
Prevalence, Virulence, and Antimicrobial Resistance of Major Mastitis Pathogens Isolated from Taiwanese Dairy Farms



Abstract
:1. Introduction
2. Results
2.1. Identification of Bacterial Isolates through 16S rRNA Gene Sequencing
2.2. Antimicrobial Susceptibility of Streptococcus spp., Staphylococcus spp., and Coliforms
2.3. Comparative Study of Phenotypic and Genotypic Antimicrobial Resistance in Streptococcus spp., Staphylococcus spp., and Coliforms
2.4. Correlation between Antimicrobial Resistance Genes and the Virulence Factors of Bacterial Isolates
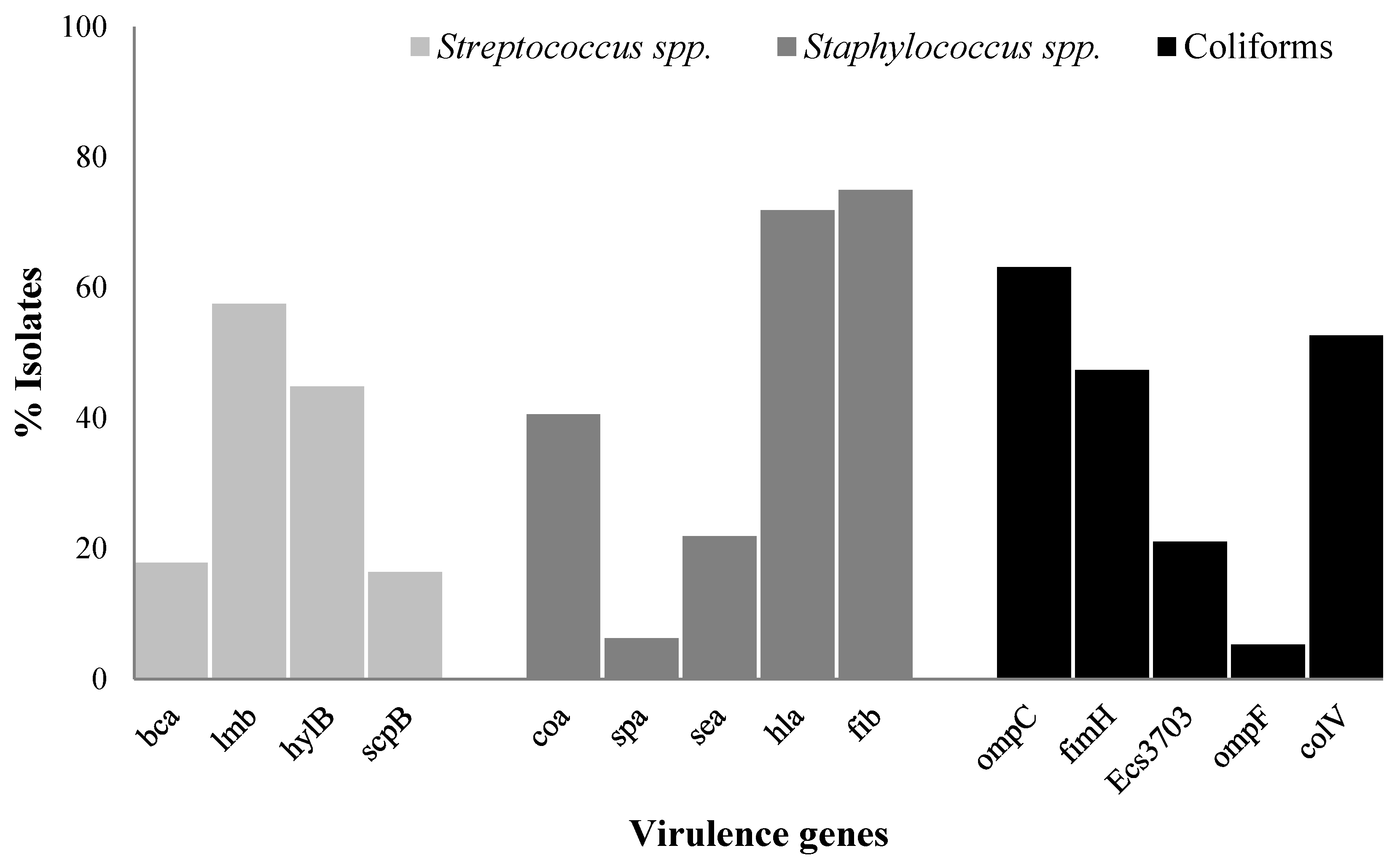
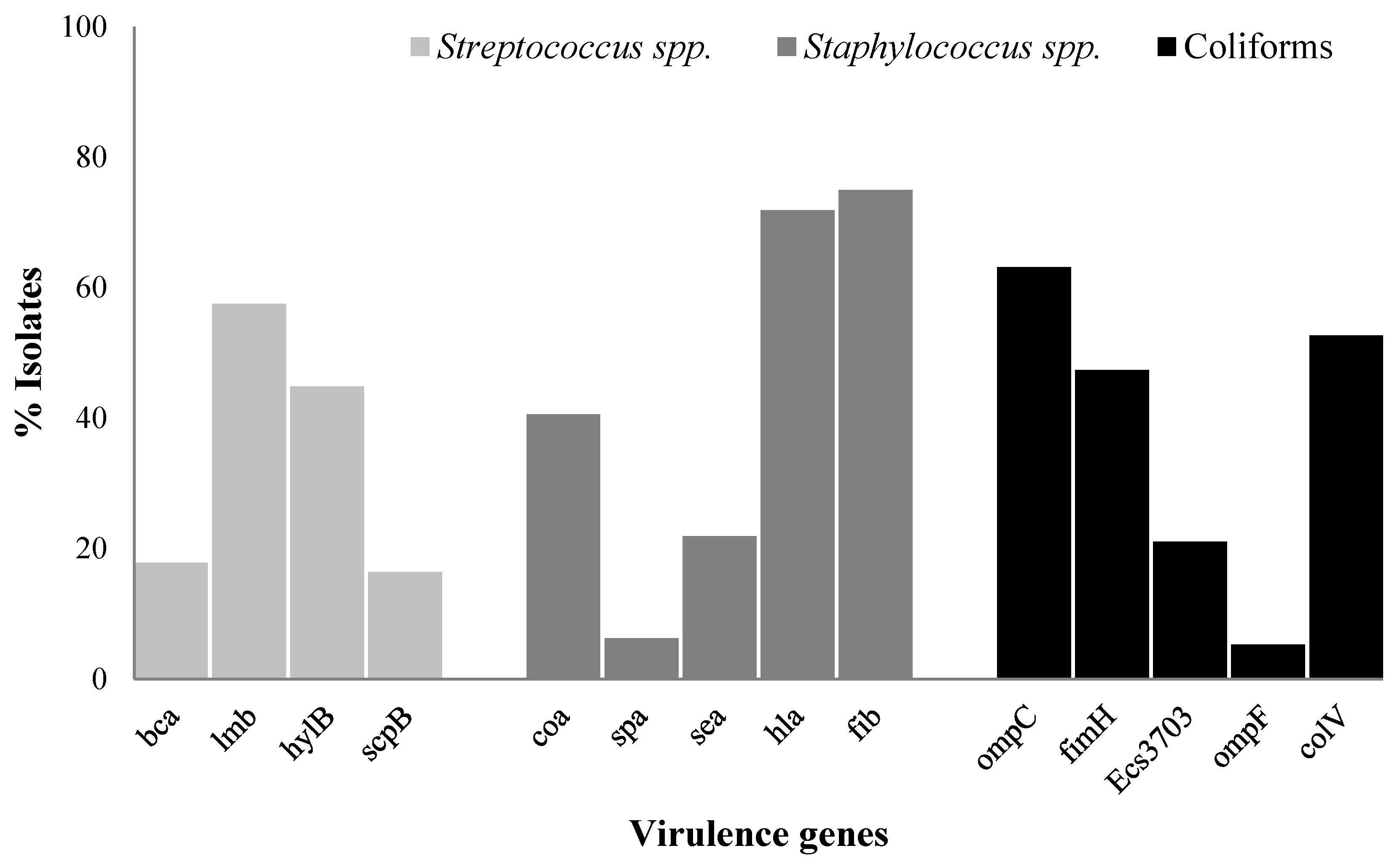
2.5. Prevalence of Virulence Genes in Streptococcus spp., Staphylococcus spp., and Coliforms
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Herd Enrollment Criteria and Milk Sample Collection
5.2. Isolation and Identification of Bacteria
5.3. Bacterial Species Identification through 16S rRNA Gene Sequencing
5.4. Antimicrobial Susceptibility Test
5.5. Identification of Antimicrobial Resistance Genes
5.6. Identification of Virulence Genes
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bradley, A. Bovine mastitis: An evolving disease. Vet. J. 2002, 164, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Barkema, H.W.; Zhang, L.; Liu, G.; Deng, Z.; Cai, L.; Shan, R.; Zhang, S.; Zou, J.; Kastelic, J.P.; et al. Incidence of clinical mastitis and distribution of pathogens on large Chinese dairy farms. J. Dairy Sci. 2017, 100, 4797–4806. [Google Scholar] [CrossRef] [PubMed]
- Sztachańska, M.; Barański, W.; Janowski, T.; Pogorzelska, J.; Zduńczyk, S. Prevalence and etiological agents of subclinical mastitis at the end of lactation in nine dairy herds in North-East Poland. Pol. J. Vet. Sci. 2016, 19, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Bergman, P.; Raqib, R.; Rekha, R.S.; Agerberth, B.; Gudmundsson, G.H. Host directed therapy against infection by boosting innate immunity. Front. Immunol. 2020, 11, 1209. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Kong, L.; Gao, H.; Cheng, X.; Wang, X. A review of current bacterial resistance to antibiotics in food animals. Front. Microbiol. 2022, 13, 822689. [Google Scholar] [CrossRef]
- WHO. Guidelines on Use of Medically Important Antimicrobials in Food-Producing Animals. 2017. Available online: http://apps.who.int/iris/bitstream/handle/10665/258970/9789241550130-eng.pdf (accessed on 12 July 2023).
- Kapoor, G.; Saigal, S.; Elongavan, A. Action and resistance mechanisms of antibiotics: A guide for clinicians. J. Anaesthesiol. Clin. Pharmacol. 2017, 33, 300–305. [Google Scholar] [CrossRef]
- McInnes, R.S.; McCallum, G.E.; Lamberte, L.E.; van Schaik, W. Horizontal transfer of antibiotic resistance genes in the human gut microbiome. Curr. Opin. Microbiol. 2020, 53, 35–43. [Google Scholar] [CrossRef]
- Monistero, V.; Barberio, A.; Biscarini, F.; Cremonesi, P.; Castiglioni, B.; Graber, H.U.; Bottini, E.; Ceballos-Marquez, A.; Kroemker, V.; Petzer, I.M.; et al. Different distribution of antimicrobial resistance genes and virulence profiles of Staphylococcus aureus strains isolated from clinical mastitis in six countries. J. Dairy Sci. 2020, 103, 3431–3446. [Google Scholar] [CrossRef]
- Patel, K.; Godden, S.M.; Royster, E.E.; Crooker, B.A.; Johnson, T.J.; Smith, E.A.; Sreevatsan, S. Prevalence, antibiotic resistance, virulence and genetic diversity of Staphylococcus aureus isolated from bulk tank milk samples of U.S. dairy herds. BMC Genom. 2021, 22, 367. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, Z.; Huang, C.; Gao, X.; Wang, Z.; Liu, Y.; Tian, C.; Hong, W.; Niu, S.; Liu, M. The phylogenetic group, antimicrobial susceptibility, and virulence genes of Escherichia coli from clinical bovine mastitis. J. Dairy Sci. 2018, 101, 572–580. [Google Scholar] [CrossRef]
- Chang, S.C.; Chen, M.W.; Lin, M.C.; Hu, O.Y.P. Antibiotic consumption in human and animals in Taiwan. Infect. Control J. 2003, 13, 334–345. [Google Scholar]
- Hsieh, J.C.; Yen, Y.S.; Chuang, S.T. Identification of Streptococcus spp. isolated from bovine milk and characterization of their antimicrobial susceptibility profiles in Taiwan. Thai J. Vet. Med. 2019, 49, 57–63. [Google Scholar] [CrossRef]
- Abureema, S.; Smooker, P.; Malmo, J.; Deighton, M. Molecular epidemiology of recurrent clinical mastitis due to Streptococcus uberis: Evidence of both an environmental source and recurring infection with the same strain. J. Dairy Sci. 2014, 97, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Monistero, V.; Graber, H.U.; Pollera, C.; Cremonesi, P.; Castiglioni, B.; Bottini, E.; Ceballos-Marquez, A.; Lasso-Rojas, L.; Kroemker, V.; Wente, N.; et al. Staphylococcus aureus isolates from bovine mastitis in eight countries: Genotypes, detection of genes encoding different toxins and other virulence genes. Toxins 2018, 10, 247. [Google Scholar] [CrossRef] [PubMed]
- Fry, P.R.; Middleton, J.R.; Dufour, S. Association of coagulase-negative staphylococcal species, mammary quarter milk somatic cell count, and persistence of intramammary infection in dairy cattle. J. Dairy Sci. 2014, 97, 4876–4885. [Google Scholar] [CrossRef] [PubMed]
- Ngu Ngwa, V.; Cuteri, V.; Awah-Ndukum, J.; Tangwa, B.V.; Manchang, K.T. Bacterial pathogens involved in bovine mastitis and their antibiotic resistance patterns in the Adamawa region of Cameroon. J. Dairy Res. Technol. 2020, 3, 012. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Zhang, J.; Yang, J.; Yi, B.; Liu, G.; Zhou, M.; Kastelic, J.P.; Han, B.; Gao, J. Klebsiella pneumoniae infection causes mitochondrial damage and dysfunction in bovine mammary epithelial cells. Vet. Res. 2021, 52, 17. [Google Scholar] [CrossRef]
- Quintana, Á.R.; Seseña, S.; Garzón, A.; Arias, R. Factors Affecting Levels of Airborne Bacteria in Dairy Farms: A Review. Animals 2020, 10, 526. [Google Scholar] [CrossRef]
- Shen, J.; Wu, X.; Yang, Y.; Lv, Y.; Li, X.; Ding, X.; Wang, S.; Yan, Z.; Yan, Y.; Yang, F.; et al. Antimicrobial Resistance and Virulence Factor of Streptococcus dysgalactiae Isolated from Clinical Bovine Mastitis Cases in Northwest China. Infect. Drug Resist. 2021, 14, 3519–3530. [Google Scholar] [CrossRef]
- Deng, J.; Liu, K.; Wang, K.; Yang, B.; Xu, H.; Wang, J.; Dai, F.; Xiao, X.; Gu, X.; Zhang, L.; et al. The prevalence of coagulase-negative staphylococcus associated with bovine mastitis in China and its antimicrobial resistance rate: A meta-analysis. J. Dairy Res. 2023, 90, 158–163. [Google Scholar] [CrossRef]
- Giguère, S. Principles of antimicrobial drug selection and Use. In Antimicrobial Therapy in Veterinary Medicine, 5th ed.; Giguère, S., Prescott, J.F., Dowling, P.M., Eds.; John Wiley & Sons: London, UK, 2013; pp. 105–115. [Google Scholar]
- Pascu, C.; Herman, V.; Iancu, I.; Costinar, L. Etiology of mastitis and antimicrobial resistance in dairy cattle farms in the western part of Romania. Antibiotics 2022, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Rana, E.A.; Fazal, M.A.; Alim, M.A. Frequently used therapeutic antimicrobials and their resistance patterns on Staphylococcus aureus and Escherichia coli in mastitis affected lactating cows. Int. J. Vet. Sci. 2022, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Qi, W.; Wang, X.; Ling, W.; Li, X.; Luo, J.; Zhang, S.; Li, H. Genetic characterization of antimicrobial resistance in Staphylococcus aureus isolated from bovine mastitis cases in Northwest China. J. Integr. Agric. 2016, 15, 2842–2847. [Google Scholar]
- Vélez, J.R.; Cameron, M.; Rodríguez-Lecompte, J.C.; Xia, F.; Heider, L.C.; Saab, M.; McClure, J.T.; Sánchez, J. Whole-genome sequence analysis of antimicrobial resistance genes in Streptococcus uberis and Streptococcus dysgalactiae isolates from Canadian Dairy Herds. Front. Vet. Sci. 2017, 4, 63. [Google Scholar] [CrossRef]
- Gao, J.; Yu, F.Q.; Luo, L.P.; He, J.Z.; Hou, R.G.; Zhang, H.Q.; Li, S.M.; Su, J.L.; Han, B. Antibiotic resistance of Streptococcus agalactiae from cows with mastitis. Vet. J. 2012, 194, 423–424. [Google Scholar] [CrossRef] [PubMed]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial resistance and virulence: A successful or deleterious association in the bacterial world? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef] [PubMed]
- Azoulay-Dupuis, E.; Rieux, V.; Muffat-Joly, M.; Bédos, J.P.; Vallée, E.; Rivier, C.; Isturiz, R.; Carbon, C.; Moine, P. Relationship between capsular type, penicillin susceptibility, and virulence of human Streptococcus pneumoniae isolates in mice. Antimicrob. Agents Chemother. 2000, 44, 1575–1577. [Google Scholar] [CrossRef]
- Linkevicius, M.; Sandegren, L.; Andersson, D.I. Mechanisms and fitness costs of tigecycline resistance in Escherichia coli. J. Antimicrob. Chemother. 2013, 68, 2809–2819. [Google Scholar] [CrossRef]
- Darmancier, H.; Domingues, C.P.F.; Rebelo, J.S.; Amaro, A.; Dionísio, F.; Pothier, J.; Serra, O.; Nogueira, T. Are virulence and antibiotic resistance genes linked? A comprehensive analysis of bacterial chromosomes and plasmids. Antibiotics 2022, 11, 706. [Google Scholar] [CrossRef]
- Ding, Y.; Zhao, J.; He, X.; Li, M.; Guan, H.; Zhang, Z.; Li, P. Antimicrobial resistance and virulence-related genes of Streptococcus obtained from dairy cows with mastitis in Inner Mongolia, China. Pharm. Biol. 2016, 54, 162–167. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, L.; Zhou, X.; He, Y.; Yong, C.; Shen, M.; Szenci, O.; Han, B. Antimicrobial susceptibility, virulence genes, and randomly amplified polymorphic DNA analysis of Staphylococcus aureus recovered from bovine mastitis in Ningxia, China. J. Dairy Sci. 2016, 99, 9560–9569. [Google Scholar] [CrossRef] [PubMed]
- Kaczorek, E.; Małaczewska, J.; Wójcik, R.; Siwicki, A.K. Biofilm production and other virulence factors in Streptococcus spp. isolated from clinical cases of bovine mastitis in Poland. BMC Vet. Res. 2017, 13, 398. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tan, X.; Zhang, X.; Xia, X.; Sun, H. The diversities of staphylococcal species, virulence and antibiotic resistance genes in the subclinical mastitis milk from a single Chinese cow herd. Microb. Pathog. 2015, 88, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Pires, R.; Rolo, D.; Gama-Norton, L. Group A Streptococci from carriage and disease in Portugal: Evolution of antimicrobial resistance and T antigenic types during 2000–2002. Microb. Drug Resist. 2005, 11, 360–370. [Google Scholar] [CrossRef]
- Meh, D.A.; Siebenlist, K.R.; Brennan, S.O.; Holyst, T.; Mosesson, M.W. The amino acid sequence in fibrin responsible for high affinity thrombin binding. Thromb. Haemost. 2001, 85, 470–474. [Google Scholar]
- Hejair, H.M.A.; Zhu, Y.; Ma, J.; Zhang, Y.; Pan, Z.; Zhang, W.; Yao, H. Functional role of ompF and ompC porins in pathogenesis of avian pathogenic Escherichia coli. Microb. Pathog. 2017, 107, 29–37. [Google Scholar] [CrossRef]
- Nossa, C.W.; Oberdorf, W.E.; Yang, L.; Aas, J.A.; Paster, B.J.; Desantis, T.Z.; Brodie, E.L.; Malamud, D.; Poles, M.A.; Pei, Z. Design of 16S rRNA gene primers for 454 pyrosequencing of the human foregut microbiome. World J. Gastroenterol. 2010, 16, 4135–4144. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals. 2018. Available online: https://clsi.org/media/2321/vet08ed4_sample.pdf (accessed on 15 July 2023).
- FDA. BD BBL™ Sensi-Disc™ Antimicrobial Susceptibility Test Discs. 2007. Available online: https://legacy.bd.com/europe/regulatory/Assets/IFU/US/8840621(1107)_No.pdf (accessed on 5 July 2023).

{kind=link}
| Genus | Species | Number (n) | Accession Number |
|---|---|---|---|
| Streptococcus (n = 73) | Strep. Uberis | 30 | NR_040820 |
| Strep. Lutetiensis | 13 | NR_037096 | |
| Strep. Dysgalactiae | 10 | NR_027517 | |
| Strep. Bovis | 9 | AJ305257 | |
| Strep. Equinus | 6 | NR_042052 | |
| Strep. Agalactiae | 5 | OP752129 | |
| Staphylococcus (n = 32) | Staph. Aureus | 14 | NR_037007 |
| Staph. Epidermidis | 7 | NR_036904 | |
| Staph. Hemolyticus | 4 | AY688062 | |
| Staph. Chromogenes | 3 | AY688044 | |
| Staph. Hyicus | 2 | NR_036905 | |
| Staph. Simulans | 1 | AY688101 | |
| Staph. Capitis | 1 | NR_027519 | |
| Coliforms (n = 19) | Escherichia coli | 8 | X80721 |
| Enterobacter aerogenes | 5 | LT221165 | |
| Klebsiella pneumoniae | 4 | NR_036794 | |
| Escherichia fergusonii | 2 | NR_027549 |
| Organism | Antimicrobial Susceptibility a | TET | NEO | BAC | AMP | OXA | CXM | CF | XNL |
|---|---|---|---|---|---|---|---|---|---|
| Streptococcus (n = 73) | Resistant (%) | 86.30 | 79.45 | 38.35 | 45.20 | 73.97 | 19.17 | 8.21 | 26.02 |
| Susceptible (%) | 13.69 | 20.54 | 61.64 | 54.79 | 26.02 | 80.82 | 91.78 | 73.97 | |
| Staphylococcus (n = 32) | Resistant (%) | 59.37 | 21.87 | 34.37 | 43.75 | 53.12 | 0.00 | 0.00 | 0.00 |
| Susceptible (%) | 40.62 | 78.12 | 65.62 | 56.25 | 46.87 | 100 | 100 | 100 | |
| Coliforms (n = 19) | Resistant (%) | 31.57 | 21.05 | 68.42 | 31.57 | 100 | 15.78 | 31.57 | 0.00 |
| Susceptible (%) | 68.42 | 78.94 | 31.57 | 68.42 | 0.00 | 84.21 | 68.42 | 100 |
| Antimicrobial Agents | Gene(s) | Characteristics of Streptococcus Isolates 1 | Association | |||||
|---|---|---|---|---|---|---|---|---|
| P+/G+ (n) | P−/G- (n) | P+/G− (n) | P−/G+ (n) | G+ (%) | r 2 | p 3 | ||
| Tetracycline | Total | 42 | 3 | 21 | 7 | 67.12 | −0.21822 | 0.0696 |
| tetM | 31 | 3 | 46.57 | |||||
| tetB | 5 | 0 | 6.84 | |||||
| tetA | 3 | 2 | 6.84 | |||||
| tetO | 3 | 2 | 6.84 | |||||
| Neomycin | Total | 39 | 10 | 19 | 5 | 60.27 | −0.19294 | 0.1298 |
| aph(3)-I | 13 | 2 | 20.54 | |||||
| aph(3)-II | 26 | 3 | 39.72 | |||||
| Bacitracin | Total | 22 | 24 | 6 | 21 | 58.90 | −0.32350 | 0.0234 * |
| bcrB | 16 | 14 | 41.09 | |||||
| bcrA | 6 | 7 | 17.80 | |||||
| Ampicillin | Total | 20 | 29 | 13 | 11 | 42.46 | −0.37388 | 0.0124 * |
| blaZ | 12 | 9 | 28.76 | |||||
| ampC | 8 | 2 | 13.69 | |||||
| Oxacillin | Total | 35 | 10 | 19 | 9 | 60.27 | −0.26827 | 0.0335 * |
| blaZ | 25 | 6 | 42.46 | |||||
| ampC | 10 | 3 | 17.80 | |||||
| Cefuroxime | Total | 9 | 42 | 5 | 17 | 35.61 | −0.48324 | 0.0059 * |
| blaZ | 6 | 16 | 30.13 | |||||
| ampC | 3 | 1 | 5.47 | |||||
| Cephalothin | Total | 2 | 55 | 4 | 12 | 19.17 | −0.75593 | 0.0003 * |
| blaZ | 2 | 12 | 19.17 | |||||
| ampC | 0 | 0 | 0.00 | |||||
| Ceftiofur | Total | 11 | 50 | 8 | 4 | 20.54 | −0.33508 | 0.1181 |
| blaZ | 7 | 2 | 12.32 | |||||
| ampC | 4 | 2 | 8.21 | |||||
| Antimicrobial Agents | Gene(s) | Characteristics of Staphylococcus Isolates 1 | Association | |||||
|---|---|---|---|---|---|---|---|---|
| P+/G+ (n) | P−/G− (n) | P+/G− (n) | P−/G+ (n) | G+ (%) | r 2 | p 3 | ||
| Tetracycline | Total | 10 | 6 | 9 | 7 | 53.12 | −0.44164 | 0.0239 * |
| tetM | 5 | 1 | 18.75 | |||||
| tetB | 0 | 2 | 6.25 | |||||
| tetO | 3 | 3 | 18.75 | |||||
| tetA | 2 | 1 | 9.37 | |||||
| Neomycin | Total | 5 | 17 | 2 | 8 | 40.62 | −0.41931 | 0.1197 |
| aph(3)-I | 2 | 3 | 15.62 | |||||
| aph(3)-II | 3 | 5 | 25.00 | |||||
| Bacitracin | Total | 10 | 13 | 1 | 8 | 56.25 | −0.20101 | 0.4093 |
| bcrB | 7 | 6 | 40.62 | |||||
| bcrA | 3 | 2 | 15.62 | |||||
| Ampicillin | Total | 9 | 13 | 5 | 5 | 43.75 | −0.35714 | 0.1333 |
| blaZ | 5 | 3 | 25.00 | |||||
| ampC | 1 | 0 | 3.12 | |||||
| mecA | 3 | 2 | 15.62 | |||||
| Oxacillin | Total | 7 | 13 | 10 | 2 | 28.12 | −0.36155 | 0.1283 |
| blaZ | 4 | 2 | 18.75 | |||||
| ampC | 1 | 0 | 3.12 | |||||
| mecA | 2 | 0 | 6.25 | |||||
| Cefuroxime | Total | 0 | 31 | 0 | 1 | 3.12 | NT | NT |
| blaZ | 0 | 1 | 3.12 | |||||
| ampC | 0 | 0 | 0.00 | |||||
| mecA | 0 | 0 | 0.00 | |||||
| Cephalothin | Total | 0 | 31 | 0 | 1 | 3.12 | NT | NT |
| blaZ | 0 | 1 | 3.12 | |||||
| ampC | 0 | 0 | 0.00 | |||||
| mecA | 0 | 0 | 0.00 | |||||
| Ceftiofur | Total | 0 | 31 | 0 | 1 | 3.12 | NT | NT |
| blaZ | 0 | 1 | 3.12 | |||||
| ampC | 0 | 0 | 0.00 | |||||
| mecA | 0 | 0 | 0.00 | |||||
| Antimicrobial Agents | Gene(s) | Characteristics of Coliforms Isolates 1 | Association | |||||
|---|---|---|---|---|---|---|---|---|
| P+/G+ (n) | P-/G- (n) | P+/G- (n) | P-/G+ (n) | G+ (%) | r 2 | p 3 | ||
| Tetracycline | Total | 5 | 5 | 1 | 8 | 68.42 | −0.32026 | 0.2643 |
| tetM | 4 | 4 | 42.10 | |||||
| tetO | 1 | 0 | 5.26 | |||||
| tetB | 0 | 2 | 10.52 | |||||
| tetA | 0 | 2 | 10.52 | |||||
| Neomycin | Total | 4 | 8 | 0 | 7 | 57.89 | NT | NT |
| aph(3)-I | 1 | 3 | 21.05 | |||||
| aph(3)-II | 3 | 4 | 36.84 | |||||
| Bacitracin | Total | 9 | 4 | 4 | 2 | 57.89 | −0.23652 | 0.3960 |
| bcrB | 9 | 2 | 57.89 | |||||
| bcrA | 0 | 0 | 0.00 | |||||
| Ampicillin | Total | 4 | 8 | 2 | 5 | 47.36 | −0.43033 | 0.1864 |
| blaZ | 4 | 5 | 47.36 | |||||
| ampC | 0 | 0 | 0.00 | |||||
| Oxacillin | Total | 7 | 0 | 12 | 0 | 36.84 | NT | NT |
| blaZ | 6 | 0 | 31.57 | |||||
| ampC | 1 | 0 | 5.26 | |||||
| Cefuroxime | Total | 2 | 16 | 1 | 0 | 10.52 | NT | NT |
| blaZ | 2 | 0 | 10.52 | |||||
| ampC | 0 | 0 | 0.00 | |||||
| Cephalothin | Total | 2 | 12 | 4 | 0 | 10.52 | NT | NT |
| blaZ | 1 | 0 | 5.26 | |||||
| ampC | 1 | 0 | 5.26 | |||||
| Ceftiofur | Total | 0 | 19 | 0 | 0 | 0.00 | NT | NT |
| blaZ | 0 | 0 | 0.00 | |||||
| ampC | 0 | 0 | 0.00 | |||||
| Bacteria Species | r 1 | p 2 |
|---|---|---|
| Streptococcus (n = 73) | −0.04193 | 0.7247 |
| Staphylococcus (n = 32) | −0.22953 | 0.2063 |
| Coliforms (n = 19) | −0.01996 | 0.9354 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhital, B.; Chuang, S.-T.; Hsieh, J.-C.; Hsieh, M.-H.; Chiang, H.-I. Prevalence, Virulence, and Antimicrobial Resistance of Major Mastitis Pathogens Isolated from Taiwanese Dairy Farms. Antibiotics 2024, 13, 36. https://doi.org/10.3390/antibiotics13010036
Dhital B, Chuang S-T, Hsieh J-C, Hsieh M-H, Chiang H-I. Prevalence, Virulence, and Antimicrobial Resistance of Major Mastitis Pathogens Isolated from Taiwanese Dairy Farms. Antibiotics. 2024; 13(1):36. https://doi.org/10.3390/antibiotics13010036
Chicago/Turabian StyleDhital, Bigya, Shih-Te Chuang, Jui-Chun Hsieh, Ming-Hsiu Hsieh, and Hsin-I Chiang. 2024. "Prevalence, Virulence, and Antimicrobial Resistance of Major Mastitis Pathogens Isolated from Taiwanese Dairy Farms" Antibiotics 13, no. 1: 36. https://doi.org/10.3390/antibiotics13010036
APA StyleDhital, B., Chuang, S.-T., Hsieh, J.-C., Hsieh, M.-H., & Chiang, H.-I. (2024). Prevalence, Virulence, and Antimicrobial Resistance of Major Mastitis Pathogens Isolated from Taiwanese Dairy Farms. Antibiotics, 13(1), 36. https://doi.org/10.3390/antibiotics13010036