


Prevalence, Genetic Homogeneity, and Antibiotic Resistance of Pathogenic Yersinia enterocolitica Strains Isolated from Slaughtered Pigs in Bulgaria

 ,
,  ,
,  , , ,
, , ,  , ,
, , 
Abstract
1. Introduction
2. Results
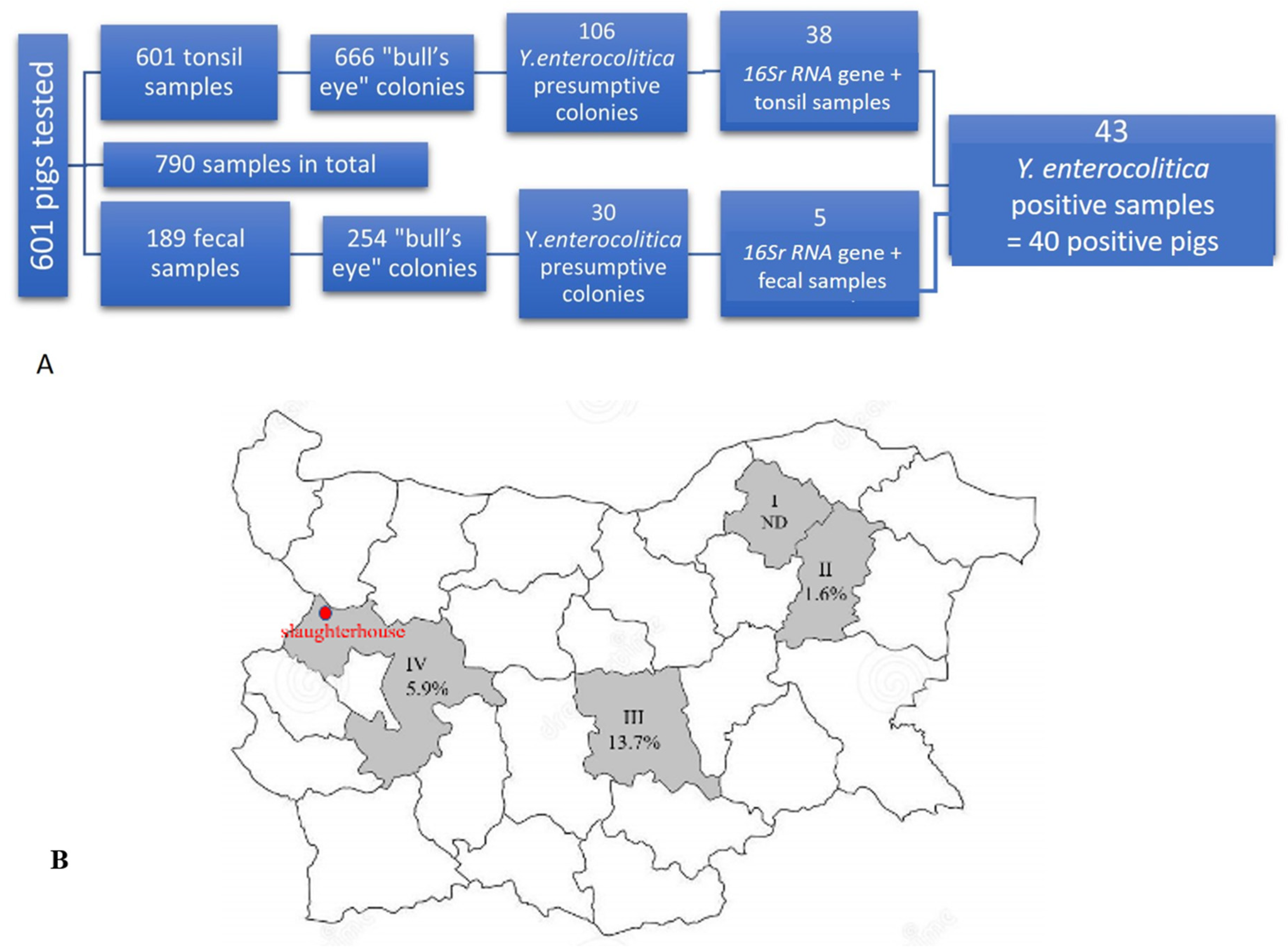
2.1. Detection of Yersinia enterocolitica
2.2. Biotyping and Serotyping
2.3. Detection of Virulence Genes
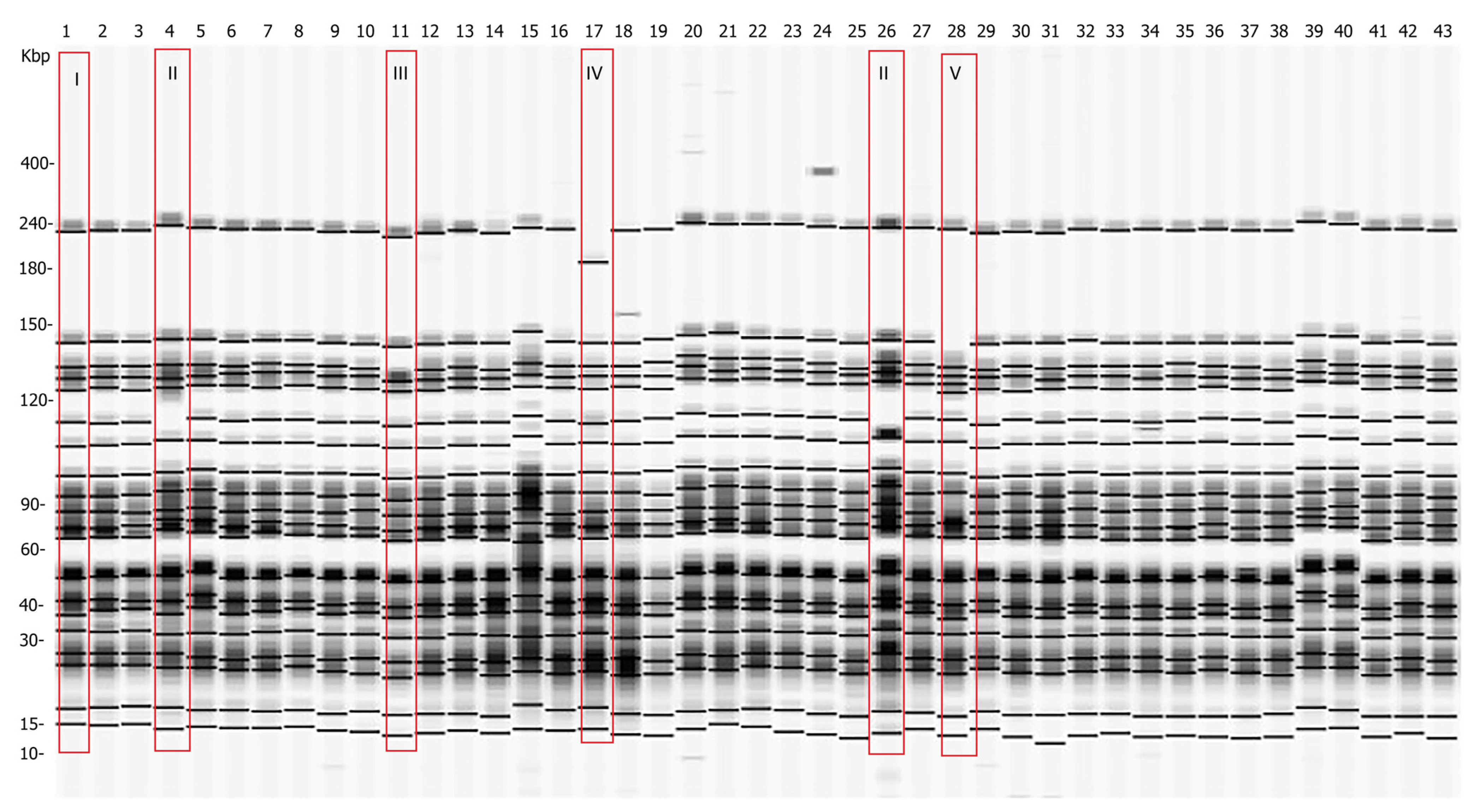
2.4. Genetic Diversity of Y. enterocolitica Strains Determined by PFGE
2.5. Antimicrobial Susceptibility of Y. enterocolitica Strains
3. Discussion
4. Materials and Methods
4.1. Animals and Samples
4.2. Microbiological and Biochemical Tests for Detection of Yersinia enterocolitica
4.3. DNA Isolation
4.4. 16S rRNA Gene Identification
4.5. Biotyping and Serotyping Methods
4.6. Phenotypic Test for Detection of Virulent Plasmid
4.7. PCR for Detection of Y. enterocolitica Virulence Genes
4.8. Pulsed-Field Gel Electrophoresis
4.9. Determination of the Antimicrobial Susceptibility of Y. enterocolitica Strains

{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primers | Tm °C | Sequence (5′ to 3′) | Amplicon Length (bp) | Reference | PCR Conditions |
|---|---|---|---|---|---|---|
| 16S rRNA Yersinia enterocolitica | YeI-6SrRNA YeII-6SrRNA | 48 °C 45 °C | ATACCGCATAACGTCTTCG TTCTTCTGCGAGTAACGTC | 328 | [44] | 95 °C for 5 min 35 cycles 94 °C for 30 s 47 °C for 30 s 72 °C for 1 min 72 °C for 10 min |
| ail | F-real 10A R-real 9A | 55 °C 55 °C | ATGATAACTGGGGAGTAATAGGTTCG CCCAGTAATCCATAAAGGCTAACATAT | 163 | [45] | 95 °C for 5 min 35 cycles 94 °C for 30 s 55 °C for 45 s 72 °C for 1 min 72 °C for 10 min |
| ystA | ystA-F ystA-R | 63 °C 57 °C | AATGCTGTCTTCATTTGGAGC ATCCCAATCACTACTGACTTC | 145 | [29] | 95 °C for 5 min 35 cycles 94 °C for 30 s 60 °C for 45 s 72 °C for 1 min 72 °C for 10 min |
| yadA | yadA-F yadA-R | 63 °C 63 °C | CTTCAGATACTGGTGTCGCTGT ATGCCTGACTAGAGCGATATCC | 849 | [29] | 95 °C for 5 min 35 cycles 94 °C for 30 s 63 °C for 45 s 72 °C for 1 min 72 °C for 10 min |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le Guern, A.-S.; Savin, C.; Angermeier, H.; Brémont, S.; Clermont, D.; Mühle, E.; Orozova, P.; Najdenski, H.; Pizarro-Cerdá, J. Yersinia artesiana sp. nov., Yersinia proxima sp. nov., Yersinia alsatica sp. nov., Yersina vastinensis sp. nov., Yersinia thracica sp. nov. and Yersinia occitanica sp. nov., isolated from humans and animals. Int. J. Syst. Evol. Microbiol. 2020, 70, 5363. [Google Scholar] [CrossRef] [PubMed]
- Bottone, E.J. Yersinia enterocolitica: Revisitation of an enduring human pathogen. Clin. Microbiol. Newsl. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, e07666. [Google Scholar] [CrossRef]
- Virdi, J.S.; Sachdeva, P. Molecular heterogeneity in Yersinia enterocolitica and ‘Y. enterocolitica-like’species—Implications for epidemiology, typing and taxonomy. FEMS Immunol. Med. Microbiol. 2005, 45, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Marimon, J.; Figueroa, R.; Idigoras, P.; Gomariz, M.; Alkorta, M.; Cilla, G.; Pérez-Trallero, E. Thirty years of human infections caused by Yersinia enterocolitica in northern Spain: 1985–2014. Epidemiol. Infect. 2017, 145, 2197–2203. [Google Scholar] [CrossRef] [PubMed]
- Morka, K.; Wałecka-Zacharska, E.; Schubert, J.; Dudek, B.; Woźniak-Biel, A.; Kuczkowski, M.; Wieliczko, A.; Bystroń, J.; Bania, J.; Bugla-Płoskońska, G. Genetic diversity and distribution of virulence-associated genes in Y. enterocolitica and Y. enterocolitica-like isolates from humans and animals in Poland. Pathogens 2021, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Terentjeva, M.; Ķibilds, J.; Gradovska, S.; Alksne, L.; Streikiša, M.; Meistere, I.; Valciņa, O. Prevalence, virulence determinants, and genetic diversity in Yersinia enterocolitica isolated from slaughtered pigs and pig carcasses. Int. J. Food Microbiol. 2022, 376, 109756. [Google Scholar] [CrossRef]
- Arsić, M.; Vićić, I.; Galić, N.; Dmitrić, M.; Kureljušić, J.; Dimitrijević, M.; Petrović, M.; Šarić, L.; Karabasil, N. Risk factors and the overall characterization of Yersinia enterocolitica as an initial model of pathogen surveillance in the pig production system in Serbia. Res. Vet. Sci. 2022, 152, 167–174. [Google Scholar] [CrossRef]
- Van Damme, I.; Berkvens, D.; Vanantwerpen, G.; Baré, J.; Houf, K.; Wauters, G.; De Zutter, L. Contamination of freshly slaughtered pig carcasses with enteropathogenic Yersinia spp.: Distribution, quantification and identification of risk factors. Int. J. Food Microbiol. 2015, 204, 33–40. [Google Scholar] [CrossRef]
- Martins, B.T.F.; Botelho, C.V.; Silva, D.A.L.; Lanna, F.G.P.A.; Grossi, J.L.; Campos-Galvão, M.E.M.; Yamatogi, R.S.; Falcão, J.P.; dos Santos Bersot, L.; Nero, L.A. Yersinia enterocolitica in a Brazilian pork production chain: Tracking of contamination routes, virulence and antimicrobial resistance. Int. J. Food Microbiol. 2018, 276, 5–9. [Google Scholar] [CrossRef]
- Vanantwerpen, G.; Van Damme, I.; De Zutter, L.; Houf, K. Within-batch prevalence and quantification of human pathogenic Yersinia enterocolitica and Y. pseudotuberculosis in tonsils of pigs at slaughter. Vet. Microbiol. 2014, 169, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson-Ahomaa, M.; Stolle, A.; Stephan, R. Prevalence of pathogenic Yersinia enterocolitica in pigs slaughtered at a Swiss abattoir. Int. J. Food Microbiol. 2007, 119, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Bancerz-Kisiel, A.; Pieczywek, M.; Łada, P.; Szweda, W. The most important virulence markers of Yersinia enterocolitica and their role during infection. Genes 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Bhaduri, S.; Smith, J.L. Virulence plasmid (pYV)-associated expression of phenotypic virulent determinants in pathogenic Yersinia species: A convenient method for monitoring the presence of pYV under culture conditions and its application for isolation/detection of Yersinia pestis in food. J. Pathog. 2011, 2011, 727313. [Google Scholar] [CrossRef] [PubMed]
- Fois, F.; Piras, F.; Torpdahl, M.; Mazza, R.; Ladu, D.; Consolati, S.G.; Spanu, C.; Scarano, C.; De Santis, E.P. Prevalence, bioserotyping and antibiotic resistance of pathogenic Yersinia enterocolitica detected in pigs at slaughter in Sardinia. Int. J. Food Microbiol. 2018, 283, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Fàbrega, A.; Ballesté-Delpierre, C.; Vila, J. Antimicrobial resistance in Yersinia enterocolitica. In Antimicrobial resistance and food safety; Elsevier: Amsterdam, The Netherlands, 2015; pp. 77–104. [Google Scholar]
- Zdolec, N.; Kiš, M.; Jankuloski, D.; Blagoevska, K.; Kazazić, S.; Pavlak, M.; Blagojević, B.; Antić, D.; Fredriksson-Ahomaa, M.; Pažin, V. Prevalence and Persistence of Multidrug-Resistant Yersinia enterocolitica 4/O: 3 in Tonsils of Slaughter Pigs from Different Housing Systems in Croatia. Foods 2022, 11, 1459. [Google Scholar] [CrossRef]
- Clinical and Labortory Standards Institute (CLSI). CLSI Supplement M100. In Performance Standard for Antimicrobial Susceptibility Testing, 31st ed.; Clinical and Labortory Standards Institute (CLSI): Wayne, PA, USA, 2021. [Google Scholar]
- Sacchini, L.; Garofolo, G.; Di Serafino, G.; Marotta, F.; Ricci, L.; Di Donato, G.; Miracco, M.G.; Perletta, F.; Di Giannatale, E. The prevalence, characterisation, and antimicrobial resistance of Yersinia enterocolitica in pigs from Central Italy. Vet. Ital. 2018, 54, 115–123. [Google Scholar] [CrossRef]
- Bonardi, S.; Bruini, I.; D’incau, M.; Van Damme, I.; Carniel, E.; Brémont, S.; Cavallini, P.; Tagliabue, S.; Brindani, F. Detection, seroprevalence and antimicrobial resistance of Yersinia enterocolitica and Yersinia pseudotuberculosis in pig tonsils in Northern Italy. Int. J. Food Microbiol. 2016, 235, 125–132. [Google Scholar] [CrossRef]
- Ibañez, T.R.; Laukkanen-Ninios, R.; Hakkinen, M.; Johansson, T.; Vilar, M.; Korkeala, H. Prevalence of pathogenic Yersinia enterocolitica in finnish slaughter pigs. J. Food Prot. 2016, 79, 677–681. [Google Scholar] [CrossRef]
- Martinez, P.O.; Fredriksson-Ahomaa, M.; Pallotti, A.; Rosmini, R.; Houf, K.; Korkeala, H. Variation in the prevalence of enteropathogenic Yersinia in slaughter pigs from Belgium, Italy, and Spain. Foodborne Pathog. Dis. 2011, 8, 445–450. [Google Scholar] [CrossRef]
- Mazzette, R.; Fois, F.; Consolati, S.G.; Salza, S.; Tedde, T.; Soro, P.; Collu, C.; Ladu, D.; Virgilio, S.; Piras, F. Detection of pathogenic Yersinia enterocolitica in slaughtered pigs by cultural methods and real-time polymerase chain reaction. Ital. J. Food Saf. 2015, 4, 4579. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson-Ahomaa, M.; Korkeala, H. Low occurrence of pathogenic Yersinia enterocolitica in clinical, food, and environmental samples: A methodological problem. Clin. Microbiol. Rev. 2003, 16, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, I.; Habib, I.; De Zutter, L. Yersinia enterocolitica in slaughter pig tonsils: Enumeration and detection by enrichment versus direct plating culture. Food Microbiol. 2010, 27, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Nesbakken, T.; Iversen, T.; Eckner, K.; Lium, B. Testing of pathogenic Yersinia enterocolitica in pig herds based on the natural dynamic of infection. Int. J. Food Microbiol. 2006, 111, 99–104. [Google Scholar] [CrossRef]
- Martínez, P.O.; Mylona, S.; Drake, I.; Fredriksson-Ahomaa, M.; Korkeala, H.; Corry, J.E. Wide variety of bioserotypes of enteropathogenic Yersinia in tonsils of English pigs at slaughter. Int. J. Food Microbiol. 2010, 139, 64–69. [Google Scholar] [CrossRef]
- Bonardi, S.; Bassi, L.; Brindani, F.; D’Incau, M.; Barco, L.; Carra, E.; Pongolini, S. Prevalence, characterization and antimicrobial susceptibility of Salmonella enterica and Yersinia enterocolitica in pigs at slaughter in Italy. Int. J. Food Microbiol. 2013, 163, 248–257. [Google Scholar] [CrossRef]
- Saraka, D.; Savin, C.; Kouassi, S.; Cissé, B.; Koffi, E.; Cabanel, N.; Brémont, S.; Faye-Kette, H.; Dosso, M.; Carniel, E. Yersinia enterocolitica, a neglected cause of human enteric infections in Cote d’Ivoire. PLoS Negl. Trop. Dis. 2017, 11, e0005216. [Google Scholar] [CrossRef]
- Martins, B.T.F.; de Azevedo, E.C.; Yamatogi, R.S.; Call, D.R.; Nero, L.A. Persistence of Yersinia enterocolitica bio-serotype 4/O: 3 in a pork production chain in Minas Gerais, Brazil. Food Microbiol. 2021, 94, 103660. [Google Scholar] [CrossRef]
- Najdenski, H.; Iteman, I.; Carniel, E. The genome of Yersinia enterocolitica is the most stable of the three pathogenic species. Contrib. Microbiol. Immunol. 1995, 13, 281–284. [Google Scholar]
- Najdenski, H.; Iteman, I.; Carniel, E. Efficient subtyping of pathogenic Yersinia enterocolitica strains by pulsed-field gel electrophoresis. J. Clin. Microbiol. 1994, 32, 2913–2920. [Google Scholar] [CrossRef]
- Raymond, P.; Houard, E.; Denis, M.; Esnault, E. Diversity of Yersinia enterocolitica isolated from pigs in a French slaughterhouse over 2 years. Microbiologyopen 2019, 8, e00751. [Google Scholar] [CrossRef] [PubMed]
- Bent, Z.W.; Young, G.M. Contribution of BlaA and BlaB β-lactamases to antibiotic susceptibility of Yersinia enterocolitica biovar 1B. Antimicrob. Agents Chemother. 2010, 54, 4000–4002. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, M.; Brodard, I.; Overesch, G. Virulence-associated gene pattern of porcine and human Yersinia enterocolitica biotype 4 isolates. Int. J. Food Microbiol. 2015, 198, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Bonardi, S.; Alpigiani, I.; Pongolini, S.; Morganti, M.; Tagliabue, S.; Bacci, C.; Brindani, F. Detection, enumeration and characterization of Yersinia enterocolitica 4/O: 3 in pig tonsils at slaughter in Northern Italy. Int. J. Food Microbiol. 2014, 177, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, P.A.; Tano, E.; Jernberg, C.; Hickman, R.A.; Guy, L.; Järhult, J.D.; Wang, H. Molecular characterization of multidrug-resistant Yersinia enterocolitica from foodborne outbreaks in Sweden. Front. Microbiol. 2021, 12, 664665. [Google Scholar] [CrossRef] [PubMed]
- Gkouletsos, T.; Patas, K.; Lambrinidis, G.; Neubauer, H.; Sprague, L.; Ioannidis, A.; Chatzipanagiotou, S. Antimicrobial resistance of Yersinia enterocolitica and presence of plasmid pYV virulence genes in human and animal isolates. New Microbes New Infect. 2019, 32, 100604. [Google Scholar] [CrossRef] [PubMed]
- Thong, K.L.; Tan, L.K.; Ooi, P.T. Genetic diversity, virulotyping and antimicrobial resistance susceptibility of Yersinia enterocolitica isolated from pigs and porcine products in Malaysia. J. Sci. Food Agric. 2018, 98, 87–95. [Google Scholar] [CrossRef]
- Hooper, D.C. Mechanisms of action of antimicrobials: Focus on fluoroquinolones. Clin. Infect. Dis. 2001, 32, S9–S15. [Google Scholar] [CrossRef]
- ISO. Microbiology of Food and Animal Feeding Stuffs-Horizontal Method for the Detection of Presumptive Pathogenic Yersinia enterocolitica; ISO (International Organization for Standardization): London, UK, 2003. [Google Scholar]
- Centers for Disease Control and Prevention. One-Day (24–28 h) Standardized Laboratory. Protocol for Molecular Subtyping of Yersinia pestis by Pulsed Field Gel Electrophoresis (PFGE); Pulse Net: Atlanta, GA, USA, 2006. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST) Breakpoint Tables for Interpretation of MICs and Zone Diameters. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_12.0_Breakpoint_Tables.pdf (accessed on 17 February 2023).
- Arnold, T.; Neubauer, H.; Nikolaou, K.; Roesler, U.; Hensel, A. Identification of Yersinia enterocolitica in minced meat: A comparative analysis of API 20E, Yersinia identification kit and a 16S rRNA-based PCR method. J. Vet. Med. Ser. B 2004, 51, 23–27. [Google Scholar] [CrossRef]
- Lambertz, S.T.; Nilsson, C.; Hallanvuo, S.; Lindblad, M. Real-time PCR method for detection of pathogenic Yersinia enterocolitica in food. Appl. Environ. Microbiol. 2008, 74, 6060–6067. [Google Scholar] [CrossRef]



| Antibiotics | Resistance Profile | |||
|---|---|---|---|---|
| Class * | Generic name | 41/43 | 1/43 | 1/43 |
| Penicillins | ampicillin | − | − | − |
| Cephems | cefamandole | − | − | − |
| ceftriaxone | + | + | + | |
| Aminoglycosides | amikacin | + | + | + |
| gentamicin | + | + | + | |
| streptomycin | + | − | + | |
| Phenicols | chloramphenicol | + | − | − |
| Tetracyclines | tetracycline | + | − | − |
| doxycycline | + | + | − | |
| Quinolones | nalidixic acid | + | − | − |
| ciprofloxacin | + | + | + | |
| levofloxacin | + | − | − | |
| Folate pathway antagonist | trimethoprim/ sulfamethoxazole | + | + | − |
| Aminocoumarins | novobiocin | − | − | − |
| Other | bacitracin | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelovska, M.; Zaharieva, M.M.; Dimitrova, L.L.; Dimova, T.; Gotova, I.; Urshev, Z.; Ilieva, Y.; Kaleva, M.D.; Kim, T.C.; Naydenska, S.; et al. Prevalence, Genetic Homogeneity, and Antibiotic Resistance of Pathogenic Yersinia enterocolitica Strains Isolated from Slaughtered Pigs in Bulgaria. Antibiotics 2023, 12, 716. https://doi.org/10.3390/antibiotics12040716
Angelovska M, Zaharieva MM, Dimitrova LL, Dimova T, Gotova I, Urshev Z, Ilieva Y, Kaleva MD, Kim TC, Naydenska S, et al. Prevalence, Genetic Homogeneity, and Antibiotic Resistance of Pathogenic Yersinia enterocolitica Strains Isolated from Slaughtered Pigs in Bulgaria. Antibiotics. 2023; 12(4):716. https://doi.org/10.3390/antibiotics12040716
Chicago/Turabian StyleAngelovska, Maya, Maya Margaritova Zaharieva, Lyudmila L. Dimitrova, Tanya Dimova, Irina Gotova, Zoltan Urshev, Yana Ilieva, Mila Dobromirova Kaleva, Tanya Chan Kim, Sevda Naydenska, and et al. 2023. "Prevalence, Genetic Homogeneity, and Antibiotic Resistance of Pathogenic Yersinia enterocolitica Strains Isolated from Slaughtered Pigs in Bulgaria" Antibiotics 12, no. 4: 716. https://doi.org/10.3390/antibiotics12040716
APA StyleAngelovska, M., Zaharieva, M. M., Dimitrova, L. L., Dimova, T., Gotova, I., Urshev, Z., Ilieva, Y., Kaleva, M. D., Kim, T. C., Naydenska, S., Dimitrov, Z., & Najdenski, H. (2023). Prevalence, Genetic Homogeneity, and Antibiotic Resistance of Pathogenic Yersinia enterocolitica Strains Isolated from Slaughtered Pigs in Bulgaria. Antibiotics, 12(4), 716. https://doi.org/10.3390/antibiotics12040716