

In Vitro Inhibitory Activity of Corilagin and Punicalagin Against Toxoplasma gondii and Their Mechanism(s) of Action
 ,
,
Abstract
1. Introduction
2. Results
2.1. In Vitro Inhibition of T. gondii Growth and Cytotoxicity
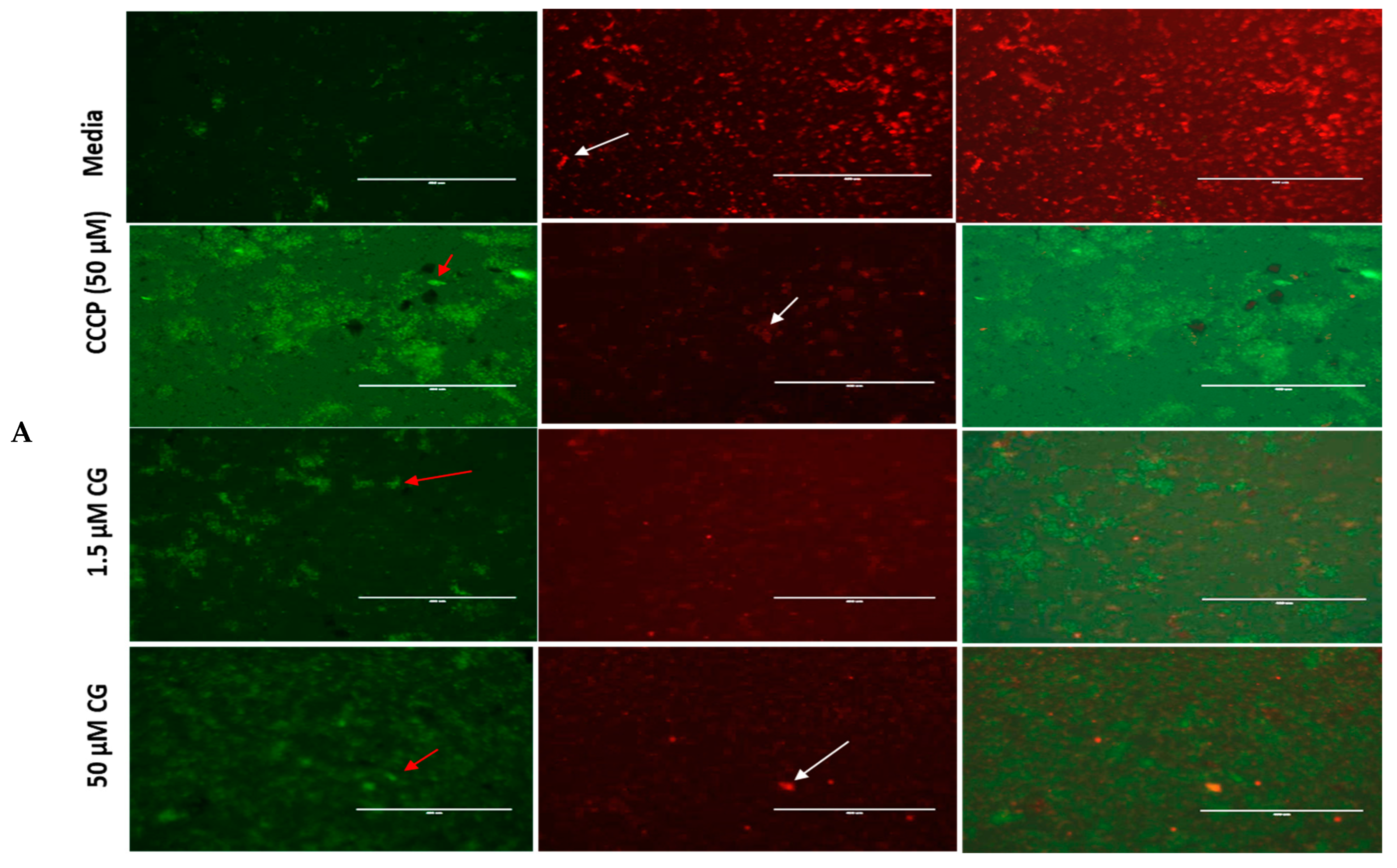
2.2. Effect of CG and PU on Mitochondria Superoxide (MitoSOX) and Reactive Oxygen Species (ROS) Production in T. gondii Tachyzoites
2.3. Mitochondrial Membrane Potential Disruption
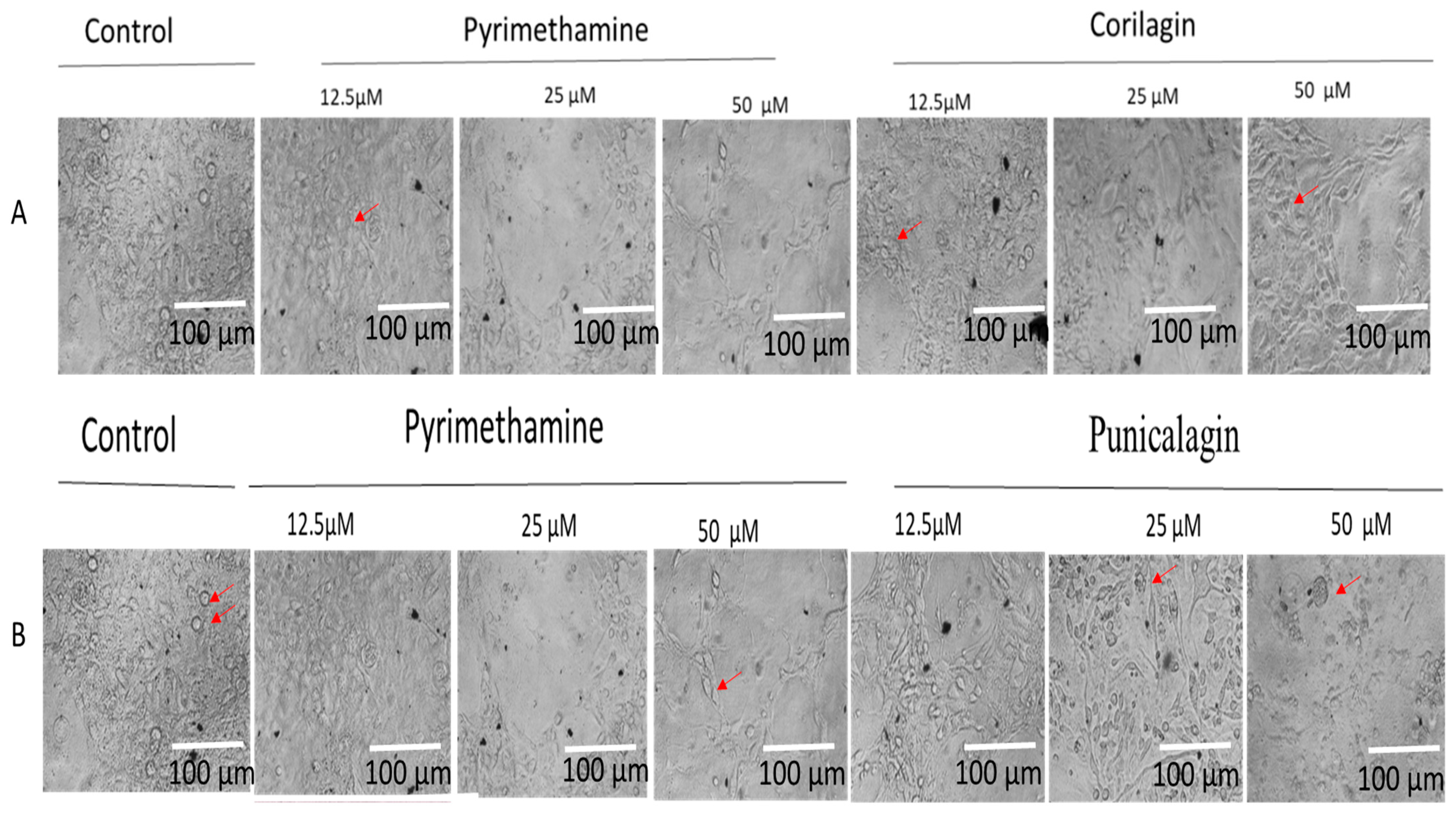
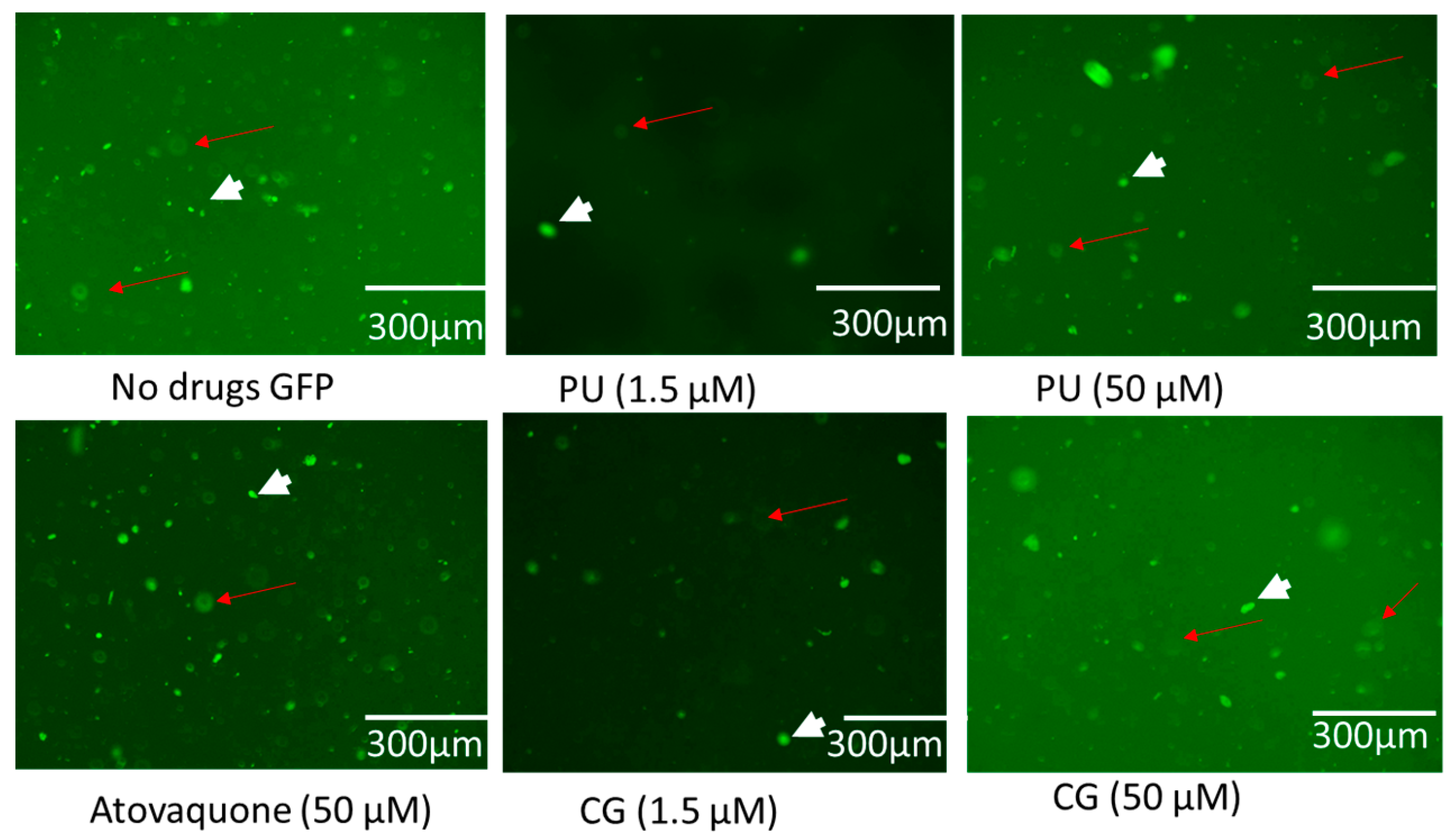
2.4. Integrity of Host Cells with Parasites and Drugs Interaction During Growth Inhibition Assay
3. Discussion
4. Materials and Methods
4.1. Chemicals Acquisition
4.2. Maintenance of Parasites and Cell Cultures
4.3. Cytotoxicity
4.4. Growth Inhibition Assays
4.5. Reactive Oxidative Species (ROS) Production
4.6. Mitochondrial Superoxide Production Assay
4.7. CG and PU Effect on T. gondii Mitochondrial Membrane Potential
4.8. Effect of PU, CG and AT on Intracellar T. gondii Tachyzoites and HFF Host Cells
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CDC. Parasites-Toxoplasmosis (Toxoplasma infection): Epidemiology and Risk; CDC: Centers for Disease Control and Prevention: Atlanta, GA, USA, 2023. [Google Scholar]
- Shiojiri, D.; Kinai, E.; Teruya, K.; Kikuchi, Y.; Oka, S. Combination of clindamycin and azithromycin as an alternative treatment for Toxoplasma gondii encephalitis. Emerg. Infect. Dis. 2019, 25, 841. [Google Scholar]
- Shammaa, A.M.; Powell, T.G.; Benmerzouga, I. Adverse outcomes associated with the treatment of Toxoplasma infections. Sci. Rep. 2021, 11, 1035. [Google Scholar] [CrossRef] [PubMed]
- Secrieru, A.; Costa, I.C.; O’Neill, P.M.; Cristiano, M.L. Antimalarial Agents as Therapeutic Tools against Toxoplasmosis—A Short Bridge between Two Distant Illnesses. Molecules 2020, 25, 1574. [Google Scholar] [CrossRef]
- Guaraldo, L.; Villar, B.B.D.L.F.; Durão, N.M.G.; Louro, V.C.; Quintana, M.D.S.B.; Curi, A.L.L.; Neves, E.S. Ocular toxoplasmosis: Adverse reactions to treatment in a Brazilian cohort. Trans. R. Soc. Trop. Med. Hyg. 2018, 112, 188–192. [Google Scholar] [PubMed]
- Bosch-Driessen, L.H.; Verbraak, F.D.; Suttorp-Schulten, M.S.; van Ruyven, R.L.; Klok, A.M.; Hoyng, C.B.; Rothova, A. A prospective, randomized trial of pyrimethamine and azithromycin vs. pyrimethamine and sulfadiazine for the treatment of ocular toxoplasmosis. Am. J. Ophthalmol. 2002, 134, 34–40. [Google Scholar] [PubMed]
- Angel, S.O.; Vanagas, L.; Ruiz, D.M.; Cristaldi, C.; Saldarriaga Cartagena, A.M.; Sullivan, W.J., Jr. Emerging Therapeutic Targets against Toxoplasma gondii: Update on DNA Repair Response Inhibitors and Genotoxic Drugs. Front. Cell. Infect. Microbiol. 2020, 10, 289. [Google Scholar]
- Mkhize, S.S.; Machaba, K.E.; Simelane, M.B.; Pooe, O.J. Mushroom-Derived Products as an Alternative Antimalarial Therapeutics: A Review. In Drug Development for Malaria: Novel Approaches for Prevention and Treatment; Wiley Online Library: Hoboken, NJ, USA, 2022; pp. 235–249. [Google Scholar]
- Leliebre-Lara, V.; Fidalgo, L.M.; Pferschy-Wenzig, E.-M.; Kunert, O.; Lima, C.N.; Bauer, R. In vitro antileishmanial activity of sterols from Trametes versicolor (Bres. Rivarden). Molecules 2016, 21, 1045. [Google Scholar] [CrossRef]
- Leliebre-Lara, V.; García, M.; Nogueiras, C.; Monzote, L. Qualitative analysis of an ethanolic extract from Trametes versicolor and biological screening against Leishmania amazonensis. Emir. J. Food Agric. 2015, 27, 592–595. [Google Scholar]
- Abugri, D.A.; Witola, W.H.; Jaynes, J.M.; Toufic, N. In vitro activity of Sorghum bicolor extracts, 3-deoxyanthocyanidins, against Toxoplasma gondii. Exp. Parasitol. 2016, 164, 12–19. [Google Scholar]
- Abugri, D.A.; Witola, W.H.; Jaynes, J.M. In vitro antagonistic and indifferent activity of combination of 3-deoxyanthocyanidins against Toxoplasma gondii. Parasitol. Res. 2017, 116, 3387–3400. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, P.; Reyes-Palomo, C.; Sanz-Fernández, S.; Rufino-Moya, P.J.; Zafra, R.; Martínez-Moreno, F.J.; Rodríguez-Estévez, V.; Díaz-Gaona, C. Antiparasitic tannin-rich plants from the south of Europe for grazing livestock: A review. Animals 2023, 13, 201. [Google Scholar] [CrossRef] [PubMed]
- Quijada, J.; Fryganas, C.; Ropiak, H.M.; Ramsay, A.; Mueller-Harvey, I.; Hoste, H. Anthelmintic activities against Haemonchus contortus or Trichostrongylus colubriformis from small ruminants are influenced by structural features of condensed tannins. J. Agric. Food Chem. 2015, 63, 6346–6354. [Google Scholar]
- Yan, F.; Cheng, D.; Wang, H.; Gao, M.; Zhang, J.; Cheng, H.; Wang, C.; Zhang, H.; Xiong, H. Corilagin ameliorates con A-induced hepatic injury by restricting M1 macrophage polarization. Front. Immunol. 2022, 12, 807509. [Google Scholar]
- Li, X.; Deng, Y.; Zheng, Z.; Huang, W.; Chen, L.; Tong, Q.; Ming, Y. Corilagin, a promising medicinal herbal agent. Biomed. Pharmacother. 2018, 99, 43–50. [Google Scholar] [PubMed]
- Wu, C.; Huang, H.; Choi, H.-Y.; Ma, Y.; Zhou, T.; Peng, Y.; Pang, K.; Shu, G.; Yang, X. Anti-esophageal cancer effect of corilagin extracted from phmllanthi fructus via the mitochondrial and endoplasmic reticulum stress pathways. J. Ethnopharmacol. 2021, 269, 113700. [Google Scholar] [PubMed]
- Deng, Y.; Li, X.; Zheng, Z.; Huang, W.; Chen, L.; Tong, Q.; Ming, Y. Corilagin induces the apoptosis of hepatocellular carcinoma cells through the mitochondrial apoptotic and death receptor pathways. Oncol. Rep. 2018, 39, 2545–2552. [Google Scholar]
- Cheng, J.T.; Lin, T.C.; Hsu, F.L. Antihypertensive effect of corilagin in the rat. Can. J. Physiol. Pharmacol. 1995, 73, 1425–1429. [Google Scholar]
- Li, N.; Luo, M.; Fu, Y.; Zu, Y.; Wang, W.; Zhang, L.; Yao, L.; Zhao, C.; Sun, Y. Effect of corilagin on membrane permeability of Escherichia coli, Staphylococcus aureus and Candida albicans. Phytother. Res. 2013, 27, 1517–1523. [Google Scholar]
- Yang, L.J.; Chen, R.H.; Hamdoun, S.; Coghi, P.; Ng, J.P.; Zhang, D.W.; Guo, X.; Xia, C.; Law, B.Y.K.; Wong, V.K.W. Corilagin prevents SARS-CoV-2 infection by targeting RBD-ACE2 binding. Phytomedicine 2021, 87, 153591. [Google Scholar]
- Xie, J.; Shang, L.; Liu, C.; Mao, J.; He, C.; Luo, M.; Fisher, D.; Hien, N.T.T.; Xu, S.; Zhao, L. Corilagin inhibits human cytomegalovirus infection and replication via activating the cGAS-STING signaling pathway in vitro and in vivo. Int. Immunopharmacol. 2024, 143, 113401. [Google Scholar]
- Xu, J.; Zhang, G.; Tong, Y.; Yuan, J.; Li, Y.; Song, G. Corilagin induces apoptosis, autophagy and ROS generation in gastric cancer cells in vitro. Int. J. Mol. Med. 2019, 43, 967–979. [Google Scholar]
- Lin, L.-T.; Chen, T.-Y.; Lin, S.-C.; Chung, C.-Y.; Lin, T.-C.; Wang, G.-H.; Anderson, R.; Lin, C.-C.; Richardson, C.D. Broad-spectrum antiviral activity of chebulagic acid and punicalagin against viruses that use glycosaminoglycans for entry. BMC Microbiol. 2013, 13, 187. [Google Scholar]
- Tito, A.; Colantuono, A.; Pirone, L.; Pedone, E.; Intartaglia, D.; Giamundo, G.; Conte, I.; Vitaglione, P.; Apone, F. A pomegranate peel extract as inhibitor of SARS-CoV-2 Spike binding to human ACE2 (in vitro): A promising source of novel antiviral drugs. Front. Chem. 2021, 9, 638187. [Google Scholar]
- Kang, B.; Kim, C.Y.; Hwang, J.; Jo, K.; Kim, S.; Suh, H.J.; Choi, H.S. Punicalagin, a pomegranate-derived ellagitannin, suppresses obesity and obesity-induced inflammatory responses via the Nrf2/Keap1 signaling pathway. Mol. Nutr. Food Res. 2019, 63, 1900574. [Google Scholar]
- Venusova, E.; Kolesarova, A.; Horky, P.; Slama, P. Physiological and immune functions of punicalagin. Nutrients 2021, 13, 2150. [Google Scholar] [CrossRef]
- Lu, L.; Peng, Y.; Yao, H.; Wang, Y.; Li, J.; Yang, Y.; Lin, Z. Punicalagin as an allosteric NSP13 helicase inhibitor potently suppresses SARS-CoV-2 replication in vitro. Antivir. Res. 2022, 206, 105389. [Google Scholar] [PubMed]
- Moon, M.J.; Kamasah, J.S.; Sharma, H.N.; Robertson, B.K.; Abugri, D.A. Apigeninidin chloride disrupts Toxoplasma gondii Mitochondrial membrane potential and induce reactive oxygen species and metabolites production. Front. Cell. Infect. Microbiol. 2024, 14, 1368019. [Google Scholar]
- Huffman, A.M.; Ayariga, J.A.; Napier, A.; Robertson, B.K.; Abugri, D.A. Inhibition of Toxoplasma gondii growth by Dihydroquinine and its Mechanisms of Action. Front. Cell. Infect. Microbiol. 2022, 12, 852889. [Google Scholar] [CrossRef]
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 3rd ed.; CRC Press: Boca Raton, Florida, USA, 2021. [Google Scholar]
- Flegr, J.; Prandota, J.; Sovičková, M.; Israili, Z.H. Toxoplasmosis–a global threat. Correlation of latent toxoplasmosis with specific disease burden in a set of 88 countries. PLoS ONE 2014, 9, e90203. [Google Scholar]
- Sun, P.R.; Gao, F.F.; Choi, H.G.; Zhou, W.; Yuk, J.M.; Kwon, J.; Lee, Y.H.; Cha, G.H. Dipenyleneiodonium induces growth inhibition of Toxoplasma gondii through ROS induction in ARPE-19 cells. Korean J. Parasitol. 2019, 57, 83. [Google Scholar]
- Zhang, J.; Si, H.; Li, B.; Zhou, X.; Zhang, J. Myrislignan exhibits activities against Toxoplasma gondii RH strain by triggering mitochondrial dysfunction. Front. Microbiol. 2019, 10, 2152. [Google Scholar] [CrossRef] [PubMed]
- Spalenka, J.; Escotte-Binet, S.; Bakiri, A.; Hubert, J.; Renault, J.H.; Velard, F.; Duchateau, S.; Aubert, D.; Huguenin, A.; Villena, I. Discovery of new inhibitors of Toxoplasma gondii via the pathogen box. Antimicrob. Agents Chemother. 2018, 62, e01640-17. [Google Scholar] [CrossRef]
- Qiu, Y.; Wang, W.; Wang, Q.; Lin, H.; Bai, Y.; Zhang, J. Effect of the flavonoid compound glabridin on tachyzoites and bradyzoites of Toxoplasma gondii. Parasites Vectors 2025, 18, 56. [Google Scholar] [CrossRef]
- Han, R.-X.; Jiang, P.-C.; Han, B.; Zhou, H.-Y.; Wang, Y.-L.; Guan, J.-Y.; Liu, Z.-R.; He, S.-Y.; Zhou, C.-X. Anti-Toxoplasma gondii effect of tylosin in vitro and in vivo. Parasites Vectors 2024, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Montazeri, M.; Sharif, M.; Sarvi, S.; Mehrzadi, S.; Ahmadpour, E.; Daryani, A. A systematic review of in vitro and in vivo activities of anti-Toxoplasma drugs and compounds (2006–2016). Front. Microbiol. 2017, 8, 228484. [Google Scholar] [CrossRef]
- Qiu, Y.; Wang, W.; Wang, Q.; Xu, J.; Dai, G.; Bai, Y.; Zhang, J. Activity Evaluation and Mode of Action of ICA Against Toxoplasma gondii In Vitro. Biomolecules 2025, 15, 202. [Google Scholar] [CrossRef]
- SaSanford, A.; Schulze, T.; Potluri, L.; Watson, G.; Darner, E.; Zach, S.; Hemsley, R.; Wallick, A.; Warner, R.; Charman, S.; et al. Derivatives of a benzoquinone acyl hydrazone with activity against Toxoplasma gondii. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 488–492. [Google Scholar] [CrossRef]
- Fonseca-Silva, F.; Inacio, J.D.; Canto-Cavalheiro, M.M.; Almeida-Amaral, E.E. Reactive oxygen species production and mitochondrial dysfunction contribute to quercetin induced death in Leishmania amazonensis. PLoS ONE 2011, 6, e14666. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Roy, A.; Dutta, N.; Majumder, H.K. Reactive oxygen species and imbalance of calcium homeostasis contributes to curcumin induced programmed cell death in Leishmania donovani. Apoptosis 2008, 13, 867–882. [Google Scholar]
- Zhang, J.; Si, H.; Lv, K.; Qiu, Y.; Sun, J.; Bai, Y.; Li, B.; Zhou, X.; Zhang, J. Licarin-B exhibits activity against the Toxoplasma gondii RH strain by damaging mitochondria and activating autophagy. Front. Cell Dev. Biol. 2021, 9, 684393. [Google Scholar] [CrossRef]
- Shi, Y.; Jiang, Y.; Qiu, H.; Hu, D.; Song, X. Mitochondrial dysfunction induced by bedaquiline as an anti-Toxoplasma alternative. Vet. Res. 2023, 54, 123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.L.; Si, H.F.; Shang, X.F.; Zhang, X.K.; Li, B.; Zhou, X.Z.; Zhang, J.Y. New Life for an Old Drug: In Vitro and In Vivo Effects of the Anthelmintic Drug Niclosamide Against Toxoplasma gondii RH Strain. Int. J. Parasitol. Drugs Drug Resist. 2019, 9, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qin, X.; Ma, W.; Jia, S.; Zhang, X.; Yang, X.; Pan, D.; Jin, F. Corilagin induces apoptosis and autophagy in NRF2-addicted U251 glioma cell line. Mol. Med. Rep. 2021, 23, 320. [Google Scholar] [CrossRef] [PubMed]
- MacRae, J.I.; Sheiner, L.; Nahid, A.; Tonkin, C.; Striepen, B.; McConville, M.J. Mitochondrial metabolism of glucose and glutamine is required for intracellular growth of Toxoplasma gondii. Cell Host Microbe 2012, 12, 682–692. [Google Scholar]
- Kakiuchi, N.; Hattori, M.; Namba, T.; Nishizawa, M.; Yamagishi, T.; Okuda, T. Inhibitory effect of tannins on reverse transcriptase from RNA tumor virus. J. Nat. Prod. 1985, 48, 614–621. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds (µM) | EC50s with 95% CL | CC50s | SI Value |
|---|---|---|---|
| PY | 0.25 (0.12 to 0.43) | Nd | Nd |
| CG | 3.09 (1.86 to 5.06) | 37.27 | 12.20 |
| PU | 19.33 (13.13 to 28.75) | 21.30 | 1.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Green-Ross, N.T.; Sharma, H.N.; Napier, A.; Robertson, B.K.; Green, R.L.; Abugri, D.A. In Vitro Inhibitory Activity of Corilagin and Punicalagin Against Toxoplasma gondii and Their Mechanism(s) of Action. Antibiotics 2025, 14, 336. https://doi.org/10.3390/antibiotics14040336
Green-Ross NT, Sharma HN, Napier A, Robertson BK, Green RL, Abugri DA. In Vitro Inhibitory Activity of Corilagin and Punicalagin Against Toxoplasma gondii and Their Mechanism(s) of Action. Antibiotics. 2025; 14(4):336. https://doi.org/10.3390/antibiotics14040336
Chicago/Turabian StyleGreen-Ross, Nicole T., Homa Nath Sharma, Audrey Napier, Boakai K. Robertson, Robert L. Green, and Daniel A. Abugri. 2025. "In Vitro Inhibitory Activity of Corilagin and Punicalagin Against Toxoplasma gondii and Their Mechanism(s) of Action" Antibiotics 14, no. 4: 336. https://doi.org/10.3390/antibiotics14040336
APA StyleGreen-Ross, N. T., Sharma, H. N., Napier, A., Robertson, B. K., Green, R. L., & Abugri, D. A. (2025). In Vitro Inhibitory Activity of Corilagin and Punicalagin Against Toxoplasma gondii and Their Mechanism(s) of Action. Antibiotics, 14(4), 336. https://doi.org/10.3390/antibiotics14040336