Abstract
Pseudomonas aeruginosa, a bacterium commonly isolated from hospital settings, exhibits intrinsic resistance to a number of antibiotics and can acquire resistance during antibiotic therapy. Resistance towards carbapenems is increasing due to its overuse in the treatment of infections caused by extended-spectrum β-lactamase (ESBL) producing organisms. Nonetheless, carbapenems are essential for the treatment of high-risk infections and are one of the remaining weapons in the fight against “extreme drug resistance” of Gram-negative/positive bacilli. Herein, we describe a case report of infections caused by P. aeruginosa strains that carry blaVIM-2 and blaKPC-2 carbapenemase genes simultaneously, identified in five patients who were admitted to a high complexity health institution in Colombia. Molecular characterization included PCR screening for blaKPC, blaGES, blaOXA-48, blaIMP, blaNDM, and blaVIM carbapenemase and other resistance genes as well as analysis of the genetic relationships by genome macro-restriction and Pulsed-Field Gel Electrophoresis (PFGE) separation. In conclusion, these infections represent a major challenge to public health due to the risk of the infection spreading compounded by the fact that limited treatment options are available, thereby increasing the risk of increased morbidity and mortality.
1. Introduction
Pseudomonas aeruginosa, a Gram-negative, non-fermenting, rod-shaped bacterium, has become a significant concern in hospital-acquired infections as it infects immunocompromised patients. Incidences of Pseudomonas aeruginosa infections are on the rise worldwide due to its mechanisms of survival, adaptation, and resistance to different types of antimicrobials [1]. The National Healthcare Safety Network (NHSN) in the United States reported that from 2011 to 2014, P. aeruginosa was the sixth most common cause of hospital-acquired infections at 7.3% of all cases. The NHSN also reported that P. aeruginosa was the second most common cause of ventilator-associated pneumonia (VAP) (16.5%), as well as the most common multidrug-resistant (MDR) Gram-negative pathogen causing VAP. Furthermore, P. aeruginosa was also implicated in 10.3% of all catheter-associated urinary tract infections and 5.7% of all surgical site infections [2,3]. P. aeruginosa is a common pathogen worldwide and is one of the five most commonly isolated bacteria in hospitals in Colombia and other regions of Latin America [4,5]. The fact that P. aeruginosa is both intrinsically resistant and can acquire resistance to a number of antibiotics during therapy limits the available therapeutic options. Therefore, knowledge of the local resistance patterns is essential in order to establish the appropriate treatment strategies [3,4].
P. aeruginosa has multiple antibiotic resistance mechanisms that have been described as intrinsic, acquired, and adaptive [6]. Acquired resistance can occur as a result of mutation(s) or acquisition of exogenous resistance determinants and can be mediated by a number of mechanisms, including enzyme degradation, reduced permeability, and active efflux [7]. Intrinsic resistance is conferred by inherent structural or functional characteristics such as low outer membrane permeability, efflux of antimicrobials, and the production of antibiotic-inactivating enzymes [6]. Adaptive resistance, on the other hand, affects the lungs of patients via the formation of biofilms that serve as a barrier against antimicrobial infiltration [8].
Lasmid-mediated extended spectrum β-lactamases (ESBLs) have been implicated in acquired resistance owing to enzyme degradation, the most commonly described antimicrobial resistance mechanism. Temoneira (TEM), Sulfhydryl reagent variable (SHV), and cefotaximase (CTX-M) ESBLs have been reported in P. aeruginosa. Vietnam extended-spectrum β-lactamase (VEB) ESBLs are prevalent in P. aeruginosa strains in East Asia and are now also found in other regions. Pseudomonas extended resistant (PER) ESBLs, widely found in Turkey, confers a high-level of resistance on antipseudomonal cephalosporins [9,10]. Carbapenem resistance in P. aeruginosa can be a result of mutations, resulting in the loss of the OprD porin, but may also be a result of the production of carbapenemases such as Guiana Extended spectrum (GES), Imipenem metallo-β-lactamase (IMP), Verona Integron-encoded metallo-β-lactamase (VIM), Sao Paulo metallo-β-lactamase (SPM), and more recently, the K. pneumoniae carbapenemase (KPC) and New Delhi metallo-β-lactamase (NDM) [9,11].
KPC, a class A carbapenemase, was initially isolated from K. pneumoniae and has also been detected in most Enterobacteria [12]. However, in 2007, a P. aeruginosa isolate harboring the blaKPC-2 gene was identified in Colombia [13], and there have since been additional reports of such isolates in other countries [14,15,16,17,18]. The blaKPC gene is mobilized on the 10 kb active Tn3-family Tn4401 transposon, which is delimited by two 39-bp inverted repeat sequences [19]. The co-presence of blaVIM-2 and blaKPC-2 genes has been more frequently reported in the species of K. pneumonia [20] compared to P. aeruginosa, of which only three reports are available. The first report of the co-expression of blaVIM-2 and blaKPC-2 in P. aeruginosa occurred in Colombia in 2012 [21,22], followed by Chile [23], and later in Puerto Rico. It must be noted that in the latter, the P. aeruginosa isolate harbored KPC and IMP-8, simultaneously [24].
Here we report, for the first time, a case series of P. aeruginosa harboring VIM and KPC concurrently, producing two carbapenemases that represent a major public health challenge due to the risk of their successful dissemination and the limited classes of antibiotics that can be used for the treatment of these multi-drug resistant (MDR) isolates.
2. Results
2.1. Case 1
A 66-year-old patient was diagnosed with abdominal sepsis secondary to mesenteric ischemia. The patient was treated with piperacillin/tazobactam, meropenem, and fluconazole and required ventilatory support as well as a vasopressor. Due to poor clinical evolution, the patient required peritoneal lavage, which resulted in the isolation of MDR P. aeruginosa in the peritoneal fluid. Colistimethate (2,700,000 UI IV every eight h) was prescribed with follow-up of renal function. After four days of antibiotic therapy, the patient presented with clinical deterioration and cardiorespiratory arrest.
2.2. Case 2
The patient was 56 years old with a polytrauma Injury Severity Score (ISS) of 24 secondaries due to a high-energy traffic accident as an automobile driver, who suffered a complete sub-trochanter fracture of the left femur. After an initial clinical deterioration, the patient required an external tutor. During this surgery, in order to test for bone necrosis, the surgeon sent a bone sample for laboratory testing which was found to contain MDR P. aeruginosa. Consequently, broad-spectrum management with colistimethate (2,000,000 UI IV every eight h), rifampicin (600 mg once daily), and doripenem (1 g every eight h) was initiated. The patient completed antibiotic treatment and in-home hospitalization for 42 days.
2.3. Case 3
The patient was 84 years old with a history of Wegener′s disease, a right hip replacement, and infection at the operative site by Proteus mirabilis that was treated with meropenem for 21 days. A surgical lavage was performed due to the patient’s clinical decline resulting in the identification of carbapenem-resistant P. aeruginosa in bone and blood cultures. Antibiotic treatment was started with colistimethate (2,000,000 UI IV every eight h), doripenem (1 g every eight h), and rifampicin (600 mg every 12 h) for 42 days, and the hip implant was also removed. On day 10 of the treatment, the patient showed clinical deterioration and later died.
2.4. Case 4
The patient was 57 years old with a history of benign prostatic hyperplasia necessitating a long-term urinary catheter, with complicated diverticular disease that required subtotal colectomy. During hospitalization, the patient required multiple invasive medical devices and approximately seven surgical lavages. The patient presented bacteraemia caused by P. aeruginosa and received treatment with colistimethate (2,400,000 UI IV every eight h) and doripenem (1 g every eight h) for 10 days with favorable clinical evolution and negative control cultures. The patient was discharged after two weeks of hospitalization.
2.5. Case 5
A 29-year-old patient with a history of epilepsy and Down syndrome presented with pulmonary septic shock due to carbapenemase-producing P. aeruginosa that was treated with colistimethate and doripenem. Following clinical improvement, the patient was discharged after 10 days but then was readmitted at day 15 with a systemic inflammatory response and deterioration of the respiratory pattern. Cultures showed a urine culture with MDR P. aeruginosa. Colistimethate (1,500,000 UI IV every eight h), doripenem (1 g every eight h), and Fosfomycin were prescribed for 12 days. The patient presented with clinical deterioration and died during hospitalization. The most important characteristics of the five patients are shown in Table 1.

Table 1.
Relevant clinical characteristics of patients.
All five isolates were resistant to meropenem, imipenem, gentamicin, ciprofloxacin, trimethoprim/sulfamethoxazole, and piperacillin/tazobactam but remained susceptible to colistin (0.5 µg/mL). Analysis of the meropenem and doripenem minimal inhibitory concentration (MIC) showed that all isolates reached a MIC value of 1024 µg/mL and 512 µg/mL, respectively. Molecular characterization revealed that the five isolates simultaneously harbored the blaVIM-2 and blaKPC-2 carbapenemase genes, and the blaTEM and aac(6’)-lb genes were also detected. In P. aeruginosa, the blaKPC-2 gene is mainly found on both complete and truncated Tn4401b transposons, within two different plasmid backbones (IncU and IncP-6 incompatibility groups). We recently reported a P. aeruginosa isolate (24Pae112) that contained a double chromosomal insertion of the Tn4401b-blaKPC-2 transposon, which was inserted into the new pathogenicity island (PAGI-17). We designed primers to amplify specific DNA fragments of these three genetic platforms (see Materials and Methods); however, they were not identified in the P. aeruginosa isolates, suggesting that the blaKPC-2 gene could be mobilized in a different platform.
The Pulsed-Field Gel Electrophoresis analysis revealed that the five isolates had an identical pulsotype, but they were different from the 24Pae112 isolate pulsotype (Figure 1), suggesting that this carbapenemase was acquired through unrelated clones. The intl1 gene and the presence of blaVIM gene into the class I integron was confirmed by PCR.
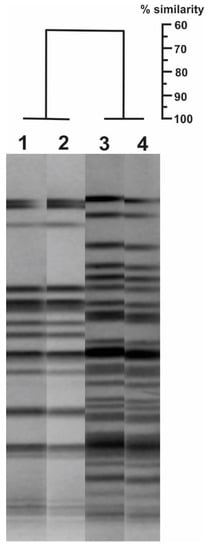
Figure 1.
Genetic relationship by Pulsed-Field Gel Electrophoresis (PFGE) of the Pseudomonas aeruginosa isolates that harbor the double chromosomal insertion of the blaKPC-2 transposon (lanes 1 and 2) and those carrying the blaVIM-2 and blaKPC-2 genes simultaneously (lanes 3 and 4). GelCompar II program (Applied Maths NV) was used, with a tolerance position of 1.5% and a Dice coefficient of 1.0%.
3. Discussion
Herein, we present a series of five intensive care unit (ICU) hospitalized patients with MDR P. aeruginosa infections with a very high MIC to carbapenems and harboring the blaKPC-2 and blaVIM-2 genes at the same time. Patients ranged in age from 29 to 84 years, and the majority were men with diverse comorbidities as well as different sites of infection. Only two patients shared a bone infection, and all treatments followed the institutional recommendation for each case. All patients received an antibiotic combination of two or three drugs with all five regimens including colistimethate. The mortality was high (60%). Bacterial strains producing two carbapenemases represent a major public health challenge due to the risk of their successful spread and difficulty of treating the infections caused by these MDR isolates [25]. To the best of our knowledge, there are no case series reports regarding this significant issue in P. aeruginosa.
Co-harboring of carbapenemases is a genetic event that, in recent years, has increased in its frequency due to the increased clinical usage of carbapenems. The co-expression of VIM and KPC enzymes has been reported more frequently in species of K. pneumoniae [22,26,27]. However, the first P. aeruginosa isolate co-harboring VIM-2 and KPC-2 was reported in Colombia in 2012 and later in Chile [23]. P. aeruginosa isolates harboring KPC and IMP-8 simultaneously were found in Puerto Rico [24]. In Colombia, the circulation of the blaKPC-2-containing P. aeruginosa isolates initiated a national public health problem because such isolates have increased in their frequency since 2007 [23]. In 2012, an analysis of 43 carbapenemase-producing P. aeruginosa isolates recovered from seven Colombian cities showed that there was a higher frequency of isolates with blaVIM-2 with respect to those with blaKPC-2 (33 vs. 9) [28]. In 2014 and 2015, two studies found a similar frequency of both the blaKPC-2- and blaVIM-2-containing P. aeruginosa isolates [20,24]. A recent study conducted in seven healthcare institutions in Bogota, Colombia, found that the blaKPC-2-containing P. aeruginosa isolates were the most frequent (4:1 ratio between blaKPC-2 and blaVIM-2, respectively) (data in publication process). Currently, the spread of P. aeruginosa co-expressing KPC and VIM presents a significant public health challenge.
The global spread of carbapenem resistance among Gram-negative organisms is explained by horizontal gene transfer, although the first carbapenemases described were chromosomally encoded and species-specific [29]. Latin America is not immune to this dissemination. Limited resources for performing the appropriate microbiological assays in the vast majority of clinical laboratories lead to an underestimation of the real problem [22]. A recent review of the epidemiology of carbapenemases in Latin America and the Caribbean identified an increased frequency of reports in both regions. This clearly illustrates the ability of these enzymes to successfully spread, becoming endemic in some countries [30].
Therapeutic options for patients with MDR bacterial infections are scarce. Treatment options include carbapenems with lower MIC and adequate penetrance to the site of infection, tigecycline, fosfomycin, amikacin, polymyxins, and some authors recommend rifampicin and daptomycin [31]. Recent advances include ceftazidime/avibactam, which was not available for use in Colombia when isolates for this study were collected. In this regard, the use of the latter is limited by the presence of metallo-β-lactamases [32], as in our cases.
These findings underscore the importance of conducting campaigns for preventing the spread of these types of carbapenemase-producing pathogens not just in our institution, but in healthcare facilities in Colombia and Latin America, especially because of their rapid dissemination.
4. Materials and Methods
4.1. Bacterial Isolates and Susceptibility Profile
The P. aeruginosa isolates were recovered from different samples using standard microbiological techniques [33] and were stored in Brain Heart Infusion (BHI) broth (Oxoid-Thermo Scientific®, Hampshire, United Kingdom) supplemented with 15% glycerol at −80 °C until use. Bacterial identification and the susceptibility profiles to meropenem, imipenem, ceftazidime, gentamicin, amikacin, ciprofloxacin, trimethoprim/sulfamethoxazole, and piperacillin/tazobactam were determined by automated VITEK®2 systems using the breakpoints defined by the Clinical and Laboratory Standards Institute, 2018 [34]. The P. aeruginosa (ATCC® 27853TM) strain was used as a susceptibility control (American Type Culture Collection, https://www.atcc.org/products/all/27853.aspx). The MIC to meropenem and colistin was established by the broth dilution method.
4.2. Detection of Resistance Genes
The blaTEM, blaSHV, blaCTX-M, blaFOX, blaACT, blaMIR blaACC, blaDHA, blaCMY, and blaMOX genes were assessed using two multiplex PCR described previously [35]. The blaIMP, blaOXA-48, blaVIM, blaGES, blaKPC, blaNDM carbapenemase genes were assessed by multiplex PCR in accordance with previously reported conditions [36]. The OXA-derived carbapenemase genes (blaOXA-23, blaOXA-24, blaOXA-43, blaOXA-51, and blaOXA-58) were also assessed by multiple PCR [37,38]. In addition, a screening of genes related to aminoglycoside and fluoroquinolone resistance was performed (aac(6´)-lb, aac(6´)-lb-cr, qnrA, qnrB, qnrS, and mcbG) [39]. The blaKPC-2 and blaVIM-2 variants were determined by sequencing of complete genes using the dideoxy chain termination method [40]. Finally, the intl1, intl2, and intl3 genes were assessed using primers previously reported [41] (Gene ID: 13906549 for intl1 gene, and GenBank accession number: KJ184348.1 and BBA94100.1 for intl2 and intl3 genes, respectively).
4.3. Establishment of the Genetic Relationship by Pulsed-Field Gel Electrophoresis (PFGE)
The genetic correlation between the isolates was determined by genome macro-restriction using the SpeI enzyme (Promega, Madison WI, USA) and PFGE separation according to previously reported methods [42]. Briefly, one colony of each isolate was grown in 5 mL of BHI broth at 37 °C for 12 h (2 × 108 cells). The bacteria were harvested by centrifugation, washed twice, and resuspended in 2 mL of TE buffer (0.2 M Tris-HCl, 20 mM EDTA (pH 7.5). Then 200 µL of bacterial suspension were mixed with 200 µL of a 1.5% agarose solution and deposited into a casting mold. The embedded cells in the agarose inserts were subjected to detergent and enzymatic lysis (50 mM Tris, 50 mM EDTA-pH 8.0, 1% Sarcosyl, and 400 µg proteinase K). The insert was washed and stored in TE buffer at 4 °C until it was ready to use. The DNA-containing agarose slices (2 mm) were subjected to digestion using 12 U SpeI enzyme (Promega) in buffer Tango® at 37 °C for five h. The DNA-fragments generated were separated using a CHEFII Pulsed Field Electrophoresis system (Biorad, California CA, USA) for 23 h at 14 °C, 120 V (6 V/cm), and with an initial and final time of 6.8 s and 35.4 s, respectively.
4.4. Assessment of the Genetic Platforms Mobilizing the Blakpc Gene
Currently, three genetic platforms have been reported mobilizing the blaKPC-2-positive Tn4401b transposon in P. aeruginosa, two different plasmid backbones (8 kb and 31.5 kb) belonging to the IncU and IncP-6 incompatibility groups [43,44], and a double chromosomal insertion within a new genomic island named PAGI-17 [45]. The specific primers were designed to amplify DNA fragments for each platform (Table 2). The optimal conditions for each PCR were established.
Table 2.
List of primers used in this study.
4.5. Ethical Approvals
Isolates were obtained as part of routine diagnostic testing and analyzed anonymously. The study was approved by the Research Ethics Committee of the Faculty of Medicine at Universidad de La Sabana (Acta 410, 9 June 2017).
5. Conclusions
We report for the first time a case series of infections caused by P. aeruginosa that concurrently harbor VIM and KPC. This represents a major public health problem considering the risk of an outbreak of such an infection in combination with the fact that we have limited therapeutic tools to treat such infections could be calamitous.
Author Contributions
Conceptualization—J.-C.G., R.H.B.-C., T.P. and J.E.-P.; Methodology—T.P., D.A. and J.E.-P.; Investigation—T.P., D.A. and J.E.-P.; Formal Analysis—J.R. and T.P.; Resources—R.H.B.-C., J.-C.G. and J.R.; Data Curation—J.R.; Writing and Original Draft Preparation—T.P., S.A., L.U., J.R. and R.H.B.-C.; Writing, Review, and Editing—T.P. and R.H.B.-C.; Supervision—R.H.B.-C. and J.-C.G.
Funding
This work was funded by the Universidad de la Sabana (MED-222-2017), Universidad El Bosque, and partially by the Departamento Administrativo de Ciencia, Tecnologıa e Innovacion, Colciencias (grant number 1308-71250819).
Acknowledgments
The authors would like to thank all the members of the Clínica Universidad de La Sabana and Bacterial Molecular Genetics Laboratory at Universidad El Bosque.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Moradali, M.F.; Ghods, S.; Rehm, B.H. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef]
- Weiner, L.M.; Webb, A.K.; Limbago, B.; Dudeck, M.A.; Patel, J.; Kallen, A.J.; Edwards, J.R.; Sievert, D.M. Antimicrobial-Resistant Pathogens Associated with Healthcare-Associated Infections: Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011–2014. Infect. Control Hosp. Epidemiol. 2016, 37, 1288–1301. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, I.; Hackel, M.; Badal, R.; Bouchillon, S.; Hawser, S.; Biedenbach, D. A Review of Ten Years of the Study for Monitoring Antimicrobial Resistance Trends (SMART) from 2002 to 2011. Pharmaceuticals 2013, 6, 1335–1346. [Google Scholar] [CrossRef] [PubMed]
- Gales, A.C.; Castanheira, M.; Jones, R.N.; Sader, H.S. Antimicrobial resistance among Gram-negative bacilli isolated from Latin America: Results from SENTRY Antimicrobial Surveillance Program (Latin America, 2008–2010). Diagn. Microbiol. Infect. Dis. 2012, 73, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, A.; Duarte, C.; Gonzalez, N.; Saavedra, S. Co-producciones de carbapenemasas un fenómeno en aumento y de difícil detección en el laboratorio de microbiología con pruebas fenotípicas. Boletín Informativo GREBO 2016, 8, 36–39. [Google Scholar]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Rossolini, G.M.; Mantengoli, E. Treatment and control of severe infections caused by multiresistant Pseudomonas aeruginosa. Clin. Microbiol. Infect. 2005, 11, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Drenkard, E. Antimicrobial resistance of Pseudomonas aeruginosa biofilms. Microbes Infect. 2003, 5, 1213–1219. [Google Scholar] [CrossRef]
- Livermore, D.M. Has the era of untreatable infections arrived? J. Antimicrob. Chemother. 2009, 64, i29–i36. [Google Scholar] [CrossRef]
- Jacoby, G.A. Beta-lactamase nomenclature. Antimicrob. Agents Chemother. 2006, 50, 1123–1129. [Google Scholar] [CrossRef]
- Poole, K. Pseudomonas aeruginosa: Resistance to the max. Front. Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef]
- Nordmann, P.; Dortet, L.; Poirel, L. Carbapenem resistance in Enterobacteriaceae: Here is the storm! Trends Mol. Med. 2012, 18, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Villegas, M.V.; Lolans, K.; Correa, A.; Kattan, J.N.; Lopez, J.A.; Quinn, J.P.; Colombian Nosocomial Resistance Study Group. First identification of Pseudomonas aeruginosa isolates producing a KPC-type carbapenem-hydrolyzing beta-lactamase. Antimicrob. Agents Chemother. 2007, 51, 1553–1555. [Google Scholar] [CrossRef] [PubMed]
- Akpaka, P.E.; Swanston, W.H.; Ihemere, H.N.; Correa, A.; Torres, J.A.; Tafur, J.D.; Montealegre, M.C.; Quinn, J.P.; Villegas, M.V. Emergence of KPC-producing Pseudomonas aeruginosa in Trinidad and Tobago. J. Clin. Microbiol. 2009, 47, 2670–2671. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Nordmann, P.; Lagrutta, E.; Cleary, T.; Munoz-Price, L.S. Emergence of KPC-producing Pseudomonas aeruginosa in the United States. Antimicrob. Agents Chemother. 2010, 54, 3072. [Google Scholar] [CrossRef] [PubMed]
- Robledo, I.E.; Aquino, E.E.; Vazquez, G.J. Detection of the KPC gene in Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Acinetobacter baumannii during a PCR-based nosocomial surveillance study in Puerto Rico. Antimicrob. Agents Chemother. 2011, 55, 2968–2970. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.; Wei, Z.; Jiang, Y.; Shen, P.; Yu, Y.; Li, L. Identification of KPC-2-producing Pseudomonas aeruginosa isolates in China. J. Antimicrob. Chemother. 2011, 66, 1184–1186. [Google Scholar] [CrossRef]
- Jacome, P.R.; Alves, L.R.; Cabral, A.B.; Lopes, A.C.; Maciel, M.A. First report of KPC-producing Pseudomonas aeruginosa in Brazil. Antimicrob. Agents Chemother. 2012, 56, 4990. [Google Scholar] [CrossRef]
- Cuzon, G.; Naas, T.; Nordmann, P. Functional characterization of Tn4401, a Tn3-based transposon involved in blaKPC gene mobilization. Antimicrob. Agents Chemother. 2011, 55, 5370–5373. [Google Scholar] [CrossRef]
- Falco, A.; Ramos, Y.; Franco, E.; Guzman, A.; Takiff, H. A cluster of KPC-2 and VIM-2-producing Klebsiella pneumoniae ST833 isolates from the pediatric service of a Venezuelan Hospital. BMC Infect. Dis. 2016, 16, 595. [Google Scholar] [CrossRef]
- Correa, A.; Montealegre, M.C.; Mojica, M.F.; Maya, J.J.; Rojas, L.J.; De La Cadena, E.P.; Ruiz, S.J.; Recalde, M.; Rosso, F.; Quinn, J.P.; et al. First report of a Pseudomonas aeruginosa isolate coharboring KPC and VIM carbapenemases. Antimicrob. Agents Chemother. 2012, 56, 5422–5423. [Google Scholar] [CrossRef]
- Vanegas, J.M.; Cienfuegos, A.V.; Ocampo, A.M.; Lopez, L.; del Corral, H.; Roncancio, G.; Sierra, P.; Echeverri-Toro, L.; Ospina, S.; Maldonado, N.; et al. Similar frequencies of Pseudomonas aeruginosa isolates producing KPC and VIM carbapenemases in diverse genetic clones at tertiary-care hospitals in Medellin, Colombia. J. Clin. Microbiol. 2014, 52, 3978–3986. [Google Scholar] [CrossRef]
- Kazmierczak, K.M.; Biedenbach, D.J.; Hackel, M.; Rabine, S.; de Jonge, B.L.; Bouchillon, S.K.; Sahm, D.F.; Bradford, P.A. Global Dissemination of blaKPC into Bacterial Species beyond Klebsiella pneumoniae and In Vitro Susceptibility to Ceftazidime-Avibactam and Aztreonam-Avibactam. Antimicrob. Agents Chemother. 2016, 60, 4490–4500. [Google Scholar] [CrossRef]
- Martinez, T.; Vazquez, G.J.; Aquino, E.E.; Ramirez-Ronda, R.; Robledo, I.E. First report of a Pseudomonas aeruginosa clinical isolate co-harbouring KPC-2 and IMP-18 carbapenemases. Int. J. Antimicrob. Agents 2012, 39, 542–543. [Google Scholar] [CrossRef]
- Correa, A.; Del Campo, R.; Perenguez, M.; Blanco, V.M.; Rodriguez-Banos, M.; Perez, F.; Maya, J.J.; Rojas, L.; Canton, R.; Arias, C.A.; et al. Dissemination of high-risk clones of extensively drug-resistant Pseudomonas aeruginosa in Colombia. Antimicrob. Agents Chemother. 2015, 59, 2421–2425. [Google Scholar] [CrossRef]
- Cuzon, G.; Naas, T.; Truong, H.; Villegas, M.V.; Wisell, K.T.; Carmeli, Y.; Gales, A.C.; Venezia, S.N.; Quinn, J.P.; Nordmann, P. Worldwide diversity of Klebsiella pneumoniae that produce beta-lactamase blaKPC-2 gene. Emerg. Infect. Dis. 2010, 16, 1349–1356. [Google Scholar] [CrossRef]
- Giakkoupi, P.; Pappa, O.; Polemis, M.; Vatopoulos, A.C.; Miriagou, V.; Zioga, A.; Papagiannitsis, C.C.; Tzouvelekis, L.S. Emerging Klebsiella pneumoniae isolates coproducing KPC-2 and VIM-1 carbapenemases. Antimicrob. Agents Chemother. 2009, 53, 4048–4050. [Google Scholar] [CrossRef]
- Saavedra, S.; Duarte, C.; Gonzalez, N.; Realpe, M. Caracterización de aislamientos de Pseudomonas aeruginosa productores de carbapenemasas de siete departamentos de Colombia. Biomedica 2014, 34, 217–223. [Google Scholar] [CrossRef][Green Version]
- Patel, G.; Bonomo, R.A. “Stormy waters ahead”: Global emergence of carbapenemases. Front. Microbiol. 2013, 4, 48. [Google Scholar] [CrossRef]
- Escandon-Vargas, K.; Reyes, S.; Gutierrez, S.; Villegas, M.V. The epidemiology of carbapenemases in Latin America and the Caribbean. Expert Rev. Anti-Infect. Ther. 2017, 15, 277–297. [Google Scholar] [CrossRef]
- Plant, A.J.; Dunn, A.; Porter, R.J. Ceftolozane-tazobactam resistance induced in vivo during the treatment of MDR Pseudomonas aeruginosa pneumonia. Expert Rev. Anti-Infect. Ther. 2018, 16, 367–368. [Google Scholar] [CrossRef]
- Zasowski, E.J.; Rybak, J.M.; Rybak, M.J. The beta-Lactams Strike Back: Ceftazidime-Avibactam. Pharmacotherapy 2015, 35, 755–770. [Google Scholar] [CrossRef]
- Leber, A.L. Clinical Microbiology Procedures Handbook, 4th ed.; American Society of Microbiology: Washington, DC, USA, 2016. [Google Scholar]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. 2018. Available online: https://clsi.org/media/1928/m07ed11_sample.pdf (accessed on 5 May 2019).
- Monstein, H.J.; Ostholm-Balkhed, A.; Nilsson, M.V.; Nilsson, M.; Dornbusch, K.; Nilsson, L.E. Multiplex PCR amplification assay for the detection of blaSHV, blaTEM and blaCTX-M genes in Enterobacteriaceae. APMIS 2007, 115, 1400–1408. [Google Scholar] [CrossRef]
- Monteiro, J.; Widen, R.H.; Pignatari, A.C.; Kubasek, C.; Silbert, S. Rapid detection of carbapenemase genes by multiplex real-time PCR. J. Antimicrob. Chemother. 2012, 67, 906–909. [Google Scholar] [CrossRef]
- Woodford, N.; Ellington, M.J.; Coelho, J.M.; Turton, J.F.; Ward, M.E.; Brown, S.; Amyes, S.G.; Livermore, D.M. Multiplex PCR for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrob. Agents 2006, 27, 351–353. [Google Scholar] [CrossRef]
- Higgins, P.G.; Lehmann, M.; Seifert, H. Inclusion of OXA-143 primers in a multiplex polymerase chain reaction (PCR) for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrob. Agents 2010, 35, 305. [Google Scholar] [CrossRef]
- Robicsek, A.; Strahilevitz, J.; Sahm, D.F.; Jacoby, G.A.; Hooper, D.C. QNR prevalence in ceftazidime-resistant Enterobacteriaceae isolates from the United States. Antimicrob. Agents Chemother. 2006, 50, 2872–2874. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Marquez, C.; Labbate, M.; Raymondo, C.; Fernandez, J.; Gestal, A.M.; Holley, M.; Borthagaray, G.; Stokes, H.W. Urinary tract infections in a South American population: Dynamic spread of class 1 integrons and multidrug resistance by homologous and site-specific recombination. J. Clin. Microbiol. 2008, 46, 3417–3425. [Google Scholar] [CrossRef]
- Herschleb, J.; Ananiev, G.; Schwartz, D.C. Pulsed-field gel electrophoresis. Nat. Protoc. 2007, 2, 677–684. [Google Scholar] [CrossRef]
- Naas, T.; Bonnin, R.A.; Cuzon, G.; Villegas, M.V.; Nordmann, P. Complete sequence of two KPC-harbouring plasmids from Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2013, 68, 1757–1762. [Google Scholar] [CrossRef]
- Dai, X.; Zhou, D.; Xiong, W.; Feng, J.; Luo, W.; Luo, G.; Wang, H.; Sun, F.; Zhou, X. The IncP-6 Plasmid p10265-KPC from Pseudomonas aeruginosa Carries a Novel DeltaISEc33-Associated bla KPC-2 Gene Cluster. Front. Microbiol. 2016, 7, 310. [Google Scholar] [CrossRef]
- Abril, D.; Marquez-Ortiz, R.A.; Castro-Cardozo, B.; Moncayo-Ortiz, J.I.; Olarte Escobar, N.M.; Corredor Rozo, Z.L.; Reyes, N.; Tovar, C.; Sanchez, H.F.; Castellanos, J.; et al. Genome plasticity favours double chromosomal Tn4401b-blaKPC-2 transposon insertion in the Pseudomonas aeruginosa ST235 clone. BMC Microbiol. 2019, 19, 45. [Google Scholar] [CrossRef]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).