

Salmonellosis: An Overview of Epidemiology, Pathogenesis, and Innovative Approaches to Mitigate the Antimicrobial Resistant Infections

 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Epidemiology of Salmonellosis
2.1. Salmonella Serotypes and Host Spectrum
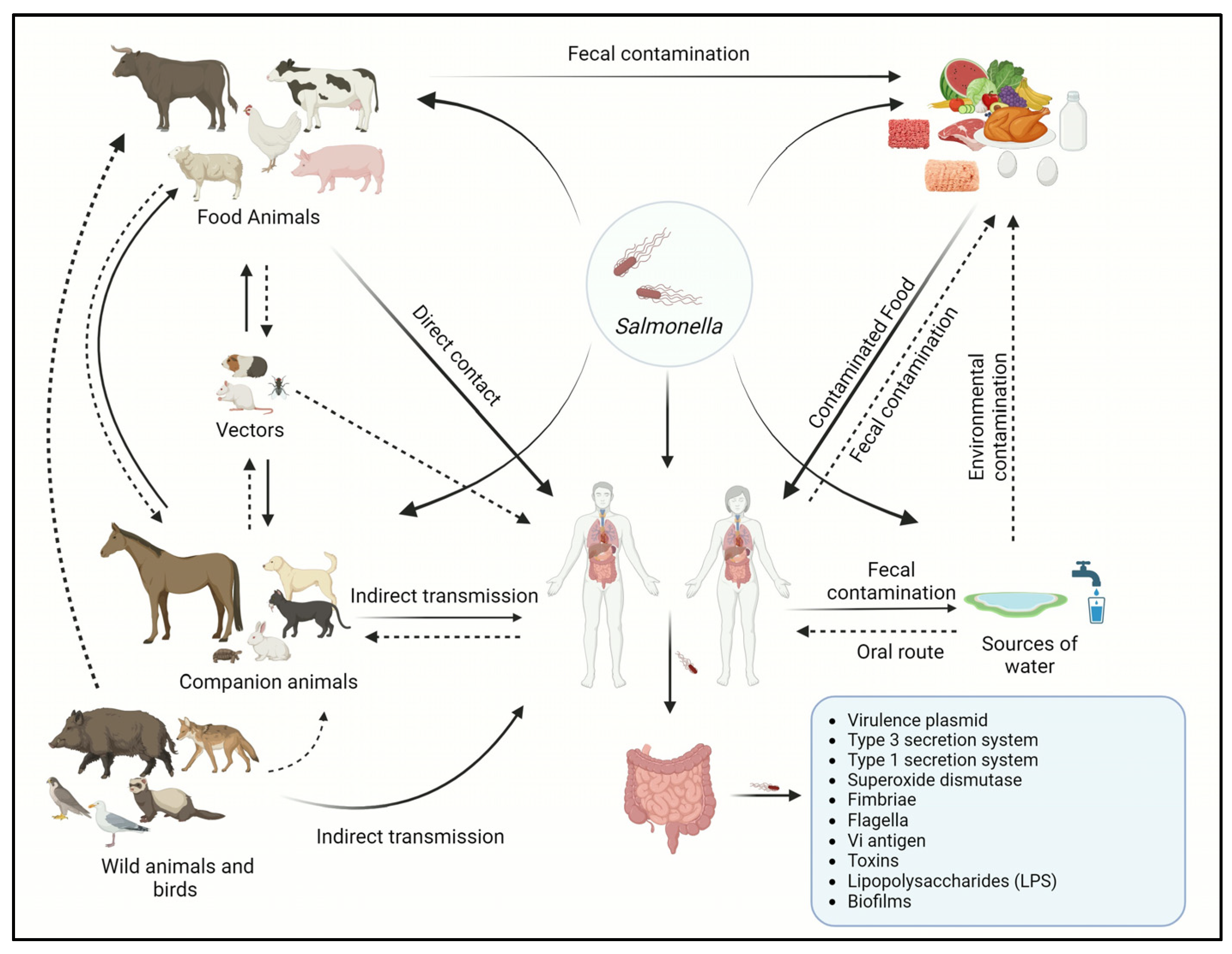
2.2. Source of Infection and Mode of Infection Transmission in Humans and Animals
2.3. Risk Factors and High-Risk Groups
2.4. Clinical Signs in Humans and Animals
2.4.1. In Humans
2.4.2. In Animals
2.5. Prevalence of Salmonellosis and the Most Recent Salmonella Outbreaks
3. Pathogenesis and Virulence Factors
3.1. Virulence Plasmid
3.2. Type III Secretion Systems
3.3. Type 1 Secretion System (T1SS)
3.4. Superoxide Dismutase
3.5. Fimbriae
3.6. Flagella
3.7. Vi Antigen
3.8. Toxins
3.9. Lipopolysaccharides (LPS)
3.10. Biofilms
4. Control Strategies for Salmonella Infections
4.1. Management and Biosecurity Measures
4.2. Vector Control and Eradication
4.3. Isolation and Quarantine
4.4. Antibiotics Used for Salmonella Treatment and Antimicrobial Resistance
4.5. Novel Antibiotic Alternatives
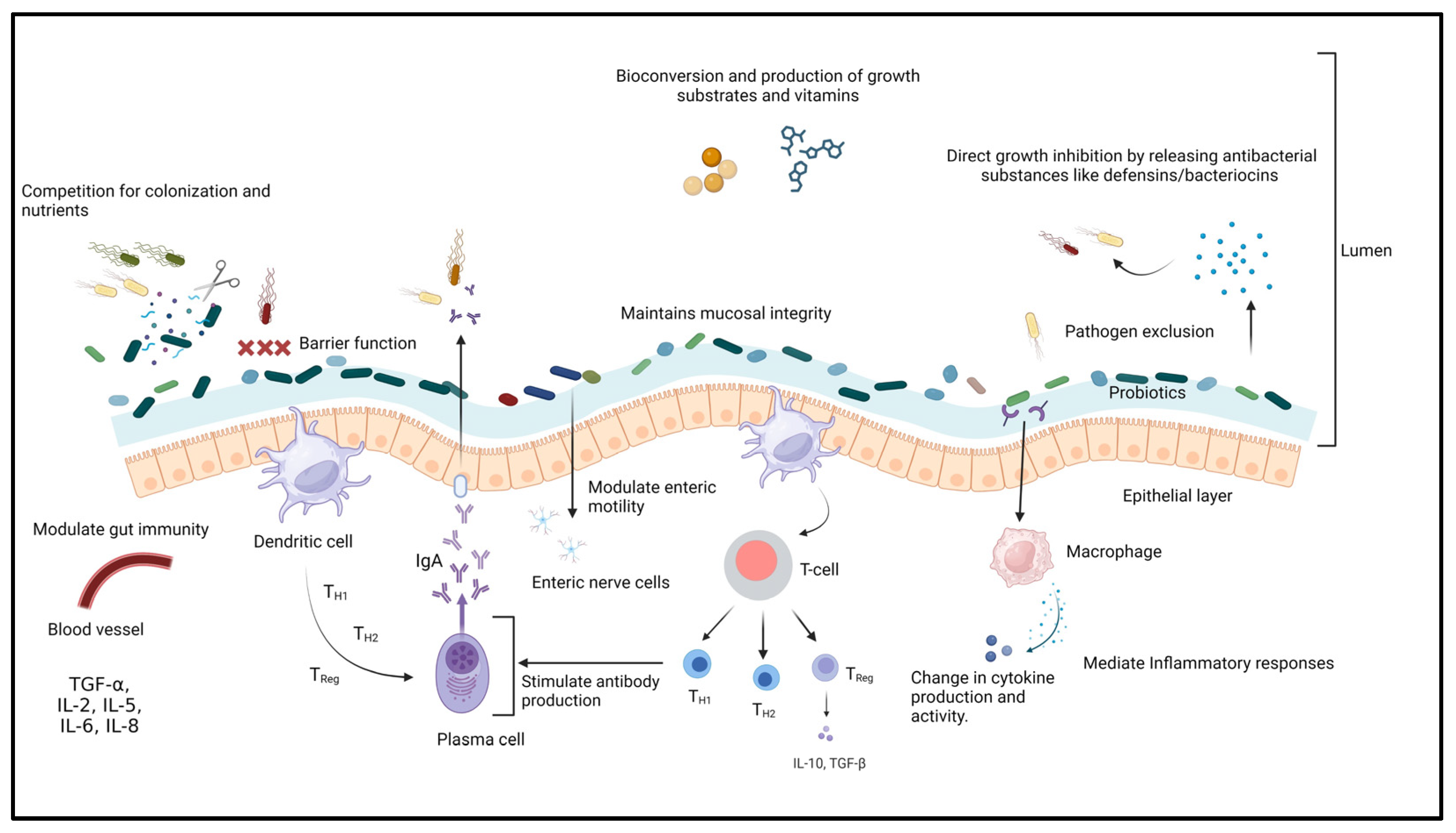
4.5.1. Probiotics

{kind=link}
{kind=link}
| Probiotics | Dose | Animal Host | Salmonella Serotype | Dose | Results | References |
|---|---|---|---|---|---|---|
| L. alvi An810, L. ingluviei An777, L. reuteri An769, and L. salivarius An63 | 107 cfu/mL | Chicken (male ISA Brown) | S. Enteritidis | 105 cfu/mL | No protective effect against S. Enteritidis in the host. | [272] |
| L. acidophilus LAP5, L. fermentum P2, Pediococcus acidilactici LS, and L. casei L21 | 107 CFU/mL | Broiler chicken | S. enterica subsp. Enterica ST19 | 108 Cfu/mL | Modulation of intestinal microbiota, increases intestinal villi height and short-chain fatty acids, restoring intestinal permeability by preventing tight junction damage. | [258] |
| L. reuteri, E. faecium, B. animalis, and P. acidilactici | 0.5 g/kg feed | Cobb broiler chickens | S. Enteritidis | 109 Cfu/mL | The growth and proliferation of S. Enteritidis decreased to 87.4–99.5% in vitro, and Salmonella load decreased by 0.85 and 1.5 log units/mL for cecal and carcass contents, respectively. | [268] |
| B. subtilis, B. licheniformis and Mannan oligosaccharide | 1.5 lbs/ton of feed | Hy-line layer hens | S. Enteritidis | 3 × 106 cfu/bird | A significant decrease (1.94 log reduction) in Salmonella colonization in the ceca. | [269] |
| E. faecium NCIMB 11181 | 4 × 108 cfu/kg of diet | Broiler chickens (Arbor | S. Typhimurium CVCC 2232 | 109 cfu/mL | Significant reduction in colonization and translocation of Salmonella in liver tissue (2.172 logs) and cecal content (4.2 logs) of infected birds pretreated with E. faecium. | [266] |
| L. salivarius CTC2197 | 105 cfu/mL | Leghorn chickens | S. Enteritidis C-114 | 108 cfu/mL | Complete clearance of Salmonella in chicken’s gut 21 days post-infection. | [262] |
| L. fermentum IKP 23, L. fermentum IKP 111 and L. salivarius IKP 333) | 107 cfu/mL | Broiler chickens | S. Enteritidis | 106 cfu/mL | Intestinal villus height was improved. Significantly high concentration of D-xylose in the plasma of broilers. | [273] |
| L. plantarum | 1.8 × 108 cfu/mL | Cobb broilers | S. Heidelberg | 2.5 × 108 cfu/mL | S. Heidelberg count was decreased in the caeca (2.1 log reduction). | [265] |
| L. salivarius L38 and L. acidophilus L36 | 109 cfu/mL | Swiss NIH mice | S. Typhimurium | 107 cfu/mL | No indication of protection against Salmonella isolates after pre-treatment with L36 or L38 probiotic strains. | [274] |
| L. reuteri R-17485, L. johnsonii R-17504 and L. vaginalis R-17362 | 2 × 108 cfu/mL | Lohmann White laying hens | S. Enteritidis | 104 cfu/mL | One-fold reduction in the cecal Salmonella count by L. reuteri R-17485, whereas significant (2-log) reduction by L. johnsonii R-17504. | [263,264] |
| L. reuteri, E. faecium, B. animalis, P. acidilactici and L. salivarius | 2 × 109 cfu/kg diet | Cobb broilers | S. Enteritidis | 6 × 105 cfu/mL | Administration of probiotics to birds resulted in 2.7 log reduction in Salmonella in the cecum. | [256] |
| L. acidophilus, B. bifidum, and Streptococcus faecalis | 1 × 105 to 1 × 106 cfu/mL | Female crossbred broiler | S. Typhimurium | 104 cfu/mL | Low- and high-dose treatment with probiotics resulted in 1.2 and 3 log reductions in S. Typhimurium load in chickens’ cecum, respectively, and decreased IFN-γ gene expression in the cecal tonsils of the treated chickens. | [275] |
| L. murinus, L. salivarius, L. pentosus, and P. pentosaceous | 4 × 109 cfu/mL | Pigs | S. Typhimurium | 108 cfu/mL | 2.4 log reduction (from 3.68 to 1.4 log CFU) in the fecal count of Salmonella. | [276] |
| L. fermentum and L. acidophilus | 108 cfu/mL | Mice | S. Typhimurium | 105 cfu/mL | No significant difference between treated and nontreated mice. | [277] |
| L. plantarum Z01 | 108 cfu/mL | Broiler chicken | S. Typhimurium | 108 cfu/0.2 mL | Significant reduction in Salmonella from the cecal content of treated chicken (5.24 out of 252 cfu × 105/g). | [278] |
| B. subtilis | 108 cfu/mL | Intestinal epithelium | S. Enteritidis, S. Typhimurium | 108 cfu/mL | High inhibition of S. Enteritidis (11–12 mm) and S. Typhimurium (11–15 mm zone of inhibition). | [257] |
| E. faecalis, C. butyricum, and B. mesentericus | 3.48 × 108, 2.0 × 107, 1.1 × 107 cfu/mL | Hospitalized infants and children | Salmonella spp. | - | Significant reduction (p < 0.0001) in diarrheal symptoms and severity of diarrhea significantly improved (p < 0.01) 3 days and no diarrhea was observed 5–7 days post-treatment. | [279] |
| B. subtilis RX7 and B. methylotrophicus C14 | 109 cfu/g | Weaned pigs | S. Typhimurium | 1011 cfu/mL | Salmonella counts in piglets after B. subtilis and B. methylotrophicus treatment have been reduced to 3.57–3.69 log cfu/g compared to the control group. | [280] |
| L. plantarum, L. casei, L. acidophilus, and E. faecium | 107 cfu/g | Horses | S. Typhimurium | - | Up to 65% reduction in fecal Salmonella shedding. | [281] |
| S. boulardii | 109 cfu/mL | Mice | S. Typhimurium | 105 cfu/mL | Enhanced survival up to 70% in treated mice as compared to 40% in untreated ones. Decreased Salmonella translocation, reduced liver damage, and decreased inflammatory cytokines | [267] |
| E. coli Nissle 1917 (EcN) | 109 cfu/mL | Day-old laying chicken | S. pullorum | 107 cfu/mL | Reduction of 2 log in the invasion of Salmonella in chicken fibroblast cells and 60% survival rate in EcN-treated group compared to 40% in the untreated ones. | [282] |
| L. lactis IBB 500, L. casei ŁOCK 0915, L. plantarum ŁOCK 0862 and S. cerevisiae | 109 cfu/mL | Ross-308 broiler chickens | S. Enteritidis | 105 cfu/mL | Reduction of 2-fold in cecal Salmonella 14 days post-infection followed by 0.5-fold reduction (p < 0.05) at 42 days post-infection. | [283,284] |
4.5.2. Prebiotics
4.5.3. Antimicrobial Peptides
4.5.4. Bacteriophages
| Phages | Target Serotypes | PFU/mL | Phase Application | Results | References |
|---|---|---|---|---|---|
| CNPSA1, CPNSA3, CNPSA4 | S. Enteritidis PT4 P125589 | 1011 | Single oral application of phage cocktail | Decrease in the occurrence of S. Enteritidis strains by 3.5 logs. | [337] |
| F1055S, F12013S | S. Enteriditis | 2 × 102 | Phage isolated and applied by aerosol spray on fertile eggs | Around 58% and 76% reduction in the cecal and visceral Salmonella count, respectively, without any loss in the body weight compared to the control group. | [339] |
| Φ st1 | S. Typhimurium and S. Hadar | 1012 | Intraclocal inoculation | Salmonella count reduced by 2.9 log10 CFU/mL within 6 h of challenge. S. Typhimurium had no trace of detection after 24 h. | [351] |
| SPGH1, SPGH3 | S. Typhimurium | 8.3 log10 | Spotted | S. Typhimurium count was significantly reduced by 4.2 log10. | [352] |
| UAB_Phi20, UAB_Phi78, UAB_Phi87 | S. Enteriditis and S. Typhimurium | 1011 | Oral | Cecal Salmonella count significantly decreased by 5.3 log upon administration of three phage cocktails one day before or after bacterial infection. | [353] |
| φ10, φ25, φ151 | S. Enteritidis P125109, Hadar 18, and Typhimurium 4/74 | 109−11 | Oral | Reduced cecal colonization by S. Enteritidis and S. Typhimurium by ≥4.2 log10 CFU and ≥2.9 log10 CFU, respectively. | [354] |
| Wide-Host-Range bacteriophages (WHR) | S. Enteritidis (SE), S. Typhimurium (ST) | 10⁹ | Sprayed with 5 mL of WHR and rinsed with sterile water | No bacteria were detected in two trials and a greater than 70% reduction was seen in the other two trials. | [355] |
| Bacteriophages of S. Typhimurium and S. enteritidis | S. Enteritidis (SE), and S. Typhimurium (ST) | 1.18 × 1011–1.03 × 102 | Oral | Moderate decrease (1 log reduction) in Salmonella loads 3 days post-infection (dpi), with a greater reduction of 2 log at 5 dpi and complete clearance of the bacteria at 7 dpi. | [356] |
| ΦCJ07 | S. Enteritidis (SE) | 105, 107 109 | Oral | After 3 weeks of treatment, no intestinal Salmonella was detected in 70% of hens treated with 109 PFU/g of bacteriophage. | [357] |
| PSE5 | S. Enteritidis (SE) | 4 × 107 | Immersion | Three logs reduction in Salmonella count was observed after 30 min of phage treatment of the contaminated eggs. | [358] |
| Pu20 | S. Pullorum | 108 or 109 | Direct inoculation | The phages demonstrated 1.06 and 1.12 log reduction in S. Pullorum in eggs stored at 4 °C and 25 °C, respectively. | [359] |
| UAB_Phi 20, UAB_Phi78, and UAB_Phi87 | S. Enteritidis (SE), S. Typhimurium (ST) | 109 and 1 × 1010 | Soaking in suspension and spraying | One log reduction in both S. Enteritidis and S. Typhimurium in chicken breast. | [343] |
| SEG5, SES8, STG2, STG5, and STS9 | S. Enteritidis, S. Typhimurium | 3 × 108 | Suspension added on the surface | S. Enteritidis and S. Typhimurium reduced by 3.06 and 2.21 log CFU/piece of chicken breast, respectively, | [344] |
| STGO-35-1 | S. Enteritidis | 4 × 106 | Direct addition | Significant reduction in Salmonella count (by 2.5 logs) in each piece of the chicken meat. | [360] |
4.5.5. Small Molecules and Quorum-Sensing Inhibitors
| Small Molecules | Action | Target Strains | Concentration/Dose | Effect on Quorum Sensing Regulatory Process/Growth Inhibition | References |
|---|---|---|---|---|---|
| Punicalagin | QSI | S. Typhimurium | 15.6 μg/mL | Downregulation of motility (flhC) and QS-associated genes (sdiA and srgE) | [388] |
| Carvacrol, Thymol, Eugenol | QSI | S. Enteritidis | 0.5 mM, 0.5 mM, 1.2 mM | Significant downregulation of genes related to host colonization (flgG, fimD, sopB, invH, and TTSS genes) and macrophage survival (ssaV and pipB) | [389] |
| Furanone | QSI | S. Typhimurium 14028 | 500 uM | Downregulation of quorum-sensing regulatory genes targets srgE and lsrA of sdiA and AI-2 followed by downregulation of genes related to flagellar biosynthesis and biofilms | [390] |
| M-gallate | QSI | S. Typhimurium | 128 μg/mL | Downregulation of QS-associated genes sdiA and srgE by 92.6 and 77.7% respectively. | [398] |
| Berberine | QSI | S. Typhimurium | 0.019 mg/mL | Reduction in AI-2 production by 73.5% compared to the control with exogenously supplied C4-HSL reporter molecule | [399] |
| Tannic acids | QSI | S. Typhi S. Paratyphi | 400 μg/mL | Drastically inhibited swarming motility, a major phenotype of quorum sensing without any impact on the growth of the bacteria | [400] |
| Xanthones | QSI | S. Typhimurium 21 SL1344 | 100 µM | A 60–70% inhibition in Ai-2 production, effective efflux pump inhibitors | [401] |
| N-(3-oxo octanoyl) DL-homoserine lactone | QSI | S. Typhimurium | 10 nM | SdiA gene downregulation and inhibition of biofilm formation | [392] |
| Homocysteine thiolactone | QSI | S. Typhimurium | 10 µM | Effect on SdiA gene expression with no effect on bacterial growth | [402] |
| Dephostatin | QSI | S. Typhimurium | 100 µM | SPI-2 virulence genes inhibitor and restoring sensitivity to the colistin | [403] |
| Fluorothiazinon | QSI | S. Typhimurium | 10 mg/kg | Suppression of Type-3 secretion system of Salmonella in vivo | [393] |
| Fusaric acid | QSI | S. Typhimurium | 100 µM | Type-3 secretion system inhibitor with anti-invasion activity | [394] |
| INP0007 and INP0403 | QSI | S. Typhimurium | 100 μM | Inhibition of Type-3 secretion system 1-associated virulence and invasion. | [395] |
| Cytosporone B | QSI | S. Typhimurium | 25 μM | Type-3 secretion system inhibition | [404] |
| Quercitrin | Growth inhibitors | S. Typhimurium | 32 μg/mL | Reduction in Salmonella adhesion, invasion, and survival in the HeLa cell lines by 70%. Blocks effector SipA translocation important for the invasion of the host cells | [370] |
| Imidazole, Methoxybenzylamine | Growth inhibitors | S. Typhimurium | 10 µM | Complete inhibition of growth and intracellular clearance of Salmonella from Caco-2, HD11, and THP-1 cell lines. Clearance of biofilm-embedded bacteria at 4 µM concentration | [371] |
| Compound 7955004 | Growth inhibitors | S. Typhimurium 14028 | 5 μM | More than 55% inhibition of preformed biofilms Complete clearance of the planktonic bacteria | [372] |
| SM4 (Imidazole class) SM 5 (Methoxybenzylamine class) | Growth inhibitors | S. Typhimurium | MIC: 10 µM and 25 µM | Bactericidal effect on WT S. Typhimurium with minimal toxicity on eukaryotic cell models including Caco-2, HD11, chicken macrophage cell lines, sheep or chicken RBCs, and complete clearance of internalized bacteria | [371] |
4.5.6. Vaccines
| Vaccines | Target Pathogens | Indications | Notable Observations | References |
|---|---|---|---|---|
| Vi Conjugate (Vi-rEPA) | S. Typhi | Human | Up to 90% efficacy in children between 2 and 5 years old. Rapid production of Vi-specific IgM and IgG with 2 logs reduction in shedding of the bacteria. | [432] |
| Modified live S. Dublin vaccine (EnterVene-d) | S. Dublin | Cattle | Stimulated cell-mediated immunity with antibody titer increased by 49% in vaccinated cows. Antibody titer increased by 88.56% in calves from the vaccinated cows, demonstrating strong horizontal transfer. | [435] |
| Ty21a | S. Typhi and S. Paratyphi B | Human | Cross-reactive multifunctional T-cell response with an increase in IgA production of 56% against S. Typhii and 38% against S. Paratyphi B compared to the control. | [424,429] |
| M01ZH09, Single dose independently attenuating deletion (S. Typhi (Ty2ΔaroCΔssaV) ZH9) | S. Typhi | Human | Rapid and high production of IgG and IgA with the fecal clearance of the bacteria within 7 days post-infection without any severe symptoms. | [434] |
| GMMA, Generalized Modules for Membrane Antigens | S. Typhimurium, S. Typhi, S. Paratyphi A | Human | Increased stimulation of peripheral blood mononuclear cells with increased IL-6 production. Elicit strong bacteriocidal anti-LPS O-antigen antibody and IgG production and complete clearance of the bacteria. | [37,40] |
| Vi Conjugate (Vi-CRM197) | S. Typhi | Human | Demonstrated 89% protective efficacy against typhoid fever and the protection lasted at least 4 years, significantly increased IgG antibody titer. | [433] |
| S. Typhi Vi polysaccharide tetanus toxoid conjugate vaccine (Tybar) | S. Typhi | Human | Robust anti-Vi IgG response in all age groups with significant protection across all age groups, including infants (children under the age of 2 years), with an efficacy of 85% without any side effects. | [431] |
| AviPro Megan Vac 1 + A12:E13 | S. Typhimurium, S. Enteritidis and S. Heidelberg | Poultry | Complete clearance of S. Enteritidis by 10 days post-infection with positive cases reduced to 6% on secondary inoculation. No vertical transfer of the antibodies observed. | [442] |
| Chitosan-adjuvanted Salmonella subunit nanoparticle vaccine (OMPs-F-CS NPs) | S. Enteritidis | Poultry | Upregulation of TLRs and Th1 and Th2 cytokine mRNA with increased OMPs-specific IgY and IgA antibodies in saliva and intestine on oral administration. Salmonella shedding was reduced by 7 times compared to the mock challenge. | [436] |
| Inactivated trivalent Salmonella enterica vaccine (Nobilis® Salenvac T; Intervet International B.V., Boxmeer, The Netherlands) | S. Typhimurium, S. Enteritidis and S. Infantis | Poultry | A 3.9 log increase in mean antibody titer upon administration of the booster dose in chicken with 2.6 log reduction in cecal shedding of S. Typhimurium and S. Enteritidis, followed by 1.3 log reduction in S. Infantis | [438] |
| Poulvac® ST (Zoetis Inc. New Jersey, USA) | S. Typhimurium, S. Kentucky, S. Enteritidis, S. Heidelberg and S. Hadar | Poultry | A % reduction in S. Kentucky, S. Enteritidis, S. Heidelberg, S. Typhimurium, and S. Hadar in liver and spleen, with cross-protection between all 5 strains. | [437] |
| Autologous killed trivalent vaccine (Tri-Vaccine) | S. Typhimurium, S. Enteritidis and S. Heidelberg | Poultry | In total, 58% of the cloacal swabs from the infected birds demonstrated complete clearance of the bacteria 8 days post-infection. | [442] |
4.5.7. Organic Acids
4.5.8. Essential Oils (EOs)
| Plant | Major Components | Salmonella Serotype | MIC | Activity | References |
|---|---|---|---|---|---|
| Thymus vulgaris | Thymol (37.5%), p-cymene (14.49%), γ-terpinene (11.15%), linalool (4.71%), and carvacrol (4.62%) | S. Typhimurium ATCC 14028 | 0.25% v/v | The zone of inhibition in the agar-well diffusion assay was found to be 25.5 mm against S. Typhimurium, with complete clearance of the bacteria at MIC 0.25% v/v | [484] |
| Origanum vulgare | Thymol- and carvacrol-based EO | S. enteritidis ATCC 13076 | 120 μg/mL (carvacrol), 130 μg/mL (Thymol) | Complete clearance of the bacteria at 120 μg/mL (carvacrol) and 130 μg/mL (Thymol) in vitro | [485] |
| Pistacia atlantica subsp. Kurdica | α-Pinene (10.8%) | S. Typhimurium ATCC 14028 | 0.26 mg/mL | Complete clearance of S. Typhimurium was found to be at 0.5 mL/mL with a zone of inhibition of 22 mm | [486] |
| Cinnamomum verum | Not identified | S. enteritidis, S. Typhimurium, S. Heidelberg | >20 μL/mL | The zone of inhibition of all of the strains was found to be higher than 20 mm on agar well diffusion assay. | [487] |
| Citrus medica L. Var. Sarcodactylis | d-Limonene terpinene | S. Typhimurium | 2.0 mg/mL | The zone of inhibition was found to be 20 mm and the inhibition of biofilm formation was found to be 90%. | [58] |
| Ocimum basilicum | linalool, 1,8-cineole, eugenol, α-terpineol, ρ-cymene, and germacrene D | S. Enteritidis | 20 μg/mL | Two log reduction in the number of Salmonella when used in food products, colonization resistance was evident | [488] |
| Allium sativum (Garlic) | diallyl disulfide | S. Typhimurium | MIC/8 (1/512) μg/mL | Inhibition of biofilm formation by 23%, downregulation of virulence genes including invA and sdiA genes | [489] |
| Commercially available Essential oils. | Thymol, carvacrol, cinnamaldehyde | S. Enteritidis | 4.6 mg/mL | Complete clearance of illeal and cecal Salmonella in broiler chicken at 10 dpi, improved illeal integrity, gut immunity modulation | [33] |
| Aniba rosaeodora | Linaloo | S. Typhimurium S. Pullorum | 4 mg/mL 8 mg/mL | In vitro: complete clearance of the bacteria at 4 mg/mL and 8 mg/mL, respectively In vivo: complete clearance and systemic protection in chicks, modulate host inflammatory process | [490] |
| Cymbopogon citrates (Lemongrass) | Neral, Citral, Geranyl acetate | S. Newport | --- | Significant reduction in bacterial population by 1 log CFU/g when co-cultured with S. Newport | [491] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Center for Disease Control and Prevention (CDC). Salmonella Infection. Available online: https://www.cdc.gov/healthypets/diseases/salmonella.html (accessed on 11 July 2023).
- Sanchez-Vargas, F.M.; Abu-El-Haija, M.A.; Gomez-Duarte, O.G. Salmonella infections: An update on epidemiology, management, and prevention. Travel Med. Infect. Dis. 2011, 9, 263–277. [Google Scholar] [CrossRef]
- Wiedemann, A.; Virlogeux-Payant, I.; Chaussé, A.-M.; Schikora, A.; Velge, P. Interactions of Salmonella with animals and plants. Front. Microbiol. 2015, 5, 791. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; El-Adawy, H.; Abdelwhab, E.M. A Comprehensive Review of Common Bacterial, Parasitic and Viral Zoonoses at the Human-Animal Interface in Egypt. Pathogens 2017, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Baptista, D.; Borsoi, A.; Reischak, D.; Nascimento, A.; Montesino, L.; Camillo, S.; Abreu, D.; Pereira, V. Salmonella Serovars Isolated from Poultry Breeding Flocks under the Brazilian Official Control Programme Between 2016 and 2018. Braz. J. Poult. Sci. 2023, 25, 1. [Google Scholar] [CrossRef]
- Balasubramanian, R.; Im, J.; Lee, J.S.; Jeon, H.J.; Mogeni, O.D.; Kim, J.H.; Rakotozandrindrainy, R.; Baker, S.; Marks, F. The global burden and epidemiology of invasive non-typhoidal Salmonella infections. Hum. Vaccines Immunother. 2019, 15, 1421–1426. [Google Scholar] [CrossRef]
- Hoffmann, S.; Batz, M.B.; Morris, J.G., Jr. Annual Cost of Illness and Quality-Adjusted Life Year Losses in the United States Due to 14 Foodborne Pathogens. J. Food Prot. 2012, 75, 1292–1302. [Google Scholar] [CrossRef]
- Chlebicz, A.; Śliżewska, K. Campylobacteriosis, salmonellosis, yersiniosis, and listeriosis as zoonotic foodborne diseases: A review. Int. J. Environ. Res. Public Health 2018, 15, 863. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention (CDC). Salmonella. Available online: https://www.cdc.gov/salmonella/index.html#:~:text=CDC%20estimates%20Salmonella%20bacteria%20cause,%2C%20fever%2C%20and%20stomach%20cramps (accessed on 11 July 2023).
- Center for Disease Control and Prevention (CDC). Drug-Resistant Nontyphoidal Salmonella. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/nt-salmonella-508.pdf (accessed on 12 July 2023).
- National Institute of Allergy and Infectious Diseases (NIAID). Available online: https://www.niaid.nih.gov/research/emerging-infectious-diseases-pathogens (accessed on 15 July 2023).
- Wei, X.; Long, L.; You, L.; Wang, M.; Wang, D.; Liu, C.; Li, S.; Wang, J. Serotype distribution, trend of multidrug resistance and prevalence of β-lactamase resistance genes in human Salmonella isolates from clinical specimens in Guizhou, China. PLoS ONE 2023, 18, e0282254. [Google Scholar] [CrossRef]
- Keestra-Gounder, A.M.; Tsolis, R.M.; Bäumler, A.J. Now you see me, now you don’t: The interaction of Salmonella with innate immune receptors. Nat. Rev. Microbiol. 2015, 13, 206–216. [Google Scholar] [CrossRef]
- Hoelzer, K.; Moreno Switt, A.I.; Wiedmann, M. Animal contact as a source of human non-typhoidal salmonellosis. Vet. Res. 2011, 42, 34. [Google Scholar] [CrossRef]
- Bula-Rudas, F.J.; Rathore, M.H.; Maraqa, N.F. Salmonella Infections in Childhood. Adv. Pediatr. 2015, 62, 29–58. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Sjolund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Epidemiology, Clinical Presentation, Laboratory Diagnosis, Antimicrobial Resistance, and Antimicrobial Management of Invasive Salmonella Infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [PubMed]
- Giannella, R.A. Salmonella. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Cohen, J.I.; Bartlett, J.A.; Corey, G.R. Extra-intestinal manifestations of salmonella infections. Medicine 1987, 66, 349–388. [Google Scholar] [CrossRef]
- Wibisono, F.M.; Wibisono, F.J.; Effendi, M.H.; Plumeriastuti, H.; Hidayatullah, A.R.; Hartadi, E.B.; Sofiana, E.D. A Review of Salmonellosis on Poultry Farms: Public Health Importance. Syst. Rev. Pharm. 2020, 11, 481–486. [Google Scholar]
- World Health Organization (WHO). Salmonella (Non-Typhoidal). Available online: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) (accessed on 12 August 2023).
- Olnood, C.G.; Beski, S.S.; Choct, M.; Iji, P.A. Use of Lactobacillus johnsonii in broilers challenged with Salmonella sofia. Anim. Nutr. 2015, 1, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Tie, K.; Yuan, Y.; Yan, S.; Yu, X.; Zhang, Q.; Xu, H.; Zhang, Y.; Gu, J.; Sun, C.; Lei, L.; et al. Isolation and identification of Salmonella pullorum bacteriophage YSP2 and its use as a therapy for chicken diarrhea. Virus Genes 2018, 54, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Sania, T.; Abdul, S.; Muhammad, H.; Areeb, A.; Ayesha, M.; Shehroz, A.; Abdelslam Masoud Abobakr, A. Salmonella in Poultry; an Overview. Int. J. Multidiscip. Sci. Arts 2022, 1, 80–84. [Google Scholar] [CrossRef]
- Velasquez, C.G.; Macklin, K.S.; Kumar, S.; Bailey, M.; Ebner, P.E.; Oliver, H.F.; Martin-Gonzalez, F.S.; Singh, M. Prevalence and antimicrobial resistance patterns of Salmonella isolated from poultry farms in southeastern United States. Poult. Sci. 2018, 97, 2144–2152. [Google Scholar] [CrossRef]
- Holschbach, C.L.; Peek, S.F. Salmonella in Dairy Cattle. Vet. Clin. Food Anim. Pract. 2018, 34, 133–154. [Google Scholar] [CrossRef]
- Chen, H.-M.; Wang, Y.; Su, L.-H.; Chiu, C.-H. Nontyphoid Salmonella Infection: Microbiology, Clinical Features, and Antimicrobial Therapy. Pediatr. Neonatol. 2013, 54, 147–152. [Google Scholar] [CrossRef]
- Galán, J.E. Salmonella Typhimurium and inflammation: A pathogen-centric affair. Nat. Rev. Microbiol. 2021, 19, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Stoycheva, M.V.; Murdjeva, M.A. Antimicrobial therapy of salmonelloses—Current state and perspectives. Folia Medica 2006, 48, 5–10. [Google Scholar] [PubMed]
- Jibril, A.H.; Okeke, I.N.; Dalsgaard, A.; Olsen, J.E. Association between antimicrobial usage and resistance in Salmonella from poultry farms in Nigeria. BMC Vet. Res. 2021, 17, 234. [Google Scholar] [CrossRef] [PubMed]
- Ashurst, J.V.; Truong, J.; Woodbury, B. Salmonella Typhi. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Hailu, W.; Helmy, Y.A.; Carney-Knisely, G.; Kauffman, M.; Fraga, D.; Rajashekara, G. Prevalence and Antimicrobial Resistance Profiles of Foodborne Pathogens Isolated from Dairy Cattle and Poultry Manure Amended Farms in Northeastern Ohio, the United States. Antibiotics 2021, 10, 1450. [Google Scholar] [CrossRef] [PubMed]
- Van Panhuis, W.G.; Grefenstette, J.; Jung, S.Y.; Chok, N.S.; Cross, A.; Eng, H.; Lee, B.Y.; Zadorozhny, V.; Brown, S.; Cummings, D. Contagious diseases in the United States from 1888 to the present. N. Engl. J. Med. 2013, 369, 2152. [Google Scholar] [CrossRef]
- Sears, K.T.; Galen, J.E.; Tennant, S.M. Advances in the development of Salmonella-based vaccine strategies for protection against Salmonellosis in humans. J. Appl. Microbiol. 2021, 131, 2640–2658. [Google Scholar] [CrossRef] [PubMed]
- Miletic, S.; Goessweiner-Mohr, N.; Marlovits, T.C. The structure of the type III secretion system needle complex. Curr. Top Microbiol. Immunol. 2020, 427, 67–90. [Google Scholar]
- Gordon, M.A.; Graham, S.M.; Walsh, A.L.; Wilson, L.; Phiri, A.; Molyneux, E.; Zijlstra, E.E.; Heyderman, R.S.; Hart, C.A.; Molyneux, M.E. Epidemics of invasive Salmonella enterica serovar enteritidis and S. enterica Serovar typhimurium infection associated with multidrug resistance among adults and children in Malawi. Clin. Infect. Dis. 2008, 46, 963–969. [Google Scholar] [CrossRef]
- Milligan, R.; Paul, M.; Richardson, M.; Neuberger, A. Vaccines for preventing typhoid fever. Cochrane Database Syst. Rev. 2018, 5, Cd001261. [Google Scholar] [CrossRef]
- Giannelli, C.; Cappelletti, E.; Di Benedetto, R.; Pippi, F.; Arcuri, M.; Di Cioccio, V.; Martin, L.B.; Saul, A.; Micoli, F. Determination of free polysaccharide in Vi glycoconjugate vaccine against typhoid fever. J. Pharm. Biomed. Anal. 2017, 139, 143–147. [Google Scholar] [CrossRef]
- De Benedetto, G.; Salvini, L.; Gotta, S.; Cescutti, P.; Micoli, F. Investigation on Sugar–Protein Connectivity in Salmonella O-Antigen Glycoconjugate Vaccines. Bioconjugate Chem. 2018, 29, 1736–1747. [Google Scholar] [CrossRef] [PubMed]
- Maddux, J.T.; Stromberg, Z.R.; Curtiss III, R.; Mellata, M. Evaluation of Recombinant Attenuated Salmonella Vaccine Strains for Broad Protection against Extraintestinal Pathogenic Escherichia coli. Front. Immunol. 2017, 8, 1280. [Google Scholar] [CrossRef]
- Mancini, F.; Micoli, F.; Necchi, F.; Pizza, M.; Berlanda Scorza, F.; Rossi, O. GMMA-Based Vaccines: The Known and The Unknown. Front. Immunol. 2021, 12, 715393. [Google Scholar] [CrossRef]
- Łojewska, E.; Sakowicz, T. An Alternative to Antibiotics: Selected Methods to Combat Zoonotic Foodborne Bacterial Infections. Curr. Microbiol. 2021, 78, 4037–4049. [Google Scholar] [CrossRef] [PubMed]
- Issenhuth-Jeanjean, S.; Roggentin, P.; Mikoleit, M.; Guibourdenche, M.; De Pinna, E.; Nair, S.; Fields, P.I.; Weill, F.-X. Supplement 2008–2010 (no. 48) to the white–Kauffmann–Le minor scheme. Res. Microbiol. 2014, 165, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.A.; Eade, C.R.; Wiedmann, M. Embracing Diversity: Differences in Virulence Mechanisms, Disease Severity, and Host Adaptations Contribute to the Success of Nontyphoidal Salmonella as a Foodborne Pathogen. Front. Microbiol. 2019, 10, 1368. [Google Scholar] [CrossRef] [PubMed]
- Townsend, S.M.; Kramer, N.E.; Edwards, R.; Baker, S.; Hamlin, N.; Simmonds, M.; Stevens, K.; Maloy, S.; Parkhill, J.; Dougan, G. Salmonella enterica serovar Typhi possesses a unique repertoire of fimbrial gene sequences. Infect. Immun. 2001, 69, 2894–2901. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.G.; Rosario, D.K.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.; Conte-Junior, C.A. Worldwide epidemiology of Salmonella serovars in animal-based foods: A meta-analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef]
- Dróżdż, M.; Małaszczuk, M.; Paluch, E.; Pawlak, A. Zoonotic potential and prevalence of Salmonella serovars isolated from pets. Infect. Ecol. Epidemiol. 2021, 11, 1975530. [Google Scholar] [CrossRef]
- Rabsch, W.; Tschäpe, H.; Bäumler, A.J. Non-typhoidal salmonellosis: Emerging problems. Microbes Infect. 2001, 3, 237–247. [Google Scholar] [CrossRef]
- Branchu, P.; Bawn, M.; Kingsley, R.A. Genome variation and molecular epidemiology of Salmonella enterica serovar Typhimurium pathovariants. Infect. Immun. 2018, 86, e00079-18. [Google Scholar] [CrossRef]
- Saraf, S.; Jafra, B.S.; Ray, P.; Rawat, A.; Verma, S. Multidrug-Resistant Nontyphoidal Salmonella Associated with Invasive Disease in an Immunocompetent Child. Indian J. Pediatr. 2021, 88, 1266. [Google Scholar] [CrossRef] [PubMed]
- Porwollik, S.; Santiviago, C.; Cheng, P.; Florea, L.; McClelland, M. Differences in gene content between Salmonella enterica serovar Enteritidis isolates and comparison to closely related serovars Gallinarum and Dublin. J. Bacteriol. 2005, 187, 6545–6555. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rabsch, W.; Andrews, H.L.; Kingsley, R.A.; Prager, R.; Tschäpe, H.; Adams, L.G.; Bäumler, A.J. Salmonella enterica serotype Typhimurium and its host-adapted variants. Infect. Immun. 2002, 70, 2249–2255. [Google Scholar] [CrossRef]
- Evangelopoulou, G.; Kritas, S.; Govaris, A.; Burriel, A.R. Animal salmonelloses: A brief review of? host adaptation and host specificity? of Salmonella spp. Vet. World 2013, 6, 703. [Google Scholar] [CrossRef]
- Kingsley, R.A.; Bäumler, A.J. Host adaptation and the emergence of infectious disease: The Salmonella paradigm. Mol. Microbiol. 2000, 36, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Langridge, G.C.; Fookes, M.; Connor, T.R.; Feltwell, T.; Feasey, N.; Parsons, B.N.; Seth-Smith, H.M.; Barquist, L.; Stedman, A.; Humphrey, T. Patterns of genome evolution that have accompanied host adaptation in Salmonella. Proc. Natl. Acad. Sci. USA 2015, 112, 863–868. [Google Scholar] [CrossRef]
- Hansen-Wester, I.; Stecher, B.r.; Hensel, M. Type III secretion of Salmonella enterica serovar Typhimurium translocated effectors and SseFG. Infect. Immun. 2002, 70, 1403–1409. [Google Scholar] [CrossRef]
- Pucciarelli, M.G.; Garcia-del Portillo, F. Salmonella intracellular lifestyles and their impact on host-to-host transmission. Microb. Transm. 2019, 5, 95–116. [Google Scholar] [CrossRef]
- Gopinath, S.; Carden, S.; Monack, D. Shedding light on Salmonella carriers. Trends Microbiol. 2012, 20, 320–327. [Google Scholar] [CrossRef]
- Zhang, H.; Lou, Z.; Chen, X.; Cui, Y.; Wang, H.; Kou, X.; Ma, C. Effect of simultaneous ultrasonic and microwave assisted hydrodistillation on the yield, composition, antibacterial and antibiofilm activity of essential oils from Citrus medica L. var. sarcodactylis. J. Food Eng. 2019, 244, 126–135. [Google Scholar] [CrossRef]
- Foster, N.; Tang, Y.; Berchieri, A.; Geng, S.; Jiao, X.; Barrow, P. Revisiting persistent Salmonella infection and the carrier state: What do we know? Pathogens 2021, 10, 1299. [Google Scholar] [CrossRef]
- Waldner, L.L.; MacKenzie, K.D.; Köster, W.; White, A.P. From exit to entry: Long-term survival and transmission of Salmonella. Pathogens 2012, 1, 128–155. [Google Scholar] [CrossRef]
- Ahmer, B.M.; Gunn, J.S. Interaction of Salmonella spp. with the intestinal microbiota. Front. Microbiol. 2011, 2, 101. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Escobedo, G.; Gunn, J.S. Gallbladder epithelium as a niche for chronic Salmonella carriage. Infect. Immun. 2013, 81, 2920–2930. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, E.J.E.; Azevedo, R.P.; Lopes, A.T.S.; Rocha, J.M.; Albuquerque, G.R.; Wenceslau, A.A.; Miranda, F.R.; Rodrigues, D.D.P.; Maciel, B.M. Salmonella spp. in Wild Free-Living Birds from Atlantic Forest Fragments in Southern Bahia, Brazil. BioMed Res. Int. 2020, 2020, 7594136. [Google Scholar] [CrossRef]
- Marin, C.; Hernandiz, A.; Lainez, M. Biofilm development capacity of Salmonella strains isolated in poultry risk factors and their resistance against disinfectants. Poult. Sci. 2009, 88, 424–431. [Google Scholar] [CrossRef]
- Morton, V.K.; Kearney, A.; Coleman, S.; Viswanathan, M.; Chau, K.; Orr, A.; Hexemer, A. Outbreaks of Salmonella illness associated with frozen raw breaded chicken products in Canada, 2015–2019. Epidemiol. Infect. 2019, 147, e254. [Google Scholar] [CrossRef]
- Chanamé Pinedo, L.; Mughini-Gras, L.; Franz, E.; Hald, T.; Pires, S.M. Sources and trends of human salmonellosis in Europe, 2015–2019: An analysis of outbreak data. Int. J. Food Microbiol. 2022, 379, 109850. [Google Scholar] [CrossRef]
- Laufer, A.S.; Grass, J.; Holt, K.; Whichard, J.M.; Griffin, P.M.; Gould, L.H. Outbreaks of Salmonella infections attributed to beef —United States, 1973–2011. Epidemiol. Infect. 2015, 143, 2003–2013. [Google Scholar] [CrossRef]
- Gutema, F.D.; Agga, G.E.; Abdi, R.D.; De Zutter, L.; Duchateau, L.; Gabriël, S. Prevalence and Serotype Diversity of Salmonella in Apparently Healthy Cattle: Systematic Review and Meta-Analysis of Published Studies, 2000–2017. Front. Vet. Sci. 2019, 6, 102. [Google Scholar] [CrossRef]
- Zając, M.; Wasyl, D.; Różycki, M.; Bilska-Zając, E.; Fafiński, Z.; Iwaniak, W.; Krajewska, M.; Hoszowski, A.; Konieczna, O.; Fafińska, P.; et al. Free-living snakes as a source and possible vector of Salmonella spp. and parasites. Eur. J. Wildl. Res. 2016, 62, 161–166. [Google Scholar] [CrossRef]
- Finley, R.; Ribble, C.; Aramini, J.; Vandermeer, M.; Popa, M.; Litman, M.; Reid-Smith, R. The risk of salmonellae shedding by dogs fed Salmonella-contaminated commercial raw food diets. Can. Vet. J. 2007, 48, 69–75. [Google Scholar] [PubMed]
- Lefebvre, S.L.; Reid-Smith, R.; Boerlin, P.; Weese, J.S. Evaluation of the risks of shedding Salmonellae and other potential pathogens by therapy dogs fed raw diets in Ontario and Alberta. Zoonoses Public Health 2008, 55, 470–480. [Google Scholar] [CrossRef]
- Younus, M.; Wilkins, M.J.; Davies, H.D.; Rahbar, M.H.; Funk, J.; Nguyen, C.; Siddiqi, A.E.; Cho, S.; Saeed, M. Case-control study of disease determinants for non-typhoidal Salmonella infections among Michigan children. BMC Res. Notes 2010, 3, 105. [Google Scholar] [CrossRef]
- Cilia, G.; Turchi, B.; Fratini, F.; Bilei, S.; Bossù, T.; De Marchis, M.L.; Cerri, D.; Pacini, M.I.; Bertelloni, F. Prevalence, Virulence and Antimicrobial Susceptibility of Salmonella spp., Yersinia enterocolitica and Listeria monocytogenes in European Wild Boar (Sus scrofa) Hunted in Tuscany (Central Italy). Pathogens 2021, 10, 93. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Gonzalez, N.; Ugarte-Ruiz, M.; Domínguez, L.; Ruiz-Fons, F. A European Perspective on the Transmission of Foodborne Pathogens at the Wildlife–Livestock–Human Interface. In Food Safety Risks from Wildlife: Challenges in Agriculture, Conservation, and Public Health; Jay-Russell, M., Doyle, M.P., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 59–88. [Google Scholar]
- Gil Molino, M.; Garcia Sanchez, A.; Risco Perez, D.; Goncalves Blanco, P.; Quesada Molina, A.; Rey Pérez, J.; Martín Cano, F.E.; Cerrato Horrillo, R.; Hermoso-de-Mendoza Salcedo, J.; Fernandez Llario, P. Prevalence of Salmonella spp. in tonsils, mandibular lymph nodes and faeces of wild boar from Spain and genetic relationship between isolates. Transbound. Emerg. Dis. 2019, 66, 1218–1226. [Google Scholar] [CrossRef] [PubMed]
- Cummings, K.J.; Rodriguez-Rivera, L.D.; Grigar, M.K.; Rankin, S.C.; Mesenbrink, B.T.; Leland, B.R.; Bodenchuk, M.J. Prevalence and Characterization of Salmonella Isolated from Feral Pigs Throughout Texas. Zoonoses Public Health 2016, 63, 436–441. [Google Scholar] [CrossRef]
- Farias, L.F.P.; Oliveira, C.J.B.; Medardus, J.J.; Molla, B.Z.; Wolfe, B.A.; Gebreyes, W.A. Phenotypic and Genotypic Characterization of Salmonella enterica in Captive Wildlife and Exotic Animal Species in Ohio, USA. Zoonoses Public Health 2015, 62, 438–444. [Google Scholar] [CrossRef]
- Olsen, A.R.; Hammack, T.S. Isolation of Salmonella spp. from the housefly, Musca domestica L., and the dump fly, Hydrotaea aenescens (Wiedemann) (Diptera: Muscidae), at caged-layer houses. J. Food Prot. 2000, 63, 958–960. [Google Scholar] [CrossRef]
- Donoso, A.; Paredes, N.; Retamal, P. Detection of Antimicrobial Resistant Salmonella enterica Strains in Larval and Adult Forms of Lesser Mealworm (Alphitobius diaperinus) From Industrial Poultry Farms. Front. Vet. Sci. 2020, 7, 577848. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Chang, Y.C.; Chuang, H.L.; Chiu, C.C.; Yeh, K.S.; Chang, C.C.; Hsuan, S.L.; Lin, W.H.; Chen, T.H. Transmission of Salmonella between swine farms by the housefly (Musca domestica). J. Food Prot. 2011, 74, 1012–1016. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Tao, S.; Hinkle, N.; Harrison, M.; Chen, J. Salmonella, including antibiotic-resistant Salmonella, from flies captured from cattle farms in Georgia, U.S.A. Sci. Total Environ. 2018, 616–617, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Henzler, D.J.; Opitz, H.M. The role of mice in the epizootiology of Salmonella enteritidis infection on chicken layer farms. Avian Dis. 1992, 36, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Lapuz, R.; Tani, H.; Sasai, K.; Shirota, K.; Katoh, H.; Baba, E. The role of roof rats (Rattus rattus) in the spread of Salmonella Enteritidis and S. Infantis contamination in layer farms in eastern Japan. Epidemiol. Infect. 2008, 136, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Davies, R.H.; Wray, C. Mice as carriers of Salmonella enteritidis on persistently infected poultry units. Vet. Rec. 1995, 137, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Meerburg, B.G.; Jacobs-Reitsma, W.F.; Wagenaar, J.A.; Kijlstra, A. Presence of Salmonella and Campylobacter spp. in wild small mammals on organic farms. Appl. Environ. Microbiol. 2006, 72, 960–962. [Google Scholar] [CrossRef]
- Pocock, M.J.; Searle, J.B.; Betts, W.B.; White, P.C. Patterns of infection by Salmonella and Yersinia spp. in commensal house mouse (Mus musculus domesticus) populations. J. Appl. Microbiol. 2001, 90, 755–760. [Google Scholar] [CrossRef]
- Le Moine, V.; Vannier, P.; Jestin, A. Microbiological studies of wild rodents in farms as carriers of pig infectious agents. Prev. Vet. Med. 1987, 4, 399–408. [Google Scholar] [CrossRef]
- Lee, K.-H.; Lee, J.-Y.; Roy, P.K.; Mizan, M.F.R.; Hossain, M.I.; Park, S.H.; Ha, S.-D. Viability of Salmonella Typhimurium biofilms on major food-contact surfaces and eggshell treated during 35 days with and without water storage at room temperature. Poult. Sci. 2020, 99, 4558–4565. [Google Scholar] [CrossRef]
- Elpers, L.; Kretzschmar, J.; Nuccio, S.P.; Bäumler, A.J.; Hensel, M. Factors Required for Adhesion of Salmonella enterica Serovar Typhimurium to Corn Salad (Valerianella locusta). Appl. Environ. Microbiol. 2020, 86, e02757-19. [Google Scholar] [CrossRef]
- Liu, H.; Whitehouse, C.A.; Li, B. Presence and Persistence of Salmonella in Water: The Impact on Microbial Quality of Water and Food Safety. Front. Public Health 2018, 6, 159. [Google Scholar] [CrossRef]
- Strawn, L.K.; Danyluk, M.D.; Worobo, R.W.; Wiedmann, M. Distributions of Salmonella subtypes differ between two U.S. produce-growing regions. Appl. Environ. Microbiol. 2014, 80, 3982–3991. [Google Scholar] [CrossRef]
- Bosilevac, J.M.; Guerini, M.N.; Kalchayanand, N.; Koohmaraie, M. Prevalence and Characterization of Salmonellae in Commercial Ground Beef in the United States. Appl. Environ. Microbiol. 2009, 75, 1892–1900. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef]
- CDC. Salmonella Outbreaks Associated with Not Ready-to-Eat Breaded, Stuffed Chicken Products—United States, 1998–2022. MMWR Morb. Mortal. Wkly. Rep. 2023 2023, 72, 484–487. [Google Scholar] [CrossRef]
- Griffith, R.W.; Carlson, S.A.; Krull, A.C. Salmonellosis. Dis. Swine 2019, 912–925. [Google Scholar] [CrossRef]
- Gantois, I.; Ducatelle, R.; Pasmans, F.; Haesebrouck, F.; Gast, R.; Humphrey, T.J.; Van Immerseel, F. Mechanisms of egg contamination by Salmonella Enteritidis. FEMS Microbiol. Rev. 2009, 33, 718–738. [Google Scholar] [CrossRef]
- Lee, J.-S.; Mogasale, V.V.; Mogasale, V.; Lee, K. Geographical distribution of typhoid risk factors in low and middle income countries. BMC Infect. Dis. 2016, 16, 732. [Google Scholar] [CrossRef]
- CDC. Foodborne Diseases Active Surveillance Network, 2011 Surveillance Report; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2011.
- Center for Disease Control and Prevention (CDC). Incidence and trends of infection with pathogens transmitted commonly through food-foodborne diseases active surveillance network, 10 U.S. sites, 1996–2012. MMWR Morb. Mortal. Wkly. Rep. 2013, 62, 283–287. [Google Scholar]
- Jones, T.F.; Ingram, L.A.; Fullerton, K.E.; Marcus, R.; Anderson, B.J.; McCarthy, P.V.; Vugia, D.; Shiferaw, B.; Haubert, N.; Wedel, S.; et al. A case-control study of the epidemiology of sporadic Salmonella infection in infants. Pediatrics 2006, 118, 2380–2387. [Google Scholar] [CrossRef]
- Bavishi, C.; Dupont, H.L. Systematic review: The use of proton pump inhibitors and increased susceptibility to enteric infection. Aliment. Pharmacol. Ther. 2011, 34, 1269–1281. [Google Scholar] [CrossRef]
- Nollet, N.; Maes, D.; De Zutter, L.; Duchateau, L.; Houf, K.; Huysmans, K.; Imberechts, H.; Geers, R.; de Kruif, A.; Van Hoof, J. Risk factors for the herd-level bacteriologic prevalence of Salmonella in Belgian slaughter pigs. Prev. Vet. Med. 2004, 65, 63–75. [Google Scholar] [CrossRef]
- Nielsen, L.R.; Schukken, Y.H.; Gröhn, Y.T.; Ersbøll, A.K. Salmonella Dublin infection in dairy cattle: Risk factors for becoming a carrier. Prev. Vet. Med. 2004, 65, 47–62. [Google Scholar] [CrossRef]
- Evans, S.; Davies, R. Case control study of multiple-resistant Salmonella typhimurium DT104 infection of cattle in Great Britain. Vet. Rec. 1996, 139, 557–558. [Google Scholar] [CrossRef]
- Kent, E.; Okafor, C.; Caldwell, M.; Walker, T.; Whitlock, B.; Lear, A. Control of Salmonella Dublin in a bovine dairy herd. J. Vet. Intern. Med. 2021, 35, 2075–2080. [Google Scholar] [CrossRef]
- Ethèves, M.A.; Choisis, N.; Alvarez, S.; Dalleau, F.; Hascoat, J.; Gallard, V.; Cardinale, E. Risk factors for Salmonella enterica subsp. enterica persistence in broiler-chicken flocks on Reunion Island. Heliyon 2021, 7, e06278. [Google Scholar] [CrossRef]
- Crump, J.A.; Braden, C.R.; Dey, M.E.; Hoekstra, R.M.; Rickelman-Apisa, J.M.; Baldwin, D.A.; De Fijter, S.J.; Nowicki, S.F.; Koch, E.M.; Bannerman, T.L.; et al. Outbreaks of Escherichia coli O157 infections at multiple county agricultural fairs: A hazard of mixing cattle, concession stands and children. Epidemiol. Infect. 2003, 131, 1055–1062. [Google Scholar] [CrossRef]
- Cardoso, M.J.; Nicolau, A.I.; Borda, D.; Nielsen, L.; Maia, R.L.; Møretrø, T.; Ferreira, V.; Knøchel, S.; Langsrud, S.; Teixeira, P. Salmonella in eggs: From shopping to consumption-A review providing an evidence-based analysis of risk factors. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2716–2741. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, J.; Ed-Dra, A.; Li, X.; Peng, X.; Xia, L.; Guo, Q.; Yao, G.; Yue, M. Prevalence and genomic investigation of Salmonella isolates recovered from animal food-chain in Xinjiang, China. Food Res. Int. 2021, 142, 110198. [Google Scholar] [CrossRef]
- Reddy, E.A.; Shaw, A.V.; Crump, J.A. Community-acquired bloodstream infections in Africa: A systematic review and meta-analysis. Lancet Infect. Dis. 2010, 10, 417–432. [Google Scholar] [CrossRef]
- Thakur, R.; Suri, C.R.; Rishi, P. Contribution of typhoid toxin in the pathogenesis of Salmonella Typhi. Microb. Pathog. 2022, 164, 105444. [Google Scholar] [CrossRef]
- Patel, T.A.; Armstrong, M.; Morris-Jones, S.D.; Wright, S.G.; Doherty, T. Imported enteric fever: Case series from the hospital for tropical diseases, London, United Kingdom. Am. J. Trop. Med. Hyg. 2010, 82, 1121–1126. [Google Scholar] [CrossRef]
- Kuvandik, C.; Karaoglan, I.; Namiduru, M.; Baydar, I. Predictive value of clinical and laboratory findings in the diagnosis of the enteric fever. New Microbiol. 2009, 32, 25–30. [Google Scholar]
- Galán, J.E. Typhoid toxin provides a window into typhoid fever and the biology of Salmonella Typhi. Proc. Natl. Acad. Sci. USA 2016, 113, 6338–6344. [Google Scholar] [CrossRef]
- Crump, J.A. Progress in Typhoid Fever Epidemiology. Clin. Infect. Dis. 2019, 68, S4–S9. [Google Scholar] [CrossRef]
- Parry, C.M.; Hien, T.T.; Dougan, G.; White, N.J.; Farrar, J.J. Typhoid fever. N. Engl. J. Med. 2002, 347, 1770–1782. [Google Scholar] [CrossRef]
- Allen, J.C.; Toapanta, F.R.; Baliban, S.M.; Sztein, M.B.; Tennant, S.M. Reduced immunogenicity of a live Salmonella enterica serovar Typhimurium vaccine in aged mice. Front. Immunol. 2023, 14, 1190339. [Google Scholar] [CrossRef]
- CDC. National Enteric Disease Surveillance: Salmonella Annual Report. Available online: https://www.cdc.gov/nationalsurveillance/salmonella-surveillance.html (accessed on 12 December 2023).
- Bakhshandeh, B.; Sorboni, S.G.; Haghighi, D.M.; Ahmadi, F.; Dehghani, Z.; Badiei, A. New analytical methods using carbon-based nanomaterials for detection of Salmonella species as a major food poisoning organism in water and soil resources. Chemosphere 2022, 287, 132243. [Google Scholar] [CrossRef]
- Turgeon, P.; Ng, V.; Murray, R.; Nesbitt, A. Forecasting the incidence of salmonellosis in seniors in Canada: A trend analysis and the potential impact of the demographic shift. PLoS ONE 2018, 13, e0208124. [Google Scholar] [CrossRef]
- Pulford, C.V.; Perez-Sepulveda, B.M.; Canals, R.; Bevington, J.A.; Bengtsson, R.J.; Wenner, N.; Rodwell, E.V.; Kumwenda, B.; Zhu, X.; Bennett, R.J.; et al. Stepwise evolution of Salmonella Typhimurium ST313 causing bloodstream infection in Africa. Nat. Microbiol. 2021, 6, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Acheson, D.; Hohmann, E.L. Nontyphoidal Salmonellosis. Clin. Infect. Dis. 2001, 32, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Ajene, A.N.; Fischer Walker, C.L.; Black, R.E. Enteric pathogens and reactive arthritis: A systematic review of Campylobacter, salmonella and Shigella-associated reactive arthritis. J. Health Popul. Nutr. 2013, 31, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Ehuwa, O.; Jaiswal, A.K.; Jaiswal, S. Salmonella, Food Safety and Food Handling Practices. Foods 2021, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- Schempp, C.M.; Schauer, F.; Huhn, C.K.; Venhoff, N.; Finzel, S. Skin inflammation associated with arthritis, synovitis and enthesitis. Part 2: Rheumatoid arthritis, reactive arthritis, Reiter’s syndrome, Lyme borreliosis, dermatomyositis and lupus erythematosus. J. Dtsch. Dermatol. Ges. 2019, 17, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, Y.; Loza, E.; Villa-Gomez, G.; Trujillo, C.C.; Baez, S.; Asai, T.; Ikoma, T.; Endoh, K.; Nakamura, K. Metagenomics of microbial communities in gallbladder bile from patients with gallbladder cancer or cholelithiasis. Asian Pac. J. Cancer Prev. 2018, 19, 961. [Google Scholar] [PubMed]
- Zha, L.; Garrett, S.; Sun, J. Salmonella infection in chronic inflammation and gastrointestinal cancer. Diseases 2019, 7, 28. [Google Scholar] [CrossRef]
- DuPont, H.L. Bacterial diarrhea. N. Engl. J. Med. 2009, 361, 1560–1569. [Google Scholar] [CrossRef]
- Serpil Kahya, D. Salmonellosis in Animals. In Salmonella—A Re-emerging Pathogen; Maria Teresa, M., Ed.; IntechOpen: Rijeka, Croatia, 2017; pp. 19–37. [Google Scholar] [CrossRef]
- Barrow, P.A.; Jones, M.A.; Smith, A.L.; Wigley, P. The long view: Salmonella—The last forty years. Avian Pathol. 2012, 41, 413–420. [Google Scholar] [CrossRef]
- Zhou, X.; Kang, X.; Zhou, K.; Yue, M. A global dataset for prevalence of Salmonella Gallinarum between 1945 and 2021. Sci. Data 2022, 9, 495. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Zhang, A.; Gu, J.; Zhao, R.; Pan, X.; Dai, Y.; Yin, L.; Zhang, Q.; Hu, X.; Wang, H.; et al. Evaluating Salmonella pullorum dissemination and shedding patterns and antibody production in infected chickens. BMC Vet. Res. 2022, 18, 240. [Google Scholar] [CrossRef] [PubMed]
- Uzzau, S.; Leori, G.S.; Petruzzi, V.; Watson, P.R.; Schianchi, G.; Bacciu, D.; Mazzarello, V.; Wallis, T.S.; Rubino, S. Salmonella enterica serovar-host specificity does not correlate with the magnitude of intestinal invasion in sheep. Infect. Immun. 2001, 69, 3092–3099. [Google Scholar] [CrossRef]
- Habrun, B.; Listes, E.; Spicic, S.; Cvetnic, Z.; Lukacevic, D.; Jemersic, L.; Lojkic, M.; Kompes, G. An outbreak of Salmonella Abortusovis abortions in sheep in south Croatia. J. Vet. Med. B Infect. Dis. Vet. Public Health 2006, 53, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Vanselow, B.A.; Hum, S.; Hornitzky, M.A.; Eamens, G.J.; Quinn, K. Salmonella Typhimurium persistence in a Hunter Valley dairy herd. Aust. Vet. J. 2007, 85, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Askari, N.; Mashhad Rafiee, S.; Amini, K. A case control study of Salmonella SPP. infection in stray dogs in Tehran shelters and the correlation between paraclinical tests results and clinical findings. Arch. Razi Inst. 2020, 75, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Feary, D.J.; Hassel, D.M. Enteritis and colitis in horses. Vet. Clin. North Am. Equine Pract. 2006, 22, 437–479. [Google Scholar] [CrossRef]
- Burgess, B.A.; Noyes, N.R.; Bolte, D.S.; Hyatt, D.R.; van Metre, D.C.; Morley, P.S. Rapid Salmonella detection in experimentally inoculated equine faecal and veterinary hospital environmental samples using commercially available lateral flow immunoassays. Equine Vet. J. 2015, 47, 119–122. [Google Scholar] [CrossRef]
- CDC. Vital signs: Incidence and trends of infection with pathogens transmitted commonly through food—Foodborne diseases active surveillance network, 10 U.S. sites, 1996–2010. MMWR Morb. Mortal. Wkly. Rep. 2011, 60, 749–755. [Google Scholar]
- Jackson, B.R.; Griffin, P.M.; Cole, D.; Walsh, K.A.; Chai, S.J. Outbreak-associated Salmonella enterica serotypes and food Commodities, United States, 1998–2008. Emerg. Infect. Dis. 2013, 19, 1239–1244. [Google Scholar] [CrossRef]
- Soto-Dávila, M.; Hossain, A.; Chakraborty, S.; Rise, M.L.; Santander, J. Aeromonas salmonicida subsp. salmonicida Early Infection and Immune Response of Atlantic Cod (Gadus morhua L.) Primary Macrophages. Front. Immunol. 2019, 10, 1237. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Food Additives and Flavourings (FAF); Younes, M.; Aquilina, G.; Engel, K.-H.; Fowler, P.; Frutos Fernandez, M.J.; Fürst, P.; Gürtler, R.; Gundert-Remy, U.; Husøy, T.; et al. Safety of use of Monk fruit extract as a food additive in different food categories. EFSA J. 2019, 17, e05921. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.K.; Murray, R.; Flockhart, L.; Pintar, K.; Pollari, F.; Fazil, A.; Nesbitt, A.; Marshall, B. Estimates of the burden of foodborne illness in Canada for 30 specified pathogens and unspecified agents, circa 2006. Foodborne Pathog. Dis. 2013, 10, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Government of Canada. National Enteric Surveillance Program (NESP) Annual Summary 2013; Government of Canada: Ottawa, ON, Canada, 2015.
- Hendriksen, R.S.; Vieira, A.R.; Karlsmose, S.; Lo Fo Wong, D.M.; Jensen, A.B.; Wegener, H.C.; Aarestrup, F.M. Global monitoring of Salmonella serovar distribution from the World Health Organization Global Foodborne Infections Network Country Data Bank: Results of quality assured laboratories from 2001 to 2007. Foodborne Pathog. Dis. 2011, 8, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Buuck, S.; Noveroske, D. Multistate outbreak of Salmonella infections linked to raw turkey products—United States, 2017–2019. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 1045–10493. [Google Scholar] [CrossRef] [PubMed]
- EFSA. The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2017. Available online: https://efsa.onlinelibrary.wiley.com/doi/10.2903/j.efsa.2018.5500 (accessed on 10 July 2023).
- European Centre for Disease, P.; Control, E.F.S.A. Multi-country outbreak of monophasic Salmonella Typhimurium sequence type 34 linked to chocolate products–first update–18 May 2022. EFSA Support. Publ. 2022, 19, 7352E. [Google Scholar] [CrossRef]
- Cevallos-Cevallos, J.M.; Gu, G.; Danyluk, M.D.; van Bruggen, A.H. Adhesion and splash dispersal of Salmonella enterica Typhimurium on tomato leaflets: Effects of rdar morphotype and trichome density. Int. J. Food Microbiol. 2012, 160, 58–64. [Google Scholar] [CrossRef]
- Ibarra, J.A.; Steele-Mortimer, O. Salmonella—The ultimate insider. Salmonella virulence factors that modulate intracellular survival. Cell. Microbiol. 2009, 11, 1579–1586. [Google Scholar] [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition in the innate immune response. Biochem. J. 2009, 420, 1–16. [Google Scholar] [CrossRef]
- Kage, H.; Takaya, A.; Ohya, M.; Yamamoto, T. Coordinated regulation of expression of Salmonella pathogenicity island 1 and flagellar type III secretion systems by ATP-dependent ClpXP protease. J. Bacteriol. 2008, 190, 2470–2478. [Google Scholar] [CrossRef]
- Lang, T.; Mansell, A. The negative regulation of Toll-like receptor and associated pathways. Immunol. Cell Biol. 2007, 85, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Everest, P.; Servos, S.; Foxwell, N.; Zähringer, U.; Brade, H.; Rietschel, E.T.; Dougan, G.; Charles, I.G.; Maskell, D.J. A lethal role for lipid A in Salmonella infections. Mol. Microbiol. 1998, 29, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Raffatellu, M.; Chessa, D.; Wilson, R.P.; Dusold, R.; Rubino, S.; Bäumler, A.J. The Vi capsular antigen of Salmonella enterica serotype Typhi reduces Toll-like receptor-dependent interleukin-8 expression in the intestinal mucosa. Infect. Immun. 2005, 73, 3367–3374. [Google Scholar] [CrossRef] [PubMed]
- Bignold, L.P.; Rogers, S.D.; Siaw, T.M.; Bahnisch, J. Inhibition of chemotaxis of neutrophil leukocytes to interleukin-8 by endotoxins of various bacteria. Infect. Immun. 1991, 59, 4255–4258. [Google Scholar] [CrossRef] [PubMed]
- Jessen, D.L.; Osei-Owusu, P.; Toosky, M.; Roughead, W.; Bradley, D.S.; Nilles, M.L. Type III secretion needle proteins induce cell signaling and cytokine secretion via Toll-like receptors. Infect. Immun. 2014, 82, 2300–2309. [Google Scholar] [CrossRef] [PubMed]
- Mambu, J.; Virlogeux-Payant, I.; Holbert, S.; Grépinet, O.; Velge, P.; Wiedemann, A. An Updated View on the Rck Invasin of Salmonella: Still Much to Discover. Front. Cell. Infect. Microbiol. 2017, 7, 500. [Google Scholar] [CrossRef]
- Mellor, K.C.; Blackwell, G.A.; Cawthraw, S.A.; Mensah, N.E.; Reid, S.W.J.; Thomson, N.R.; Petrovska, L.; Mather, A.E. Contrasting long-term dynamics of antimicrobial resistance and virulence plasmids in Salmonella typhimurium from animals. Microb. Genom. 2022, 8, 000826. [Google Scholar] [CrossRef]
- Zuo, L.; Zhou, L.; Wu, C.; Wang, Y.; Li, Y.; Huang, R.; Wu, S. Salmonella spvC Gene Inhibits Pyroptosis and Intestinal Inflammation to Aggravate Systemic Infection in Mice. Front. Microbiol. 2020, 11, 562491. [Google Scholar] [CrossRef]
- Rodríguez-Beltrán, J.; DelaFuente, J.; León-Sampedro, R.; MacLean, R.C.; San Millán, Á. Beyond horizontal gene transfer: The role of plasmids in bacterial evolution. Nat. Rev. Microbiol. 2021, 19, 347–359. [Google Scholar] [CrossRef]
- Sengupta, M.; Austin, S. Prevalence and significance of plasmid maintenance functions in the virulence plasmids of pathogenic bacteria. Infect. Immun. 2011, 79, 2502–2509. [Google Scholar] [CrossRef]
- Kosarewicz, A.; Königsmaier, L.; Marlovits, T.C. The blueprint of the type-3 injectisome. Philos. Trans. R. Soc. Lond B Biol. Sci. 2012, 367, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, C.V.; Mercado-Lubo, R.; Hallstrom, K.; McCormick, B.A. Salmonella effector proteins and host-cell responses. Cell. Mol. Life Sci. 2011, 68, 3687–3697. [Google Scholar] [CrossRef] [PubMed]
- LaRock, D.L.; Chaudhary, A.; Miller, S.I. Salmonellae interactions with host processes. Nat. Rev. Microbiol. 2015, 13, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Figueira, R.; Holden, D.W. Functions of the Salmonella pathogenicity island 2 (SPI-2) type III secretion system effectors. Microbiology 2012, 158, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Sellin, M.E.; Müller, A.A.; Felmy, B.; Dolowschiak, T.; Diard, M.; Tardivel, A.; Maslowski, K.M.; Hardt, W.D. Epithelium-intrinsic NAIP/NLRC4 inflammasome drives infected enterocyte expulsion to restrict Salmonella replication in the intestinal mucosa. Cell Host Microbe 2014, 16, 237–248. [Google Scholar] [CrossRef] [PubMed]
- McGhie, E.J.; Brawn, L.C.; Hume, P.J.; Humphreys, D.; Koronakis, V. Salmonella takes control: Effector-driven manipulation of the host. Curr. Opin. Microbiol. 2009, 12, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Lenders, M.H.H.; Weidtkamp-Peters, S.; Kleinschrodt, D.; Jaeger, K.-E.; Smits, S.H.J.; Schmitt, L. Directionality of substrate translocation of the hemolysin A Type I secretion system. Sci. Rep. 2015, 5, 12470. [Google Scholar] [CrossRef]
- Li, X.; Bleumink-Pluym, N.M.C.; Luijkx, Y.M.C.A.; Wubbolts, R.W.; van Putten, J.P.M.; Strijbis, K. MUC1 is a receptor for the Salmonella SiiE adhesin that enables apical invasion into enterocytes. PLOS Pathog. 2019, 15, e1007566. [Google Scholar] [CrossRef]
- Main-Hester, K.L.; Colpitts, K.M.; Thomas, G.A.; Fang, F.C.; Libby, S.J. Coordinate regulation of Salmonella pathogenicity island 1 (SPI1) and SPI4 in Salmonella enterica serovar Typhimurium. Infect. Immun. 2008, 76, 1024–1035. [Google Scholar] [CrossRef]
- Fang, L.; Shen, H.; Tang, Y.; Fang, W. Superoxide dismutase of Streptococcus suis serotype 2 plays a role in anti-autophagic response by scavenging reactive oxygen species in infected macrophages. Vet. Microbiol. 2015, 176, 328–336. [Google Scholar] [CrossRef]
- Felmy, B.; Songhet, P.; Slack, E.M.; Müller, A.J.; Kremer, M.; Van Maele, L.; Cayet, D.; Heikenwalder, M.; Sirard, J.C.; Hardt, W.D. NADPH oxidase deficient mice develop colitis and bacteremia upon infection with normally avirulent, TTSS-1- and TTSS-2-deficient Salmonella typhimurium. PLoS ONE 2013, 8, e77204. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Elbediwi, M.; Nambiar, R.B.; Yang, H.; Lin, J.; Yue, M. Genomic Characterization of Antimicrobial-Resistant Salmonella enterica in Duck, Chicken, and Pig Farms and Retail Markets in Eastern China. Microbiol. Spectr. 2022, 10, e0125722. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Deng, L.; Huang, F.; Wang, Z.; Lu, Q.; Xu, C. Flagellar Motility Is Critical for Salmonella enterica Serovar Typhimurium Biofilm Development. Front. Microbiol. 2020, 11, 1695. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, R.; Kim, B.; Mollo, E.A.; Imlay, J.A.; Slauch, J.M. Structural properties of periplasmic SodCI that correlate with virulence in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2007, 189, 4343–4352. [Google Scholar] [CrossRef] [PubMed]
- Rehman, T.; Yin, L.; Latif, M.B.; Chen, J.; Wang, K.; Geng, Y.; Huang, X.; Abaidullah, M.; Guo, H.; Ouyang, P. Adhesive mechanism of different Salmonella fimbrial adhesins. Microb. Pathog. 2019, 137, 103748. [Google Scholar] [CrossRef]
- Kolenda, R.; Ugorski, M.; Grzymajlo, K. Everything You Always Wanted to Know About Salmonella Type 1 Fimbriae, but Were Afraid to Ask. Front. Microbiol. 2019, 10, 1017. [Google Scholar] [CrossRef] [PubMed]
- Grzymajło, K.; Ugorski, M.; Kolenda, R.; Kędzierska, A.; Kuźmińska-Bajor, M.; Wieliczko, A. FimH adhesin from host unrestricted Salmonella Enteritidis binds to different glycoprotein ligands expressed by enterocytes from sheep, pig and cattle than FimH adhesins from host restricted Salmonella Abortus-ovis, Salmonella Choleraesuis and Salmonella Dublin. Vet. Microbiol. 2013, 166, 550–557. [Google Scholar] [CrossRef]
- Uchiya, K.I.; Kamimura, Y.; Jusakon, A.; Nikai, T. Salmonella Fimbrial Protein FimH Is Involved in Expression of Proinflammatory Cytokines in a Toll-Like Receptor 4-Dependent Manner. Infect. Immun. 2019, 87, e00881-18. [Google Scholar] [CrossRef]
- Sano, G.; Takada, Y.; Goto, S.; Maruyama, K.; Shindo, Y.; Oka, K.; Matsui, H.; Matsuo, K. Flagella facilitate escape of Salmonella from oncotic macrophages. J. Bacteriol. 2007, 189, 8224–8232. [Google Scholar] [CrossRef]
- Wang, G.; Song, Q.; Huang, S.; Wang, Y.; Cai, S.; Yu, H.; Ding, X.; Zeng, X.; Zhang, J. Effect of antimicrobial peptide microcin J25 on growth performance, immune regulation, and intestinal microbiota in broiler chickens challenged with Escherichia coli and Salmonella. Animals 2020, 10, 345. [Google Scholar] [CrossRef]
- Sokaribo, A.S.; Hansen, E.G.; McCarthy, M.; Desin, T.S.; Waldner, L.L.; MacKenzie, K.D.; Mutwiri, G., Jr.; Herman, N.J.; Herman, D.J.; Wang, Y.; et al. Metabolic Activation of CsgD in the Regulation of Salmonella Biofilms. Microorganisms 2020, 8, 964. [Google Scholar] [CrossRef] [PubMed]
- Haiko, J.; Westerlund-Wikström, B. The role of the bacterial flagellum in adhesion and virulence. Biology 2013, 2, 1242–1267. [Google Scholar] [CrossRef] [PubMed]
- Horstmann, J.A.; Zschieschang, E.; Truschel, T.; de Diego, J.; Lunelli, M.; Rohde, M.; May, T.; Strowig, T.; Stradal, T.; Kolbe, M.; et al. Flagellin phase-dependent swimming on epithelial cell surfaces contributes to productive Salmonella gut colonisation. Cell. Microbiol. 2017, 19, e12739. [Google Scholar] [CrossRef]
- Liston, S.D.; Ovchinnikova, O.G.; Whitfield, C. Unique lipid anchor attaches Vi antigen capsule to the surface of Salmonella enterica serovar Typhi. Proc. Natl. Acad. Sci. USA 2016, 113, 6719–6724. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.J.; O’Shaughnessy, C.M.; Siggins, M.K.; Bobat, S.; Kingsley, R.A.; Goulding, D.A.; Crump, J.A.; Reyburn, H.; Micoli, F.; Dougan, G.; et al. Differential Killing of Salmonella enterica Serovar Typhi by Antibodies Targeting Vi and Lipopolysaccharide O:9 Antigen. PLoS ONE 2016, 11, e0145945. [Google Scholar] [CrossRef] [PubMed]
- Hiyoshi, H.; Tiffany, C.R.; Bronner, D.N.; Bäumler, A.J. Typhoidal Salmonella serovars: Ecological opportunity and the evolution of a new pathovar. FEMS Microbiol. Rev. 2018, 42, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Chong, A.; Lee, S.; Yang, Y.A.; Song, J. The Role of Typhoid Toxin in Salmonella Typhi Virulence. Yale J. Biol. Med. 2017, 90, 283–290. [Google Scholar]
- Needham, B.D.; Trent, M.S. Fortifying the barrier: The impact of lipid A remodelling on bacterial pathogenesis. Nat. Rev. Microbiol. 2013, 11, 467–481. [Google Scholar] [CrossRef]
- Richards, S.M.; Strandberg, K.L.; Gunn, J.S. Salmonella-regulated lipopolysaccharide modifications. Subcell. Biochem. 2010, 53, 101–122. [Google Scholar] [CrossRef]
- Kong, Q.; Yang, J.; Liu, Q.; Alamuri, P.; Roland, K.L.; Curtiss, R., 3rd. Effect of deletion of genes involved in lipopolysaccharide core and O-antigen synthesis on virulence and immunogenicity of Salmonella enterica serovar typhimurium. Infect. Immun. 2011, 79, 4227–4239. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Tsen, H.-Y.; Lin, C.-L.; Yu, B.; Chen, C.-S. Oral administration of a combination of select lactic acid bacteria strains to reduce the Salmonella invasion and inflammation of broiler chicks. Poult. Sci. 2012, 91, 2139–2147. [Google Scholar] [CrossRef] [PubMed]
- Römling, U. Characterization of the rdar morphotype, a multicellular behaviour in Enterobacteriaceae. Cell. Mol. Life Sci. 2005, 62, 1234–1246. [Google Scholar] [CrossRef] [PubMed]
- Collinson, S.K.; Clouthier, S.C.; Doran, J.L.; Banser, P.A.; Kay, W.W. Salmonella enteritidis agfBAC operon encoding thin, aggregative fimbriae. J. Bacteriol. 1996, 178, 662–667. [Google Scholar] [CrossRef]
- Zakikhany, K.; Harrington, C.R.; Nimtz, M.; Hinton, J.C.; Römling, U. Unphosphorylated CsgD controls biofilm formation in Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2010, 77, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Kader, A.; Simm, R.; Gerstel, U.; Morr, M.; Römling, U. Hierarchical involvement of various GGDEF domain proteins in rdar morphotype development of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2006, 60, 602–616. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; Russell, J.B.; Flythe, M.D.; Gantois, I.; Timbermont, L.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R. The use of organic acids to combat Salmonella in poultry: A mechanistic explanation of the efficacy. Avian Pathol. 2006, 35, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Galipó, E.; Zoche-Golob, V.; Sassu, E.L.; Prigge, C.; Sjölund, M.; Tobias, T.; Rzeżutka, A.; Smith, R.P.; Burow, E. Prioritization of pig farm biosecurity for control of Salmonella and hepatitis E virus infections: Results of a European expert opinion elicitation. Porc. Health Manag. 2023, 9, 8. [Google Scholar] [CrossRef]
- Madec, F. Good Practices for Biosecurity in the Pig Sector: Issues and Options in Developing and Transition Countries. Available online: https://www.fao.org/3/i1435e/i1435e00.htm (accessed on 15 June 2023).
- Sharma, B. Poultry Production, Management and Bio-Security Measures. J. Agric. Environ. 2010, 11, 120–125. [Google Scholar] [CrossRef]
- Jensen, A.N.; Dalsgaard, A.; Stockmarr, A.; Nielsen, E.M.; Baggesen, D.L. Survival and transmission of Salmonella enterica serovar typhimurium in an outdoor organic pig farming environment. Appl. Environ. Microbiol. 2006, 72, 1833–1842. [Google Scholar] [CrossRef]
- Mannion, C.; Leonard, F.C.; Lynch, P.B.; Egan, J. Efficacy of cleaning and disinfection on pig farms in Ireland. Vet. Rec. 2007, 161, 371–375. [Google Scholar] [CrossRef]
- Trampel, D.W.; Holder, T.G.; Gast, R.K. Integrated farm management to prevent Salmonella Enteritidis contamination of eggs. J. Appl. Poult. Res. 2014, 23, 353–365. [Google Scholar] [CrossRef]
- Lestari, V.S.; Sirajuddin, S.N.; Kasim, K. Adoption of biosecurity measures by layer smallholders. J. Indones. Trop. Anim. Agric. 2011, 36, 297–302. [Google Scholar] [CrossRef]
- Mee, J.F.; Geraghty, T.; O’Neill, R.; More, S.J. Bioexclusion of diseases from dairy and beef farms: Risks of introducing infectious agents and risk reduction strategies. Vet. J. 2012, 194, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Renault, V.; Damiaans, B.; Sarrazin, S.; Humblet, M.F.; Dewulf, J.; Saegerman, C. Biosecurity practices in Belgian cattle farming: Level of implementation, constraints and weaknesses. Transbound. Emerg. Dis. 2018, 65, 1246–1261. [Google Scholar] [CrossRef] [PubMed]
- Tilli, G.; Laconi, A.; Galuppo, F.; Mughini-Gras, L.; Piccirillo, A. Assessing Biosecurity Compliance in Poultry Farms: A Survey in a Densely Populated Poultry Area in North East Italy. Animals 2022, 12, 1409. [Google Scholar] [CrossRef] [PubMed]
- Busani, L.; Dalla Pozza, M.; Bonfanti, L.; Toson, M.; Ferrè, N.; Marangon, S. Intervention strategies for low-pathogenic avian influenza control in Italy. Avian Dis. 2007, 51, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Gelaude, P.; Schlepers, M.; Verlinden, M.; Laanen, M.; Dewulf, J. Biocheck.UGent: A quantitative tool to measure biosecurity at broiler farms and the relationship with technical performances and antimicrobial use. Poult. Sci. 2014, 93, 2740–2751. [Google Scholar] [CrossRef] [PubMed]
- Van Limbergen, T.; Dewulf, J.; Klinkenberg, M.; Ducatelle, R.; Gelaude, P.; Méndez, J.; Heinola, K.; Papasolomontos, S.; Szeleszczuk, P.; Maes, D. Scoring biosecurity in European conventional broiler production. Poult. Sci. 2018, 97, 74–83. [Google Scholar] [CrossRef]
- Maunsell, F.; Donovan, G.A. Biosecurity and risk management for dairy replacements. Vet. Clin. North Am. Food Anim. Pract. 2008, 24, 155–190. [Google Scholar] [CrossRef]
- Monitoring, N.A.H. Dairy 2002:Animal Disease Exclusion Practices on U.S. Dairy Operations; USDA: Fort Collins, CO, USA, 2004.
- Verwoerd, D.W. Definition of a vector and a vector-borne disease. Rev. Sci. Tech. 2015, 34, 29–39. [Google Scholar] [CrossRef]
- Spalding, M.G. Diseases of Poultry, 12th ed.; Wildlife disease Association: Lawrence, KS, USA, 2009. [Google Scholar]
- Meerburg, B.G.; Kijlstra, A. Role of rodents in transmission of Salmonella and Campylobacter. J. Sci. Food Agric. 2007, 87, 2774–2781. [Google Scholar] [CrossRef]
- Rebeca, Z.-S.; Andrea Molina, A. Preharvest Salmonella Risk Contamination and the Control Strategies. In Current Topics in Salmonella and Salmonellosis; Mihai, M., Ed.; IntechOpen: Rijeka, Croatia, 2017; pp. 193–213. [Google Scholar] [CrossRef]
- Axtell, R.C. Fly Management in Poultry Production: Cultural, Biological, and Chemical1. Poult. Sci. 1986, 65, 657–667. [Google Scholar] [CrossRef]
- Balaraman, V.; Drolet, B.S.; Mitzel, D.N.; Wilson, W.C.; Owens, J.; Gaudreault, N.N.; Meekins, D.A.; Bold, D.; Trujillo, J.D.; Noronha, L.E.; et al. Mechanical transmission of SARS-CoV-2 by house flies. Parasites Vectors 2021, 14, 214. [Google Scholar] [CrossRef]
- Azizi-Lalabadi, M.; Rahimzadeh-Sani, Z.; Feng, J.; Hosseini, H.; Jafari, S.M. The impact of essential oils on the qualitative properties, release profile, and stimuli-responsiveness of active food packaging nanocomposites. Crit. Rev. Food Sci. Nutr. 2023, 63, 1822–1845. [Google Scholar] [CrossRef]
- Ong, S.Q.; Ab Majid, A.H.; Ahmad, H. Insecticide Residues on Poultry Manures: Field Efficacy Test on Selected Insecticides in Managing Musca Domestica Population. Trop. Life Sci. Res. 2017, 28, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Bioinsecticides based on plant essential oils: A short overview. Z. Naturforsch. C J. Biosci. 2020, 75, 179–182. [Google Scholar] [CrossRef]
- Weese, J.S. Barrier precautions, isolation protocols, and personal hygiene in veterinary hospitals. Vet. Clin. N. Am. Equine Pract. 2004, 20, 543–559. [Google Scholar] [CrossRef]
- Wierup, M. The control of microbial diseases in animals: Alternatives to the use of antibiotics. Int. J. Antimicrob. Agents 2000, 14, 315–319. [Google Scholar] [CrossRef]
- Martelli, F.; Lambert, M.; Butt, P.; Cheney, T.; Tatone, F.A.; Callaby, R.; Rabie, A.; Gosling, R.J.; Fordon, S.; Crocker, G.; et al. Evaluation of an enhanced cleaning and disinfection protocol in Salmonella contaminated pig holdings in the United Kingdom. PLoS ONE 2017, 12, e0178897. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). Interventions for the Control of Non-Typhoidal Salmonella spp. in Beef and Pork; FAO: Rome, Italy, 2015. [Google Scholar]
- Braden, C.R. Salmonella enterica serotype Enteritidis and eggs: A national epidemic in the United States. Clin. Infect. Dis. 2006, 43, 512–517. [Google Scholar] [CrossRef]
- Jørgensen, F.; Bailey, R.; Williams, S.; Henderson, P.; Wareing, D.R.; Bolton, F.J.; Frost, J.A.; Ward, L.; Humphrey, T.J. Prevalence and numbers of Salmonella and Campylobacter spp. on raw, whole chickens in relation to sampling methods. Int. J. Food Microbiol. 2002, 76, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, M.; Poelstra, J.W.; Kauffman, M.; Varghese, B.; Helmy, Y.A.; Scaria, J.; Rajashekara, G. Genomic Diversity, Antimicrobial Resistance, Plasmidome, and Virulence Profiles of Salmonella Isolated from Small Specialty Crop Farms Revealed by Whole-Genome Sequencing. Antibiotics 2023, 12, 1637. [Google Scholar] [CrossRef]
- Wright, G.D. Antibiotic resistance in the environment: A link to the clinic? Curr. Opin. Microbiol. 2010, 13, 589–594. [Google Scholar] [CrossRef]
- Liljebjelke, K.A.; Hofacre, C.L.; White, D.G.; Ayers, S.; Lee, M.D.; Maurer, J.J. Diversity of antimicrobial resistance phenotypes in Salmonella isolated from commercial poultry farms. Front. Vet. Sci. 2017, 4, 96. [Google Scholar] [CrossRef] [PubMed]
- Sneeringer, S.; MacDonald, J.; Key, N.; McBride, W. Economics of Antibiotic Use in US. Livestock Production; CreateSpace Independent Publishing Platform: Scotts Valley, CA, USA, 2015; pp. 1–100. [Google Scholar]
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef]
- Gebreyes, W.A.; Thakur, S. Multidrug-resistant Salmonella enterica serovar Muenchen from pigs and humans and potential interserovar transfer of antimicrobial resistance. Antimicrob. Agents Chemother. 2005, 49, 503–511. [Google Scholar] [CrossRef]
- Elsayed, M.M.; El-Basrey, Y.F.H.; El-Baz, A.H.; Dowidar, H.A.; Shami, A.; Al-Saeed, F.A.; Alsamghan, A.; Salem, H.M.; Alhazmi, W.A.; El-Tarabily, K.A.; et al. Ecological incidence, genetic diversity, multidrug resistance of Salmonella enteritidis recovered from broiler and layer chicken farms. Poult. Sci. 2023, 103, 103320. [Google Scholar] [CrossRef]
- Srednik, M.E.; Morningstar-Shaw, B.R.; Hicks, J.A.; Tong, C.; Mackie, T.A.; Schlater, L.K. Whole-genome sequencing and phylogenetic analysis capture the emergence of a multi-drug resistant Salmonella enterica serovar Infantis clone from diagnostic animal samples in the United States. Front. Microbiol. 2023, 14, 1166908. [Google Scholar] [CrossRef] [PubMed]
- Leon, I.M.; Lawhon, S.D.; Norman, K.N.; Threadgill, D.S.; Ohta, N.; Vinasco, J.; Scott, H.M. Serotype Diversity and Antimicrobial Resistance among Salmonella enterica Isolates from Patients at an Equine Referral Hospital. Appl. Environ. Microbiol. 2018, 84, e02829-17. [Google Scholar] [CrossRef]
- Gebreyes, W.A.; Thakur, S.; Morrow, W.M. Comparison of prevalence, antimicrobial resistance, and occurrence of multidrug-resistant Salmonella in antimicrobial-free and conventional pig production. J. Food Prot. 2006, 69, 743–748. [Google Scholar] [CrossRef]
- Lynne, A.M.; Dorsey, L.L.; David, D.E.; Foley, S.L. Characterisation of antibiotic resistance in host-adapted Salmonella enterica. Int. J. Antimicrob. Agents 2009, 34, 169–172. [Google Scholar] [CrossRef]
- Alagawany, M.; Abd El-Hack, M.E.; Farag, M.R.; Sachan, S.; Karthik, K.; Dhama, K. The use of probiotics as eco-friendly alternatives for antibiotics in poultry nutrition. Environ. Sci. Pollut. Res. 2018, 25, 10611–10618. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Taha-Abdelaziz, K.; Hawwas, H.A.E.; Ghosh, S.; AlKafaas, S.S.; Moawad, M.M.M.; Saied, E.M.; Kassem, I.I.; Mawad, A.M.M. Antimicrobial Resistance and Recent Alternatives to Antibiotics for the Control of Bacterial Pathogens with an Emphasis on Foodborne Pathogens. Antibiotics 2023, 12, 274. [Google Scholar] [CrossRef] [PubMed]
- de Melo Pereira, G.V.; de Oliveira Coelho, B.; Júnior, A.I.M.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, R.R.; Gaghan, C.; Gorrell, K.; Sharif, S.; Taha-Abdelaziz, K. Probiotics as alternatives to antibiotics for the prevention and control of necrotic enteritis in chickens. Pathogens 2022, 11, 692. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, M.; Bavananthasivam, J.; Shojadoost, B.; Astill, J.; Taha-Abdelaziz, K.; Alqazlan, N.; Boodhoo, N.; Shoja Doost, J.; Sharif, S. In ovo and oral administration of probiotic lactobacilli modulate cell-and antibody-mediated immune responses in newly hatched chicks. Front. Immunol. 2021, 12, 664387. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Tuomola, E.M.; Ouwehand, A.C.; Salminen, S.J. The effect of probiotic bacteria on the adhesion of pathogens to human intestinal mucus. FEMS Immunol. Med. Microbiol. 1999, 26, 137–142. [Google Scholar] [CrossRef]
- Liu, D.; Jiang, X.-Y.; Zhou, L.-S.; Song, J.-H.; Zhang, X. Effects of probiotics on intestinal mucosa barrier in patients with colorectal cancer after operation: Meta-analysis of randomized controlled trials. Medicine 2016, 95, e3342. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in Medicine: A Long Debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef] [PubMed]
- Timmerman, H.M.; Koning, C.J.; Mulder, L.; Rombouts, F.M.; Beynen, A.C. Monostrain, multistrain and multispecies probiotics—A comparison of functionality and efficacy. Int. J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-C.; Hu, S.-Y.; Chiu, C.-S.; Liu, C.-H. Multiple-strain probiotics appear to be more effective in improving the growth performance and health status of white shrimp, Litopenaeus vannamei, than single probiotic strains. Fish Shellfish Immunol. 2019, 84, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Kathayat, D.; Closs, G.; Helmy, Y.A.; Deblais, L.; Srivastava, V.; Rajashekara, G. In Vitro and In Vivo Evaluation of Lacticaseibacillus rhamnosus GG and Bifidobacterium lactis Bb12 Against Avian Pathogenic Escherichia coli and Identification of Novel Probiotic-Derived Bioactive Peptides. Probiotics Antimicrob. Proteins 2022, 14, 1012–1028. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Invernici, M.M.; Furlaneto, F.A.; Messora, M.R. Effectiveness of multi-strain versus single-strain probiotics: Current status and recommendations for the future. J. Clin. Gastroenterol. 2018, 52, S35–S40. [Google Scholar] [CrossRef] [PubMed]
- Puvanasundram, P.; Chong, C.M.; Sabri, S.; Yusoff, M.S.M.; Lim, K.C.; Karim, M. Efficacy of Single and Multi-Strain Probiotics on In Vitro Strain Compatibility, Pathogen Inhibition, Biofilm Formation Capability, and Stress Tolerance. Biology 2022, 11, 1644. [Google Scholar] [CrossRef]
- Mountzouris, K.; Balaskas, C.; Xanthakos, I.; Tzivinikou, A.; Fegeros, K. Effects of a multi-species probiotic on biomarkers of competitive exclusion efficacy in broilers challenged with Salmonella enteritidis. Br. Poult. Sci. 2009, 50, 467–478. [Google Scholar] [CrossRef]
- Thirabunyanon, M.; Thongwittaya, N. Protection activity of a novel probiotic strain of Bacillus subtilis against Salmonella Enteritidis infection. Res. Vet. Sci. 2012, 93, 74–81. [Google Scholar] [CrossRef]
- Chang, C.H.; Teng, P.Y.; Lee, T.T.; Yu, B. Effects of multi-strain probiotic supplementation on intestinal microbiota, tight junctions, and inflammation in young broiler chickens challenged with Salmonella enterica subsp. enterica. Asian-Australas. J. Anim. Sci. 2020, 33, 1797. [Google Scholar] [CrossRef]
- Acharjee, M.; Hasan, F.; Islam, T.; Nur, I.T.; Begum, N.; Mazumder, C.; Lubna, M.A.; Zerin, N.; Shahriar, A.; Mahmud, M.R. In-vitro antibacterial activity of commercially available probiotics on food-borne pathogens along with their synergistic effects with synthetic drugs. Metab. Open 2022, 14, 100187. [Google Scholar] [CrossRef]
- Bhogoju, S.; Nahashon, S.; Wang, X.; Darris, C.; Kilonzo-Nthenge, A. A comparative analysis of microbial profile of Guinea fowl and chicken using metagenomic approach. PLoS ONE 2018, 13, e0191029. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, H.; Tahoun, A.; Rizk, A.M.; Suzuki, T.; Elmonir, W.; Nassef, E.; Shukry, M.; Germoush, M.O.; Farrag, F.; Bin-Jumah, M.; et al. Evaluation of Bifidobacteria and Lactobacillus Probiotics as Alternative Therapy for Salmonella typhimurium Infection in Broiler Chickens. Animals 2020, 10, 1023. [Google Scholar] [CrossRef] [PubMed]
- Pascual, M.; Hugas, M.; Badiola, J.I.; Monfort, J.M.; Garriga, M. Lactobacillus salivarius CTC2197 prevents Salmonella enteritidis colonization in chickens. Appl. Environ. Microbiol. 1999, 65, 4981–4986. [Google Scholar] [CrossRef]
- Van Coillie, E.; Goris, J.; Cleenwerck, I.; Grijspeerdt, K.; Botteldoorn, N.; Van Immerseel, F.; De Buck, J.; Vancanneyt, M.; Swings, J.; Herman, L. Identification of lactobacilli isolated from the cloaca and vagina of laying hens and characterization for potential use as probiotics to control Salmonella Enteritidis. J. Appl. Microbiol. 2007, 102, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Gil De Los Santos, J.; Storch, O.; Gil-Turnes, C. Bacillus cereus var. toyoii and Saccharomyces boulardii increased feed efficiency in broilers infected with Salmonella enteritidis. Br. Poult. Sci. 2005, 46, 494–497. [Google Scholar] [CrossRef]
- Okamoto, A.S.; Andreatti Filho, R.L.; Milbradt, E.L.; Moraes, A.C.I.; Vellano, I.H.B.; Guimarães-Okamoto, P.T.C. Bacterial communication between Lactobacillus spp. isolated from poultry in the inhibition of Salmonella Heidelberg—Proof of concept. Poult. Sci. 2018, 97, 2708–2712. [Google Scholar] [CrossRef]
- Shao, Y.; Zhen, W.; Guo, F.; Hu, Z.; Zhang, K.; Kong, L.; Guo, Y.; Wang, Z. Pretreatment with probiotics Enterococcus faecium NCIMB 11181 attenuated Salmonella Typhimurium-induced gut injury through modulating intestinal microbiome and immune responses with barrier function in broiler chickens. J. Anim. Sci. Biotechnol. 2022, 13, 130. [Google Scholar] [CrossRef]
- Martins, F.S.; Vieira, A.T.; Elian, S.D.; Arantes, R.M.; Tiago, F.C.; Sousa, L.P.; Araújo, H.R.; Pimenta, P.F.; Bonjardim, C.A.; Nicoli, J.R. Inhibition of tissue inflammation and bacterial translocation as one of the protective mechanisms of Saccharomyces boulardii against Salmonella infection in mice. Microbes Infect. 2013, 15, 270–279. [Google Scholar] [CrossRef]
- Shanmugasundaram, R.; Mortada, M.; Cosby, D.; Singh, M.; Applegate, T.; Syed, B.; Pender, C.; Curry, S.; Murugesan, G.; Selvaraj, R. Synbiotic supplementation to decrease Salmonella colonization in the intestine and carcass contamination in broiler birds. PLoS ONE 2019, 14, e0223577. [Google Scholar] [CrossRef]
- Kimminau, E.; Karnezos, T.; Berghaus, R.; Jones, M.; Baxter, J.; Hofacre, C. Combination of probiotic and prebiotic impacts Salmonella Enteritidis infection in layer hens. J. Appl. Poult. Res. 2021, 30, 100200. [Google Scholar] [CrossRef]
- Medina, M.; Izquierdo, E.; Ennahar, S.; Sanz, Y. Differential immunomodulatory properties of Bifidobacterium logum strains: Relevance to probiotic selection and clinical applications. Clin. Exp. Immunol. 2007, 150, 531–538. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Forsythe, S.J.; El-Nezami, H. Probiotics interaction with foodborne pathogens: A potential alternative to antibiotics and future challenges. Crit. Rev. Food Sci. Nutr. 2019, 59, 3320–3333. [Google Scholar] [CrossRef] [PubMed]
- Juricova, H.; Matiasovicova, J.; Faldynova, M.; Sebkova, A.; Kubasova, T.; Prikrylova, H.; Karasova, D.; Crhanova, M.; Havlickova, H.; Rychlik, I. Probiotic Lactobacilli Do Not Protect Chickens against Salmonella Enteritidis Infection by Competitive Exclusion in the Intestinal Tract but in Feed, Outside the Chicken Host. Microorganisms 2022, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Nawaz, M.; Anjum, A.A.; Ahmad, M.-u.-D.; Mehmood, A.; Rabbani, M.; Mustafa, A.; Ali, M.A. Effect of indigenous probiotics on gut morphology and intestinal absorption capacity in broiler chicken challenged with Salmonella enteritidis. Pak. J. Zool. 2020, 52, 1825. [Google Scholar] [CrossRef]
- Steinberg, R.S.; Silva, L.C.; Souza, T.C.; Lima, M.T.; De Oliveira, N.L.; Vieira, L.Q.; Arantes, R.M.; Miyoshi, A.; Nicoli, J.R.; Neumann, E. Safety and protective effectiveness of two strains of Lactobacillus with probiotic features in an experimental model of salmonellosis. Int. J. Environ. Res. Public Health 2014, 11, 8755–8776. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, H.R.; Abdul-Careem, M.F.; Dara, R.A.; Chambers, J.R.; Sharif, S. Cytokine gene expression in chicken cecal tonsils following treatment with probiotics and Salmonella infection. Vet. Microbiol. 2008, 126, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Casey, P.G.; Gardiner, G.E.; Casey, G.; Bradshaw, B.; Lawlor, P.G.; Lynch, P.B.; Leonard, F.C.; Stanton, C.; Ross, R.P.; Fitzgerald, G.F. A five-strain probiotic combination reduces pathogen shedding and alleviates disease signs in pigs challenged with Salmonella enterica serovar Typhimurium. Appl. Environ. Microbiol. 2007, 73, 1858–1863. [Google Scholar] [CrossRef]
- Truusalu, K.; Naaber, P.; Kullisaar, T.; Tamm, H.; Mikelsaar, R.-H.; Zilmer, K.; Rehema, A.; Zilmer, M.; Mikelsaar, M. The influence of antibacterial and antioxidative probiotic lactobacilli on gut mucosa in a mouse model of Salmonella infection. Microb. Ecol. Health Dis. 2004, 16, 180–187. [Google Scholar]
- Chen, Q.; Tong, C.; Ma, S.; Zhou, L.; Zhao, L.; Zhao, X. Involvement of microRNAs in probiotics-induced reduction of the cecal inflammation by Salmonella typhimurium. Front. Immunol. 2017, 8, 704. [Google Scholar] [CrossRef]
- Huang, Y.-F.; Liu, P.-Y.; Chen, Y.-Y.; Nong, B.-R.; Huang, I.-F.; Hsieh, K.-S.; Chen, K.-T. Three-combination probiotics therapy in children with salmonella and rotavirus gastroenteritis. J. Clin. Gastroenterol. 2014, 48, 37–42. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Shanmugam, S.K.; Kang, D.K.; Kim, I.H. Preliminary assessment on potentials of probiotic B. subtilis RX7 and B. methylotrophicus C14 strains as an immune modulator in Salmonella-challenged weaned pigs. Trop. Anim. Health Prod. 2017, 49, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Ward, M.; Alinovi, C.; Couetil, L.; Glickman, L.; Wu, C. randomized clinical trial using probiotics to prevent Salmonella fecal shedding in hospitalized horses. J. Equine Vet. Sci. 2004, 24, 242–247. [Google Scholar] [CrossRef]
- Sun, C.; Gao, X.; Sun, M.; Wang, Z.; Wang, Y.; Zhao, X.; Jia, F.; Zhang, T.; Ge, C.; Zhang, X.; et al. Protective effects of E. coli Nissle 1917 on chickens infected with Salmonella pullorum. Microb. Pathog. 2022, 172, 105768. [Google Scholar] [CrossRef] [PubMed]
- Smialek, M.; Kaczorek, E.; Szczucińska, E.; Burchardt, S.; Kowalczyk, J.; Tykałowski, B.; Koncicki, A. Evaluation of Lactobacillus spp. and yeast based probiotic (Lavipan) supplementation for the reduction of Salmonella Enteritidis after infection of broiler chickens. Pol. J. Vet. Sci. 2019, 22, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Savino, F.; Fornasero, S.; Ceratto, S.; De Marco, A.; Mandras, N.; Roana, J.; Tullio, V.; Amisano, G. Probiotics and gut health in infants: A preliminary case–control observational study about early treatment with Lactobacillus reuteri DSM 17938. Clin. Chim. Acta 2015, 451, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Pineiro, M.; Asp, N.-G.; Reid, G.; Macfarlane, S.; Morelli, L.; Brunser, O.; Tuohy, K. FAO Technical meeting on prebiotics. J. Clin. Gastroenterol. 2008, 42, S156–S159. [Google Scholar] [CrossRef] [PubMed]
- Cazzola, M.; Pham-Thi, N.; Kerihuel, J.-C.; Durand, H.; Bohbot, S. Efficacy of a synbiotic supplementation in the prevention of common winter diseases in children: A randomized, double-blind, placebo-controlled pilot study. Ther. Adv. Respir. Dis. 2010, 4, 271–278. [Google Scholar] [CrossRef]
- Ojansivu, I.; Ferreira, C.L.; Salminen, S. Yacon, a new source of prebiotic oligosaccharides with a history of safe use. Trends Food Sci. Technol. 2011, 22, 40–46. [Google Scholar] [CrossRef]
- Passeron, T.; Lacour, J.P.; Fontas, E.; Ortonne, J.P. Prebiotics and synbiotics: Two promising approaches for the treatment of atopic dermatitis in children above 2 years. Allergy 2006, 61, 431–437. [Google Scholar] [CrossRef]
- Macfarlane, S.; Macfarlane, G.T.; Cummings, J.H. Review article: Prebiotics in the gastrointestinal tract. Aliment Pharmacol. Ther. 2006, 24, 701–714. [Google Scholar] [CrossRef]
- De Vrese, M.; Schrezenmeir, J. Probiotics, prebiotics, and synbiotics. Food Biotechnol. 2008, 111, 1–66. [Google Scholar]
- Gibson, G.R.; Probert, H.M.; Van Loo, J.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Closs, G., Jr.; Jung, K.; Kathayat, D.; Vlasova, A.; Rajashekara, G. Effect of Probiotic E. coli Nissle 1917 Supplementation on the Growth Performance, Immune Responses, Intestinal Morphology, and Gut Microbes of Campylobacter jejuni Infected Chickens. Infect. Immun. 2022, 90, e0033722. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Kassem, I.I.; Kumar, A.; Rajashekara, G. In Vitro Evaluation of the Impact of the Probiotic E. coli Nissle 1917 on Campylobacter jejuni’s Invasion and Intracellular Survival in Human Colonic Cells. Front. Microbiol. 2017, 8, 1588. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Kassem, I.I.; Rajashekara, G. Immuno-modulatory effect of probiotic E. coli Nissle 1917 in polarized human colonic cells against Campylobacter jejuni infection. Gut Microbes 2021, 13, 1–16. [Google Scholar] [CrossRef]
- Mawad, A.; Helmy, Y.A.; Shalkami, A.G.; Kathayat, D.; Rajashekara, G.E. coli Nissle microencapsulation in alginate-chitosan nanoparticles and its effect on Campylobacter jejuni in vitro. Appl. Microbiol. Biotechnol. 2018, 102, 10675–10690. [Google Scholar] [CrossRef]
- Al-Sheraji, S.H.; Ismail, A.; Manap, M.Y.; Mustafa, S.; Yusof, R.M.; Hassan, F.A. Prebiotics as functional foods: A review. J. Funct. Foods 2013, 5, 1542–1553. [Google Scholar] [CrossRef]
- Gibson, G.R.; Scott, K.P.; Rastall, R.A.; Tuohy, K.M.; Hotchkiss, A.; Dubert-Ferrandon, A.; Gareau, M.; Murphy, E.F.; Saulnier, D.; Loh, G. Dietary prebiotics: Current status and new definition. Food Sci. Technol. Bull. Funct. Foods 2010, 7, 1–19. [Google Scholar] [CrossRef]
- Chen, T. Effect of adding chicory fructans in feed on fecal and intestinal microflora and excreta volatile ammonia. Int. J. Poult. Sci. 2003, 2, 188–194. [Google Scholar]
- Bailey, J.; Blankenship, L.; Cox, N. Effect of fructooligosaccharide on Salmonella colonization of the chicken intestine. Poult. Sci. 1991, 70, 2433–2438. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Namkung, H.; Paik, I. Effects of dietary fructooligosaccharides on the suppression of intestinal colonization of Salmonella typhimurium in broiler chickens. Korean J. Anim. Sci. 1994, 82, 1030–1036. [Google Scholar]
- Vicente, J.; Wolfenden, A.; Torres-Rodriguez, A.; Higgins, S.; Tellez, G.; Hargis, B. Effect of a Lactobacillus species-based probiotic and dietary lactose prebiotic on turkey poult performance with or without Salmonella enteritidis challenge. J. Appl. Poult. Res. 2007, 16, 361–364. [Google Scholar] [CrossRef]
- Sobotik, E.B.; Ramirez, S.; Roth, N.; Tacconi, A.; Pender, C.; Murugesan, R.; Archer, G.S. Evaluating the effects of a dietary synbiotic or synbiotic plus enhanced organic acid on broiler performance and cecal and carcass Salmonella load. Poult. Sci. 2021, 100, 101508. [Google Scholar] [CrossRef]
- Petersen, A.; Heegaard, P.M.; Pedersen, A.L.; Andersen, J.B.; Sørensen, R.B.; Frøkiær, H.; Lahtinen, S.J.; Ouwehand, A.C.; Poulsen, M.; Licht, T.R. Some putative prebiotics increase the severity of Salmonella enterica serovar Typhimurium infection in mice. BMC Microbiol. 2009, 9, 245. [Google Scholar] [CrossRef]
- Ribeiro, A.M.L.; Vogt, L.K.; Canal, C.W.; Cardoso, M.d.I.; Labres, R.V.; Streck, A.F.; Bessa, M.C. Effects of prebiotics and probiotics on the colonization and immune response of broiler chickens challenged with Salmonella enteritidis. Braz. J. Poult. Sci. 2007, 9, 193–200. [Google Scholar] [CrossRef]
- Rodríguez-Sorrento, A.; Castillejos, L.; López-Colom, P.; Cifuentes-Orjuela, G.; Rodríguez-Palmero, M.; Moreno-Muñoz, J.A.; Martin-Orue, S.M. Effects of Bifidobacterium longum subsp. infantis CECT 7210 and Lactobacillus rhamnosus HN001, combined or not with oligofructose-enriched inulin, on weaned pigs orally challenged with Salmonella typhimurium. Front. Microbiol. 2020, 11, 2012. [Google Scholar] [CrossRef]
- Murate, L.S.; Paião, F.G.; de Almeida, A.M.; Berchieri, A., Jr.; Shimokomaki, M. Efficacy of prebiotics, probiotics, and synbiotics on laying hens and broilers challenged with Salmonella Enteritidis. J. Poult. Sci. 2015, 52, 52–56. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Lai, Y.; Villaruz, A.E.; Li, M.; Cha, D.J.; Sturdevant, D.E.; Otto, M. The human anionic antimicrobial peptide dermcidin induces proteolytic defence mechanisms in staphylococci. Mol. Microbiol. 2007, 63, 497–506. [Google Scholar] [CrossRef]
- Malkoski, M.; Dashper, S.G.; O’Brien-Simpson, N.M.; Talbo, G.H.; Macris, M.; Cross, K.J.; Reynolds, E.C. Kappacin, a novel antibacterial peptide from bovine milk. Antimicrob. Agents Chemother. 2001, 45, 2309–2315. [Google Scholar] [CrossRef] [PubMed]
- Oren, Z.; Shai, Y. Mode of action of linear amphipathic alpha-helical antimicrobial peptides. Biopolymers 1998, 47, 451–463. [Google Scholar] [CrossRef]
- Lohner, K.; Prossnigg, F. Biological activity and structural aspects of PGLa interaction with membrane mimetic systems. Biochim. Biophys. Acta 2009, 1788, 1656–1666. [Google Scholar] [CrossRef] [PubMed]
- Mardirossian, M.; Pérébaskine, N.; Benincasa, M.; Gambato, S.; Hofmann, S.; Huter, P.; Müller, C.; Hilpert, K.; Innis, C.A.; Tossi, A. The dolphin proline-rich antimicrobial peptide Tur1A inhibits protein synthesis by targeting the bacterial ribosome. Cell Chem. Biol. 2018, 25, 530–539.E7. [Google Scholar] [CrossRef] [PubMed]
- Le, C.-F.; Gudimella, R.; Razali, R.; Manikam, R.; Sekaran, S.D. Transcriptome analysis of Streptococcus pneumoniae treated with the designed antimicrobial peptides, DM3. Sci. Rep. 2016, 6, 26828. [Google Scholar] [CrossRef]
- He, S.-w.; Zhang, J.; Li, N.-q.; Zhou, S.; Yue, B.; Zhang, M. A TFPI-1 peptide that induces degradation of bacterial nucleic acids, and inhibits bacterial and viral infection in half-smooth tongue sole, Cynoglossus semilaevis. Fish Shellfish Immunol. 2017, 60, 466–473. [Google Scholar] [CrossRef]
- Le, C.-F.; Fang, C.-M.; Sekaran, S.D. Intracellular targeting mechanisms by antimicrobial peptides. Antimicrob. Agents Chemother. 2017, 61, e02340-16. [Google Scholar] [CrossRef]
- Cruz, G.F.; de Araujo, I.; Torres, M.D.; de la Fuente-Nunez, C.; Oliveira, V.X.; Ambrosio, F.N.; Lombello, C.B.; Almeida, D.V.; Silva, F.D.; Garcia, W. Photochemically-generated silver chloride nanoparticles stabilized by a peptide inhibitor of cell division and its antimicrobial properties. J. Inorg. Organomet. Polym. Mater. 2020, 30, 2464–2474. [Google Scholar] [CrossRef]
- Almarwani, B.; Phambu, N.; Hamada, Y.Z.; Sunda-Meya, A. Interactions of an anionic antimicrobial peptide with Zinc (II): Application to bacterial mimetic membranes. Langmuir 2020, 36, 14554–14562. [Google Scholar] [CrossRef]
- Festa, R.; Ambrosio, R.L.; Lamas, A.; Gratino, L.; Palmieri, G.; Franco, C.M.; Cepeda, A.; Anastasio, A. A study on the antimicrobial and antibiofilm peptide 1018-K6 as potential alternative to antibiotics against food-pathogen Salmonella enterica. Foods 2021, 10, 1372. [Google Scholar] [CrossRef]
- Sengkhui, S.; Klubthawee, N.; Aunpad, R. A novel designed membrane-active peptide for the control of foodborne Salmonella enterica serovar Typhimurium. Sci. Rep. 2023, 13, 3507. [Google Scholar] [CrossRef] [PubMed]
- Mangmee, S.; Reamtong, O.; Kalambaheti, T.; Roytrakul, S.; Sonthayanon, P. Antimicrobial Peptide Modifications against Clinically Isolated Antibiotic-Resistant Salmonella. Molecules 2021, 26, 4654. [Google Scholar] [CrossRef] [PubMed]
- Klubthawee, N.; Aunpad, R. A Thermostable, Modified Cathelicidin-Derived Peptide With Enhanced Membrane-Active Activity Against Salmonella enterica serovar Typhimurium. Front. Microbiol. 2021, 11, 592220. [Google Scholar] [CrossRef] [PubMed]
- Forkus, B.; Ritter, S.; Vlysidis, M.; Geldart, K.; Kaznessis, Y.N. Antimicrobial probiotics reduce Salmonella enterica in turkey gastrointestinal tracts. Sci. Rep. 2017, 7, 40695. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Q.; Dong, M.; Song, H.; Hang, B.; Sun, Y.; Zhang, H.; Hu, J. Evaluation of the efficacy of the antimicrobial peptide HJH-3 in chickens infected with Salmonella Pullorum. Front. Microbiol. 2023, 14, 1102789. [Google Scholar] [CrossRef] [PubMed]
- Yeom, J.-H.; Lee, B.; Kim, D.; Lee, J.-k.; Kim, S.; Bae, J.; Park, Y.; Lee, K. Gold nanoparticle-DNA aptamer conjugate-assisted delivery of antimicrobial peptide effectively eliminates intracellular Salmonella enterica serovar Typhimurium. Biomaterials 2016, 104, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Kumaresan, V.; Bhatt, P.; Ganesh, M.-R.; Harikrishnan, R.; Arasu, M.; Al-Dhabi, N.A.; Pasupuleti, M.; Marimuthu, K.; Arockiaraj, J. A novel antimicrobial peptide derived from fish goose type lysozyme disrupts the membrane of Salmonella enterica. Mol. Immunol. 2015, 68, 421–433. [Google Scholar] [CrossRef]
- Tuxpan-Pérez, A.; Ibarra-Valencia, M.A.; Estrada, B.E.; Clement, H.; Corrales-García, L.L.; Espino-Solis, G.P.; Corzo, G. Antimicrobial and Immunomodulatory Effects of Selected Chemokine and Antimicrobial Peptide on Cytokine Profile during Salmonella Typhimurium Infection in Mouse. Antibiotics 2022, 11, 607. [Google Scholar] [CrossRef]
- Roque-Borda, C.A.; Pereira, L.P.; Guastalli, E.A.L.; Soares, N.M.; Mac-Lean, P.A.B.; Salgado, D.D.A.; Meneguin, A.B.; Chorilli, M.; Vicente, E.F. Hpmcp-coated microcapsules containing the ctx (Ile21)-ha antimicrobial peptide reduce the mortality rate caused by resistant Salmonella enteritidis in laying hens. Antibiotics 2021, 10, 616. [Google Scholar] [CrossRef]
- Bailleul, G.; Guabiraba, R.; Virlogeux-Payant, I.; Lantier, I.; Trotereau, J.; Gilbert, F.B.; Wiedemann, A.; Trotereau, A.; Velge, P.; Schouler, C. Systemic administration of avian defensin 7: Distribution, cellular target, and antibacterial potential in mice. Front. Microbiol. 2019, 10, 541. [Google Scholar] [CrossRef]
- Maiti, S.; Patro, S.; Purohit, S.; Jain, S.; Senapati, S.; Dey, N. Effective control of Salmonella infections by employing combinations of recombinant antimicrobial human β-defensins hBD-1 and hBD-2. Antimicrob. Agents Chemother. 2014, 58, 6896–6903. [Google Scholar] [CrossRef] [PubMed]
- Milona, P.; Townes, C.L.; Bevan, R.M.; Hall, J. The chicken host peptides, gallinacins 4, 7, and 9 have antimicrobial activity against Salmonella serovars. Biochem. Biophys. Res. Commun. 2007, 356, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Britannica, T. (Ed.) Argon. In Encyclopedia Britannica; Encyclopædia Britannica, Inc.: Chicago, IL, USA, 2020. [Google Scholar]
- Kasman, L.M.; Porter, L.D. Bacteriophages. In StatPearls; StatPearls Publishing: St. Petersburg, FL, USA, 2021. [Google Scholar]
- Doron, S.; Melamed, S.; Ofir, G.; Leavitt, A.; Lopatina, A.; Keren, M.; Amitai, G.; Sorek, R. Systematic discovery of antiphage defense systems in the microbial pangenome. Science 2018, 359, eaar4120. [Google Scholar] [CrossRef]
- Campbell, A. The future of bacteriophage biology. Nat. Rev. Genet. 2003, 4, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Wernicki, A.; Nowaczek, A.; Urban-Chmiel, R. Bacteriophage therapy to combat bacterial infections in poultry. Virol. J. 2017, 14, 179. [Google Scholar] [CrossRef] [PubMed]
- Fiorentin, L.; Vieira, N.D.; Barioni, W., Jr. Oral treatment with bacteriophages reduces the concentration of Salmonella Enteritidis PT4 in caecal contents of broilers. Avian Pathol. 2005, 34, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.S.; Rahman, S.R. Use of Phages to Treat Antimicrobial-Resistant Salmonella Infections in Poultry. Vet. Sci. 2022, 9, 438. [Google Scholar] [CrossRef]
- Henriques, A.; Sereno, R.; Almeida, A. Reducing Salmonella horizontal transmission during egg incubation by phage therapy. Foodborne Pathog. Dis. 2013, 10, 718–722. [Google Scholar] [CrossRef]
- Lee, S.; Kwon, T.; Chae, S.-J.; Kim, J.-H.; Kang Yeon, H.; Chung Gyung, T.; Kim, D.-W.; Lee, D.-Y. Complete Genome Sequence of Bacteriophage MA12, Which Infects both Campylobacter jejuni and Salmonella enterica Serovar Enteritidis. Genome Announc. 2016, 4, e00810–e00816. [Google Scholar] [CrossRef]
- Lorenzo-Rebenaque, L.; Malik, D.J.; Catalá-Gregori, P.; Marin, C.; Sevilla-Navarro, S. In Vitro and In Vivo Gastrointestinal Survival of Non-Encapsulated and Microencapsulated Salmonella Bacteriophages: Implications for Bacteriophage Therapy in Poultry. Pharmaceuticals 2021, 14, 434. [Google Scholar] [CrossRef]
- Pelyuntha, W.; Sanguankiat, A.; Kovitvadhi, A.; Vongkamjan, K. Broad lytic spectrum of novel Salmonella phages on ciprofloxacin-resistant Salmonella contaminated in the broiler production chain. Vet. World 2022, 15, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Spricigo, D.A.; Bardina, C.; Cortés, P.; Llagostera, M. Use of a bacteriophage cocktail to control Salmonella in food and the food industry. Int. J. Food Microbiol. 2013, 165, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Duc, H.M.; Son, H.M.; Honjoh, K.-I.; Miyamoto, T. Isolation and application of bacteriophages to reduce Salmonella contamination in raw chicken meat. LWT 2018, 91, 353–360. [Google Scholar] [CrossRef]
- Hyman, P.; Abedon, S.T. Bacteriophage host range and bacterial resistance. Adv. Appl. Microbiol. 2010, 70, 217–248. [Google Scholar] [PubMed]
- Gill, J.J.; Hyman, P. Phage choice, isolation, and preparation for phage therapy. Curr. Pharm. Biotechnol. 2010, 11, 2–14. [Google Scholar] [CrossRef]
- Callaway, T.R.; Edrington, T.S.; Brabban, A.D.; Anderson, R.C.; Rossman, M.L.; Engler, M.J.; Carr, M.A.; Genovese, K.J.; Keen, J.E.; Looper, M.L. Bacteriophage isolated from feedlot cattle can reduce Escherichia coli O157: H7 populations in ruminant gastrointestinal tracts. Foodborne Pathog. Dis. 2008, 5, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Tetz, G.V.; Ruggles, K.V.; Zhou, H.; Heguy, A.; Tsirigos, A.; Tetz, V. Bacteriophages as potential new mammalian pathogens. Sci. Rep. 2017, 7, 7043. [Google Scholar] [CrossRef]
- Merril, C.R. Interaction of bacteriophages with animals. In Bacteriophage Ecology; Cambridge University Press: Cambridge, UK, 2008; pp. 332–352. [Google Scholar]
- Kosznik-Kwaśnicka, K.; Podlacha, M.; Grabowski, Ł.; Stasiłojć, M.; Nowak-Zaleska, A.; Ciemińska, K.; Cyske, Z.; Dydecka, A.; Gaffke, L.; Mantej, J.; et al. Biological aspects of phage therapy versus antibiotics against Salmonella enterica serovar Typhimurium infection of chickens. Front. Cell. Infect. Microbiol. 2022, 12, 941867. [Google Scholar] [CrossRef]
- Wong, C.L.; Sieo, C.C.; Tan, W.S.; Abdullah, N.; Hair-Bejo, M.; Abu, J.; Ho, Y.W. Evaluation of a lytic bacteriophage, Φ st1, for biocontrol of Salmonella enterica serovar Typhimurium in chickens. Int. J. Food. Microbiol. 2014, 172, 92–101. [Google Scholar] [CrossRef]
- Esmael, A.; Azab, E.; Gobouri, A.A.; Nasr-Eldin, M.A.; Moustafa, M.M.A.; Mohamed, S.A.; Badr, O.A.M.; Abdelatty, A.M. Isolation and Characterization of Two Lytic Bacteriophages Infecting a Multi-Drug Resistant Salmonella Typhimurium and Their Efficacy to Combat Salmonellosis in Ready-to-Use Foods. Microorganisms 2021, 9, 423. [Google Scholar] [CrossRef]
- Bardina, C.; Spricigo, D.A.; Cortés, P.; Llagostera, M. Significance of the bacteriophage treatment schedule in reducing Salmonella colonization of poultry. Appl. Environ. Microbiol. 2012, 78, 6600–6607. [Google Scholar] [CrossRef]
- Atterbury, R.J.; Van Bergen, M.; Ortiz, F.; Lovell, M.; Harris, J.; De Boer, A.; Wagenaar, J.; Allen, V.; Barrow, P. Bacteriophage therapy to reduce Salmonella colonization of broiler chickens. Appl. Environ. Microbiol. 2007, 73, 4543–4549. [Google Scholar] [CrossRef] [PubMed]
- Bielke, L.R.; Higgins, S.E.; Donoghue, A.M.; Donoghue, D.J.; Hargis, B.M.; Tellez, G. Use of Wide-Host-Range Bacteriophages to Reduce Salmonella on Poultry Products. Int. J. Poult. Sci. 2007, 6, 754–757. [Google Scholar]
- Nabil, N.M.; Tawakol, M.M.; Hassan, H.M. Assessing the impact of bacteriophages in the treatment of Salmonella in broiler chickens. Infect. Ecol. Epidemiol. 2018, 8, 1539056. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.H.; Kim, M.S.; Lee, D.H.; Lee, Y.N.; Park, J.K.; Youn, H.N.; Lee, H.J.; Yang, S.Y.; Cho, Y.W.; Lee, J.B.; et al. Use of bacteriophage for biological control of Salmonella Enteritidis infection in chicken. Res. Vet. Sci. 2012, 93, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Sonalika, J.; Srujana, A.; Akhila, D.; Juliet, M.; Santhosh, K. Application of bacteriophages to control Salmonella Enteritidis in raw eggs. Iran. J. Vet. Res. 2020, 21, 221. [Google Scholar]
- Zhang, Y.; Ding, Y.; Li, W.; Zhu, W.; Wang, J.; Wang, X. Application of a novel lytic podoviridae phage Pu20 for biological control of drug-resistant Salmonella in liquid eggs. Pathogens 2021, 10, 34. [Google Scholar] [CrossRef]
- Rivera, D.; Moreno-Switt, A.I.; Denes, T.G.; Hudson, L.K.; Peters, T.L.; Samir, R.; Aziz, R.K.; Noben, J.-P.; Wagemans, J.; Dueñas, F. Novel Salmonella phage, vB_Sen_STGO-35-1, characterization and evaluation in chicken meat. Microorganisms 2022, 10, 606. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Maurer, C.K.; Lu, C.; Empting, M.; Hartmann, R.W. Synthetic quorum sensing inhibitors (QSIs) blocking receptor signaling or signal molecule biosynthesis in Pseudomonas aeruginosa. In Quorum Sensing vs Quorum Quenching: A Battle with No End in Sight; Springer: Berlin/Heidelberg, Germany, 2014; pp. 303–317. [Google Scholar]
- Tirkkonen, H.; Brown, K.V.; Niemczura, M.; Faudemer, Z.; Brown, C.; Ponomareva, L.V.; Helmy, Y.A.; Thorson, J.S.; Nybo, S.E.; Metsä-Ketelä, M. Engineering BioBricks for Deoxysugar Biosynthesis and Generation of New Tetracenomycins. ACS Omega 2023, 8, 21237–21253. [Google Scholar] [CrossRef]
- Alshawwa, S.Z.; Alshallash, K.S.; Ghareeb, A.; Elazzazy, A.M.; Sharaf, M.; Alharthi, A.; Abdelgawad, F.E.; El-Hossary, D.; Jaremko, M.; Emwas, A.-H.; et al. Assessment of Pharmacological Potential of Novel Exopolysaccharide Isolated from Marine Kocuria sp. Strain AG5: Broad-Spectrum Biological Investigations. Life 2022, 12, 1387. [Google Scholar] [CrossRef] [PubMed]
- Ta, C.A.; Arnason, J.T. Mini Review of Phytochemicals and Plant Taxa with Activity as Microbial Biofilm and Quorum Sensing Inhibitors. Molecules 2015, 21, E29. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Muciño, E.; Arenas-Hernández, M.M.P.; Luna-Guevara, M.L. Mechanisms of Inhibition of Quorum Sensing as an Alternative for the Control of E. coli and Salmonella. Microorganisms 2022, 10, 884. [Google Scholar] [CrossRef]
- Guo, M.; Gamby, S.; Zheng, Y.; Sintim, H.O. Small molecule inhibitors of AI-2 signaling in bacteria: State-of-the-art and future perspectives for anti-quorum sensing agents. Int. J. Mol. Sci. 2013, 14, 17694–17728. [Google Scholar] [CrossRef]
- Witsø, I.L.; Valen Rukke, H.; Benneche, T.; Aamdal Scheie, A. Thiophenone attenuates enteropathogenic Escherichia coli O103: H2 virulence by interfering with AI-2 signaling. PLoS ONE 2016, 11, e0157334. [Google Scholar] [CrossRef] [PubMed]
- Vinothkannan, R.; Muthu Tamizh, M.; David Raj, C.; Adline Princy, S. Fructose furoic acid ester: An effective quorum sensing inhibitor against uropathogenic Escherichia coli. Bioorganic Chem. 2018, 79, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, L.; Xu, J.; Liu, S.; Song, Z.; Chen, T.; Deng, X.; Wang, J.; Lv, Q. Quercitrin Is a Novel Inhibitor of Salmonella enterica Serovar Typhimurium Type III Secretion System. Molecules 2023, 28, 5455. [Google Scholar] [CrossRef]
- Deblais, L.; Helmy, Y.A.; Kathayat, D.; Huang, H.-C.; Miller, S.A.; Rajashekara, G. Novel Imidazole and Methoxybenzylamine Growth Inhibitors Affecting Salmonella Cell Envelope Integrity and its Persistence in Chickens. Sci. Rep. 2018, 8, 13381. [Google Scholar] [CrossRef]
- Koopman Jacob, A.; Marshall Joanna, M.; Bhatiya, A.; Eguale, T.; Kwiek Jesse, J.; Gunn John, S. Inhibition of Salmonella enterica Biofilm Formation Using Small-Molecule Adenosine Mimetics. Antimicrob. Agents Chemother. 2014, 59, 76–84. [Google Scholar] [CrossRef]
- Nagy, T.A.; Quintana, J.L.J.; Reens, A.L.; Crooks, A.L.; Detweiler, C.S. Autophagy Induction by a Small Molecule Inhibits Salmonella Survival in Macrophages and Mice. Antimicrob. Agents Chemother. 2019, 63, e01536-19. [Google Scholar] [CrossRef]
- Reens, A.L.; Crooks, A.L.; Su, C.-C.; Nagy, T.A.; Reens, D.L.; Podoll, J.D.; Edwards, M.E.; Yu, E.W.; Detweiler, C.S. A cell-based infection assay identifies efflux pump modulators that reduce bacterial intracellular load. PLoS Pathog. 2018, 14, e1007115. [Google Scholar] [CrossRef]
- Bakowski, M.A.; Braun, V.; Brumell, J.H. Salmonella-containing vacuoles: Directing traffic and nesting to grow. Traffic 2008, 9, 2022–2031. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.M.; Schuster, M.; Rumbaugh, K.P. Working together for the common good: Cell-cell communication in bacteria. J. Bacteriol. 2012, 194, 2131–2141. [Google Scholar] [CrossRef] [PubMed]
- Helmy Yosra, A.; Kathayat, D.; Deblais, L.; Srivastava, V.; Closs, G.; Tokarski Robert, J.; Ayinde, O.; Fuchs James, R.; Rajashekara, G. Evaluation of Novel Quorum Sensing Inhibitors Targeting Auto-Inducer 2 (AI-2) for the Control of Avian Pathogenic Escherichia coli Infections in Chickens. Microbiol. Spectr. 2022, 10, e0028622. [Google Scholar] [CrossRef] [PubMed]
- Linciano, P.; Cavalloro, V.; Martino, E.; Kirchmair, J.; Listro, R.; Rossi, D.; Collina, S. Tackling antimicrobial resistance with small molecules targeting LsrK: Challenges and opportunities. J. Med. Chem. 2020, 63, 15243–15257. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Deblais, L.; Kassem, I.I.; Kathayat, D.; Rajashekara, G. Novel small molecule modulators of quorum sensing in avian pathogenic Escherichia coli (APEC). Virulence 2018, 9, 1640–1657. [Google Scholar] [CrossRef]
- Winzer, K.; Hardie, K.R.; Williams, P. LuxS and autoinducer-2: Their contribution to quorum sensing and metabolism in bacteria. Adv. Appl. Microbiol. 2003, 53, 291–396. [Google Scholar] [CrossRef] [PubMed]
- Sholpan, A.; Lamas, A.; Cepeda, A.; Franco, C.M. Salmonella spp. quorum sensing: An overview from environmental persistence to host cell invasion. AIMS Microbiol. 2021, 7, 238–256. [Google Scholar] [CrossRef]
- Smith, D.; Wang, J.-H.; Swatton, J.E.; Davenport, P.; Price, B.; Mikkelsen, H.; Stickland, H.; Nishikawa, K.; Gardiol, N.; Spring, D.R. Variations on a theme: Diverse N-acyl homoserine lactone-mediated quorum sensing mechanisms in gram-negative bacteria. Sci. Prog. 2006, 89, 167–211. [Google Scholar] [CrossRef]
- Johnson Jeremiah, G.; Yuhas, C.; McQuade Thomas, J.; Larsen Martha, J.; DiRita Victor, J. Narrow-Spectrum Inhibitors of Campylobacter jejuni Flagellar Expression and Growth. Antimicrob. Agents Chemother. 2015, 59, 3880–3886. [Google Scholar] [CrossRef]
- Choi, J.; Shin, D.; Ryu, S. Implication of quorum sensing in Salmonella enterica serovar Typhimurium virulence: The luxS gene is necessary for expression of genes in pathogenicity island 1. Infect. Immun. 2007, 75, 4885–4890. [Google Scholar] [CrossRef] [PubMed]
- Deblais, L.; Helmy, Y.A.; Kumar, A.; Antwi, J.; Kathayat, D.; Acuna, U.M.; Huang, H.-c.; de Blanco, E.C.; Fuchs, J.R.; Rajashekara, G. Novel narrow spectrum benzyl thiophene sulfonamide derivatives to control Campylobacter. J. Antibiot. 2019, 72, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Shank, J.M.; Johnson, J.G. Current and Potential Treatments for Reducing Campylobacter Colonization in Animal Hosts and Disease in Humans. Front. Microbiol. 2017, 8, 487. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Kathayat, D.; Closs, G., Jr.; Galgozy, K.; Fuchs, J.R.; Rajashekara, G. Efficacy of quorum sensing and growth inhibitors alone and in combination against avian pathogenic Escherichia coli infection in chickens. Poult. Sci. 2023, 102, 102543. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yan, C.; Xu, Y.; Feng, Y.; Wu, Q.; Lv, X.; Yang, B.; Wang, X.; Xia, X. Punicalagin inhibits Salmonella virulence factors and has anti-quorum-sensing potential. Appl. Environ. Microbiol. 2014, 80, 6204–6211. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, I.; Upadhyay, A.; Kollanoor-Johny, A.; Darre, M.J.; Venkitanarayanan, K. Effect of plant derived antimicrobials on Salmonella enteritidis adhesion to and invasion of primary chicken oviduct epithelial cells in vitro and virulence gene expression. Int. J. Mol. Sci. 2013, 14, 10608–10625. [Google Scholar] [CrossRef]
- Janssens, J.C.; Steenackers, H.; Robijns, S.; Gellens, E.; Levin, J.; Zhao, H.; Hermans, K.; De Coster, D.; Verhoeven, T.L.; Marchal, K.; et al. Brominated furanones inhibit biofilm formation by Salmonella enterica serovar Typhimurium. Appl. Environ. Microbiol. 2008, 74, 6639–6648. [Google Scholar] [CrossRef] [PubMed]
- Birhanu, B.T.; Park, N.-H.; Lee, S.-J.; Hossain, M.A.; Park, S.-C. Inhibition of Salmonella Typhimurium adhesion, invasion, and intracellular survival via treatment with methyl gallate alone and in combination with marbofloxacin. Vet. Res. 2018, 49, 101. [Google Scholar] [CrossRef]
- Styles, M.J.; Blackwell, H.E. Non-native autoinducer analogs capable of modulating the SdiA quorum sensing receptor in Salmonella enterica serovar Typhimurium. Beilstein J. Org. Chem. 2018, 14, 2651–2664. [Google Scholar] [CrossRef]
- Nesterenko, L.N.; Zigangirova, N.A.; Zayakin, E.S.; Luyksaar, S.I.; Kobets, N.V.; Balunets, D.V.; Shabalina, L.A.; Bolshakova, T.N.; Dobrynina, O.Y.; Gintsburg, A.L. A small-molecule compound belonging to a class of 2,4-disubstituted 1,3,4-thiadiazine-5-ones suppresses Salmonella infection in vivo. J. Antibiot. 2016, 69, 422–427. [Google Scholar] [CrossRef]
- Song, Y.; Xu, G.; Li, C.; Li, Z.; Lu, C.; Shen, Y. Structural optimization of natural product fusaric acid to discover novel T3SS inhibitors of Salmonella. Biochem. Biophys. Res. Commun. 2021, 582, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Hudson Debra, L.; Layton Abigail, N.; Field Terry, R.; Bowen Alison, J.; Wolf-Watz, H.; Elofsson, M.; Stevens Mark, P.; Galyov Edouard, E. Inhibition of Type III Secretion in Salmonella enterica Serovar Typhimurium by Small-Molecule Inhibitors. Antimicrob. Agents Chemother. 2007, 51, 2631–2635. [Google Scholar] [CrossRef] [PubMed]
- Defoirdt, T. Quorum-sensing systems as targets for antivirulence therapy. Trends Microbiol. 2018, 26, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Krzyżek, P. Challenges and Limitations of Anti-quorum Sensing Therapies. Front. Microbiol. 2019, 10, 2473. [Google Scholar] [CrossRef] [PubMed]
- Mechesso, A.F.; Yixian, Q.; Park, S.C. Methyl gallate and tylosin synergistically reduce the membrane integrity and intracellular survival of Salmonella Typhimurium. PLoS ONE 2019, 14, e0221386. [Google Scholar] [CrossRef] [PubMed]
- Aswathanarayan, J.B.; Vittal, R.R. Inhibition of biofilm formation and quorum sensing mediated phenotypes by berberine in Pseudomonas aeruginosa and Salmonella typhimurium. RSC Adv. 2018, 8, 36133–36141. [Google Scholar] [CrossRef]
- Sivasankar, C.; Jha, N.K.; Ghosh, R.; Shetty, P.H. Anti quorum sensing and anti virulence activity of tannic acid and it’s potential to breach resistance in Salmonella enterica Typhi/Paratyphi A clinical isolates. Microb. Pathog. 2020, 138, 103813. [Google Scholar] [CrossRef]
- Durães, F.; Resende, D.; Palmeira, A.; Szemerédi, N.; Pinto, M.M.M.; Spengler, G.; Sousa, E. Xanthones Active against Multidrug Resistance and Virulence Mechanisms of Bacteria. Antibiotics 2021, 10, 600. [Google Scholar] [CrossRef]
- Janssens Joost, C.A.; Metzger, K.; Daniels, R.; Ptacek, D.; Verhoeven, T.; Habel Lothar, W.; Vanderleyden, J.; De Vos Dirk, E.; De Keersmaecker Sigrid, C.J. Synthesis of N-Acyl Homoserine Lactone Analogues Reveals Strong Activators of SdiA, the Salmonella enterica Serovar Typhimurium LuxR Homologue. Appl. Environ. Microbiol. 2007, 73, 535–544. [Google Scholar] [CrossRef]
- Tsai, C.N.; MacNair, C.R.; Cao, M.P.T.; Perry, J.N.; Magolan, J.; Brown, E.D.; Coombes, B.K. Targeting Two-Component Systems Uncovers a Small-Molecule Inhibitor of Salmonella Virulence. Cell Chem. Biol. 2020, 27, 793–805.E7. [Google Scholar] [CrossRef]
- Li, J.; Lv, C.; Sun, W.; Li, Z.; Han, X.; Li, Y.; Shen, Y. Cytosporone B, an Inhibitor of the Type III Secretion System of Salmonella enterica Serovar Typhimurium. Antimicrob. Agents Chemother. 2013, 57, 2191–2198. [Google Scholar] [CrossRef] [PubMed]
- CDC. Immunization: The Basics. Available online: https://www.cdc.gov/vaccines/vac-gen/imz-basics.htm (accessed on 14 June 2023).
- Plotkin, S.A. Updates on immunologic correlates of vaccine-induced protection. Vaccine 2020, 38, 2250–2257. [Google Scholar] [CrossRef] [PubMed]
- Allen, H.K.; Levine, U.Y.; Looft, T.; Bandrick, M.; Casey, T.A. Treatment, promotion, commotion: Antibiotic alternatives in food-producing animals. Trends Microbiol. 2013, 21, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Elaish, M.; Ngunjiri, J.M.; Ali, A.; Xia, M.; Ibrahim, M.; Jang, H.; Hiremath, J.; Dhakal, S.; Helmy, Y.A.; Jiang, X. Supplementation of inactivated influenza vaccine with norovirus P particle-M2e chimeric vaccine enhances protection against heterologous virus challenge in chickens. PLoS ONE 2017, 12, e0171174. [Google Scholar] [CrossRef] [PubMed]
- Fawzy, M.; Helmy, Y.A. The One Health Approach is Necessary for the Control of Rift Valley Fever Infections in Egypt: A Comprehensive Review. Viruses 2019, 11, 139. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Fawzy, M.; Elaswad, A.; Sobieh, A.; Kenney, S.P.; Shehata, A.A. The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and Control. J. Clin. Med. 2020, 9, 1225. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Singh, M.; Sharif, S.; Sharma, S.; Kulkarni, R.R.; Alizadeh, M.; Yitbarek, A.; Helmy, Y.A. Intervention Strategies to Control Campylobacter at Different Stages of the Food Chain. Microorganisms 2023, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Lucero, M.S.; Chimeno Zoth, S.; Jaton, J.; Gravisaco, M.J.; Pinto, S.; Richetta, M.; Berinstein, A.; Gómez, E. Oral Immunization With Plant-Based Vaccine Induces a Protective Response Against Infectious Bursal Disease. Front. Plant Sci. 2021, 12, 741469. [Google Scholar] [CrossRef]
- Yadav, D.K.; Yadav, N.; Khurana, S.M.P. Vaccines: Present status and applications. In Animal Biotechnology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 523–542. [Google Scholar]
- Tumpey, T.M.; Alvarez, R.; Swayne, D.E.; Suarez, D.L. Diagnostic approach for differentiating infected from vaccinated poultry on the basis of antibodies to NS1, the nonstructural protein of influenza A virus. J. Clin. Microbiol. 2005, 43, 676–683. [Google Scholar] [CrossRef]
- Francis, M.J. Recent Advances in Vaccine Technologies. Vet. Clin. N. Am. Small Anim. Pract. 2018, 48, 231–241. [Google Scholar] [CrossRef]
- Vanderslott, S.; Dattani, S.; Spooner, F.; Roser, M. Vaccination; Our World Data: Oxford, UK, 2013. [Google Scholar]
- Parham, P. The Immune System: Fifth International Student Edition with Registration Card; WW Norton & Company: New York, NY, USA, 2021. [Google Scholar]
- Lidder, P.; Sonnino, A. Biotechnologies for the management of genetic resources for food and agriculture. Adv. Genet. 2012, 78, 1–167. [Google Scholar] [PubMed]
- Micoli, F.; MacLennan, C.A. Outer membrane vesicle vaccines. In Proceedings of the Seminars in Immunology; 2020; p. 101433. [Google Scholar]
- Rappuoli, R. Glycoconjugate vaccines: Principles and mechanisms. Sci. Transl. Med. 2018, 10, eaat4615. [Google Scholar] [CrossRef] [PubMed]
- Clarke, E.T.; Williams, N.A.; Dull, P.M.; Findlow, J.; Borrow, R.; Finn, A.; Heyderman, R.S. Polysaccharide–protein conjugate vaccination induces antibody production but not sustained B-cell memory in the human nasopharyngeal mucosa. Mucosal Immunol. 2013, 6, 288–296. [Google Scholar] [CrossRef]
- Ura, T.; Okuda, K.; Shimada, M. Developments in viral vector-based vaccines. Vaccines 2014, 2, 624–641. [Google Scholar] [CrossRef] [PubMed]
- Gheibi Hayat, S.M.; Darroudi, M. Nanovaccine: A novel approach in immunization. J. Cell. Physiol. 2019, 234, 12530–12536. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, C.A.; Martin, L.B.; Micoli, F. Vaccines against invasive Salmonella disease: Current status and future directions. Hum. Vaccines Immunother. 2014, 10, 1478–1493. [Google Scholar] [CrossRef] [PubMed]
- Germanier, R.; Fiirer, E. Isolation and characterization of Gal E mutant Ty 21a of Salmonella typhi: A candidate strain for a live, oral typhoid vaccine. J. Infect. Dis. 1975, 131, 553–558. [Google Scholar] [CrossRef]
- Levine, M.M.; Ferreccio, C.; Black, R.E.; Lagos, R.; Martin, O.S.; Blackwelder, W.C. Ty21a live oral typhoid vaccine and prevention of paratyphoid fever caused by Salmonella enterica Serovar Paratyphi B. Clin. Infect. Dis. 2007, 45, S24–S28. [Google Scholar] [CrossRef]
- Tagliabue, A.; Villa, L.; De Magistris, M.T.; Romano, M.; Silvestri, S.; Boraschi, D.; Nencioni, L. IgA-driven T cell-mediated anti-bacterial immunity in man after live oral Ty 21a vaccine. J. Immunol. 1986, 137, 1504–1510. [Google Scholar] [CrossRef]
- Lindow, J.C.; Fimlaid, K.A.; Bunn, J.Y.; Kirkpatrick, B.D. Antibodies in action: Role of human opsonins in killing Salmonella enterica serovar Typhi. Infect. Immun. 2011, 79, 3188–3194. [Google Scholar] [CrossRef]
- Wahid, R.; Simon, R.; Zafar, S.J.; Levine, M.M.; Sztein, M.B. Live oral typhoid vaccine Ty21a induces cross-reactive humoral immune responses against Salmonella enterica serovar Paratyphi A and S. Paratyphi B in humans. Clin. Vaccine Immunol. 2012, 19, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Chinnasami, B.; Sadasivam, K.; Vivekanandhan, A.; Arunachalam, P.; Pasupathy, S. A study on longevity of immune response after vaccination with Salmonella Typhi Vi conjugate vaccine (Pedatyph™) in children. J. Clin. Diagn. Res. 2015, 9, SC01. [Google Scholar] [CrossRef]
- Crump, J.A.; Oo, W.T. Salmonella Typhi Vi polysaccharide conjugate vaccine protects infants and children against typhoid fever. Lancet 2021, 398, 643–644. [Google Scholar] [CrossRef] [PubMed]
- Jossi, S.E.; Arcuri, M.; Alshayea, A.; Persaud, R.R.; Marcial-Juárez, E.; Palmieri, E.; Di Benedetto, R.; Pérez-Toledo, M.; Pillaye, J.; Channell, W.M.; et al. Vi polysaccharide and conjugated vaccines afford similar early, IgM or IgG-independent control of infection but boosting with conjugated Vi vaccines sustains the efficacy of immune responses. Front. Immunol. 2023, 14, 1139329. [Google Scholar] [CrossRef]
- Micoli, F.; Rondini, S.; Pisoni, I.; Giannelli, C.; Di Cioccio, V.; Costantino, P.; Saul, A.; Martin, L. Production of a conjugate vaccine for Salmonella enterica serovar Typhi from Citrobacter Vi. Vaccine 2012, 30, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Lyon, C.E.; Sadigh, K.S.; Carmolli, M.P.; Harro, C.; Sheldon, E.; Lindow, J.C.; Larsson, C.J.; Martinez, T.; Feller, A.; Ventrone, C.H.; et al. In a randomized, double-blinded, placebo-controlled trial, the single oral dose typhoid vaccine, M01ZH09, is safe and immunogenic at doses up to 1.7×1010 colony-forming units. Vaccine 2010, 28, 3602–3608. [Google Scholar] [CrossRef]
- Smith, G.W.; Smith, F.; Zuidhof, S.; Foster, D.M. Short communication: Characterization of the serologic response induced by vaccination of late-gestation cows with a Salmonella Dublin vaccine. J. Dairy Sci. 2015, 98, 2529–2532. [Google Scholar] [CrossRef]
- Renu, S.; Han, Y.; Dhakal, S.; Lakshmanappa, Y.S.; Ghimire, S.; Feliciano-Ruiz, N.; Senapati, S.; Narasimhan, B.; Selvaraj, R.; Renukaradhya, G.J. Chitosan-adjuvanted Salmonella subunit nanoparticle vaccine for poultry delivered through drinking water and feed. Carbohydr. Polym. 2020, 243, 116434. [Google Scholar] [CrossRef]
- Muniz, E.C.; Verdi, R.; Leão, J.A.; Back, A.; Nascimento, V.P.d. Evaluation of the effectiveness and safety of a genetically modified live vaccine in broilers challenged with Salmonella Heidelberg. Avian Pathol. 2017, 46, 676–682. [Google Scholar] [CrossRef]
- Crouch, C.F.; Nell, T.; Reijnders, M.; Donkers, T.; Pugh, C.; Patel, A.; Davis, P.; van Hulten, M.C.W.; de Vries, S.P.W. Safety and efficacy of a novel inactivated trivalent Salmonella enterica vaccine in chickens. Vaccine 2020, 38, 6741–6750. [Google Scholar] [CrossRef]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Moyle, P.M.; Toth, I. Modern subunit vaccines: Development, components, and research opportunities. ChemMedChem 2013, 8, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Tekle, Y.I.; Nielsen, K.M.; Liu, J.; Pettigrew, M.M.; Meyers, L.A.; Galvani, A.P.; Townsend, J.P. Controlling antimicrobial resistance through targeted, vaccine-induced replacement of strains. PLoS ONE 2012, 7, e50688. [Google Scholar] [CrossRef] [PubMed]
- Huberman, Y.D.; Velilla, A.V.; Terzolo, H.R. Evaluation of different live Salmonella enteritidis vaccine schedules administered during layer hen rearing to reduce excretion, organ colonization, and egg contamination. Poult. Sci. 2019, 98, 2422–2431. [Google Scholar] [CrossRef] [PubMed]
- Baranyi, J.; Roberts, T.A. A dynamic approach to predicting bacterial growth in food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef]
- Theron, M.M.; Lues, J.F.R. Organic Acids and Meat Preservation: A Review. Food Rev. Int. 2007, 23, 141–158. [Google Scholar] [CrossRef]
- Taylor, M.; Joerger, R.; Palou, E.; López-Malo, A.; Avila-Sosa, R.; Calix-Lara, T. Alternatives to traditional antimicrobials for organically processed meat and poultry. Org. Meat Prod. Process. 2012, 1, 211–237. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.-J. Antibacterial free fatty acids and monoglycerides: Biological activities, experimental testing, and therapeutic applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef]
- Gómez-García, M.; Sol, C.; de Nova, P.J.G.; Puyalto, M.; Mesas, L.; Puente, H.; Mencía-Ares, Ó.; Miranda, R.; Argüello, H.; Rubio, P.; et al. Antimicrobial activity of a selection of organic acids, their salts and essential oils against swine enteropathogenic bacteria. Porc. Health Manag. 2019, 5, 32. [Google Scholar] [CrossRef]
- Ng, W.-K.; Koh, C.-B. The utilization and mode of action of organic acids in the feeds of cultured aquatic animals. Rev. Aquac. 2017, 9, 342–368. [Google Scholar] [CrossRef]
- Ricke, S.C. Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult. Sci. 2003, 82, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Dittoe, D.K.; Ricke, S.C.; Kiess, A.S. Organic Acids and Potential for Modifying the Avian Gastrointestinal Tract and Reducing Pathogens and Disease. Front. Vet. Sci. 2018, 5, 216. [Google Scholar] [CrossRef]
- Ricke, S.C.; Dittoe, D.K.; Richardson, K.E. Formic Acid as an Antimicrobial for Poultry Production: A Review. Front. Vet. Sci. 2020, 7, 563. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef]
- Hensel, M. Evolution of pathogenicity islands of Salmonella enterica. Int. J. Med. Microbiol. 2004, 294, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Sprong, R.C.; Hulstein, M.F.; Van der Meer, R. Bactericidal activities of milk lipids. Antimicrob. Agents Chemother. 2001, 45, 1298–1301. [Google Scholar] [CrossRef]
- Immerseel, F.V.; Buck, J.D.; Smet, I.D.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R. Interactions of butyric acid–and acetic acid–treated Salmonella with chicken primary Cecal epithelial cells in vitro. Avian Dis. 2004, 48, 384–391. [Google Scholar] [CrossRef]
- Durant, J.A.; Corrier, D.E.; Ricke, S.C. Short-chain volatile fatty acids modulate the expression of the hilA and invF genes of Salmonella typhimurium. J. Food Prot. 2000, 63, 573–578. [Google Scholar] [CrossRef]
- Koyuncu, S.; Andersson, M.G.; Löfström, C.; Skandamis, P.N.; Gounadaki, A.; Zentek, J.; Häggblom, P. Organic acids for control of Salmonella in different feed materials. BMC Vet. Res. 2013, 9, 81. [Google Scholar] [CrossRef]
- Menconi, A.; Reginatto, A.; Londero, A.; Pumford, N.; Morgan, M.; Hargis, B.; Tellez, G. Effect of organic acids on Salmonella Typhimurium infection in broiler chickens. Int. J. Poult. Sci. 2013, 12, 72–75. [Google Scholar] [CrossRef]
- Ruhnke, I.; Röhe, I.; Goodarzi Boroojeni, F.; Knorr, F.; Mader, A.; Hafeez, A.; Zentek, J. Feed supplemented with organic acids does not affect starch digestibility, nor intestinal absorptive or secretory function in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2015, 99, 29–35. [Google Scholar] [CrossRef]
- Lynch, H.; Leonard, F.C.; Walia, K.; Lawlor, P.G.; Duffy, G.; Fanning, S.; Markey, B.K.; Brady, C.; Gardiner, G.E.; Argüello, H. Investigation of in-feed organic acids as a low cost strategy to combat Salmonella in grower pigs. Prev. Vet. Med. 2017, 139, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-C.; Hsih, H.-Y.; Chiu, H.-H.; Lai, Y.-Y.; Liu, J.-H.; Yu, B.; Tsen, H.-Y. Antagonistic activity against Salmonella infection in vitro and in vivo for two Lactobacillus strains from swine and poultry. Int. J. Food Microbiol. 2005, 102, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Gunal, M.; Yayli, G.; Kaya, O.; Karahan, N.; Sulak, O. The effects of antibiotic growth promoter, probiotic or organic acid supplementation on performance, intestinal microflora and tissue of broilers. Int. J. Poult. Sci. 2006, 5, 149–155. [Google Scholar]
- Warnecke, T.; Gill, R.T. Organic acid toxicity, tolerance, and production in Escherichia coli biorefining applications. Microb. Cell Factories 2005, 4, 25. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.H.; Iqbal, J. Recent advances in the role of organic acids in poultry nutrition. J. Appl. Anim. Res. 2016, 44, 359–369. [Google Scholar] [CrossRef]
- Brah, A.S.; Armah, F.A.; Obuah, C.; Akwetey, S.A.; Adokoh, C.K. Toxicity and therapeutic applications of citrus essential oils (CEOs): A review. Int. J. Food Prop. 2023, 26, 301–326. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Angioni, A.; Barra, A.; Coroneo, V.; Dessi, S.; Cabras, P. Chemical composition, seasonal variability, and antifungal activity of Lavandula stoechas L. ssp. stoechas essential oils from stem/leaves and flowers. J. Agric. Food Chem. 2006, 54, 4364–4370. [Google Scholar] [CrossRef]
- Inouye, S.; Abe, S.; Yamaguchi, H.; Asakura, M. Comparative study of antimicrobial and cytotoxic effects of selected essential oils by gaseous and solution contacts. Int. J. Aromather. 2003, 13, 33–41. [Google Scholar] [CrossRef]
- Costa, D.C.; Costa, H.; Albuquerque, T.G.; Ramos, F.; Castilho, M.C.; Sanches-Silva, A. Advances in phenolic compounds analysis of aromatic plants and their potential applications. Trends Food Sci. Technol. 2015, 45, 336–354. [Google Scholar] [CrossRef]
- Lubbe, A.; Verpoorte, R. Cultivation of medicinal and aromatic plants for specialty industrial materials. Ind. Crops. Prod. 2011, 34, 785–801. [Google Scholar] [CrossRef]
- Bansal, T. Benefits of essential oil. J. Chem. Pharm. Res. 2016, 8, 143–149. [Google Scholar]
- O’Brien, T.F. Emergence, spread, and environmental effect of antimicrobial resistance: How use of an antimicrobial anywhere can increase resistance to any antimicrobial anywhere else. Clin. Infect. Dis. 2002, 34, S78–S84. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial Properties of Plant Essential Oils against Human Pathogens and Their Mode of Action: An Updated Review. Evid.-Based Complement. Altern. Med. 2016, 2016, 3012462. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B.; Kang, J.H.; Song, K.B. Antibacterial activities of a cinnamon essential oil with cetylpyridinium chloride emulsion against Escherichia coli O157:H7 and Salmonella Typhimurium in basil leaves. Food Sci. Biotechnol. 2018, 27, 47–55. [Google Scholar] [CrossRef]
- Raybaudi-Massilia, R.M.; Mosqueda-Melgar, J.; Martin-Belloso, O. Antimicrobial Activity of Essential Oils on Salmonella Enteritidis, Escherichia coli, and Listeria innocua in Fruit Juices. J. Food Prot. 2006, 69, 1579–1586. [Google Scholar] [CrossRef]
- Ebani, V.V.; Nardoni, S.; Bertelloni, F.; Tosi, G.; Massi, P.; Pistelli, L.; Mancianti, F. In Vitro Antimicrobial Activity of Essential Oils Against Salmonella enterica Serotypes Enteritidis and Typhimurium Strains Isolated from Poultry. Molecules 2019, 24, 900. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Al-Holy, M.A.; Abu Ghoush, M.H.; Al-Nabulsi, A.A.; Osaili, T.M.; Holley, R.A. Inhibitory effects of cinnamon and thyme essential oils against Salmonella spp. in hummus (chickpea dip). J. Food Process. Preserv. 2019, 43, e13925. [Google Scholar] [CrossRef]
- Thanissery, R.; Kathariou, S.; Smith, D.P. Rosemary oil, clove oil, and a mix of thyme-orange essential oils inhibit Salmonella and Campylobacter in vitro. J. Appl. Poult. Res. 2014, 23, 221–227. [Google Scholar] [CrossRef]
- Puškárová, A.; Bučková, M.; Kraková, L.; Pangallo, D.; Kozics, K. The antibacterial and antifungal activity of six essential oils and their cyto/genotoxicity to human HEL 12469 cells. Sci. Rep. 2017, 7, 8211. [Google Scholar] [CrossRef] [PubMed]
- Slameňová, D.; Horváthová, E.; Kováčiková, Z.; Kozics, K.; Hunáková, Ľ. Essential rosemary oil protects testicular cells against DNA-damaging effects of H2O2 and DMNQ. Food Chem. 2011, 129, 64–70. [Google Scholar] [CrossRef]
- Maurya, A.; Prasad, J.; Das, S.; Dwivedy, A.K. Essential Oils and Their Application in Food Safety. Front. Sustain. Food Syst. 2021, 5, 133. [Google Scholar] [CrossRef]
- Millet, Y.; Jouglard, J.; Steinmetz, M.; Tognetti, P.; Joanny, P.; Arditti, J. Toxicity of some essential plant oils. Clinical and experimental study. Clin. Toxicol. 1981, 18, 1485–1498. [Google Scholar] [CrossRef]
- Posadzki, P.; Alotaibi, A.; Ernst, E. Adverse effects of aromatherapy: A systematic review of case reports and case series. Int. J. Risk Saf. Med. 2012, 24, 147–161. [Google Scholar] [CrossRef]
- Fadil, M.; Fikri-Benbrahim, K.; Rachiq, S.; Ihssane, B.; Lebrazi, S.; Chraibi, M.; Haloui, T.; Farah, A. Combined treatment of Thymus vulgaris L., Rosmarinus officinalis L. and Myrtus communis L. essential oils against Salmonella typhimurium: Optimization of antibacterial activity by mixture design methodology. Eur. J. Pharm. Biopharm. 2018, 126, 211–220. [Google Scholar] [CrossRef]
- Barbosa, L.N.; Alves, F.C.B.; Andrade, B.F.M.T.; Albano, M.; Rall, V.L.M.; Fernandes, A.A.H.; Buzalaf, M.A.R.; Leite, A.d.L.; de Pontes, L.G.; dos Santos, L.D.; et al. Proteomic analysis and antibacterial resistance mechanisms of Salmonella Enteritidis submitted to the inhibitory effect of Origanum vulgare essential oil, thymol and carvacrol. J. Proteom. 2020, 214, 103625. [Google Scholar] [CrossRef]
- Hasheminya, S.-M.; Dehghannya, J. Composition, phenolic content, antioxidant and antimicrobial activity of Pistacia atlantica subsp. kurdica hulls’ essential oil. Food Biosci. 2020, 34, 100510. [Google Scholar] [CrossRef]
- Alibi, S.; Selma, W.B.; Ramos-Vivas, J.; Smach, M.A.; Touati, R.; Boukadida, J.; Navas, J.; Mansour, H.B. Anti-oxidant, antibacterial, anti-biofilm, and anti-quorum sensing activities of four essential oils against multidrug-resistant bacterial clinical isolates. Curr. Res. Transl. Med. 2020, 68, 59–66. [Google Scholar] [CrossRef]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Antimicrobial Activity of Basil (Ocimum basilicum) Oil against Salmonella Enteritidis in Vitro and in Food. Biosci. Biotechnol. Biochem. 2010, 74, 1200–1204. [Google Scholar] [CrossRef]
- Morshdy, A.; El-Tahlawy, A.S.; Qari, S.H.; Qumsani, A.T.; Bay, D.H.; Sami, R.; Althubaiti, E.H.; Mansour, A.M.A.; Aljahani, A.H.; Hafez, A.E.E.; et al. Anti-Biofilms’ Activity of Garlic and Thyme Essential Oils against Salmonella typhimurium. Molecules 2022, 27, 2182. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Liu, L.; Guo, F.; Huang, J.; Qiao, J.; Bi, R.; Huang, J.; Zhang, K.; Guo, Y.; Wang, Z. Dietary supplemental coated essential oils and organic acids mixture improves growth performance and gut health along with reduces Salmonella load of broiler chickens infected with Salmonella Enteritidis. J. Anim. Sci. Biotechnol. 2023, 14, 95. [Google Scholar] [CrossRef] [PubMed]
- Moore-Neibel, K.; Gerber, C.; Patel, J.; Friedman, M.; Ravishankar, S. Antimicrobial activity of lemongrass oil against Salmonella enterica on organic leafy greens. J. Appl. Microbiol. 2012, 112, 485–492. [Google Scholar] [CrossRef] [PubMed]


| Year | Number of Outbreaks | Number of Illnesses | Identified Serotypes | Source |
|---|---|---|---|---|
| 2012 | 9 | 1217 | Bredeney, Braenderup, Typhimurium, Newport, Enteritidis, Bareilly, Nchanga, Hadar, Infantis, Newport, Lille | Peanut butter, mangoes, cantaloupe, ground beef, raw scraped ground tuna product, hedgehogs, live poultry |
| 2013 | 9 | 2278 | Sandiego, Pomona, Poona, Heidelberg, Montevideo, Mbandaka, Saintpaul, Typhimurium, Infantis, Lille, and Newport | Small turtles, foster farms brand chicken, tahini sesame paste, cucumber, ground beef, live poultry |
| 2014 | 8 | 429 | Cotham, Heidelberg, Stanley, Typhimurium, Newport, Hartford, Oranienburg, and Braenderup | Bearded dragons, chicken, organic sprouted chia powder, nut butter, raw cashew cheese, frozen rodent feed, cucumbers |
| 2015 | 8 | 1512 | Enteritidis, Paratyphi B variant L (+) tartrate (+), Weltevreden, Sandiego, Poona, Hadar, Indiana, and Muenchen. | Bean sprouts, raw sprouted nut butter spreads, cucumbers, raw, frozen, stuffed chicken entrees, frozen raw tuna, live poultry, and small turtles |
| 2016 | 5 | 114 | Oranienburg, Reading, Abony, Montevideo, Senftenberg, Muenchen, Kentucky, Virchow, and Heidelberg. | Shell eggs, alfalfa sprouts, pistachios, organic shake and meal products, dairy calves, and live poultry |
| 2017 | 5 | 2171 | Agbeni, and Typhimurium | Pet turtles, live poultry, laboratory exposure. |
| 2019 | 9 | 1632 | Javiana, Dublin, Uganda, Concord, Carrau, Schwarzengrund, Oranienburg, Typhimurium | Cut fruit, ground beef, papayas, kawaran brand tahini, pre-cut melon, butterball, brand ground turkey, pet turtles, backyard poultry, and hedgehogs |
| 2020 | 8 | 3107 | Stanley, Enteritidis, Newport, Muenster, Typhimurium, Hadar | Wood ear mushrooms, peaches, onions, pet bearded dragons, pet hedgehogs, backyard poultry, and small pet turtles |
| 2021 | 10 | 2575 | Thompson, Oranienburg, Typhimurium, Weltevreden, Infantis, Enteritidis, Hadar | Seafood, pet turtles, Italian-style meats, onions, prepackaged salads, frozen cooked shrimp, raw frozen breaded stuffed chicken products, cashew brie, ground turkey, backyard poultry, wild songbirds |
| 2022 | 7 | 1469 | Typhimurium, Litchfield, Senftenberg, Stanley, and Uganda | Alfalfa sprouts, fish, peanut butter, pet bearded dragons, small turtles, poultry |
| 2023 | 8 | 1527 | Enteritidis, Thompson, Saint Paul, and Infantis | Raw cookie dough, flour, ground beef, fresh diced onion, cantaloupes, small turtles, dry dog food, and poultry |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamichhane, B.; Mawad, A.M.M.; Saleh, M.; Kelley, W.G.; Harrington, P.J., II; Lovestad, C.W.; Amezcua, J.; Sarhan, M.M.; El Zowalaty, M.E.; Ramadan, H.; et al. Salmonellosis: An Overview of Epidemiology, Pathogenesis, and Innovative Approaches to Mitigate the Antimicrobial Resistant Infections. Antibiotics 2024, 13, 76. https://doi.org/10.3390/antibiotics13010076
Lamichhane B, Mawad AMM, Saleh M, Kelley WG, Harrington PJ II, Lovestad CW, Amezcua J, Sarhan MM, El Zowalaty ME, Ramadan H, et al. Salmonellosis: An Overview of Epidemiology, Pathogenesis, and Innovative Approaches to Mitigate the Antimicrobial Resistant Infections. Antibiotics. 2024; 13(1):76. https://doi.org/10.3390/antibiotics13010076
Chicago/Turabian StyleLamichhane, Bibek, Asmaa M. M. Mawad, Mohamed Saleh, William G. Kelley, Patrick J. Harrington, II, Cayenne W. Lovestad, Jessica Amezcua, Mohamed M. Sarhan, Mohamed E. El Zowalaty, Hazem Ramadan, and et al. 2024. "Salmonellosis: An Overview of Epidemiology, Pathogenesis, and Innovative Approaches to Mitigate the Antimicrobial Resistant Infections" Antibiotics 13, no. 1: 76. https://doi.org/10.3390/antibiotics13010076
APA StyleLamichhane, B., Mawad, A. M. M., Saleh, M., Kelley, W. G., Harrington, P. J., II, Lovestad, C. W., Amezcua, J., Sarhan, M. M., El Zowalaty, M. E., Ramadan, H., Morgan, M., & Helmy, Y. A. (2024). Salmonellosis: An Overview of Epidemiology, Pathogenesis, and Innovative Approaches to Mitigate the Antimicrobial Resistant Infections. Antibiotics, 13(1), 76. https://doi.org/10.3390/antibiotics13010076