Unraveling Nutritional Regulation of Tacrolimus Biosynthesis in Streptomyces tsukubaensis through omic Approaches


Abstract
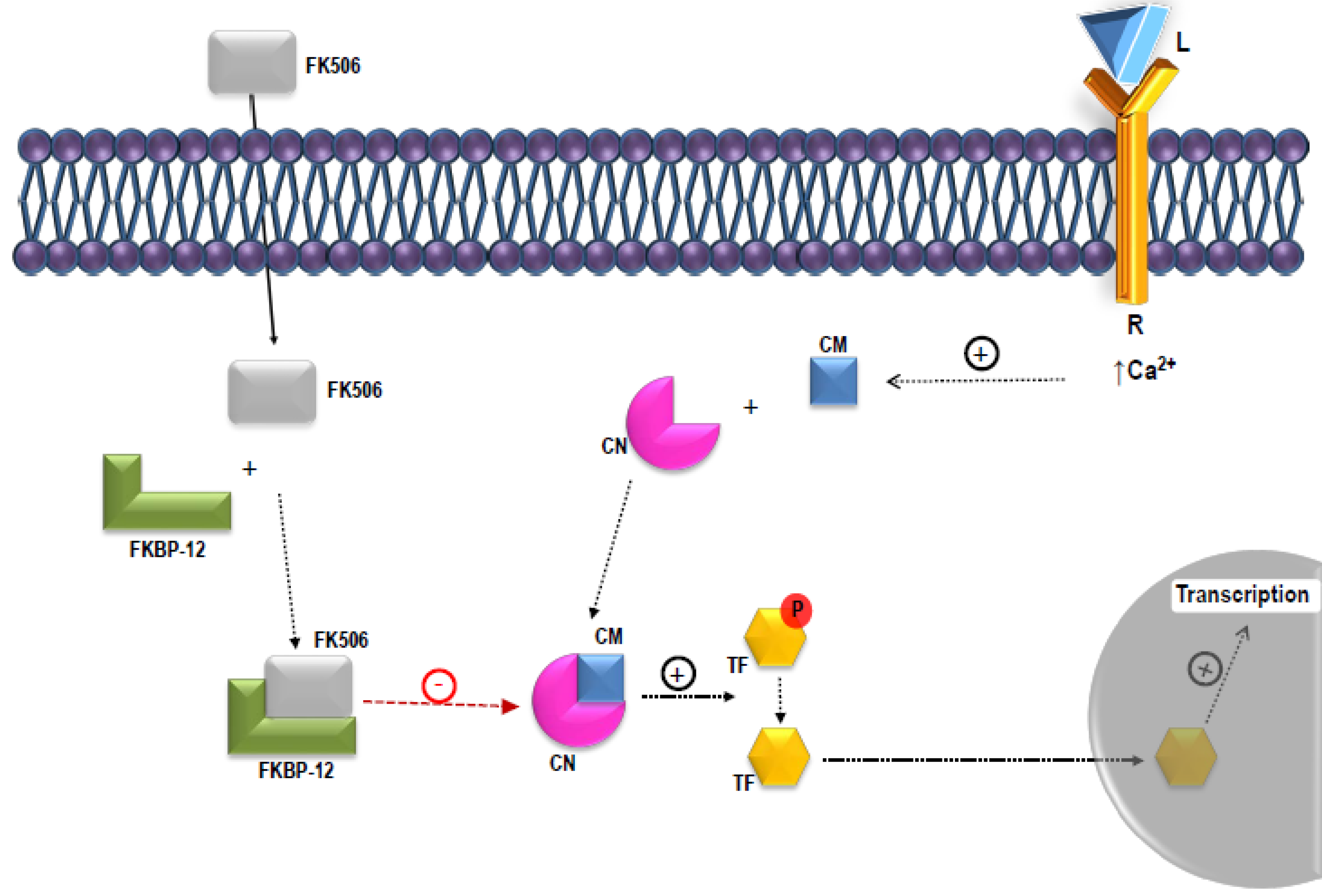
1. Discovery of S. tsukubaensis and Tacrolimus Use in Current Clinics
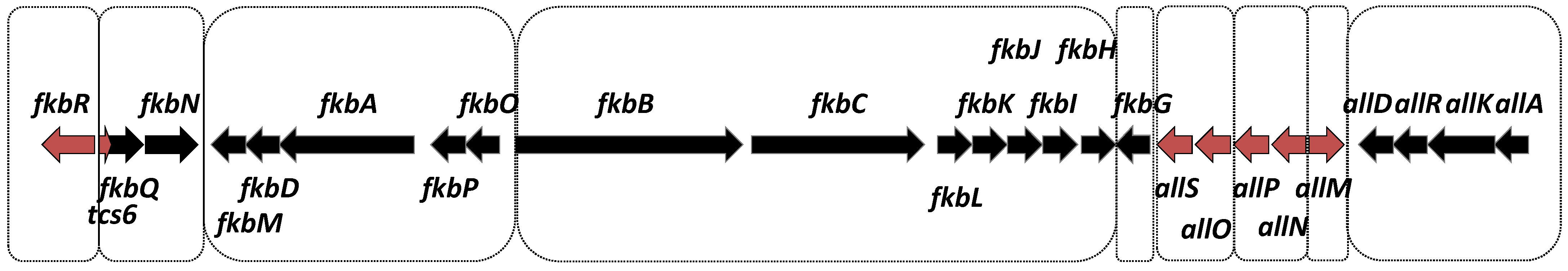
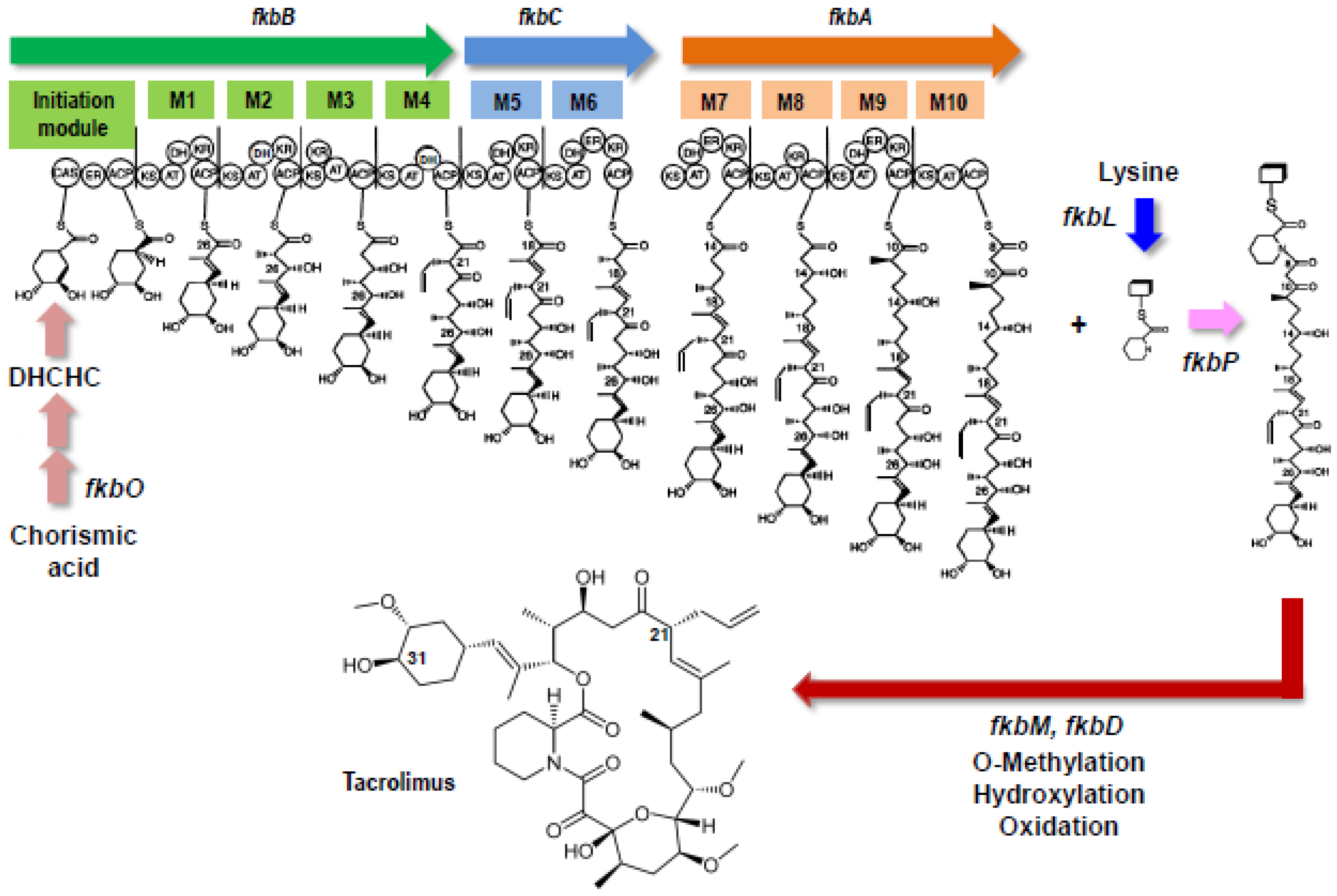
2. Biosynthetic Pathway and Gene Cluster
3. Transcriptional Regulators and Recent Insights through Transcriptomic and RNAseq Studies
3.1. Characterization of fkb Cluster Transcriptional Subunits
3.2. Genes Located Outside of the Tacrolimus Gene Cluster Regulated by FkbN
4. Classical Strategies to Increase Tacrolimus Production
5. Omic Approaches in S. Tsukubaensis and Their Application in Tacrolimus Production
5.1. Metabolomic and Proteomic Studies
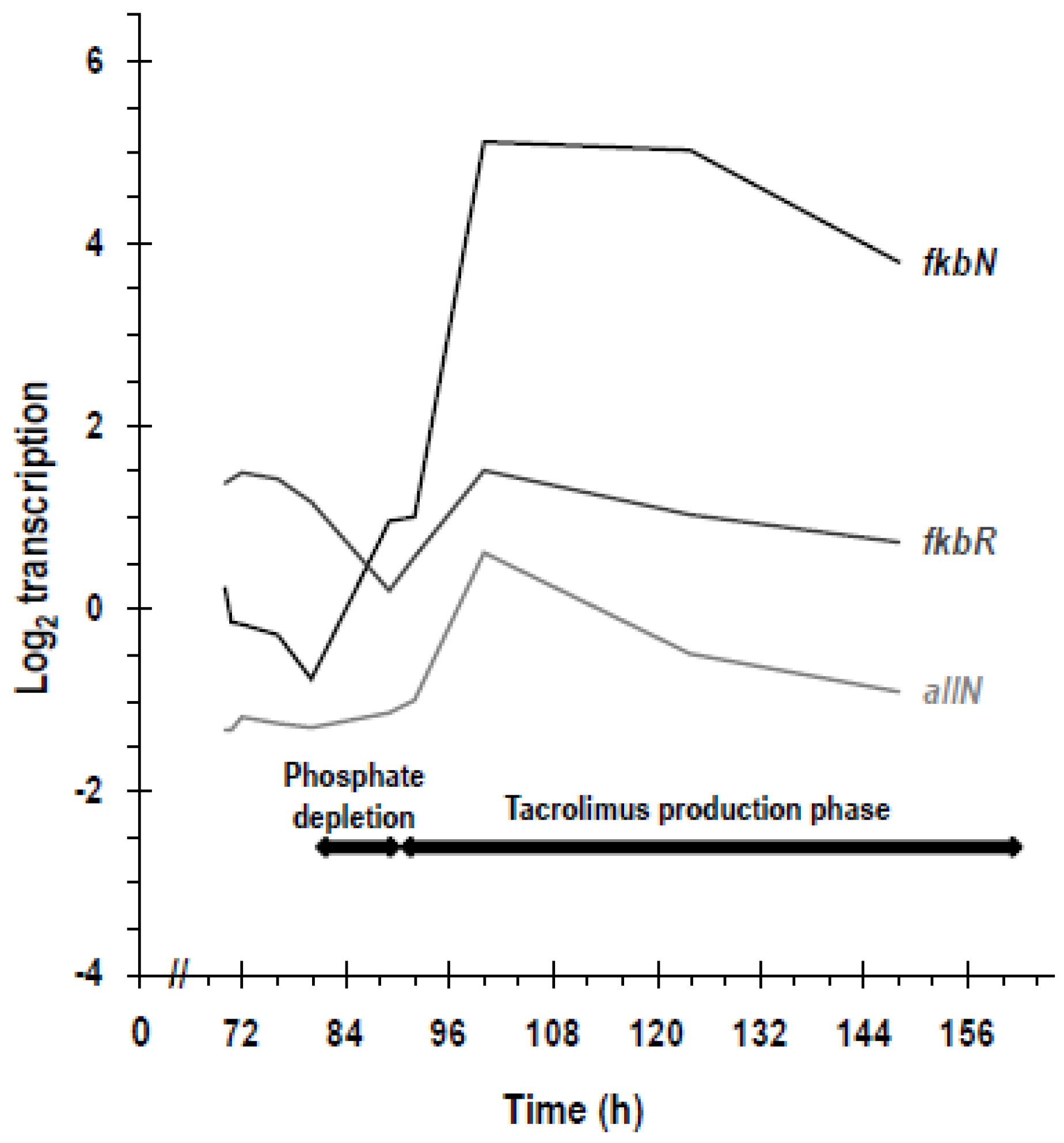
5.2. Transcriptomic Studies on Phosphate Regulation of the fkb Cluster
5.2.1. Transcriptomics of Carbon Catabolite Regulation of Tacrolimus Biosynthesis
5.2.2. Transcriptomics of N-acetylglucosamine Addition in Tacrolimus Biosynthesis
6. Conclusions and Future Prospective
Author Contributions
Acknowledgements
Conflicts of interest
References
- Kino, T.; Hatanaka, H.; Hashimoto, M.; Nishiyama, M.; Goto, T.; Okuhara, M.; Kohsaka, M.; Aoki, H.; Imanaka, H. FK-506, a novel immunosuppressant isolated from a Streptomyces. I. Fermentation, isolation, and physico-chemical and biological characteristics. J. Antibiot. 1987, 40, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Kino, T.; Hatanaka, H.; Miyata, S.; Inamura, N.; Nishiyama, M.; Yajima, T.; Goto, T.; Okuhara, M.; Kohsaka, M.; Aoki, H. FK-506, a novel immunosuppressant isolated from a Streptomyces. II. Immunosuppressive effect of FK-506 in vitro. J. Antibiot. 1987, 40, 1256–1265. [Google Scholar] [CrossRef] [PubMed]
- Wallemacq, P.E.; Reding, R. FK506 (tacrolimus), a novel immunosuppressant in organ transplantation: Clinical, biomedical, and analytical aspects. Clin. Chem. 1993, 39, 2219–2228. [Google Scholar] [PubMed]
- Liu, J.; Farmer, J., Jr.; Lane, W.S.; Friedman, J.; Weissman, I.; Schreiber, S.L. Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. Cell 1991, 66, 807–815. [Google Scholar] [CrossRef]
- Harding, M.W.; Galat, A.; Uehling, D.E.; Schreiber, S.L. A receptor for the immunosuppressant FK506 is a cis-trans peptidyl-prolyl isomerase. Nature 1989, 341, 758–760. [Google Scholar] [CrossRef] [PubMed]
- Tocci, M.J.; Matkovich, D.A.; Collier, K.A.; Kwok, P.; Dumont, F.; Lin, S.; Degudicibus, S.; Siekierka, J.J.; Chin, J.; Hutchinson, N.I. The immunosuppressant FK506 selectively inhibits expression of early T cell activation genes. J. Immunol. 1989, 143, 718–726. [Google Scholar] [PubMed]
- Foor, F.; Parent, S.A.; Morin, N.; Dahl, A.M.; Ramadan, N.; Chrebet, G.; Bostian, K.A.; Nielsen, J.B. Calcineurin mediates inhibition by FK506 and cyclosporin of recovery from alpha-factor arrest in yeast. Nature 1992, 360, 682–684. [Google Scholar] [PubMed]
- Kunz, J.; Hall, M.N.; Cyclosporin, A. FK506 and rapamycin: More than just immunosuppression. Trends Biochem. Sci. 1993, 18, 334–338. [Google Scholar] [CrossRef]
- Breuder, T.; Hemenway, C.S.; Movva, N.R.; Cardenas, M.E.; Heitman, J. Calcineurin is essential in cyclosporin A- and FK506-sensitive yeast strains. Proc. Natl. Acad. Sci. USA 1994, 91, 5372–5376. [Google Scholar] [CrossRef] [PubMed]
- Trede, N.S.; Warwick, A.B.; Rosoff, P.M.; Rohrer, R.; Bierer, B.E.; Guinan, E. Tacrolimus (FK506) in allogeneic bone marrow transplantation for severe aplastic anemia following orthotopic liver transplantation. Bone Marrow Transplant. 1997, 20, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kriesche, H.-U.; Li, S.; Gruessner, R.W.G.; Fung, J.J.; Bustami, R.T.; Barr, M.L.; Leichtman, A.B. Immunosuppression: Evolution in practice and trends, 1994–2004. Am. J. Transplant. 2006, 6, 1111–1131. [Google Scholar] [CrossRef] [PubMed]
- McCormack, P.L.; Keating, G.M. Tacrolimus: In heart transplant recipients. Drugs 2006, 66, 2269–2279. [Google Scholar] [CrossRef] [PubMed]
- Ingram, J.R.; Martin, J.A.; Finlay, A.Y. Impact of topical calcineurin inhibitors on quality of life in patients with atopic dermatitis. Am. J. Clin. Dermatol. 2009, 10, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Remitz, A.; Reitamo, S. Long-term safety of tacrolimus ointment in atopic dermatitis. Expert Opin. Drug Saf. 2009, 8, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.; Jensen, M.R.; McAlpine, J. Toward the optimization of stent-based treatment for coronary artery disease. Curr. Opin. Drug Discov. Devel. 2010, 13, 157–158. [Google Scholar] [PubMed]
- Akimoto, K.; Kusunoki, Y.; Nishio, S.; Takagi, K.; Kawai, S. Safety profile of tacrolimus in patients with rheumatoid arthritis. Clin. Rheumatol. 2008, 27, 1393–1397. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.; Barrett, T.; Sparberg, M.; Buchman, A.L. Efficacy and safety of tacrolimus in refractory ulcerative colitis and Crohn’s disease: A single-center experience. Inflamm. Bowel Dis. 2008, 14, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Reis, S.A.; Moussatché, N.; Damaso, C.R.A. FK506, a secondary metabolite produced by Streptomyces, presents a novel antiviral activity against Orthopoxvirus infection in cell culture. J. Appl. Microbiol. 2006, 100, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Karpas, A.; Lowdell, M.; Jacobson, S.K.; Hill, F. Inhibition of human immunodeficiency virus and growth of infected T cells by the immunosuppressive drugs cyclosporin A and FK 506. Proc. Natl. Acad. Sci. USA 1992, 89, 8351–8355. [Google Scholar] [CrossRef] [PubMed]
- Briggs, C.J.; Ott, D.E.; Coren, L.V.; Oroszlan, S.; Tözsér, J. Comparison of the effect of FK506 and cyclosporin A on virus production in H9 cells chronically and newly infected by HIV-1. Arch. Virol. 1999, 144, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Mortola, E.; Endo, Y.; Ohno, K.; Watari, T.; Tsujimoto, H.; Hasegawa, A. The use of two immunosuppressive drugs, cyclosporin A and tacrolimus, to inhibit virus replication and apoptosis in cells infected with feline immunodeficiency virus. Vet. Res. Commun. 1998, 22, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Jiang, H.; Kato, R. Stimulation of hair growth by topical application of FK506, a potent immunosuppressive agent. J. Investig. Dermatol. 1994, 102, 160–164. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Klettner, A.; Herdegen, T. FK506 and its analogs—Therapeutic potential for neurological disorders. Curr. Drug Targets CNS Neurol. Disord. 2003, 2, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Sierra-Paredes, G.; Sierra-Marcuño, G. Ascomycin and FK506: Pharmacology and therapeutic potential as anticonvulsants and neuroprotectants. CNS Neurosci. Ther. 2008, 14, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Konofaos, P.; Terzis, J.K. FK506 and nerve regeneration: Past, present, and future. J. Reconstr. Microsurg. 2013, 29, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Periyasamy, S.; Warrier, M.; Tillekeratne, M.P.M.; Shou, W.; Sanchez, E.R. The immunophilin ligands cyclosporin A and FK506 suppress prostate cancer cell growth by androgen receptor-dependent and -independent mechanisms. Endocrinology 2007, 148, 4716–4726. [Google Scholar] [CrossRef] [PubMed]
- Barot, R.K.; Shitole, S.C.; Bhagat, N.; Patil, D.; Sawant, P.; Patil, K. Therapeutic effect of 0.1% Tacrolimus Eye Ointment in Allergic Ocular Diseases. JCDR 2016, 10, NC05-9. [Google Scholar] [CrossRef] [PubMed]
- Pirsch, J.D.; Miller, J.; Deierhoi, M.H.; Vincenti, F.; Filo, R.S. A comparison of tacrolimus (FK506) and cyclosporine for immunosuppression after cadaveric renal transplantation. FK506 Kidney Transplant Study Group. Transplantation 1997, 63, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Kobayashi, M. Differences between cyclosporin A and tacrolimus in organ transplantation. Transplant. Proc. 1999, 31, 1978–1980. [Google Scholar] [CrossRef]
- Motamedi, H.; Shafiee, A. The biosynthetic gene cluster for the macrolactone ring of the immunosuppressant FK506. Eur. J. Biochem. 1998, 256, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Motamedi, H.; Shafiee, A.; Cai, S.J.; Streicher, S.L.; Arison, B.H.; Miller, R.R. Characterization of methyltransferase and hydroxylase genes involved in the biosynthesis of the immunosuppressants FK506 and FK520. J. Bacteriol. 1996, 178, 5243–5248. [Google Scholar] [CrossRef] [PubMed]
- Motamedi, H.; Cai, S.J.; Shafiee, A.; Elliston, K.O. Structural organization of a multifunctional polyketide synthase involved in the biosynthesis of the macrolide immunosuppressant FK506. Eur. J. Biochem. 1997, 244, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Shafiee, A.; Motamedi, H.; Chen, T. Enzymology of FK-506 biosynthesis. Purification and characterization of 31-O-desmethylFK-506 O:methyltransferase from Streptomyces sp. MA6858. Eur. J. Biochem. 1994, 225, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, C.; Martínez-Castro, M. Trends in the biosynthesis and production of the immunosuppressant tacrolimus (FK506). Appl. Microbiol. Biotechnol. 2014, 98, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Zong, G.; Zhong, C.; Fu, J.; Qin, R.; Cao, G. Draft genome sequence of the tacrolimus-producing bacterium Streptomyces tsukubaensis F601. Genome Announc. 2017, 5, e00385-17. [Google Scholar] [CrossRef] [PubMed]
- Goranovič, D.A.; Blažič, I.M.; Magdevska, V.; Horvat, J.; Kuščer, E.; Polak, T.; Santos-Aberturas, J.; Martínez-Castro, M.; Barreiro, C.; Mrak, P.; et al. FK506 biosynthesis is regulated by two positive regulatory elements in Streptomyces tsukubaensis. BMC Microbiol. 2012, 12, 238. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Kim, D.H.; Lee, J.H.; Park, J.W.; Basnet, D.B.; Ban, Y.H.; Yoo, Y.J.; Chen, S.-W.; Park, S.R.; Choi, E.A.; et al. Biosynthesis of the allylmalonyl-CoA extender unit for the FK506 polyketide synthase proceeds through a dedicated polyketide synthase and facilitates the mutasynthesis of analogues. J. Am. Chem. Soc. 2011, 133, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez-Robles, M.; Rodríguez-García, A.; Martín, J.F. Target genes of the Streptomyces tsukubaensis FkbN regulator include most of the tacrolimus biosynthesis genes, a phosphopantetheinyl transferase and other PKS genes. Appl. Microbiol. Biotechnol. 2016, 100, 8091–8103. [Google Scholar] [CrossRef] [PubMed]
- Bauer, J.S.; Fillinger, S.; Förstner, K.; Herbig, A.; Jones, A.C.; Flinspach, K.; Sharma, C.; Gross, H.; Nieselt, K.; Apel, A.K. dRNA-seq transcriptional profiling of the FK506 biosynthetic gene cluster in Streptomyces tsukubaensis NRRL18488 and general analysis of the transcriptome. RNA Biol. 2017, 14, 1617–1626. [Google Scholar] [CrossRef] [PubMed]
- Andexer, J.N.; Kendrew, S.G.; Nur-e-Alam, M.; Lazos, O.; Foster, T.A.; Zimmermann, A.-S.; Warneck, T.D.; Suthar, D.; Coates, N.J.; Koehn, F.E.; et al. Biosynthesis of the immunosuppressants FK506, FK520, and rapamycin involves a previously undescribed family of enzymes acting on chorismate. Proc. Natl. Acad. Sci. USA 2011, 108, 4776–4781. [Google Scholar] [CrossRef] [PubMed]
- Goulet, M.T.; Rupprecht, K.M.; Sinclair, P.J.; Wyvratt, M.J.; Parsons, W.H. The medicinal chemistry of FK-506. Perspect. Drug Discov. Des. 1994, 2, 145–162. [Google Scholar] [CrossRef]
- Carroll, B.J.; Moss, S.J.; Bai, L.; Kato, Y.; Toelzer, S.; Yu, T.-W.; Floss, H.G. Identification of a set of genes involved in the formation of the substrate for the incorporation of the unusual “glycolate” chain extension unit in ansamitocin biosynthesis. J. Am. Chem. Soc. 2002, 124, 4176–4177. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Bai, L.; Xue, Q.; Revill, W.P.; Yu, T.-W.; Floss, H.G. Functional expression of genes involved in the biosynthesis of the novel polyketide chain extension unit, methoxymalonyl-acyl carrier protein, and engineered biosynthesis of 2-desmethyl-2-methoxy-6-deoxyerythronolide B. J. Am. Chem. Soc. 2002, 124, 5268–5269. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Chung, L.; Revill, W.P.; Katz, L.; Reeves, C.D. The FK520 gene cluster of Streptomyces hygroscopicus var. ascomyceticus (ATCC 14891) contains genes for biosynthesis of unusual polyketide extender units. Gene 2000, 251, 81–90. [Google Scholar] [PubMed]
- Chen, D.; Zhang, L.; Pang, B.; Chen, J.; Xu, Z.; Abe, I.; Liu, W. FK506 maturation involves a cytochrome p450 protein-catalyzed four-electron C-9 oxidation in parallel with a C-31 o-methylation. J. Bacteriol. 2013, 195, 1931–1939. [Google Scholar] [CrossRef] [PubMed]
- Goranovič, D.; Kosec, G.; Mrak, P.; Fujs, S.; Horvat, J.; Kuščer, E.; Kopitar, G.; Petković, H. Origin of the allyl group in FK506 biosynthesis. J. Biol. Chem. 2010, 285, 14292–14300. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wang, Y.Y.; Guo, Y.Y.; Shen, J.J.; Zhang, X.S.; Luo, H.D.; Ren, N.N.; Jiang, X.H.; Li, Y.Q. An acyltransferase domain of FK506 polyketide synthase recognizing both an acyl carrier protein and coenzyme A as acyl donors to transfer allylmalonyl and ethylmalonyl units. FEBS J. 2015, 282, 2527–2539. [Google Scholar] [CrossRef] [PubMed]
- Byrne, K.; Shafiee, A.; Nielsen, J.; Arison, B.; Monaghan, R.; Kaplan, L. The biosynthesis and enzymology of an immunosuppressant, immunomycin, produced by Streptomyces hygroscopicus var. ascomyceticus. Dev. Ind. Microbiol. 1993, 32, 29–45. [Google Scholar]
- Gatto, G.J., Jr.; McLoughlin, S.M.; Kelleher, N.L.; Walsh, C.T. Elucidating the substrate specificity and condensation domain activity of FkbP, the FK520 pipecolate-incorporating enzyme. Biochemistry 2005, 44, 5993–6002. [Google Scholar] [CrossRef] [PubMed]
- Gatto, G.J., Jr.; Boyne, M.T., 2nd; Kelleher, N.L.; Walsh, C.T. Biosynthesis of pipecolic acid by RapL, a lysine cyclodeaminase encoded in the rapamycin gene cluster. J. Am. Chem. Soc. 2006, 128, 3838–3847. [Google Scholar] [CrossRef] [PubMed]
- Van Duyne, G.D.; Standaert, R.F.; Karplus, P.A.; Schreiber, S.L.; Clardy, J. Atomic structure of FKBP-FK506, an immunophilin-immunosuppressant complex. Science 1991, 252, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.W.; Rotonda, J.; McKeever, B.M.; Chan, H.K.; Marcy, A.I.; Wiederrecht, G.; Hermes, J.D.; Springer, J.P. FK-506-binding protein: Three-dimensional structure of the complex with the antagonist L-685,818. J. Biol. Chem. 1993, 268, 11335–11339. [Google Scholar] [PubMed]
- Shafiee, A.; Motamedi, H.; Dumont, F.J.; Arison, B.H.; Miller, R.R. Chemical and biological characterization of two FK506 analogs produced by targeted gene disruption in Streptomyces sp. MA6548. J. Antibiot. 1997, 50, 418–423. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ban, Y.H.; Shinde, P.B.; Hwang, J.-Y.; Song, M.-C.; Kim, D.H.; Lim, S.-K.; Sohng, J.K.; Yoon, Y.J. Characterization of FK506 biosynthetic intermediates involved in post-PKS elaboration. J. Nat. Prod. 2013, 76, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Schrijver, A.D.; Mot, R.D. A subfamily of MalT-related ATP-dependent regulators in the LuxR family. Microbiology 1999, 145, 1287–1288. [Google Scholar] [CrossRef] [PubMed]
- Bibb, M.J. Regulation of secondary metabolism in Streptomycetes. Curr. Opin. Microbiol. 2005, 8, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Molnár, I.; Aparicio, J.F.; Haydock, S.F.; Khaw, L.E.; Schwecke, T.; König, A.; Staunton, J.; Leadlay, P.F. Organisation of the biosynthetic gene cluster for rapamycin in Streptomyces hygroscopicus: Analysis of genes flanking the polyketide synthase. Gene 1996, 169, 1–7. [Google Scholar] [CrossRef]
- Wilson, D.J.; Xue, Y.; Reynolds, K.A.; Sherman, D.H. Characterization and analysis of the PikD regulatory factor in the pikromycin biosynthetic pathway of Streptomyces venezuelae. J. Bacteriol. 2001, 183, 3468–3475. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Lei, J.; Liu, Y.; Wang, Y. The LuxR family members GdmRI and GdmRII are positive regulators of geldanamycin biosynthesis in Streptomyces hygroscopicus 17997. Arch. Microbiol. 2008, 189, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Huang, X.; Zhou, X.; Bai, L.; He, J.; Jeong, K.J.; Lee, S.Y.; Deng, Z. Organizational and Mutational Analysis of a Complete FR-008/Candicidin Gene Cluster Encoding a Structurally Related Polyene Complex. Chem. Biol. 2003, 10, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Aparicio, J.F. Biosynthesis Enzymology of the Polyenes Pimaricin and Candicidin. Methods Enzymol. 2009, 459, 215–242. [Google Scholar] [PubMed]
- Santos-Aberturas, J.; Payero, T.D.; Vicente, C.M.; Guerra, S.M.; Cañibano, C.; Martín, J.F.; Aparicio, J.F. Functional conservation of PAS-LuxR transcriptional regulators in polyene macrolide biosynthesis. Metab. Eng. 2011, 13, 756–757. [Google Scholar] [CrossRef] [PubMed]
- Santos-Aberturas, J.; Vicente, C.M.; Guerra, S.M.; Payero, T.D.; Martín, J.F.; Aparicio, J.F. Molecular control of polyene macrolide biosynthesis: Direct binding of the regulator PimM to eight promoters of pimaricin genes and identification of binding boxes. J. Biol. Chem. 2011, 286, 9150–9161. [Google Scholar] [CrossRef] [PubMed]
- Brautaset, T.; Sekurova, O.N.; Sletta, H.; Ellingsen, T.E.; Strøm, A.R.; Valla, S.; Zotchev, S.B. Biosynthesis of the polyene antifungal antibiotic nystatin in Streptomyces noursei ATCC 11455: Analysis of the gene cluster and deduction of the biosynthetic pathway. Chem. Biol. 2000, 7, 395–403. [Google Scholar] [CrossRef]
- Carmody, M.; Byrne, B.; Murphy, B.; Breen, C.; Lynch, S.; Flood, E.; Finnan, S.; Caffrey, P. Analysis andmanipulation of amphotericin biosynthetic genes by means of modified phage KC515 transduction techniques. Gene 2004, 343, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Ikeda, H.; Ishikawa, J.; Hanamoto, A.; Takahashi, C.; Shinose, M.; Hattori, M. Genome Sequence of an Industrial Microorganism Streptomyces avermitilis: Deducing the Ability of Producing Secondary Metabolites. Proc. Natl. Acad. Sci. USA 2001, 98, 12215–12220. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, H.; Ishikawa, J.; Hanamoto, A.; Shinose, M.; Kikuchi, H.; Shiba, T.; Sakaki, Y.; Hattori, M.; Omura, S. Complete genome sequence and comparative analysis of the industrial microorganism Streptomyces avermitilis. Nat. Biotechnol. 2003, 21, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Wei, L.; Liu, J.; Wang, J.; Qi, H.; Wen, J. Engineering of the LysR family transcriptional regulator FkbR1 and its target gene to improve ascomycin production. Appl. Microbiol. Biotechnol. 2017, 101, 4581–4592. [Google Scholar] [CrossRef] [PubMed]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.-M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3. Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.A. Molecular biology of the LysR family of transcriptional regulators. Annu. Rev. Microbiol. 1993, 47, 597–626. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, S.E.; Oyston, P.C. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154 (Pt 12), 3609–3623. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.; Nuñez, L.E.; Braña, A.F.; Mendez, C.; Salas, J.A.; Blanco, G. Identification of transcriptional activators for thienamycin and cephamycin C biosynthetic genes within the thienamycin gene cluster from Streptomyces cattleya. Mol. Microbiol. 2008, 69, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Redondo, R.; Rodríguez-García, A.; Martín, J.F.; Liras, P. The claR gene of Streptomyces clavuligerus, encoding a LysR-type regulatory protein controlling clavulanic acid biosynthesis, is linked to the clavulanate-9-aldehyde reductase (car) gene. Gene 1998, 211, 311–321. [Google Scholar] [CrossRef]
- Kölling, R.; Lother, H. AsnC: An autogenlously regulated activator of asparragine syntetase A transcription in Eschericha coli. J. Bacteriol. 1985, 164, 310–315. [Google Scholar] [PubMed]
- Zhang, X.S.; Luo, H.D.; Tao, Y.; Wang, Y.Y.; Jiang, X.H.; Jiang, H.; Li, Y.Q. FkbN and Tcs7 are pathway-specific regulators of the FK506 biosynthetic gene cluster in Streptomyces tsukubaensis L19’. J. Ind. Microbiol. Biotechnol. 2016, 43, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Yoo, Y.J.; Ban, Y.H.; Lee, S.-K.; Kim, E.; Suh, J.-W.; Yoon, Y.J. Roles of fkbN in positive regulation and tcs7 in negative regulation of FK506 biosynthesis in Streptomyces sp. strain KCTC 11604BP. Appl. Environ. Microbiol. 2012, 78, 2249–2255. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.C.; Gust, B.; Kulik, A.; Heide, L.; Buttner, M.J.; Bibb, M.J. Phage p1-derived artificial chromosomes facilitate heterologous expression of the FK506 gene cluster. PLoS ONE 2013, 8, e69319. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular regulation of antibiotic biosynthesis in Streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Burgo, Y.; Álvarez-Álvarez, R.; Rodríguez-García, A.; Liras, P. The Pathway-Specific Regulator ClaR of Streptomyces clavuligerus has a Global Effect on the Expression of Genes for Secondary Metabolism and Differentiation. Appl. Environ. Microbiol. 2015, 81, 6637–6648. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.W.; San Roman, A.K.; Gehring, A.M. Role of Phosphopantetheinyl Transferase Genes in Antibiotic Production by Streptomyces coelicolor. J. Bacteriol. 2008, 190, 6903–6908. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, Y.Y.; Zhang, X.S.; Luo, H.D.; Ren, N.N.; Jiang, X.H.; Jiang, H.; Li, Y.Q. Characterization of discrete phosphopantetheinyl transferases in Streptomyces tsukubaensis L19 unveils a complicate phosphopantetheinylation network. Sci. Rep. 2016, 6, 24255. [Google Scholar] [CrossRef] [PubMed]
- Blazic, M.; Starcevic, A.; Lisfi, M.; Baranasic, D.; Goranovic, D.; Fujs, S.; Kuščer, E.; Kosec, G.; Petkovic, H.; Cullum, J.; et al. Annotation of the modular polyketide synthase and nonribosomal peptide synthetase gene clusters in the genome of Streptomyces tsukubaensis NRRL18488. Appl. Environ. Microbiol. 2012, 78, 8183–8190. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.D. Information content of individual genetic sequences. J. Theor. Biol. 1997, 189, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Vockenhuber, M.P.; Sharma, C.M.; Statt, M.G.; Schmidt, D.; Xu, Z.; Dietrich, S.; Liesegang, H.; Mathews, D.H.; Suess, B. Deep sequencing based identification of small non-coding RNAs in Streptomyces coelicolor. RNA Biol. 2011, 8, 468–477. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hatanaka, H.; Kino, T.; Asano, M.; Goto, T.; Tanaka, H.; Okuhara, M. FK-506 related compounds produced by Streptomyces tsukubaensis No. 9993. J. Antibiot. 1989, 42, 620–622. [Google Scholar] [CrossRef] [PubMed]
- Kosec, G.; Goranovič, D.; Mrak, P.; Fujs, S.; Kuščer, E.; Horvat, J.; Kopitar, G.; Petković, H. Novel chemobiosynthetic approach for exclusive production of FK506. Metab. Eng. 2012, 14, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Mo, S.-J.; Park, S.R.; Ban, Y.-H.; Yoo, Y.J.; Yoon, Y.J. Liquid chromatography-mass spectrometry characterization of FK506 biosynthetic intermediates in Streptomyces clavuligerus KCTC 10561BP. Anal. Biochem. 2009, 393, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuka, M.; Ragan, J.A.; Sammakia, T.; Smith, D.B.; Uehling, D.E.; Schreiber, S.L. Total synthesis of FK506 and an FKBP probe reagent, [C,C-13C2]-FK506. J. Am. Chem. Soc. 1990, 112, 5583–5601. [Google Scholar] [CrossRef]
- Ireland, R.E.; Gleason, J.L.; Gegnas, L.D.; Highsmith, T.K. A Total Synthesis of FK-506. J. Org. Chem. 1996, 61, 6856–6872. [Google Scholar] [CrossRef] [PubMed]
- Ban, Y.H.; Park, S.R.; Yoon, Y.J. The biosynthetic pathway of FK506 and its engineering: From past achievements to future prospects. J. Ind. Microbiol. Biotechnol. 2016, 43, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.F.; Tao, Y.; Jin, M.Y.; Jiang, H. Improvement of FK506 production by synthetic biology approaches. Biotechnol. Lett. 2016, 38, 2015–2021. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.J.; Choi, C.Y. Nutrient Effects on FK-506, a New Immunosuppressant, Production by Streptomyces sp. in a Defined Medium. J. Ferment. Bioeng. 1997, 83, 599–603. [Google Scholar] [CrossRef]
- Martínez-Castro, M.; Salehi-Najafabadi, Z.; Romero, F.; Pérez-Sanchiz, R.; Fernández-Chimeno, R.I.; Martín, J.F.; Barreiro, C. Taxonomy and chemically semi-defined media for the analysis of the tacrolimus producer ‘Streptomyces tsukubaensis’. Appl. Microbiol. Biotechnol. 2013, 97, 2139–2152. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez-Robles, M.; Santos-Beneit, F.; Rodríguez-García, A.; Martín, J.F. Analysis of the PHO regulon in Streptomyces tsukubaensis. Microbiol. Res. 2017, 205, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez-Robles, M.; Santos-Beneit, F.; Albillos, S.M.; Liras, P.; Martín, J.F.; Rodríguez-García, A. Streptomyces tsukubaensis as a new model for carbon repression: Transcriptomic response to tacrolimus repressing carbon sources. Appl. Microbiol. Biotechnol. 2017, 101, 8181–8195. [Google Scholar] [CrossRef] [PubMed]
- Reeves, A.R.; Cernota, W.H.; Brikun, I.A.; Wesley, R.K.; Weber, J.M. Engineering precursor flow for increased erythromycin production in Aeromicrobium erythreum. Metab. Eng. 2004, 6, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.P.; Behera, B.K. Regulation of tacrolimus production by altering primary source of carbons and amino acids. Lett. Appl. Microbiol. 2009, 49, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Verma, S. Optimization of process parameters for tacrolimus (FK 506) production by new isolate of Streptomyces sp. using response surface methodology. J. Biochem. Technol. 2012, 3, 419–425. [Google Scholar]
- Xia, M.; Huang, D.; Li, S.; Wen, J.; Jia, X.; Chen, Y. Enhanced FK506 production in Streptomyces tsukubaensis by rational feeding strategies based on comparative metabolic profiling analysis. Biotechnol. Bioeng. 2013, 110, 2717–2730. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Xia, M.; Li, S.; Wen, J.; Jia, X. Enhancement of FK506 production by engineering secondary pathways of Streptomyces tsukubaensis and exogenous feeding strategies. J. Ind. Microbiol. Biotechnol. 2013, 40, 1023–1037. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Huang, D.; Xia, M.; Wen, J.; Huang, M. Improved FK506 production by the precursors and product-tolerant mutant of Streptomyces tsukubaensis based on genome shuffling and dynamic fed-batch strategies. J. Ind. Microbiol. Biotechnol. 2014, 41, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, J.; Liu, H.; Liang, S.; Wen, J. Combining metabolomics and network analysis to improve tacrolimus productionin Streptomyces tsukubaensis using different exogenous feedings. J. Ind. Microbiol. Biotechnol. 2017, 44, 1527–1540. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, H.; Huang, D.; Jin, L.; Wang, C.; Wen, J. Comparative proteomic and metabolomic analysis of Streptomyces tsukubaensis reveals the metabolic mechanism of FK506 overproduction by feeding soybean oil. Appl. Microbiol. Biotechnol. 2017, 101, 2447–2465. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Ban, Y.-H.; Park, J.W.; Yoo, Y.J.; Yoon, Y.J. Enhanced FK506 production in Streptomyces clavuligerus CKD1119 by engineering the supply of methylmalonyl-CoA precursor. J. Ind. Microbiol. Biotechnol. 2009, 36, 1473–1482. [Google Scholar] [CrossRef] [PubMed]
- Turlo, J.; Gajzlerska, W.; Klimaszewska, M.; Król, M.; Dawidowski, M.; Gutkowska, B. Enhancement of tacrolimus productivity in Streptomyces tsukubaensis by the use of novel precursors for biosynthesis. Enzyme Microb. Technol. 2012, 51, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Gajzlerska, W.; Kurkowiak, J.; Turlo, J. Use of three-carbon chain compounds as biosynthesis precursors to enhance tacrolimus production in Streptomyces tsukubaensis. New Biotechnol. 2014, 32, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhang, W.; Chen, X.; Wu, H.; Duan, Y.; Xu, Z. Generation of high rapamycin producing strain via rational metabolic pathway-based mutagenesis and further titer improvement with fed-batch bioprocess optimization. Biotechnol. Bioeng. 2010, 107, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhang, Q.; Zhang, Q.; Cen, P.; Xu, Z.; Liu, W. Improvement of FK506 production in Streptomyces tsukubaensis by genetic enhancement of the supply of unusual polyketide extender units via utilization of two distinct site-specific recombination systems. Appl. Environ. Microbiol. 2012, 78, 5093–5103. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wang, G.Y.; Li, X.; Waters, B.; Davies, J. Enhanced production of microbial metabolites in the presence of dimethyl sulfoxide. J. Antibiot. 2000, 53, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.R.; Cundliffe, E. Influence of dimethylsulfoxide on tylosin production in Streptomyces fradiae. J. Ind. Microbiol. Biotechnol. 2001, 27, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Li, S.; Xia, M.; Wen, J.; Jia, X. Genome-scale metabolic network guided engineering of Streptomyces tsukubaensis for FK506 production improvement. Microb. Cell. Fact. 2013, 12, 52. [Google Scholar] [CrossRef] [PubMed]
- Pei, G.; Chen, L.; Zhang, W. Chapter nine-WGCNA application to proteomic and metabolomic data analysis. Methods Enzymol. 2017, 585, 135–158. [Google Scholar] [PubMed]
- Wang, C.; Liu, J.; Liu, H.; Wang, J.; Wen, J. A genome-scale dynamic flux balance analysis model of Streptomyces tsukubaensis NRRL18488 to predict the targets for increasing FK506 production. Biochem. Eng. J. 2017, 123, 45–56. [Google Scholar] [CrossRef]
- Höffner, K.; Harwood, S.; Barton, P. A reliable simulator for dynamic flux balance analysis. Biotechnol. Bioeng 2013, 110, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Sola-Landa, A.; Moura, R.S.; Martín, J.F. The two-component PhoR-PhoP system controls both primary metabolism and secondary metabolite biosynthesis in Streptomyces lividans. Proc. Natl. Acad. Sci. USA 2003, 100, 6133–6138. [Google Scholar] [CrossRef] [PubMed]
- Sola-Landa, A.; Rodríguez-García, A.; Franco-Domínguez, E.; Martín, J.F. Binding of PhoP to promoters of phosphate-regulated genes in Streptomyces coelicolor: Identification of PHO boxes. Mol. Microbiol. 2005, 56, 1373–1385. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F. Phosphate control of the biosynthesis of antibiotics and other secondary metabolites is mediated by the PhoR-PhoP system: An unfinished story. J. Bacteriol. 2004, 186, 5197–5201. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Santos-Beneit, F.; Rodríguez-García, A.; Sola-Landa, A.; Smith, M.C.M.; Ellingsen, T.E.; Nieselt, K.; Burroughs, N.J.; Wellington, E.M.H. Transcriptomic studies of phosphate control of primary and secondary metabolism in Streptomyces coelicolor. Appl. Microbiol. Biotechnol. 2012, 95, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Santos-Beneit, F. The Pho regulon: A huge regulatory network in bacteria. Front. Microbiol. 2015, 6, 402. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Rodríguez-García, A.; Liras, P. The master regulator PhoP coordinates phosphate and nitrogen metabolism, respiration, cell differentiation and antibiotic biosynthesis: Comparison in Streptomyces coelicolor and Streptomyces avermitilis. J. Antibiot. 2017, 70, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Makino, K.; Shinagawa, H.; Amemura, M.; Kimura, S.; Nakata, A.; Ishihama, A. Regulation of the phosphate regulon of Escherichia coli Activation of pstS transcription by PhoB protein in vitro. J. Mol. Biol. 1988, 203, 85–95. [Google Scholar] [CrossRef]
- Ruiz, B.; Chávez, A.; Forero, A.; García-Huante, Y.; Romero, A.; Sánchez, M.; Rocha, D.; Sánchez, B.; Rodríguez-Sanoja, R.; Sánchez, S.; et al. Production of microbial secondary metabolites: Regulation by the carbon source. Crit. Rev. Microbiol. 2010, 36, 146–167. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, S.; Chávez, A.; Forero, A.; García-Huante, Y.; Romero, A.; Sánchez, M.; Rocha, D.; Sánchez, B.; Avalos, M.; Guzmán-Trampe, S.; et al. Carbon source regulation of antibiotic production. J. Antibiot. 2010, 63, 442–459. [Google Scholar] [CrossRef] [PubMed]
- Strakova, E.; Zikova, A.; Vohradsky, J. Inference of sigma factor controlled networks by using numerical modeling applied to microarray time series data of the germinating prokaryote. Nucl. Acids Res. 2014, 42, 748–763. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, S.; Watanabe, M.; Saito, N.; Hesketh, A.; Vachalova, K.; Matsubara, K.; Ochi, K. Molecular and functional analyses of the gene (eshA) encoding the 52-kilodalton protein of Streptomyces coelicolor A3 required for antibiotic production. J. Bacteriol. 2001, 183, 6009–6016. [Google Scholar] [CrossRef] [PubMed]
- Saito, N.; Xu, J.; Hosaka, T.; Okamoto, S.; Aoki, H.; Bibb, M.J.; Ochi, K. EshA accentuates ppGpp accumulation and is conditionally required for antibiotic production in Streptomyces coelicolor A3. J. Bacteriol. 2006, 188, 4952–4961. [Google Scholar] [CrossRef] [PubMed]
- Uguru, G.C.; Stephens, K.E.; Stead, J.A.; Towle, J.E.; Baumberg, S.; McDowall, K.J. Transcriptional activation of the pathway-specific regulator of the actinorhodin biosynthetic genes in Streptomyces coelicolor. Mol. Microbiol. 2005, 58, 131–150. [Google Scholar] [CrossRef] [PubMed]
- Vujaklija, D.; Horinouchi, S.; Beppu, T. Detection of an A-factor-responsive protein that binds to the upstream activation sequence of strR, a regulatory gene for streptomycin biosynthesis in Streptomyces griseus. J. Bacteriol. 1993, 175, 2652–2661. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rigali, S.; Nothaft, H.; Noens, E.E.E.; Schlicht, M.; Colson, S.; Müller, M.; Joris, B.; Koerten, H.K.; Hopwood, D.A.; Titgemeyer, F.; et al. The sugar phosphotransferase system of Streptomyces coelicolor is regulated by the GntR-family regulator DasR and links N-acetylglucosamine metabolism to the control of development. Mol. Microbiol. 2006, 61, 1237–1251. [Google Scholar] [CrossRef] [PubMed]
- Rigali, S.; Titgemeyer, F.; Barends, S.; Mulder, S.; Thomae, A.W.; Hopwood, D.A.; van Wezel, G.P. Feast or famine: The global regulator DasR links nutrient stress to antibiotic production by Streptomyces. EMBO Rep. 2008, 9, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, C.; Prieto, C.; Sola-Landa, A.; Solera, E.; Martínez-Castro, M.; Pérez-Redondo, R.; García-Estrada, C.; Aparicio, J.F.; Fernández-Martínez, L.T.; Santos-Aberturas, J.; et al. Draft genome of Streptomyces tsukubaensis NRRL 18488, the producer of the clinically important immunosuppressant tacrolimus (FK506). J. Bacteriol. 2012, 194, 3756–3757. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Liras, P. Novel Antimicrobial and other Bioactive Metabolites obtained from Silent Gene Clusters. In Antibiotics: Current Innovations and Future Trends; Demain, A.L., Sánchez, S., Eds.; Horizon Scientific Press and Caister Academic Press: Norkfolk, UK, 2015; pp. 275–292. ISBN 978-1-908230-54-6. [Google Scholar]
- Werner, G.; Hagenmaier, H.; Drautz, H.; Baumgartner, A.; Zähner, H. Metabolic products of microorganisms. 224. Bafilomycins, a new group of macrolide antibiotics. Production, isolation, chemical structure and biological activity. J. Antibiot. 1984, 37, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.M.; Mironenko, T.; Sun, Y.; Hong, H.; Deng, Z.; Leadlay, P.F.; Weissman, K.J.; Haydock, S.F. Insights into polyether biosynthesis from analysis of the nigericin biosynthetic gene cluster in Streptomyces sp. DSM4137. Chem. Biol. 2007, 14, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Zabriskie, T.M. The enduracidin biosynthetic gene cluster from Streptomyces fungicidicus. Microbiology 2006, 152 Pt 10, 2969–2983. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Precursor | Strain | Reference |
|---|---|---|
| Soybean oil | Streptomyces sp. MA6858 B3178 | [97,98,99,100,101] |
| S. tsukubaensis TJ-04 | ||
| S. tsukubaensis D852 | ||
| l-lysine | Streptomyces sp. MA6858 B3178 | [93,97,98,100,101,102,103] |
| S. tsukubaensis D852 | ||
| S. tsukubaensis NRRL18488 | ||
| Methyl-oleate | S. clavuligerus CKD1119 | [98,104] |
| Pipecolic acid | S. tsukubaensis NRRL18488 | [100,101,105] |
| S. tsukubaensis D852 | ||
| Picolinic acid | S. tsukubaensis NRRL18488 | [105] |
| Nicotinamide | S. tsukubaensis NRRL18488 | [105] |
| Nicotinic acid | S. tsukubaensis NRRL18488 | [105] |
| Chorismate | S. tsukubaensis D852 | [100,101] |
| Shikimate | S. tsukubaensis D852 | [99,100,101,102,103] |
| S. tsukubaensis TJ-04 | ||
| S. tsukubaensis NRRL18488 | ||
| Lactate | S. tsukubaensis D852 | [92,100,101,102] |
| Streptomyces sp. MA6858 | ||
| Succinate | S. tsukubaensis D852 | [99,100,101,103] |
| S. tsukubaensis TJ-04 | ||
| S. tsukubaensis NRRL18488 | ||
| Isoleucine | S. tsukubaensis D852 | [100,101] |
| Valine | S. tsukubaensis D852 | [99,100,101] |
| S. tsukubaensis TJ-04 | ||
| Proline | S. tsukubaensis TJ-04 | [99] |
| Leucine | S. tsukubaensis TJ-04 | [99] |
| Threonine | S. tsukubaensis TJ-04 | [99] |
| Propilenglycol | S. tsukubaensis FERM BP-927 | [106] |
| Propanol | S. tsukubaensis FERM BP-927 | [106] |
| Propionic acid | S. tsukubaensis FERM BP-927 | [106] |
| Malonate | S. tsukubaensis D852 | [102,103] |
| S. tsukubaensis NRRL18488 | ||
| Citrate | S. tsukubaensis D852 | [102,103] |
| S. tsukubaensis NRRL18488 |
| Gene/Modification | Strain | Reference |
|---|---|---|
| fkbO/overexpression | S. tsukubaensis D852 | [100] |
| fkbL/overexpression | S. tsukubaensis D852 | [100] |
| fkbM/overexpression | S. tsukubaensis D852 | [100] |
| fkbP/overexpression | S. tsukubaensis D852 | [100] |
| fkbD/overexpression | S. tsukubaensis D852 | [100] |
| gdhA/inactivation | S. tsukubaensis D852 | [111] |
| ppc/inactivation | S. tsukubaensis D852 | [111] |
| dahp/overexpression | S. tsukubaensis D852 | [111] |
| pntAB/overexpression | S. tsukubaensis D852 | [111] |
| accA2/overexpression | S. tsukubaensis D852 | [111] |
| zwf2/overexpression | S. tsukubaensis D852 | [111] |
| fkbD/overexpression | S. tsukubaensis D852 | [111] |
| aroC/overexpression | S. tsukubaensis D852 | [102] |
| dapA/overexpression | S. tsukubaensis D852 | [102] |
| gcdh/inactivation | S. tsukubaensis NRRL 18488 | [113] |
| tktB/overexpression | S. tsukubaensis NRRL 18488 | [113] |
| msdh/overexpression | S. tsukubaensis NRRL 18488 | [113] |
| ask/overexpression | S. tsukubaensis NRRL 18488 | [113] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ordóñez-Robles, M.; Santos-Beneit, F.; Martín, J.F. Unraveling Nutritional Regulation of Tacrolimus Biosynthesis in Streptomyces tsukubaensis through omic Approaches. Antibiotics 2018, 7, 39. https://doi.org/10.3390/antibiotics7020039
Ordóñez-Robles M, Santos-Beneit F, Martín JF. Unraveling Nutritional Regulation of Tacrolimus Biosynthesis in Streptomyces tsukubaensis through omic Approaches. Antibiotics. 2018; 7(2):39. https://doi.org/10.3390/antibiotics7020039
Chicago/Turabian StyleOrdóñez-Robles, María, Fernando Santos-Beneit, and Juan F. Martín. 2018. "Unraveling Nutritional Regulation of Tacrolimus Biosynthesis in Streptomyces tsukubaensis through omic Approaches" Antibiotics 7, no. 2: 39. https://doi.org/10.3390/antibiotics7020039
APA StyleOrdóñez-Robles, M., Santos-Beneit, F., & Martín, J. F. (2018). Unraveling Nutritional Regulation of Tacrolimus Biosynthesis in Streptomyces tsukubaensis through omic Approaches. Antibiotics, 7(2), 39. https://doi.org/10.3390/antibiotics7020039