
Peptidome Profiling of Bubalus bubalis Urine and Assessment of Its Antimicrobial Activity against Mastitis-Causing Pathogens

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
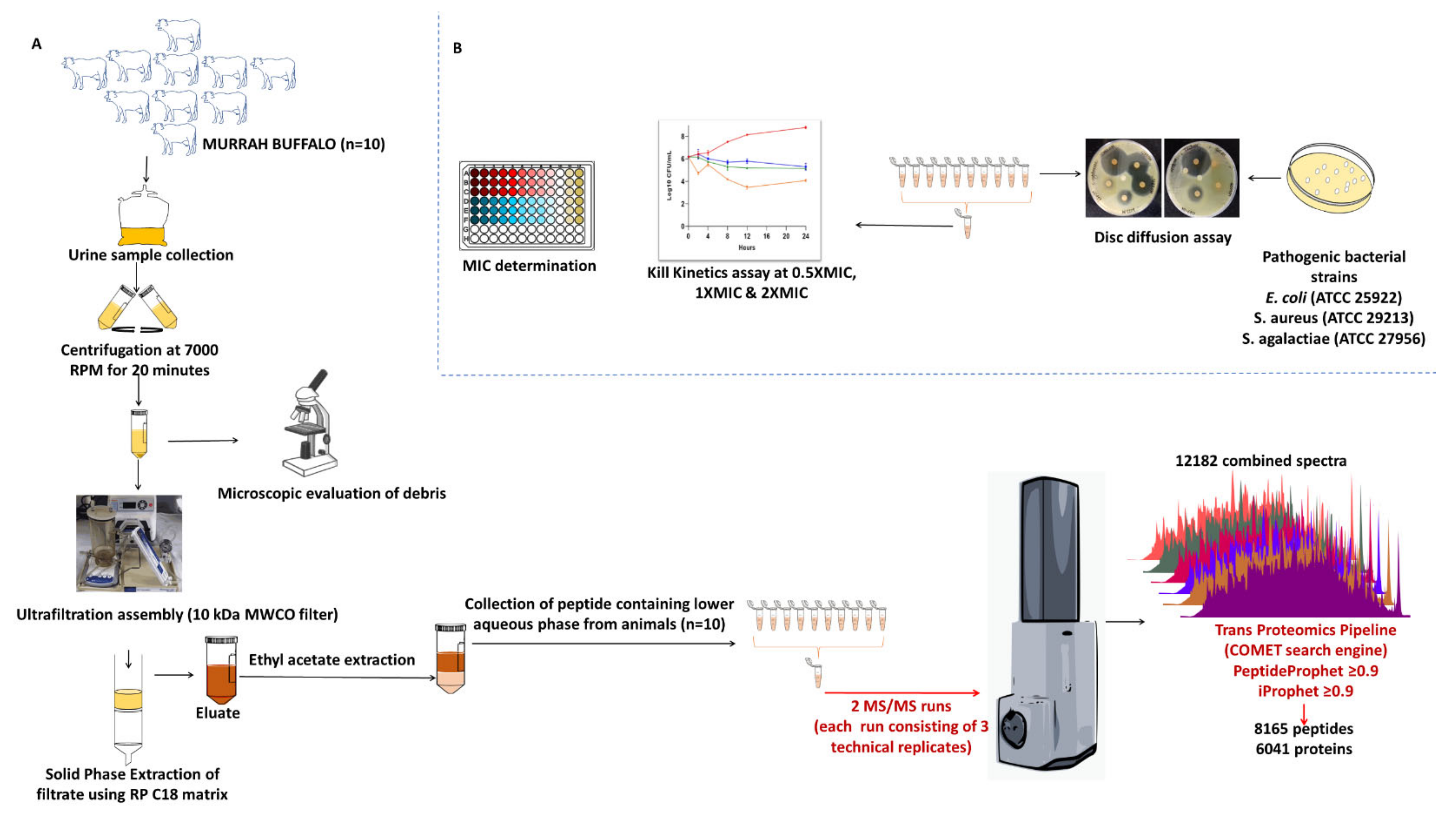
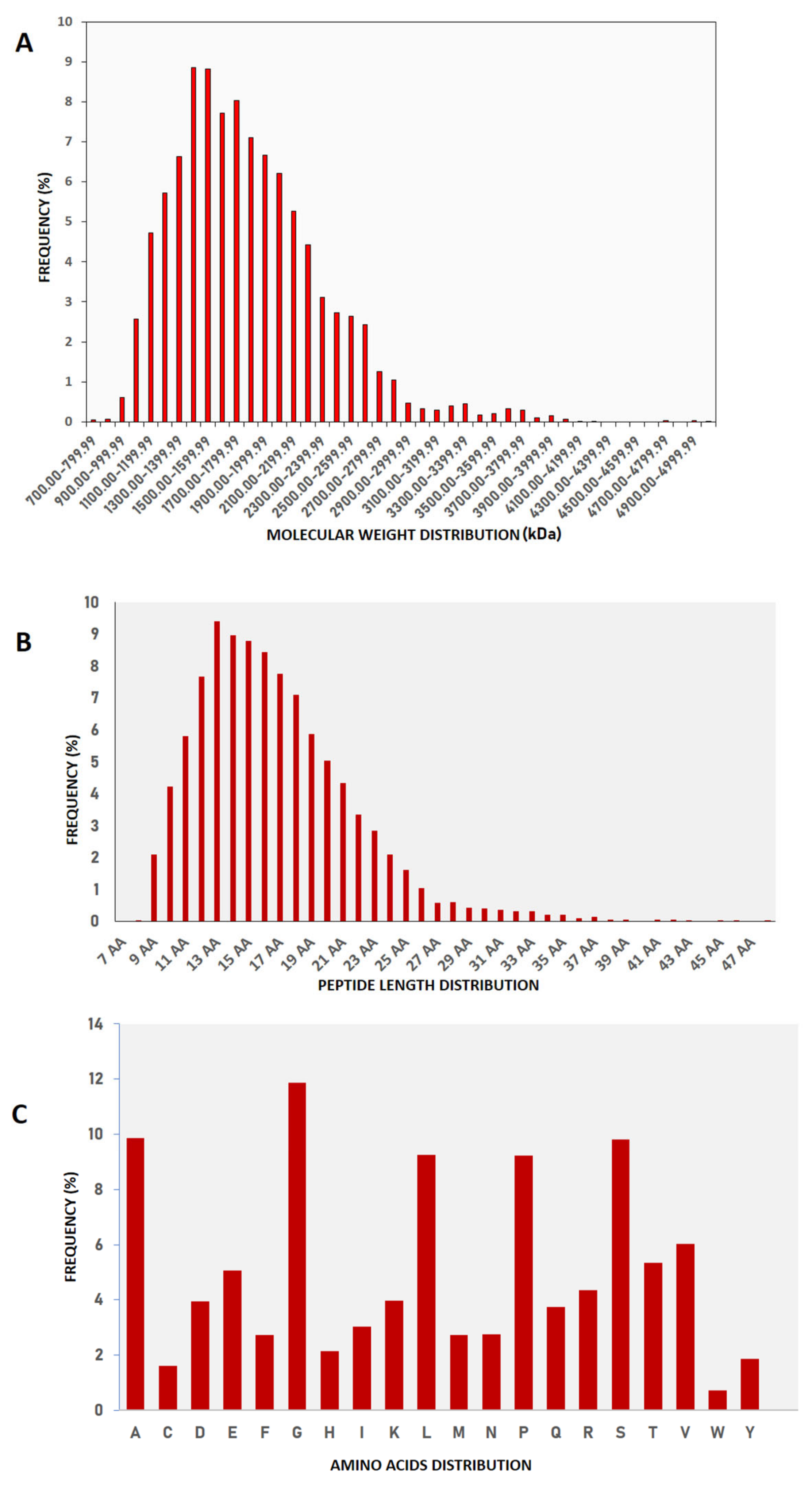
2.1. Molecular Nature of Buffalo Urinary Peptides
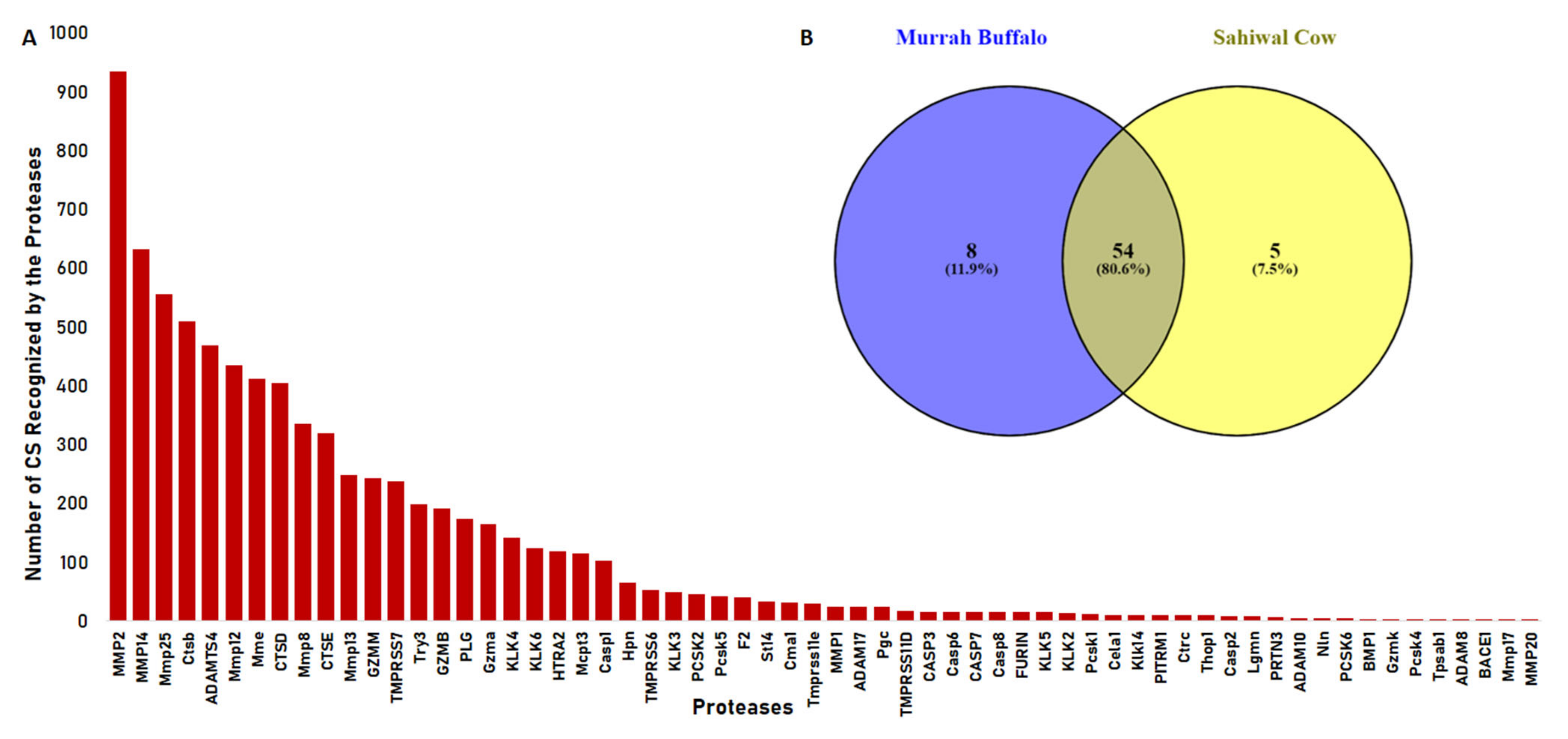
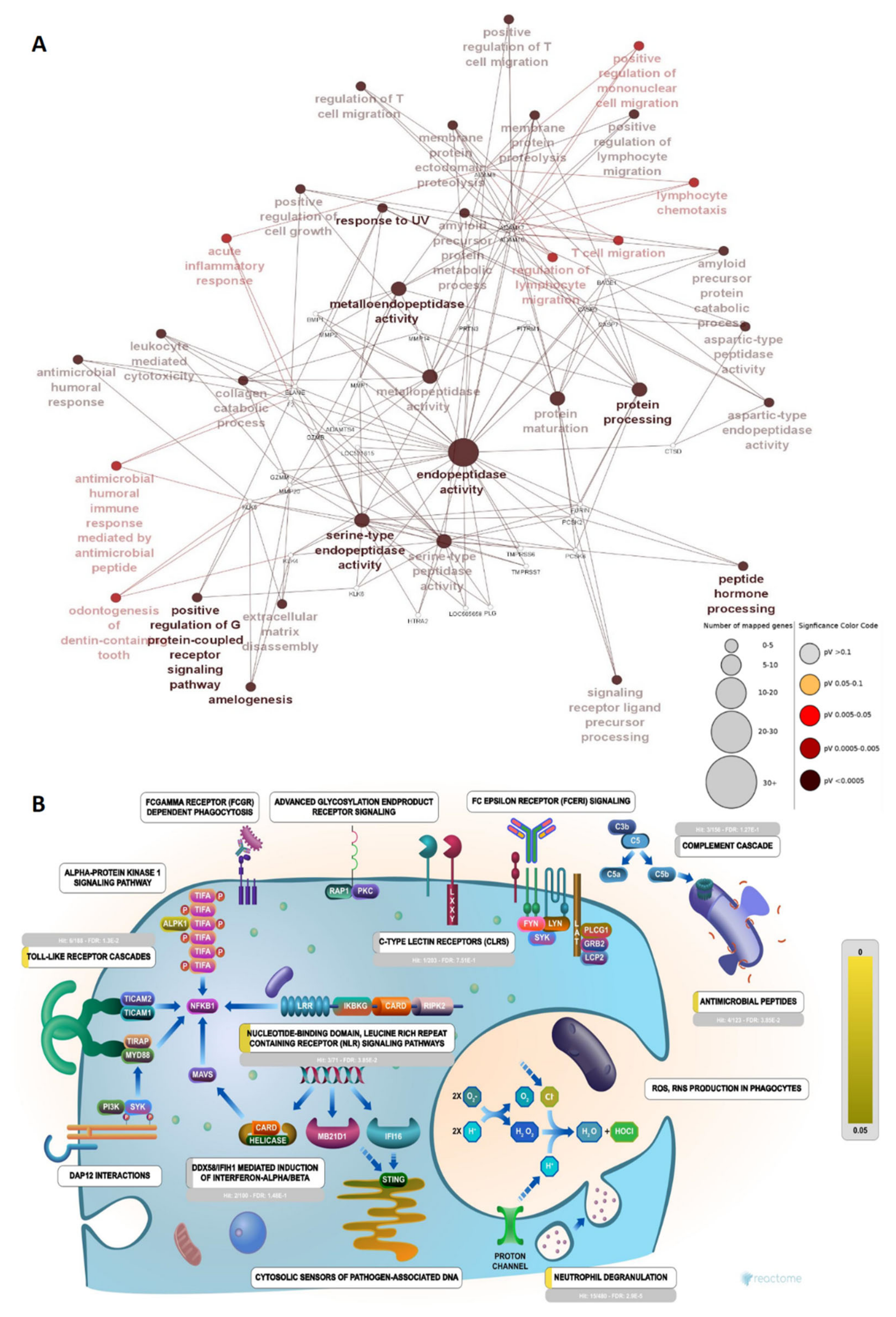
2.2. Proteases Involved in Urinary Peptide Generation
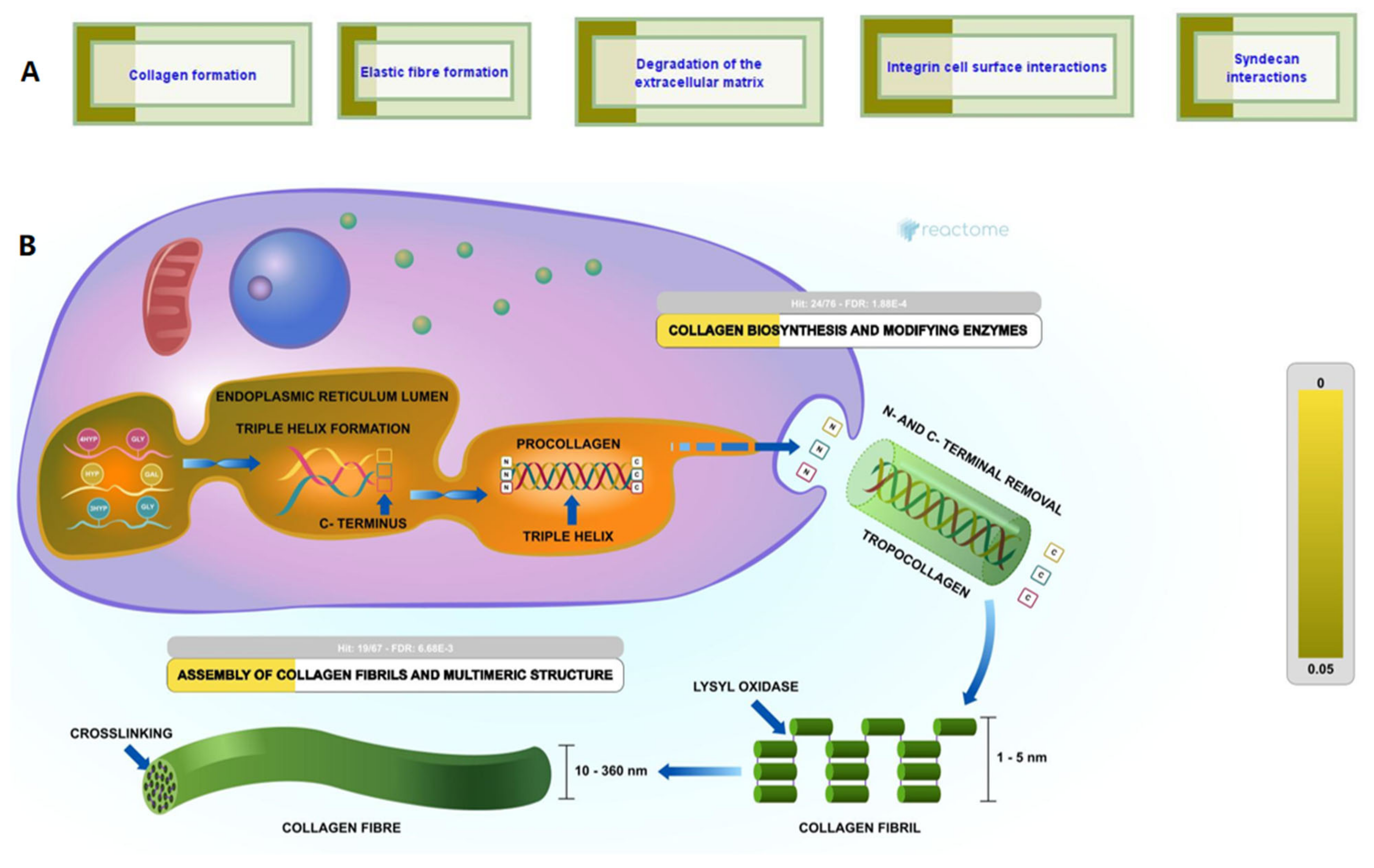
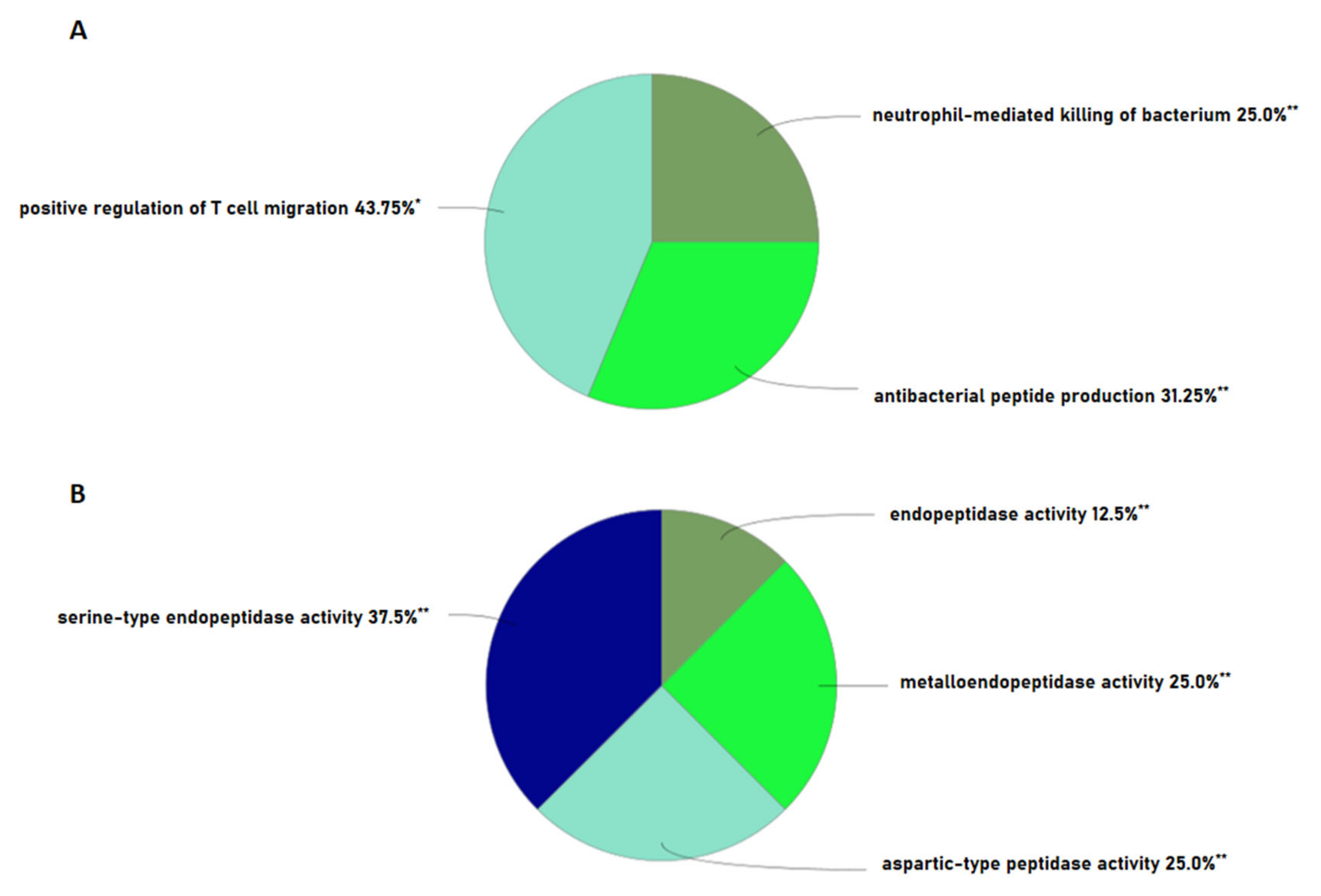
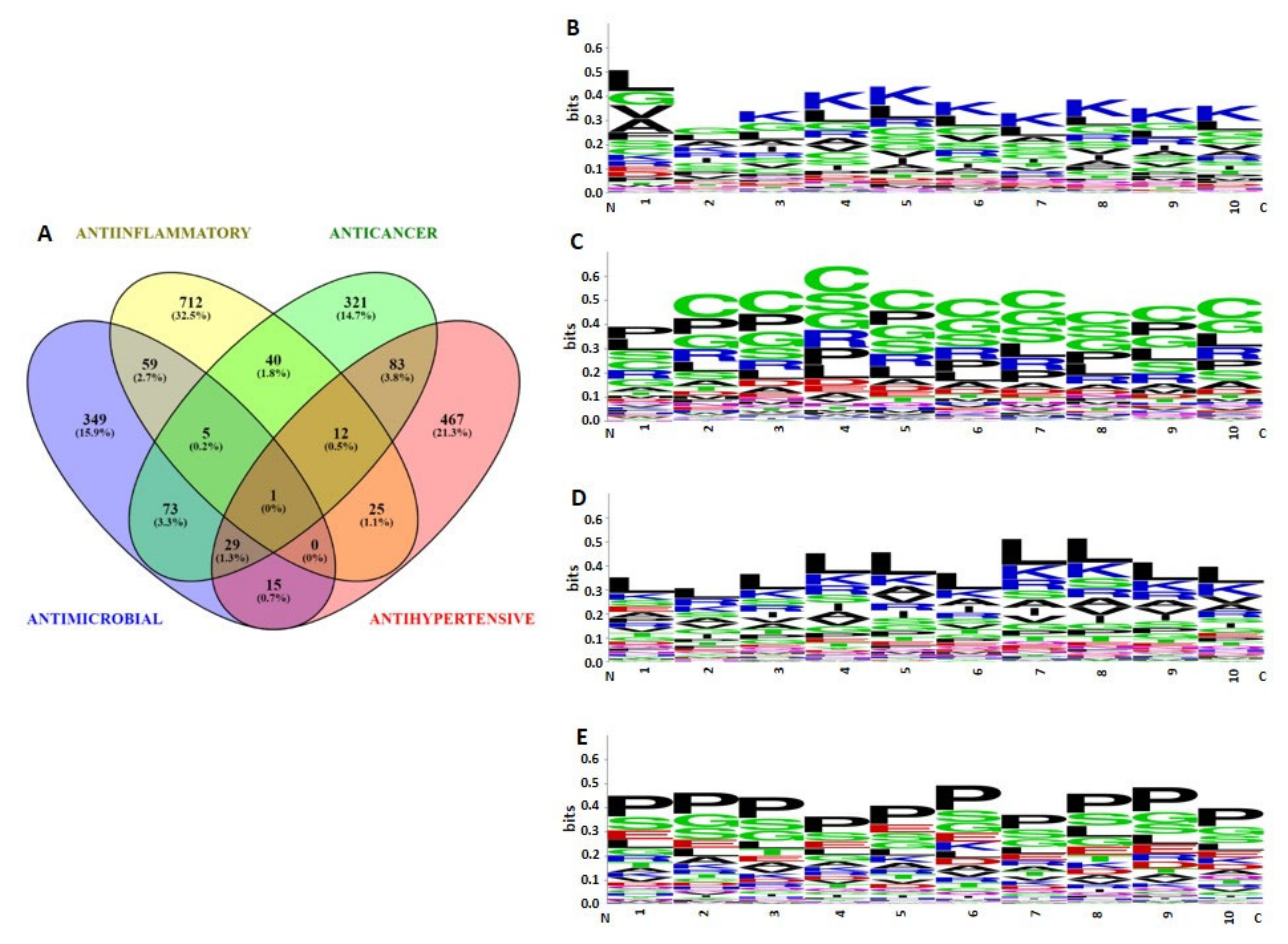
2.3. Bioactivities and Compositional Biases of the Predicted Functions
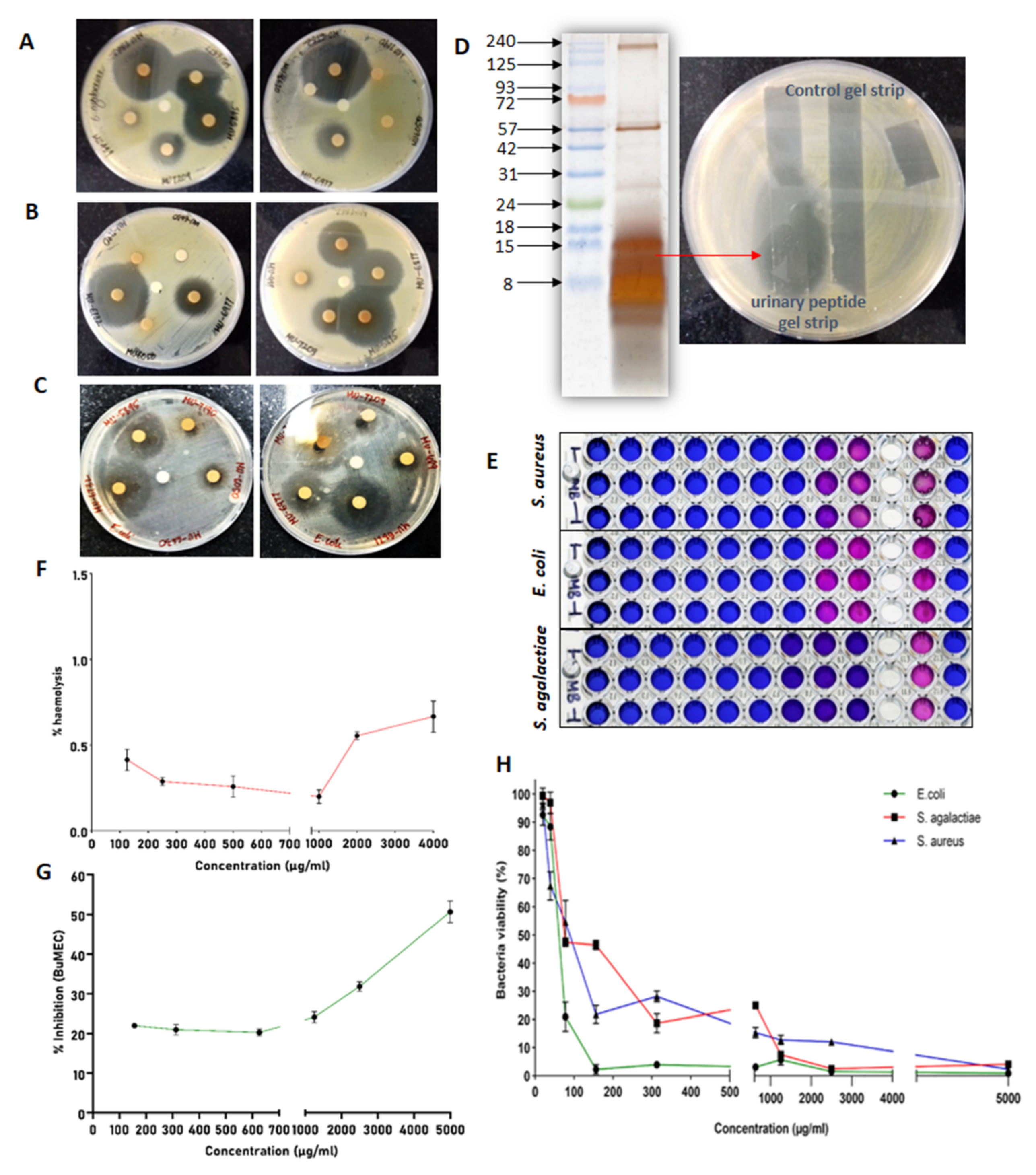
2.4. Antimicrobial Assay
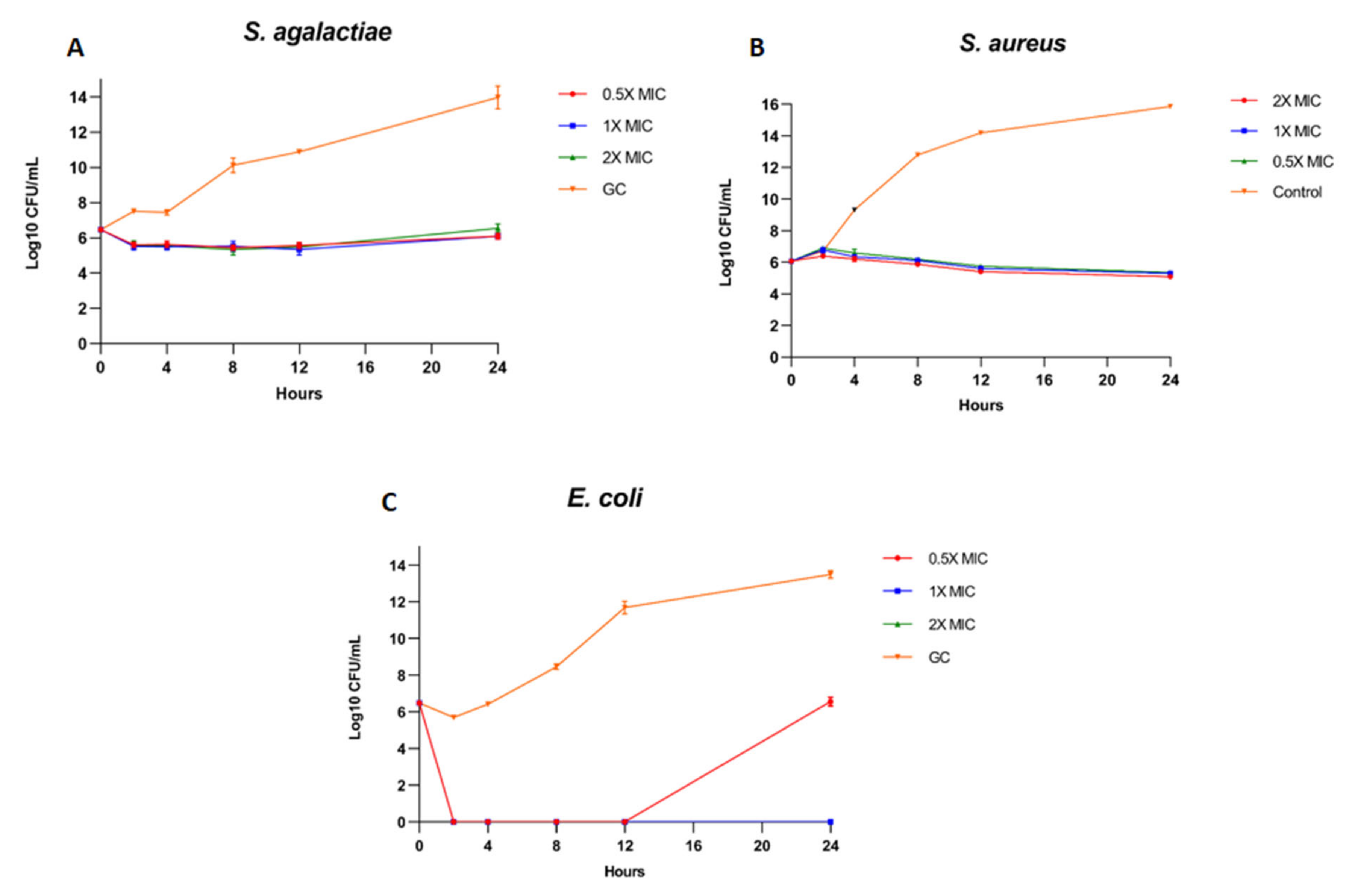
2.5. Kill Kinetics Assay
2.6. Haemolysis and Cytotoxicity Assay
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Processing
4.2. Peptide Extraction
4.3. LC-MS/MS Acquisition
4.4. Data Processing
4.5. Protease Prediction Using MEROPS Database
4.6. Bioactive Peptide Classification Using SVM Algorithm-Based Prediction Platforms
4.7. Antimicrobial Assay and MIC Determination
4.8. Kill Kinetics
4.9. Haemolysis Assay
4.10. Cytotoxicity Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez, V.; Ibernón, M.; López, D.; Pastor, M.C.; Navarro, M.; Navarro-Muñoz, M.; Bonet, J.; Romero, R. Urinary Peptide Profiling to Differentiate between Minimal Change Disease and Focal Segmental Glomerulosclerosis. PLoS ONE 2014, 9, e87731. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Pejchinovski, M.; Mullen, W.; Beige, J.; Mischak, H.; Jankowski, V.; He, T.; Pejchinovski, M.; Mischak, H.; Jankowski, V.; et al. Peptides in Plasma, Urine, and Dialysate: Toward Unravelling Renal Peptide Handling. Proteom.–Clin. Appl. 2021, 15, 2000029. [Google Scholar] [CrossRef]
- Martens, D.S.; Thijs, L.; Latosinska, A.; Trenson, S.; Siwy, J.; Zhang, Z.-Y.; Wang, C.; Beige, J.; Vlahou, A.; Janssens, S.; et al. Urinary Peptidomic Profiles to Address Age-Related Disabilities: A Prospective Population Study. Lancet Health Longev. 2021, 2, e690–e703. [Google Scholar] [CrossRef] [PubMed]
- Bathla, S.; Rawat, P.; Baithalu, R.; Yadav, M.L.; Naru, J.; Tiwari, A.; Kumar, S.; Balhara, A.K.; Singh, S.; Chaudhary, S.; et al. Profiling of Urinary Proteins in Karan Fries Cows Reveals More than 1550 Proteins. J. Proteom. 2015, 127, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Juanes, F.; Muñiz, M.C.; Raposo, C.; Rodríguez-Prieto, S.; Paradela, A.; Quiros, Y.; López-Hernández, F.; González-Buitrago, J.M.; Ferreira, L. Unveiling the Rat Urinary Proteome with Three Complementary Proteomics Approaches. Electrophoresis 2013, 34, 2473–2483. [Google Scholar] [CrossRef]
- Good, D.M.; Thongboonkerd, V.; Novak, J.; Bascands, J.L.; Schanstra, J.P.; Coon, J.J.; Dominiczak, A.; Mischak, H. Body Fluid Proteomics for Biomarker Discovery: Lessons from the Past Hold the Key to Success in the Future. J. Proteome Res. 2007, 6, 4549–4555. [Google Scholar] [CrossRef]
- Frantzi, M.; Van Kessel, K.E.; Zwarthoff, E.C.; Marquez, M.; Rava, M.; Malats, N.; Merseburger, A.S.; Katafigiotis, I.; Stravodimos, K.; Mullen, W.; et al. Development and Validation of Urine-Based Peptide Biomarker Panels for Detecting Bladder Cancer in a Multi-Center Study. Clin. Cancer Res. 2016, 22, 4077–4086. [Google Scholar] [CrossRef]
- Stalmach, A.; Johnsson, H.; McInnes, I.B.; Husi, H.; Klein, J.; Dakna, M.; Mullen, W.; Mischak, H.; Porter, D. Identification of Urinary Peptide Biomarkers Associated with Rheumatoid Arthritis. PLoS ONE 2014, 9, e104625. [Google Scholar] [CrossRef]
- Good, D.M.; Zürbig, P.; Argilés, À.; Bauer, H.W.; Behrens, G.; Coon, J.J.; Dakna, M.; Decramer, S.; Delles, C.; Dominiczak, A.F.; et al. Naturally Occurring Human Urinary Peptides for Use in Diagnosis of Chronic Kidney Disease. Mol. Cell Proteom. 2010, 9, 2424–2437. [Google Scholar] [CrossRef]
- Chromek, M.; Slamová, Z.; Bergman, P.; Kovács, L.; Podracká, L.; Ehrén, I.; Hökfelt, T.; Gudmundsson, G.H.; Gallo, R.L.; Agerberth, B.; et al. The Antimicrobial Peptide Cathelicidin Protects the Urinary Tract against Invasive Bacterial Infection. Nat. Med. 2006, 12, 636–641. [Google Scholar] [CrossRef]
- Spencer, J.D.; Schwaderer, A.L.; Dirosario, J.D.; McHugh, K.M.; McGillivary, G.; Justice, S.S.; Carpenter, A.R.; Baker, P.B.; Harder, J.; Hains, D.S. Ribonuclease 7 Is a Potent Antimicrobial Peptide within the Human Urinary Tract. Kidney Int. 2011, 80, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Da Spencer, J.; Hains, D.S.; Porter, E.; Bevins, C.L.; DiRosario, J.; Becknell, B.; Wang, H.; Schwaderer, A.L. Human Alpha Defensin 5 Expression in the Human Kidney and Urinary Tract. PLoS ONE 2012, 7, e31712. [Google Scholar] [CrossRef]
- Eichler, T.; Bender, K.; Murtha, M.J.; Schwartz, L.; Metheny, J.; Solden, L.; Jaggers, R.M.; Bailey, M.T.; Gupta, S.; Mosquera, C.; et al. Ribonuclease 7 Shields the Kidney and Bladder from Invasive Uropathogenic Escherichia Coli Infection. J. Am. Soc. Nephrol. 2019, 30, 1385–1397. [Google Scholar] [CrossRef]
- Babikir, I.H.; Abugroun, E.A.; Bilal, N.E.; Alghasham, A.A.; Abdalla, E.E.; Adam, I. The Impact of Cathelicidin, the Human Antimicrobial Peptide LL-37 in Urinary Tract Infections. BMC Infect. Dis. 2018, 18, 17. [Google Scholar] [CrossRef]
- Kumar, R.; Ali, S.A.; Singh, S.K.; Bhushan, V.; Mathur, M.; Jamwal, S.; Mohanty, A.K.; Kaushik, J.K.; Kumar, S. Antimicrobial Peptides in Farm Animals: An Updated Review on Its Diversity, Function, Modes of Action and Therapeutic Prospects. Vet. Sci. 2020, 7, 206. [Google Scholar] [CrossRef]
- Mookherjee, N.; Wilson, H.L.; Doria, S.; Popowych, Y.; Falsafi, R.; Yu, J.J.; Li, Y.; Veatch, S.; Roche, F.M.; Brown, K.L.; et al. Bovine and Human Cathelicidin Cationic Host Defense Peptides Similarly Suppress Transcriptional Responses to Bacterial Lipopolysaccharide. J. Leukoc. Biol. 2006, 80, 1563–1574. [Google Scholar] [CrossRef]
- Elsbach, P. What Is the Real Role of Antimicrobial Polypeptides That Can Mediate Several Other Inflammatory Responses? J. Clin. Investig. 2003, 111, 1643–1645. [Google Scholar] [CrossRef]
- Torres, P.; DÍaz, J.; Arce, M.; Silva, P.; Mendoza, P.; Lois, P.; Molina-Berríos, A.; Owen, G.I.; Palma, V.; Torres, V.A. The Salivary Peptide Histatin-1 Promotes Endothelial Cell Adhesion, Migration, and Angiogenesis. FASEB J. 2017, 31, 4946–4958. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, K.; Okumura, K.; Ogawa, H.; Niyonsaba, F. An Antimicrobial Peptide with Angiogenic Properties, AG-30/5C, Activates Human Mast Cells through the MAPK and NF-ΚB Pathways. Immunol. Res. 2016, 64, 594–603. [Google Scholar] [CrossRef]
- Zuyderduyn, S.; Ninaber, D.K.; Hiemstra, P.S.; Rabe, K.F. The Antimicrobial Peptide LL-37 Enhances IL-8 Release by Human Airway Smooth Muscle Cells. J. Allergy Clin. Immunol. 2006, 117, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Gamero, E.J.; Martins, M.N.C.; Cappabianco, F.A.M.; Ide, J.S.; Miranda, A. Characterization of Dual Effects Induced by Antimicrobial Peptides: Regulated Cell Death or Membrane Disruption. Biochim. Biophys. Acta-Gen. Subj. 2012, 1820, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zheng, Y.; Jia, J.; Li, C.; Duan, Q.; Li, R.; Wang, X.; Shao, Y.; Chen, C.; Yan, H. Antimicrobial Peptide LL-37 Promotes the Viability and Invasion of Skin Squamous Cell Carcinoma by Upregulating YB-1. Exp. Ther. Med. 2017, 14, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Mu, L.; Tang, J.; Liu, H.; Shen, C.; Rong, M.; Zhang, Z.; Lai, R. A Potential Wound-Healing-Promoting Peptide from Salamander Skin. FASEB J. 2014, 28, 3919–3929. [Google Scholar] [CrossRef]
- Imperlini, E.; Massaro, F.; Buonocore, F. Antimicrobial Peptides against Bacterial Pathogens: Innovative Delivery Nanosystems for Pharmaceutical Applications. Antibiotics 2023, 12, 184. [Google Scholar] [CrossRef]
- Sampaio de Oliveira, K.B.; Leite, M.L.; Rodrigues, G.R.; Duque, H.M.; da Costa, R.A.; Cunha, V.A.; de Loiola Costa, L.S.; da Cunha, N.B.; Franco, O.L.; Dias, S.C. Strategies for Recombinant Production of Antimicrobial Peptides with Pharmacological Potential. Expert Rev. Clin. Pharmacol. 2020, 13, 367–390. [Google Scholar] [CrossRef]
- Hoelscher, M.P.; Forner, J.; Calderone, S.; Krämer, C.; Taylor, Z.; Loiacono, F.V.; Agrawal, S.; Karcher, D.; Moratti, F.; Kroop, X.; et al. Expression Strategies for the Efficient Synthesis of Antimicrobial Peptides in Plastids. Nat. Commun. 2022, 13, 5856. [Google Scholar] [CrossRef]
- Ma, Y.; Guo, Z.; Xia, B.; Zhang, Y.; Liu, X.; Yu, Y.; Tang, N.; Tong, X.; Wang, M.; Ye, X.; et al. Identification of Antimicrobial Peptides from the Human Gut Microbiome Using Deep Learning. Nat. Biotechnol. 2022, 40, 921–931. [Google Scholar] [CrossRef] [PubMed]
- University of Glasgow-University News-Archive of News-2016-July-Scientists Identify Biomarkers of Cow Mastitis. Available online: https://www.gla.ac.uk/news/archiveofnews/2016/july/headline_475363_en.html (accessed on 3 February 2023).
- Banal, B.K.; Gupta, D.K. Economic Analysis of Bovine Mastitis in India and Punjab—A Review. Indian J. Dairy Sci. 2009, 62, 337–345. [Google Scholar]
- Shaheen, M.; Ha, T. A Treatise on Bovine Mastitis: Disease and Disease Economics, Etiological Basis, Risk Factors, Impact on Human Health, Therapeutic Management, Prevention and Control Strategy. Adv. Dairy Res. 2015, 4, 1–10. [Google Scholar] [CrossRef]
- Hu, S.; Loo, J.A.; Wong, D.T. Human Body Fluid Proteome Analysis. Proteomics 2006, 6, 6326–6353. [Google Scholar] [CrossRef]
- Kumar, R.; Ali, S.A.; Singh, S.K.; Bhushan, V.; Kaushik, J.K.; Mohanty, A.K.; Kumar, S. Peptide Profiling in Cow Urine Reveals Molecular Signature of Physiology-Driven Pathways and in-Silico Predicted Bioactive Properties. Sci. Rep. 2021, 11, 12427. [Google Scholar] [CrossRef] [PubMed]
- Coon, J.J.; Zürbig, P.; Dakna, M.; Dominiczak, A.F.; Decramer, S.; Fliser, D.; Frommberger, M.; Golovko, I.; Good, D.M.; Herget-Rosenthal, S.; et al. CE-MS Analysis of the Human Urinary Proteome for Biomarker Discovery and Disease Diagnostics. Proteom. Clin. Appl. 2008, 2, 964. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, P.; Pontillo, C.; Pejchinovski, M.; Siwy, J.; Krochmal, M.; Makridakis, M.; Carrick, E.; Klein, J.; Mullen, W.; Jankowski, J.; et al. Comparison of Urine and Plasma Peptidome Indicates Selectivity in Renal Peptide Handling. Proteom.-Clin. Appl. 2018, 12, 1700163. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.L.; Howard, P.S.; Koo, H.P.; Macarak, E.J. Role of Type III Collagen in Bladder Filling. Neurourol. Urodyn. 1998, 17, 135–145. [Google Scholar] [CrossRef]
- Macarak, E.J.; Howard, P.S. The Role of Collagen in Bladder Filling. Adv. Exp. Med. Biol. 1999, 462, 215–223. [Google Scholar] [CrossRef]
- Slack, M.A.; Gordon, S.M. Protease Activity in Vascular Disease. Arterioscler. Thromb. Vasc. Biol. 2019, 39, E210–E218. [Google Scholar] [CrossRef]
- Motta, J.-P.; Martin, L.; Vergnolle, N. Proteases/Antiproteases in Inflammatory Bowel Diseases. Proteases Their Recept. Inflamm. 2011, 173–215. [Google Scholar] [CrossRef]
- Twigg, M.S.; Brockbank, S.; Lowry, P.; Fitzgerald, S.P.; Taggart, C.; Weldon, S. The Role of Serine Proteases and Antiproteases in the Cystic Fibrosis Lung. Mediat. Inflamm. 2015, 2015, 293053. [Google Scholar] [CrossRef]
- Bastos, P.; Trindade, F.; Ferreira, R.; Casteleiro, M.A.; Stevens, R.; Klein, J.; Vitorino, R. Unveiling Antimicrobial Peptide–Generating Human Proteases Using PROTEASIX. J. Proteom. 2018, 171, 53–62. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and Function of Matrix Metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef]
- Shoombuatong, W.; Schaduangrat, N.; Nantasenamat, C. Unraveling the Bioactivity of Anticancer Peptides as Deduced from Machine Learning. EXCLI J. 2018, 17, 734–752. [Google Scholar] [CrossRef]
- Shanahan, M.T.; Vidrich, A.; Shirafuji, Y.; Dubois, C.L.; Henschen-Edman, A.; Hagen, S.J.; Cohn, S.M.; Ouellette, A.J. Elevated Expression of Paneth Cell CRS4C in Ileitis-Prone Samp1/Yitfc Mice: Regional Distribution, Subcellular Localization, And Mechanism Of Action. J. Biol. Chem. 2010, 285, 7493–7504. [Google Scholar] [CrossRef]
- Manavalan, B.; Shin, T.H.; Kim, M.O.; Lee, G. AIPpred: Sequence-Based Prediction of Anti-Inflammatory Peptides Using Random Forest. Front. Pharmacol. 2018, 9, 276. [Google Scholar] [CrossRef]
- Khatun, M.S.; Hasan, M.M.; Kurata, H. PreAIP: Computational Prediction of Anti-Inflammatory Peptides by Integrating Multiple Complementary Features. Front. Genet. 2019, 10, 129. [Google Scholar] [CrossRef]
- Singh, O.; Hsu, W.L.; Su, E.C.Y. Co-AMPpred for in Silico-Aided Predictions of Antimicrobial Peptides by Integrating Composition-Based Features. BMC Bioinform. 2021, 22, 389. [Google Scholar] [CrossRef]
- Chung, C.R.; Jhong, J.H.; Wang, Z.; Chen, S.; Wan, Y.; Horng, J.T.; Lee, T.Y. Characterization and Identification of Natural Antimicrobial Peptides on Different Organisms. Int. J. Mol. Sci. 2020, 21, 986. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.Y.; Lin, T.P.; Shih, L.Y.; Wang, C.K. Analysis and Prediction of the Critical Regions of Antimicrobial Peptides Based on Conditional Random Fields. PLoS ONE 2015, 10, e0119490. [Google Scholar] [CrossRef] [PubMed]
- Eom, K.H.; Li, S.; Lee, E.G.; Kim, J.H.; Kim, J.R.; Kim, I. Synthetic Polypeptides with Cationic Arginine Moieties Showing High Antimicrobial Activity in Similar Mineral Environments to Blood Plasma. Polymers 2022, 14, 1868. [Google Scholar] [CrossRef]
- Ma, R.; Mahadevappa, R.; Kwok, H.F.; Ma, R.; Mahadevappa, R.; Kwok, H.F. Venom-Based Peptide Therapy: Insights into Anti-Cancer Mechanism. Oncotarget 2017, 8, 100908–100930. [Google Scholar] [CrossRef]
- Shim, D.W.; Heo, K.H.; Kim, Y.K.; Sim, E.J.; Kang, T.B.; Choi, J.W.; Sim, D.W.; Cheong, S.H.; Lee, S.H.; Bang, J.K.; et al. Anti-Inflammatory Action of an Antimicrobial Model Peptide That Suppresses the TRIF-Dependent Signaling Pathway via Inhibition of Toll-like Receptor 4 Endocytosis in Lipopolysaccharide-Stimulated Macrophages. PLoS ONE 2015, 10, e0126871. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Yang, J.; He, X.; Mo, G.; Hong, J.; Yan, X.; Lin, D.; Lai, R. Structure and Function of a Potent Lipopolysaccharide-Binding Antimicrobial and Anti-Inflammatory Peptide. J. Med. Chem. 2013, 56, 3546–3556. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Cai, X.; Shi, W.; Bi, X.; Su, X.; Pan, M.; Li, H.; Lin, H.; Huang, W.; Qian, H. Pro-Apoptotic Cationic Host Defense Peptides Rich in Lysine or Arginine to Reverse Drug Resistance by Disrupting Tumor Cell Membrane. Amin. Acids 2017, 49, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Nasiri, F.; Atanaki, F.F.; Behrouzi, S.; Kavousi, K.; Bagheri, M. CpACpP: In Silico Cell-Penetrating Anticancer Peptide Prediction Using a Novel Bioinformatics Framework. ACS Omega 2021, 6, 19846–19859. [Google Scholar] [CrossRef]
- Ma, R.; Wong, S.W.; Ge, L.; Shaw, C.; Siu, S.W.I.; Kwok, H.F. In Vitro and MD Simulation Study to Explore Physicochemical Parameters for Antibacterial Peptide to Become Potent Anticancer Peptide. Mol. Ther. Oncolytics 2020, 16, 7–19. [Google Scholar] [CrossRef]
- Burdukiewicz, M.; Sidorczuk, K.; Rafacz, D.; Pietluch, F.; Bąkała, M.; Słowik, J.; Gagat, P. CancerGram: An Effective Classifier for Differentiating Anticancer from Antimicrobial Peptides. Pharmaceutics 2020, 12, 1045. [Google Scholar] [CrossRef] [PubMed]
- Tomar, V.; Nigam, R.; Pandey, V.; Singh, A.P.; Roy, D.; Sharma, A. Evaluation of in Vitro Anti-Microbial Activity of Goat Urine Peptides. J. Animal Res. 2018, 8, 33–37. [Google Scholar]
- Joseph, S.; Karnik, S.; Nilawe, P.; Jayaraman, V.K.; Idicula-Thomas, S. ClassAMP: A Prediction Tool for Classification of Antimicrobial Peptides. IEEE/ACM Trans. Comput. Biol. Bioinform. 2012, 9, 1535–1538. [Google Scholar] [CrossRef]
- Sharma, A.; Kapoor, P.; Gautam, A.; Chaudhary, K.; Kumar, R.; Chauhan, J.S.; Tyagi, A.; Raghava, G.P.S. Computational Approach for Designing Tumor Homing Peptides. Sci. Rep. 2013, 3, 1607. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, P.; Kumar, R.; Bhardwaj, A. dPABBs: A Novel in Silico Approach for Predicting and Designing Anti-Biofilm Peptides. Sci. Rep. 2016, 6, 21839. [Google Scholar] [CrossRef]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula-Thomas, S. CAMPR3: A Database on Sequences, Structures and Signatures of Antimicrobial Peptides. Nucleic Acids Res. 2016, 44, D1094. [Google Scholar] [CrossRef]
- Kumar, R.; Chaudhary, K.; Sharma, M.; Nagpal, G.; Chauhan, S.; Singh, S.; Gautam, A.; Raghava, G.P.S. AHTPDB: A Comprehensive Platform for Analysis and Presentation of Antihypertensive Peptides. Nucleic Acids Res. 2015, 43, 956–962. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, R.; Tyagi, N.; Nagpal, A.; Kaushik, J.K.; Mohanty, A.K.; Kumar, S. Peptidome Profiling of Bubalus bubalis Urine and Assessment of Its Antimicrobial Activity against Mastitis-Causing Pathogens. Antibiotics 2024, 13, 299. https://doi.org/10.3390/antibiotics13040299
Kumar R, Tyagi N, Nagpal A, Kaushik JK, Mohanty AK, Kumar S. Peptidome Profiling of Bubalus bubalis Urine and Assessment of Its Antimicrobial Activity against Mastitis-Causing Pathogens. Antibiotics. 2024; 13(4):299. https://doi.org/10.3390/antibiotics13040299
Chicago/Turabian StyleKumar, Rohit, Nikunj Tyagi, Anju Nagpal, Jai Kumar Kaushik, Ashok Kumar Mohanty, and Sudarshan Kumar. 2024. "Peptidome Profiling of Bubalus bubalis Urine and Assessment of Its Antimicrobial Activity against Mastitis-Causing Pathogens" Antibiotics 13, no. 4: 299. https://doi.org/10.3390/antibiotics13040299
APA StyleKumar, R., Tyagi, N., Nagpal, A., Kaushik, J. K., Mohanty, A. K., & Kumar, S. (2024). Peptidome Profiling of Bubalus bubalis Urine and Assessment of Its Antimicrobial Activity against Mastitis-Causing Pathogens. Antibiotics, 13(4), 299. https://doi.org/10.3390/antibiotics13040299