the prevention, control, and treatment of antibiotic-resistant Campylobacter. Transl. Res. 2020, 223, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Gibreel, A.; Kos, V.N.; Keelan, M.; Trieber, C.A.; Levesque, S.; Michaud, S.; Taylor, D.E. Macrolide Resistance in Campylobacter jejuni and Campylobacter coli: Molecular Mechanism and Stability of the Resistance Phenotype. Antimicrob. Agents Chemother. 2005, 49, 2753–2759. [Google Scholar] [CrossRef]
- Pratt, A.; Korolik, V. Tetracycline resistance of Australian Campylobacter jejuni and Campylobacter coli isolates. J. Antimicrob. Chemother. 2005, 55, 452–460. [Google Scholar] [CrossRef]
- Elhadidy, M.; Miller, W.G.; Arguello, H.; Álvarez-Ordóñez, A.; Duarte, A.; Dierick, K.; Botteldoorn, N. Genetic Basis and Clonal Population Structure of Antibiotic Resistance in Campylobacter jejuni Isolated From Broiler Carcasses in Belgium. Front. Microbiol. 2018, 9, 1014. [Google Scholar] [CrossRef]
- Burki, T.K. Superbugs: An Arms Race Against Bacteria. Lancet Respir. Med. 2018, 6, 668. [Google Scholar] [CrossRef]
- Sproston, E.L.; Wimalarathna, H.M.L.; Sheppard, S.K. Trends in fluoroquinolone resistance in Campylobacter. Microb. Genom. 2018, 4, e000198. [Google Scholar] [CrossRef]
- Sasirekha, B.; Gahamanyi, N. Photosensitizers: An Effective Alternative Approach to Microbial Pathogen. Front. Clin. Drug Res. (Anti Infect.) 2017, 3, 187–205. [Google Scholar]
- Cos, P.; Vlietinck, A.J.; Berghe, D.V.; Maes, L. Anti-infective potential of natural products: How to develop a stronger in vitro ‘proof-of-concept’. J. Ethnopharmacol. 2006, 106, 290–302. [Google Scholar] [CrossRef]
- Kürekci, C.; Padmanabha, J.; Bishop-Hurley, S.L.; Hassan, E.; Al Jassim, R.A.; McSweeney, C.S. Antimicrobial activity of essential oils and five terpenoid compounds against Campylobacter jejuni in pure and mixed culture experiments. Int. J. Food Microbiol. 2013, 166, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Falowo, A.B.; Fayemi, P.O.; Muchenje, V. Natural antioxidants against lipid–protein oxidative deterioration in meat and meat products: A review. Food Res. Int. 2014, 64, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, M.; Saki, K.; Shahsavari, S.; Rafieian-Kopaei, M.; Sepahvand, R.; Adineh, A. Identification of medicinal plants effective in infectious diseases in Urmia, northwest of Iran. Asian Pac. J. Trop. Biomed. 2015, 5, 858–864. [Google Scholar] [CrossRef]
- Ayaz, M.; Ullah, F.; Sadiq, A.; Ullah, F.; Ovais, M.; Ahmed, J.; Devkota, H.P. Synergistic interactions of phytochemicals with antimicrobial agents: Potential strategy to counteract drug resistance. Chem. Interactions 2019, 308, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Ooi, L.S.M.; Li, Y.; Kam, S.-L.; Wang, H.; Wong, E.Y.L.; Ooi, V.E.C. Antimicrobial Activities of Cinnamon Oil and Cinnamaldehyde from the Chinese Medicinal Herb Cinnamomum cassia Blume. Am. J. Chin. Med. 2006, 34, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Hu, N.; Wang, K.-X. Study of Scutellaria baicalensis and Baicalin against antimicrobial susceptibility of Helicobacter pylori strains in vitro. Zhong Yao Cai = Zhongyaocai = J. Chin. Med. Mater. 2008, 31, 707–710. [Google Scholar]
- Kim, H.-O.; Park, S.-W.; Park, H. Inactivation of Escherichia coli O157:H7 by cinnamic aldehyde purified from Cinnamomum cassia shoot. Food Microbiol. 2004, 21, 105–110. [Google Scholar] [CrossRef]
- Hassan, W.H.; Abdel-Ghany, A.E.; Afifi, S.I.; Sedik, S.H. Genotypic Characterization of Campylobacter Species Isolated from Livestock and Poultry and Evaluation of some Herbal Oil Antimicrobial Effect against Selected Campylobacter Species. Adv. Anim. Vet. Sci. 2019, 7, 1083–1092. [Google Scholar] [CrossRef]
- Imai, H.; Osawa, K.; Yasuda, H.; Hamashima, H.; Arai, T.; Sasatsu, M. Inhibition by the essential oils of peppermint and spearmint of the growth of pathogenic bacteria. Microbios 2001, 106, 31–39. [Google Scholar]
- Wannissorn, B.; Jarikasem, S.; Siriwangchai, T.; Thubthimthed, S. Antibacterial properties of essential oils from Thai medicinal plants. Fitoterapia 2005, 76, 233–236. [Google Scholar] [CrossRef]
- Thawkar, B.S. Phytochemical and pharmacological review of Mentha arvensis. Int. J. Green Pharm. 2016, 10. [Google Scholar] [CrossRef]
- Murata, T.; Miyase, T.; Yoshizaki, F. Hyaluronidase Inhibitory Rosmarinic Acid Derivatives from Meehania urticifolia. Chem. Pharm. Bull. 2011, 59, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Kovács, J.K.; Felső, P.; Makszin, L.; Pápai, Z.; Horváth, G.; Ábrahám, H.; Palkovics, T.; Böszörményi, A.; Emődy, L.; Schneider, G. Antimicrobial and Virulence-Modulating Effects of Clove Essential Oil on the Foodborne Pathogen Campylobacter jejuni. Appl. Environ. Microbiol. 2016, 82, 6158–6166. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Ghosh, A.; Hazra, B. Evaluation of the antibacterial activity of Ventilago madraspatana Gaertn., Rubia cordifolia Linn. And Lantana camara Linn.: Isolation of emodin and physcion as active antibacterial agents. Phytother. Res. 2005, 19, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Sohn, H.-Y.; Son, K.; Kwon, C.-S.; Kang, S. Antimicrobial and cytotoxic activity of 18 prenylated flavonoids isolated from medicinal plants: Morus alba L., Morus mongolica Schneider, Broussnetia papyrifera (L.) Vent, Sophora flavescens Ait and Echinosophora koreensis Nakai. Phytomedicine 2004, 11, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.-J.; Jeong, J.-H.; Lee, S.W.; Ryu, Y.B.; Jeong, H.J.; Jung, K.; Lim, J.S.; Cho, K.-O.; Lee, W.S.; Rho, M.-C.; et al. In vitro anti-reovirus activity of kuraridin isolated from Sophora flavescens against viral replication and hemagglutination. J. Pharmacol. Sci. 2015, 128, 159–169. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, Antimicrobial Mechanisms, and Antibiotic Activities of Cinnamaldehyde against Pathogenic Bacteria in Animal Feeds and Human Foods. J. Agric. Food Chem. 2017, 65, 10406–10423. [Google Scholar] [CrossRef]
- Gyawali, R.; Ibrahim, S.A. Natural products as antimicrobial agents. Food Control. 2014, 46, 412–429. [Google Scholar] [CrossRef]
- Roila, R.; Valiani, A.; Galarini, R.; Servili, M.; Branciari, R. Antimicrobial and anti-biofilm activity of olive oil by-products against Campylobacter spp. isolated from chicken meat. Acta Sci. Pol. Technol. Aliment. 2019, 18, 43–52. [Google Scholar] [CrossRef]
- Cho, E.; Yokozawa, T.; Rhyu, D.; Kim, S.; Shibahara, N.; Park, J. Study on the inhibitory effects of Korean medicinal plants and their main compounds on the 1,1-diphenyl-2-picrylhydrazyl radical. Phytomedicine 2003, 10, 544–551. [Google Scholar] [CrossRef]
- WHO. WHO Global Report on Traditional and Complementary Medicine 2019; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Hlashwayo, D.F.; Barbosa, F.; Langa, S.; Sigaúque, B.; Bila, C.G. A Systematic Review of In Vitro Activity of Medicinal Plants from Sub-Saharan Africa against Campylobacter spp. Evidence-Based Complement. Altern. Med. 2020, 2020, 1–13. Available online: https://www.hindawi.com/journals/ecam/2020/9485364/ (accessed on 4 June 2020). [CrossRef] [PubMed]
- Klančnik, A.; Šimunović, K.; Sterniša, M.; Ramić, D.; Možina, S.S.; Bucar, F. Anti-adhesion activity of phytochemicals to prevent Campylobacter jejuni biofilm formation on abiotic surfaces. Phytochem. Rev. 2020, 1–30. [Google Scholar] [CrossRef]
- Hossan, S.; Jindal, H.; Maisha, S.; Raju, C.S.; Sekaran, S.D.; Nissapatorn, V.; Kaharudin, F.; Yi, L.S.; Khoo, T.J.; Rahmatullah, M.; et al. Antibacterial effects of 18 medicinal plants used by the Khyang tribe in Bangladesh. Pharm. Biol. 2018, 56, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, J.; Hiremath, N.; Jacob, H. Antimicrobial, Rheological, and Thermal Properties of Plasticized Polylactide Films Incorporated with Essential Oils to Inhibit Staphylococcus aureus and Campylobacter jejuni. J. Food Sci. 2016, 81, E419–E429. [Google Scholar] [CrossRef] [PubMed]
- Micciche, A.; Rothrock, M.J.J.; Yang, Y.; Ricke, S.C. Essential Oils as an Intervention Strategy to Reduce Campylobacter in Poultry Production: A Review. Front. Microbiol. 2019, 10, 1058. [Google Scholar] [CrossRef]
- Shene, C.; Reyes, A.K.; Villarroel, M.; Sineiro, J.; Pinelo, M.; Rubilar, M. Plant location and extraction procedure strongly alter the antimicrobial activity of murta extracts. Eur. Food Res. Technol. 2008, 228, 467–475. [Google Scholar] [CrossRef]
- Kuete, V. Potential of Cameroonian Plants and Derived Products against Microbial Infections: A Review. Planta Medica 2010, 76, 1479–1491. [Google Scholar] [CrossRef]
- Murata, T.; Miyase, T.; Yoshizaki, F. Meehanines L−W, Spermidine Alkaloidal Glycosides fromMeehania urticifolia. J. Nat. Prod. 2009, 72, 1937–1943. [Google Scholar] [CrossRef]
- Murata, T.; Miyase, T.; Yoshizaki, F. New phenolic compounds from Meehania urticifolia. J. Nat. Med. 2011, 65, 385–390. [Google Scholar] [CrossRef]
- Lee, J.-H.; Park, J.-S. Antibacterial effect of traditional food ingredients for healthcare on Helicobacter pylori. Technol. Health Care 2019, 27, 509–518. [Google Scholar] [CrossRef]
- Lu, Y.; Joerger, R.; Wu, C. Study of the Chemical Composition and Antimicrobial Activities of Ethanolic Extracts from Roots of Scutellaria baicalensis Georgi. J. Agric. Food Chem. 2011, 59, 10934–10942. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.C.; Ip, M.; Lau, C.B.; Lui, S.L.; Jolivalt, C.; Ganem-Elbaz, C.; Litaudon, M.; Reiner, N.E.; Gong, H.; See, R.H.; et al. Synergistic effects of baicalein with ciprofloxacin against NorA over-expressed methicillin-resistant Staphylococcus aureus (MRSA) and inhibition of MRSA pyruvate kinase. J. Ethnopharmacol. 2011, 137, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.-Y.; Wan, X.-H.; Niu, F.-J.; Xie, S.-M.; Guo, H.; Yang, Y.-Y.; Guo, L.-Y.; Zhou, C.-Z. Salvia plebeia R. Br.: An overview about its traditional uses, chemical constituents, pharmacology and modern applications. Biomed. Pharmacother. 2020, 121, 109589. [Google Scholar] [CrossRef] [PubMed]
- Yamaki, M.; Kashihara, M.; Takagi, S. Activity of Ku Shen compounds against Staphylococcus aureus and Streptococcus mutans. Phytotherapy Res. 1990, 4, 235–236. [Google Scholar] [CrossRef]
- Chan, B.C.-L.; Yu, H.; Wong, C.-W.; Lui, S.-L.; Jolivalt, C.; Ganem-Elbaz, C.; Paris, J.-M.; Morleo, B.; Litaudon, M.; Lau, C.B.-S.; et al. Quick identification of kuraridin, a noncytotoxic anti-MRSA (methicillin-resistant Staphylococcus aureus) agent from Sophora flavescens using high-speed counter-current chromatography. J. Chromatogr. B 2012, 880, 157–162. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, M.Y.; Kim, S.J.; Jeon, S.-E.; Cha, I.; Hong, S.; Chung, G.T.; Huh, M.-J.; Kang, Y.-H.; Yoo, C.-K. High-Level Ciprofloxacin-Resistant Campylobacter jejuni Isolates Circulating in Humans and Animals in Incheon, Republic of Korea. Zoonoses Public Health 2016, 63, 545–554. [Google Scholar] [CrossRef]
- Gahamanyi, N.; Mboera, L.E.G.; Matee, M.I.; Mutangana, D.; Komba, E.V.G. Prevalence, Risk Factors, and Antimicrobial Resistance Profiles of Thermophilic Campylobacter Species in Humans and Animals in Sub-Saharan Africa: A Systematic Review. Int. J. Microbiol. 2020, 2020, 2092478. [Google Scholar] [CrossRef]
- Iovine, N.M. Resistance mechanisms inCampylobacter jejuni. Virulence 2013, 4, 230–240. [Google Scholar] [CrossRef]
- Ku, B.K.; Kim, H.J.; Lee, Y.-J.; Kim, Y.I.; Choi, J.S.; Park, M.Y.; Kwon, J.W.; Nam, H.-M.; Kim, Y.H.; Jung, S.-C.; et al. Genetic Characterization and Antimicrobial Susceptibility of Campylobacter spp. Isolated from Domestic and Imported Chicken Meats and Humans in Korea. Foodborne Pathog. Dis. 2011, 8, 381–386. [Google Scholar] [CrossRef]
- Wei, B.; Cha, S.-Y.; Kang, M.; Roh, J.-H.; Seo, H.-S.; Yoon, R.-H.; Jang, H.-K. Antimicrobial Susceptibility Profiles and Molecular Typing of Campylobacter jejuni and Campylobacter coli Isolates from Ducks in South Korea. Appl. Environ. Microbiol. 2014, 80, 7604–7610. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, W.M.; Taylor, D. Cloning and nucleotide sequence of the Campylobacter jejuni gyrA gene and characterization of quinolone resistance mutations. Antimicrob. Agents Chemother. 1993, 37, 457–463. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bachoual, R.; Ouabdesselam, S.; Mory, F.; Lascols, C.; Soussy, C.-J.; Tankovic, J. Single or Double Mutational Alterations of gyrA Associated with Fluoroquinolone Resistance in Campylobacter jejuni and Campylobacter coli. Microb. Drug Resist. 2001, 7, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J.V.; Ricci, V.; Pumbwe, L.; Everett, M.J.; Griggs, D.J. Fluoroquinolone resistance in Campylobacter species from man and animals: Detection of mutations in topoisomerase genes. J. Antimicrob. Chemother. 2002, 51, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.; Schmidt, A.-M.; Escher, U.; Kittler, S.; Kehrenberg, C.; Thunhorst, E.; Bereswill, S.; Heimesaat, M.M. Carvacrol ameliorates acute campylobacteriosis in a clinical murine infection model. Gut Pathog. 2020, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Jeon, B. Synergistic anti-Campylobacter jejuni activity of fluoroquinolone and macrolide antibiotics with phenolic compounds. Front. Microbiol. 2015, 6, 1129. [Google Scholar] [CrossRef]
- Kapperud, G.; Espeland, G.; Wahl, E.; Walde, A.; Herikstad, H.; Gustavsen, S.; Tveit, I.; Natås, O.; Bevanger, L.; Digranes, A. Factors Associated with Increased and Decreased Risk of Campylobacter Infection: A Prospective Case-Control Study in Norway. Am. J. Epidemiol. 2003, 158, 234–242. [Google Scholar] [CrossRef]
- Al-Nasrawi, H.A.-H.A. Phylogenetic analysis of Campylobacter jejuni from human and birds sources in Iraq. Afr. J. Microbiol. Res. 2016, 10, 752–758. [Google Scholar] [CrossRef][Green Version]
- Sibanda, N.; McKenna, A.; Richmond, A.; Ricke, S.C.; Callaway, T.; Stratakos, A.C.; Gundogdu, O.; Corcionivoschi, N. A Review of the Effect of Management Practices on Campylobacter Prevalence in Poultry Farms. Front. Microbiol. 2018, 9, 2002. [Google Scholar] [CrossRef]
- Humphrey, T.; O’Brien, S.; Madsen, M. Campylobacters as zoonotic pathogens: A food production perspective. Int. J. Food Microbiol. 2007, 117, 237–257. [Google Scholar] [CrossRef]
- Guévremont, E.; Nadeau, E.; Sirois, M.; Quessy, S. Antimicrobial susceptibilities of thermophilic Campylobacter from humans, swine, and chicken broilers. Can. J. Vet. Res. Rev. Can. Rech. Vet. 2006, 70, 81–86. [Google Scholar]
- L-Edany, A.A.A.; Khudor, M.H.; Radhi, L.Y. Isolation, identification and toxigenic aspects of Campylobacter jejuni isolated from slaughtered cattle and sheep at Basrah city. Bas. J. Vet. Res. 2015, 14, 316–327. [Google Scholar]
- Yamazaki-Matsune, W.; Taguchi, M.; Seto, K.; Kawahara, R.; Kawatsu, K.; Kumeda, Y.; Kitazato, M.; Nukina, M.; Misawa, N.; Tsukamoto, T. Development of a multiplex PCR assay for identification of Campylobacter coli, Campylobacter fetus, Campylobacter hyointestinalis subsp. hyointestinalis, Campylobacter jejuni, Campylobacter lari and Campylobacter upsaliensis. J. Med Microbiol. 2007, 56, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Linton, D.; Lawson, A.J.; Owen, R.J.; Stanley, J. PCR detection, identification to species level, and fingerprinting of Campylobacter jejuni and Campylobacter coli direct from diarrheic samples. J. Clin. Microbiol. 1997, 35, 2568–2572. [Google Scholar] [CrossRef] [PubMed]
- Burnett, T.A.; Hornitzky, M.A.; Kuhnert, P.; Djordjevic, S.P. Speciating Campylobacter jejuni and Campylobacter coli isolates from poultry and humans using six PCR-based assays. FEMS Microbiol. Lett. 2002, 216, 201–209. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pajaniappan, M.; Hall, J.E.; Cawthraw, S.A.; Newell, D.G.; Gaynor, E.C.; Fields, J.A.; Rathbun, K.M.; Agee, W.A.; Burns, C.M.; Hall, S.J.; et al. A temperature-regulated Campylobacter jejuni gluconate dehydrogenase is involved in respiration-dependent energy conservation and chicken colonization. Mol. Microbiol. 2008, 68, 474–491. [Google Scholar] [CrossRef]
- Price, L.B.; Johnson, E.; Vailes, R.; Silbergeld, E. Fluoroquinolone-Resistant Campylobacter Isolates from Conventional and Antibiotic-Free Chicken Products. Environ. Health Perspect. 2005, 113, 557–560. [Google Scholar] [CrossRef]
- Obeng, A.; Rickard, H.; Sexton, M.; Pang, Y.; Peng, H.; Barton, M. Antimicrobial susceptibilities and resistance genes in Campylobacter strains isolated from poultry and pigs in Australia. J. Appl. Microbiol. 2012, 113, 294–307. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Bezek, K.; Kurinčič, M.; Knauder, E.; Klančnik, A.; Raspor, P.; Bucar, F.; Možina, S.S. Attenuation of Adhesion, Biofilm Formation and Quorum Sensing of Campylobacter jejuni by Euodia ruticarpa. Phytother. Res. 2016, 30, 1527–1532. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Eloff, J.N. Avoiding pitfalls in determining antimicrobial activity of plant extracts and publishing the results. BMC Complement. Altern. Med. 2019, 19, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Madden, T. The BLAST Sequence Analysis Tool; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2013. [Google Scholar]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Library Code | Family | Scientific Name | Common Name | Collection Site | Collection Date | Part of Plant | Extraction Solvent |
|---|---|---|---|---|---|---|---|
| BE0005B1 | Lamiaceae | Meehania urticifolia (Miq.) Makino | Nettle-leaf mint | Gangneung, Gangwon | 2016 | Aerial part | Ethanol |
| BE0165A1 | Lamiaceae | Scutellaria baicalensis Georgi | Skullcap | Yeosu, Jeonnam | 2017 | Root | Ethanol |
| BE0167A1 | Lamiaceae | Mentha canadensis L. | Wild mint | Andong, Gyeongbuk | 2017 | Aerial part | Ethanol |
| BE1192A1 | Lamiaceae | Salvia plebeia R.Br. | Common sage | Paju, | 2015 | Whole plant | Ethanol |
| Gyeonggi | |||||||
| BEA585A1 | Lauraceae | Cinnamomum cassia (L.) J.Presl | Cinnamon | Gyeongdong Seoul | 2015 | Bark | Ethyl acetate |
| NP/Antibiotic | CJ–RS | CC–RS | CJ–CI | CC–CI | ||||
|---|---|---|---|---|---|---|---|---|
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | |
| M. urticifolia | 400 | 800 | 400 | 800 | 400 | 800 | 400 | 800 |
| S. baicalensis | 400 | 800 | 400 | 800 | 400 | 800 | 400 | 800 |
| M. canadensis | 400 | 800 | 400 | 800 | 800 | 1600 | 400 | 800 |
| S. plebeia | 400 | 800 | 400 | 800 | 800 | 1600 | 400 | 800 |
| C. cassia | 200 | 400 | 200 | 400 | 200 | 400 | 200 | 400 |
| Clove oil | 50 | 100 | 100 | 400 | 50 | 100 | 200 | 400 |
| Cinnamon oil | 25 | 25 | 50 | 100 | 25 | 50 | 50 | 100 |
| Eugenol | 50 | 100 | 100 | 200 | 50 | 100 | 100 | 200 |
| (E)-Cinnamaldehyde | 25 | 25 | 50 | 50 | 25 | 50 | 50 | 50 |
| Baicalein | 32 | 64 | 64 | 64 | 32 | 64 | 64 | 64 |
| Kuraridin | 48 | ND | 48 | ND | 48 | ND | 48 | ND |
| Emodin | 50 | ND | 200 | ND | 50 | ND | 200 | ND |
| Ciprofloxacin | 0.125 | 1 | 0.5 | 1 | 32 | 64 | 64 | 128 |
| Erythromycin | 0.5 | 1 | 1 | 4 | 0.5 | 1 | 2 | 4 |
| Gentamicin | 2 | 8 | 2 | 8 | 1 | 2 | 1 | 8 |
| Tetracycline | 1 | 4 | 1 | 4 | 256 | 512 | 64 | 128 |
| Nalidixic acid | 16 | 32 | 8 | 32 | 128 | 256 | 64 | 128 |
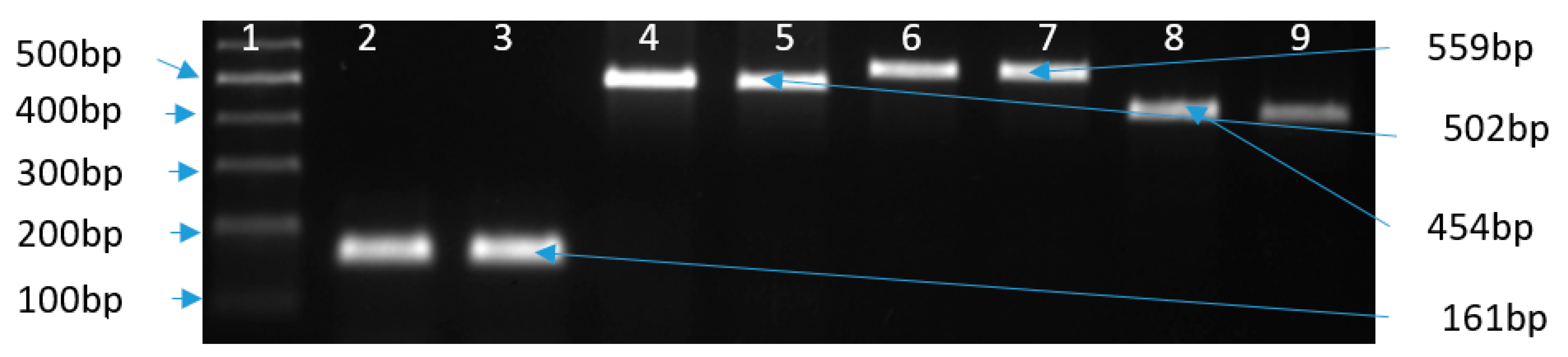
| Target Gene | Primer Name | Sequence (5’–3’) | Amplicon Size | Annealing T (°C) | Reference |
|---|---|---|---|---|---|
| 16S rRNA | C412F | GGATGACACTTTTCGGAGC | 816 | 55 | [73] |
| C1228R | CATTGTAGCACGTGTGTC | ||||
| cj0414 | C1F | CAAATAAAGTTAGAGGTAGAATGT | 161 | ||
| C3R | CCATAAGCACTAGCTAGCTGAT | ||||
| ask | CC18F | GGTATGATTTCTACAAAGCGAG | 502 | ||
| CC519R | ATAAAAGACTATCGTCGCGTG | ||||
| tet(O) | tet(O)F | GCGTTTTGTTTATGTGCG | 559 | [77,78] | |
| tet(O)R | ATGGACAACCCGACAGAAG | ||||
| cjgyrA | QRDRF | GCCTGACGCAAGAGATGGTTTA | 454 | ||
| QRDRR | TATGAGGCGGGATGTTTGTCG | ||||
| cmeB | cmeBF | TCCTAGCAGCACAATATG | 241 | ||
| cmeBR | AGCTTCGATAGCTGCATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gahamanyi, N.; Song, D.-G.; Cha, K.H.; Yoon, K.-Y.; Mboera, L.E.G.; Matee, M.I.; Mutangana, D.; Amachawadi, R.G.; Komba, E.V.G.; Pan, C.-H. Susceptibility of Campylobacter Strains to Selected Natural Products and Frontline Antibiotics. Antibiotics 2020, 9, 790. https://doi.org/10.3390/antibiotics9110790
Gahamanyi N, Song D-G, Cha KH, Yoon K-Y, Mboera LEG, Matee MI, Mutangana D, Amachawadi RG, Komba EVG, Pan C-H. Susceptibility of Campylobacter Strains to Selected Natural Products and Frontline Antibiotics. Antibiotics. 2020; 9(11):790. https://doi.org/10.3390/antibiotics9110790
Chicago/Turabian StyleGahamanyi, Noel, Dae-Geun Song, Kwang Hyun Cha, Kye-Yoon Yoon, Leonard E.G. Mboera, Mecky I. Matee, Dieudonné Mutangana, Raghavendra G. Amachawadi, Erick V.G. Komba, and Cheol-Ho Pan. 2020. "Susceptibility of Campylobacter Strains to Selected Natural Products and Frontline Antibiotics" Antibiotics 9, no. 11: 790. https://doi.org/10.3390/antibiotics9110790
APA StyleGahamanyi, N., Song, D.-G., Cha, K. H., Yoon, K.-Y., Mboera, L. E. G., Matee, M. I., Mutangana, D., Amachawadi, R. G., Komba, E. V. G., & Pan, C.-H. (2020). Susceptibility of Campylobacter Strains to Selected Natural Products and Frontline Antibiotics. Antibiotics, 9(11), 790. https://doi.org/10.3390/antibiotics9110790