
Cytotoxicity and Antimicrobial Resistance of Aeromonas Strains Isolated from Fresh Produce and Irrigation Water

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
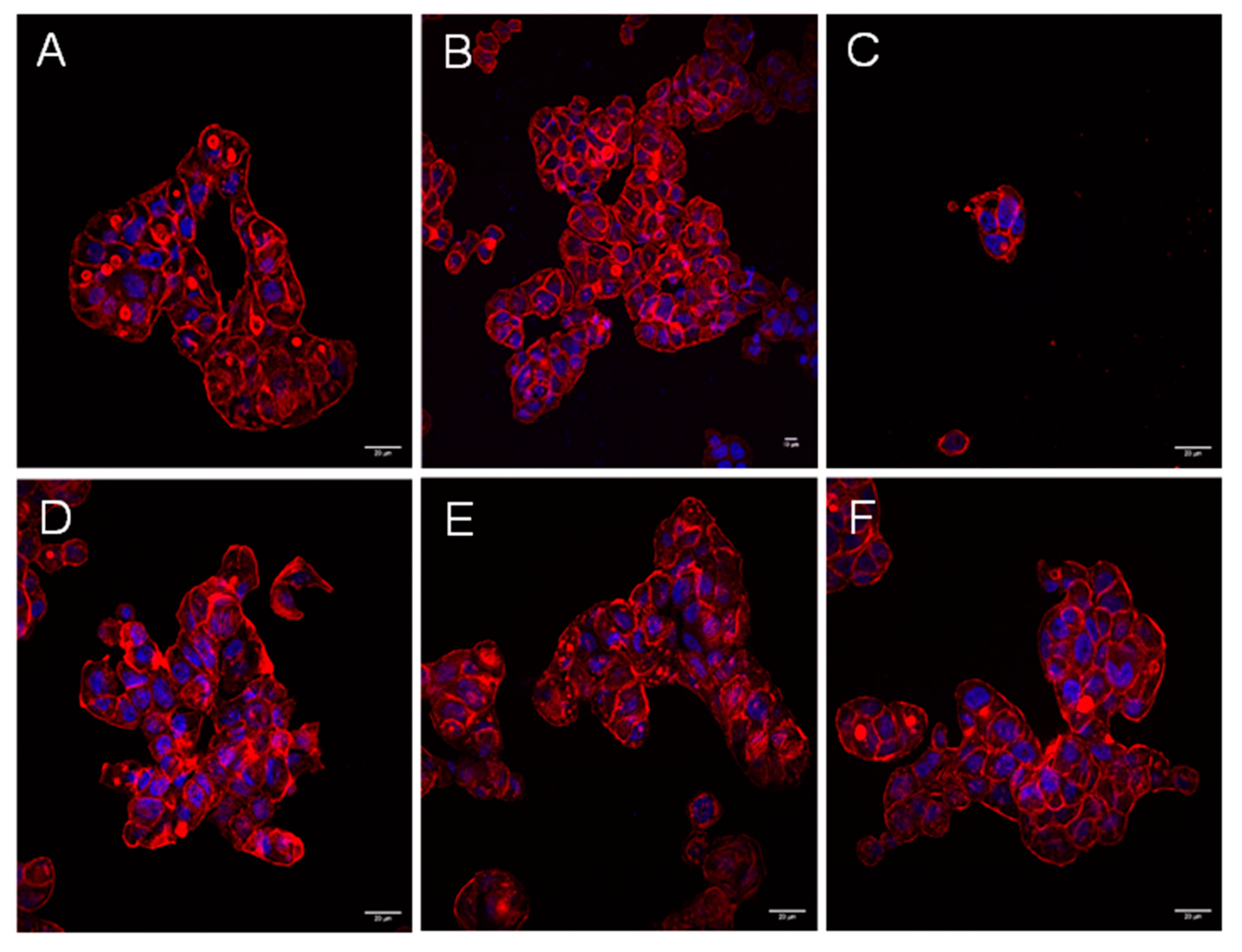
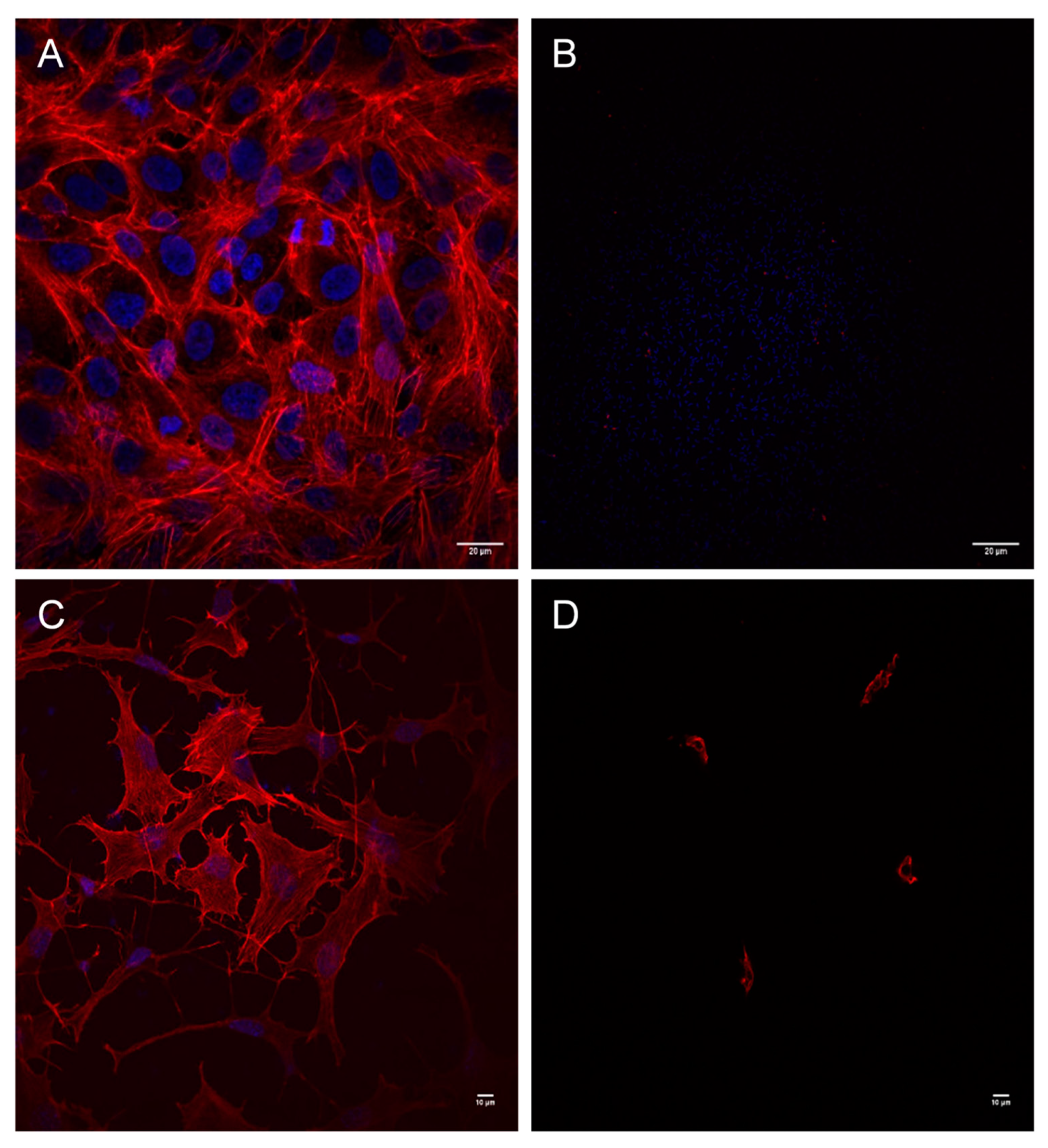
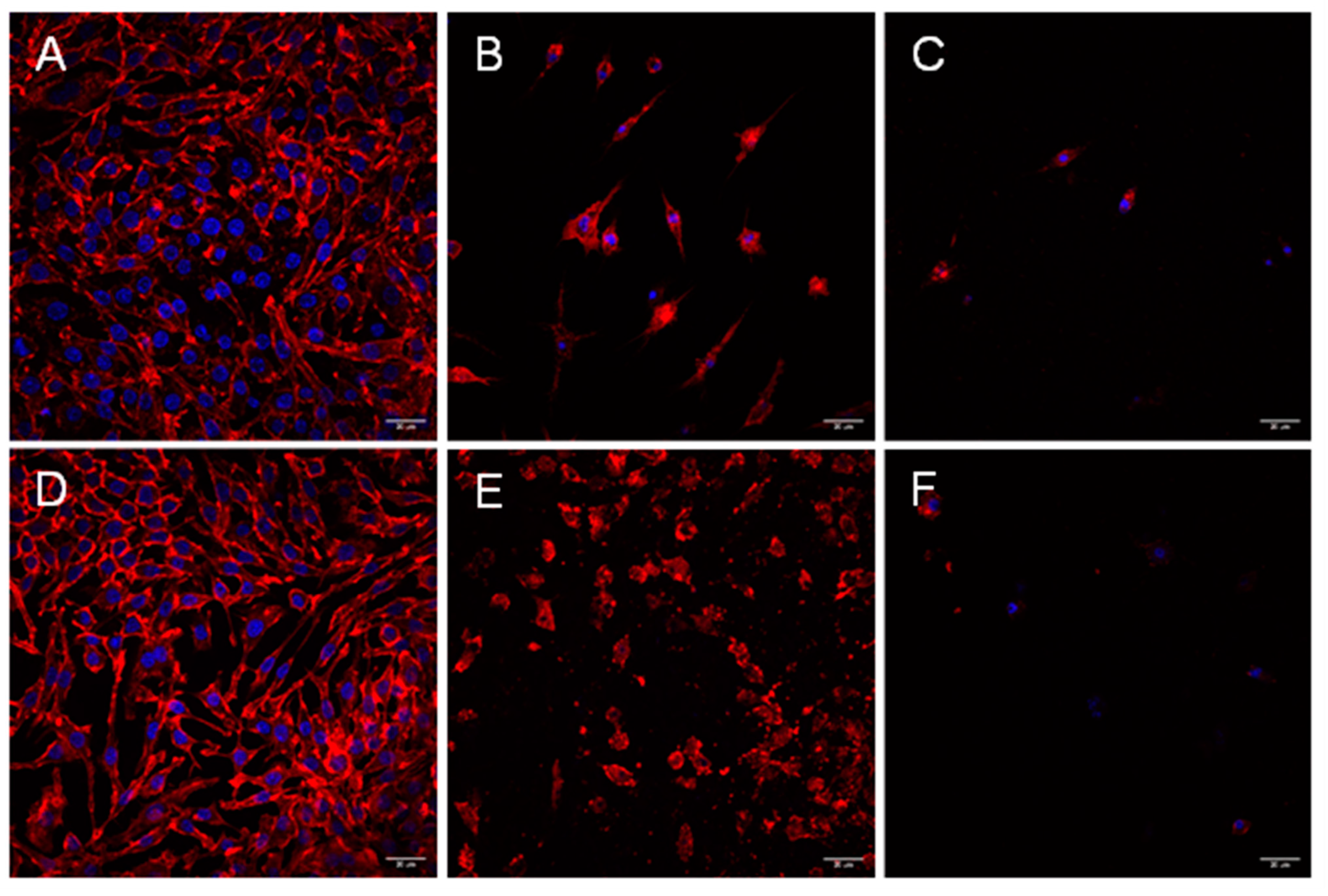
2.1. Cytotoxic Effects of Aeromonas Strains on Several Cell Lines
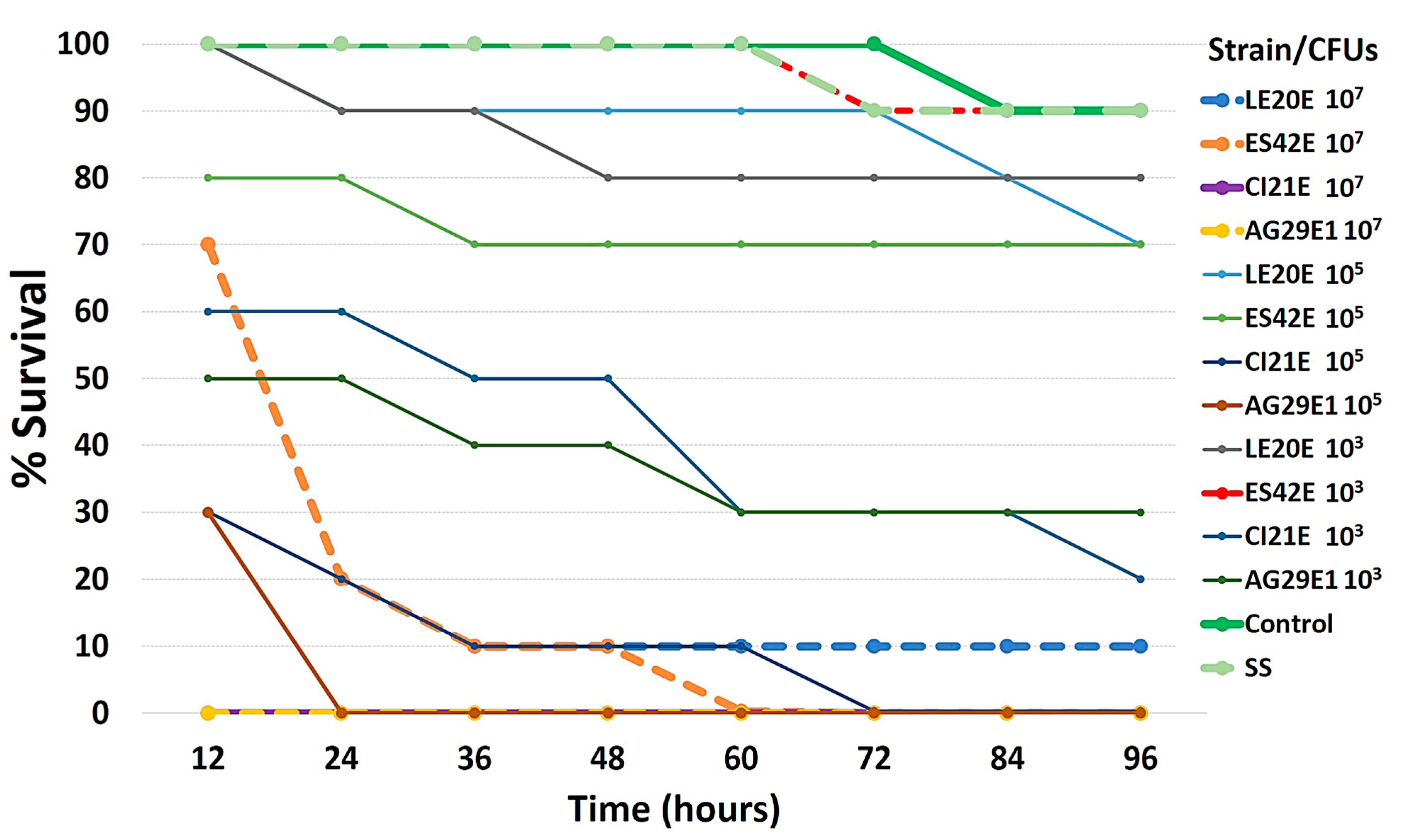
2.2. Virulence in Galleria mellonella
2.3. Antimicrobial Susceptibility Testing
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Cell Lines
4.3. Fluorescence Assays
4.4. Cytotoxicity of Bacterial Extracellular Products
4.5. Bacterial Cell-Contact Cytotoxicity
4.6. Galleria mellonella Killing Assays
4.7. Antimicrobial Susceptibility Testing
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azzam-Sayuti, M.; Ina-Salwany, M.Y.; Zamri-Saad, M.; Annas, S.; Yusof, M.T.; Monir, M.S.; Mohamad, A.; Muhamad-Sofie, M.H.N.; Lee, J.Y.; Chin, Y.K.; et al. Comparative Pathogenicity of Aeromonas Spp. in Cultured Red Hybrid Tilapia (Oreochromis niloticus × O. Mossambicus). Biology 2021, 10, 1192. [Google Scholar] [CrossRef] [PubMed]
- Grilo, M.L.; Amaro, G.; Chambel, L.; Marques, C.S.; Marques, T.A.; Gil, F.; Sousa-Santos, C.; Robalo, J.I.; Oliveira, M. Aeromonas Spp. Prevalence, Virulence, and Antimicrobial Resistance in an Ex Situ Program for Threatened Freshwater Fish-A Pilot Study with Protective Measures. Animals 2022, 12, 436. [Google Scholar] [CrossRef]
- Dhanapala, P.M.; Kalupahana, R.S.; Kalupahana, A.W.; Wijesekera, D.P.H.; Kottawatta, S.A.; Jayasekera, N.K.; Silva-Fletcher, A.; Jagoda, S.S.S.d.S. Characterization and Antimicrobial Resistance of Environmental and Clinical Aeromonas Species Isolated from Fresh Water Ornamental Fish and Associated Farming Environment in Sri Lanka. Microorganisms 2021, 9, 2106. [Google Scholar] [CrossRef] [PubMed]
- Hoel, S.; Vadstein, O.; Jakobsen, A.N. The Significance of Mesophilic Aeromonas spp. in Minimally Processed Ready-to-Eat Seafood. Microorganisms 2019, 7, E91. [Google Scholar] [CrossRef] [PubMed]
- Hammad, A.M.; Moustafa, A.-E.H.; Mansour, M.M.; Fahmy, B.M.; Hamada, M.G.; Shimamoto, T.; Shimamoto, T. Molecular and Phenotypic Analysis of Hemolytic Aeromonas Strains Isolated from Food in Egypt Revealed Clinically Important Multidrug Resistance and Virulence Profiles. J. Food Prot. 2018, 81, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Schuetz, A.N. Emerging Agents of Gastroenteritis: Aeromonas, Plesiomonas, and the Diarrheagenic Pathotypes of Escherichia Coli. Semin. Diagn. Pathol. 2019, 36, 187–192. [Google Scholar] [CrossRef]
- Chopra, A.K.; Houston, C.W. Enterotoxins in Aeromonas-Associated Gastroenteritis. Microbes Infect. 1999, 1, 1129–1137. [Google Scholar] [CrossRef]
- Saleh, A.; Elkenany, R.; Younis, G. Virulent and Multiple Antimicrobial Resistance Aeromonas Hydrophila Isolated from Diseased Nile Tilapia Fish (Oreochromis Niloticus) in Egypt with Sequencing of Some Virulence-Associated Genes. Biocontrol. Sci. 2021, 26, 167–176. [Google Scholar] [CrossRef]
- Gonzalez-Avila, L.U.; Loyola-Cruz, M.A.; Hernández-Cortez, C.; Bello-López, J.M.; Castro-Escarpulli, G. Colistin Resistance in Aeromonas spp. Int. J. Mol. Sci. 2021, 22, 5974. [Google Scholar] [CrossRef]
- Lamy, B.; Baron, S.; Barraud, O. Aeromonas: The Multifaceted Middleman in the One Health World. Curr. Opin. Microbiol. 2022, 65, 24–32. [Google Scholar] [CrossRef]
- Vivas, J.; Sáa, A.I.; Tinajas, A.; Barbeyto, L.; Rodríguez, L.A. Identification of Motile Aeromonas Strains with the MicroScan WalkAway System in Conjunction with the Combo Negative Type 1S Panels. Appl. Environ. Microbiol. 2000, 66, 1764–1766. [Google Scholar] [CrossRef] [PubMed]
- Park, S.M.; Kim, H.W.; Choi, C.; Rhee, M.S. Pathogenicity and Seasonal Variation of Aeromonas Hydrophila Isolated from Seafood and Ready-to-Eat Sushi in South Korea. Food Res. Int. 2021, 147, 110484. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Hoel, S.; Lunestad, B.-T.; Lerfall, J.; Jakobsen, A.N. Aeromonas Spp. Isolated from Ready-to-Eat Seafood on the Norwegian Market: Prevalence, Putative Virulence Factors and Antimicrobial Resistance. J. Appl. Microbiol. 2021, 130, 1380–1393. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.; Rasheduzzaman, M.; Darling, A.; Krometis, L.-A.; Edwards, M.; Brown, T.; Ahmed, T.; Wettstone, E.; Pholwat, S.; Taniuchi, M.; et al. Bottled and Well Water Quality in a Small Central Appalachian Community: Household-Level Analysis of Enteric Pathogens, Inorganic Chemicals, and Health Outcomes in Rural Southwest Virginia. Int. J. Environ. Res. Public Health 2022, 19, 8610. [Google Scholar] [CrossRef]
- Yu, Q.; Feng, T.; Yang, J.; Su, W.; Zhou, R.; Wang, Y.; Zhang, H.; Li, H. Seasonal Distribution of Antibiotic Resistance Genes in the Yellow River Water and Tap Water, and Their Potential Transmission from Water to Human. Environ. Pollut. 2022, 292, 118304. [Google Scholar] [CrossRef]
- Miyagi, K.; Sano, K.; Hirai, I. Sanitary Evaluation of Domestic Water Supply Facilities with Storage Tanks and Detection of Aeromonas, Enteric and Related Bacteria in Domestic Water Facilities in Okinawa Prefecture of Japan. Water Res. 2017, 119, 171–177. [Google Scholar] [CrossRef]
- Didugu, H.; Thirtham, M.; Nelapati, K.; Reddy, K.K.; Kumbhar, B.S.; Poluru, A.; Pothanaboyina, G. A Study on the Prevalence of Aeromonas Spp. and Its Enterotoxin Genes in Samples of Well Water, Tap Water, and Bottled Water. Vet. World 2015, 8, 1237–1242. [Google Scholar] [CrossRef]
- Miyagi, K.; Shimoji, N.; Shimoji, S.; Tahara, R.; Uechi, A.; Tamaki, I.; Oshiro, H.; Komiyama, A.; Tedokon, M.; Hirai, I. Comparison of Species, Virulence Genes and Clones of Aeromonas Isolates from Clinical Specimens and Well Water in Okinawa Prefecture, Japan. J. Appl. Microbiol. 2021, 131, 1515–1530. [Google Scholar] [CrossRef]
- Vivas, J.; Razquin, B.E.; López-Fierro, P.; Naharro, G.; Villena, A. Correlation between Production of Acyl Homoserine Lactones and Proteases in an Aeromonas Hydrophila AroA Live Vaccine. Vet. Microbiol. 2004, 101, 167–176. [Google Scholar] [CrossRef]
- Romero, A.; Saraceni, P.R.; Merino, S.; Figueras, A.; Tomás, J.M.; Novoa, B. The Animal Model Determines the Results of Aeromonas Virulence Factors. Front. Microbiol. 2016, 7, 1574. [Google Scholar] [CrossRef]
- Kingombe, C.I.; Huys, G.; Tonolla, M.; Albert, M.J.; Swings, J.; Peduzzi, R.; Jemmi, T. PCR Detection, Characterization, and Distribution of Virulence Genes in Aeromonas spp. Appl. Environ. Microbiol. 1999, 65, 5293–5302. [Google Scholar] [CrossRef] [PubMed]
- Körkoca, H.; Alan, Y.; Bozari, S.; Berktas, M.; Goz, Y. Detection of Putative Virulence Genes in Aeromonas Isolates from Humans and Animals. J. Infect. Dev. Ctries 2014, 8, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Dahanayake, P.S.; Hossain, S.; Wickramanayake, M.V.K.S.; Heo, G.-J. Prevalence of Virulence and Antimicrobial Resistance Genes in Aeromonas Species Isolated from Marketed Cockles (Tegillarca Granosa) in Korea. Lett. Appl. Microbiol. 2020, 71, 94–101. [Google Scholar] [CrossRef]
- Snowden, L.; Wernbacher, L.; Stenzel, D.; Tucker, J.; McKay, D.; O’Brien, M.; Katouli, M. Prevalence of Environmental Aeromonas in South East Queensland, Australia: A Study of Their Interactions with Human Monolayer Caco-2 Cells. J. Appl. Microbiol. 2006, 101, 964–975. [Google Scholar] [CrossRef]
- Thornley, J.P.; Shaw, J.G.; Gryllos, I.A.; Eley, A. Adherence of Aeromonas Caviae to Human Cell Lines Hep-2 and Caco-2. J. Med. Microbiol. 1996, 45, 445–451. [Google Scholar] [CrossRef]
- dos Santos, P.A.; Pereira, A.C.M.; Ferreira, A.F.; de Mattos Alves, M.A.; Rosa, A.C.P.; Freitas-Almeida, A.C. Adhesion, Invasion, Intracellular Survival and Cytotoxic Activity of Strains of Aeromonas Spp. in HEp-2, Caco-2 and T-84 Cell Lines. Antonie Van Leeuwenhoek 2015, 107, 1225–1236. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Yeh, W.-H.; Tang, H.-J.; Chen, J.-W.; Shu, H.-Y.; Su, Y.-C.; Wang, S.-T.; Kuo, C.-J.; Chuang, Y.-C.; Chen, C.-C.; et al. UvrY Is Required for the Full Virulence of Aeromonas Dhakensis. Virulence 2020, 11, 502–520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, Z.; Yang, H.; Wang, G.; Liu, G.; Wang, Y.; Bello, B.K.; Zhao, P.; Liang, W.; Dong, J. Aeromonas Sobria Induces Proinflammatory Cytokines Production in Mouse Macrophages via Activating NLRP3 Inflammasome Signaling Pathways. Front. Cell Infect. Microbiol. 2021, 11, 691445. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhao, X.; Jiang, X.; Zhang, J.; Li, L.; Pei, C.; Zhou, Y.; Zeng, L.; Kong, X. Contribution of Flagellar Cap Gene in Virulence and Pathogenicity of Aeromonas Veronii. J. Fish. Dis. 2023, 46, 247–259. [Google Scholar] [CrossRef]
- Fernández-Bravo, A.; Figueras, M.J. An Update on the Genus Aeromonas: Taxonomy, Epidemiology, and Pathogenicity. Microorganisms 2020, 8, 129. [Google Scholar] [CrossRef]
- Gonçalves Pessoa, R.B.; de Oliveira, W.F.; Marques, D.S.C.; Dos Santos Correia, M.T.; de Carvalho, E.V.M.M.; Coelho, L.C.B.B. The Genus Aeromonas: A General Approach. Microb. Pathog. 2019, 130, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Menanteau-Ledouble, S.; Kumar, G.; Saleh, M.; El-Matbouli, M. Aeromonas Salmonicida: Updates on an Old Acquaintance. Dis. Aquat. Organ. 2016, 120, 49–68. [Google Scholar] [CrossRef] [PubMed]
- Beaz-Hidalgo, R.; Figueras, M.J. Aeromonas Spp. Whole Genomes and Virulence Factors Implicated in Fish Disease. J. Fish. Dis. 2013, 36, 371–388. [Google Scholar] [CrossRef] [PubMed]
- Wiklund, T.; Dalsgaard, I. Occurrence and Significance of Atypical Aeromonas Salmonicida in Non-Salmonid and Salmonid Fish Species: A Review. Dis. Aquat. Organ. 1998, 32, 49–69. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, U.D.; Bhattacharjee, S. Bacteriological, Clinical and Virulence Aspects of Aeromonas-Associated Diseases in Humans. Pol. J. Microbiol. 2018, 67, 137–149. [Google Scholar] [CrossRef]
- Aeromonas Dhakensis, an Increasingly Recognized Human Pathogen—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/27303382/ (accessed on 12 November 2022).
- Liakopoulos, V.; Arampatzis, S.; Kourti, P.; Tsolkas, T.; Zarogiannis, S.; Eleftheriadis, T.; Giannopoulou, M.; Stefanidis, I. Aeromonas Hydrophila as a Causative Organism in Peritoneal Dialysis-Related Peritonitis: Case Report and Review of the Literature. Clin. Nephrol. 2011, 75 (Suppl. S1), 65–68. [Google Scholar]
- Janda, J.M.; Abbott, S.L. The Genus Aeromonas: Taxonomy, Pathogenicity, and Infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef]
- Kobayashi, H.; Seike, S.; Yamaguchi, M.; Ueda, M.; Takahashi, E.; Okamoto, K.; Yamanaka, H. Aeromonas sobria serine protease decreases epithelial barrier function in T84 cells and accelerates bacterial translocation across the T84 monolayer in vitro. PLoS ONE 2019, 14, e0221344. [Google Scholar] [CrossRef]
- Couto, C.R.A.; Oliveira, S.S.; Queiroz, M.L.P.; Freitas-Almeida, A.C. Interactions of Clinical and Environmental Aeromonas Isolates with Caco-2 and HT29 Intestinal Epithelial Cells. Lett. Appl. Microbiol. 2007, 45, 405–410. [Google Scholar] [CrossRef]
- Sierra, J.C.; Suarez, G.; Sha, J.; Foltz, S.M.; Popov, V.L.; Galindo, C.L.; Garner, H.R.; Chopra, A.K. Biological Characterization of a New Type III Secretion System Effector from a Clinical Isolate of Aeromonas Hydrophila-Part II. Microb. Pathog. 2007, 43, 147–160. [Google Scholar] [CrossRef]
- Sha, J.; Wang, S.F.; Suarez, G.; Sierra, J.C.; Fadl, A.A.; Erova, T.E.; Foltz, S.M.; Khajanchi, B.K.; Silver, A.; Graf, J.; et al. Further Characterization of a Type III Secretion System (T3SS) and of a New Effector Protein from a Clinical Isolate of Aeromonas Hydrophila--Part I. Microb. Pathog. 2007, 43, 127–146. [Google Scholar] [CrossRef] [PubMed]
- Galindo, C.L.; Sha, J.; Ribardo, D.A.; Fadl, A.A.; Pillai, L.; Chopra, A.K. Identification of Aeromonas Hydrophila Cytotoxic Enterotoxin-Induced Genes in Macrophages Using Microarrays. J. Biol. Chem. 2003, 278, 40198–40212. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, J.A.; Chopra, A.K. Modulation of Host Immune Defenses by Aeromonas and Yersinia Species: Convergence on Toxins Secreted by Various Secretion Systems. Front. Cell Infect. Microbiol. 2013, 3, 70. [Google Scholar] [CrossRef] [PubMed]
- Remuzgo-Martínez, S.; Aranzamendi-Zaldunbide, M.; Pilares-Ortega, L.; Icardo, J.M.; Acosta, F.; Martínez-Martínez, L.; Ramos-Vivas, J. Interaction of Macrophages with a Cytotoxic Serratia Liquefaciens Human Isolate. Microbes Infect. 2013, 15, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Hoel, S.; Vadstein, O.; Jakobsen, A.N. Species Distribution and Prevalence of Putative Virulence Factors in Mesophilic Aeromonas Spp. Isolated from Fresh Retail Sushi. Front. Microbiol. 2017, 8, 931. [Google Scholar] [CrossRef]
- Castilho, M.C.B.; Castro, T.L.A.; Araújo, V.S.; Trajano, R.S.; Santos, P.A.; Pimenta, P.M.C.; Lucheze, K.; Melo, J.T.B.; Gonçalves, A.M.; Nogueira, R.T.; et al. High Frequency of Hemolytic and Cytotoxic Activity in Aeromonas Spp. Isolated from Clinical, Food and Environmental in Rio de Janeiro, Brazil. Antonie Van Leeuwenhoek 2009, 96, 53–61. [Google Scholar] [CrossRef]
- Fernández-Bravo, A.; Figueras, M.J. Immune Response of the Monocytic Cell Line THP-1 Against Six Aeromonas spp. Front. Immunol. 2022, 13, 875689. [Google Scholar] [CrossRef]
- Silver, A.C.; Williams, D.; Faucher, J.; Horneman, A.J.; Gogarten, J.P.; Graf, J. Complex Evolutionary History of the Aeromonas Veronii Group Revealed by Host Interaction and DNA Sequence Data. PLoS ONE 2011, 6, e16751. [Google Scholar] [CrossRef]
- Sebastião, F.A.; Majolo, C.; Martins, V.F.S.; Boijink, C.L.; Brandão, F.R.; Pereira, S.L.A.; Fujimoto, R.Y.; Chagas, E.C. Antimicrobial Resistance Profile of Aeromonas Spp. Isolated from Asymptomatic Colossoma Macropomum Cultured in the Amazonas State, Brazil. Braz. J. Biol. 2023, 82, e260773. [Google Scholar] [CrossRef]
- Hossain, S.; Heo, G.-J. Detection of Antimicrobial and Heavy-Metal Resistance Genes in Aeromonas Spp. Isolated from Hard-Shelled Mussel (Mytilus Coruscus). Microb. Drug. Resist. 2022, 28, 127–135. [Google Scholar] [CrossRef]
- Dubey, S.; Ager-Wick, E.; Kumar, J.; Karunasagar, I.; Karunasagar, I.; Peng, B.; Evensen, Ø.; Sørum, H.; Munang’andu, H.M. Aeromonas Species Isolated from Aquatic Organisms, Insects, Chicken, and Humans in India Show Similar Antimicrobial Resistance Profiles. Front. Microbiol. 2022, 13, 1008870. [Google Scholar] [CrossRef] [PubMed]
- Dubey, S.; Ager-Wick, E.; Peng, B.; Evensen, Ø.; Sørum, H.; Munang’andu, H.M. Characterization of Virulence and Antimicrobial Resistance Genes of Aeromonas Media Strain SD/21-15 from Marine Sediments in Comparison with Other Aeromonas spp. Front. Microbiol. 2022, 13, 1022639. [Google Scholar] [CrossRef] [PubMed]
- De Silva, L.A.D.S.; Wickramanayake, M.V.K.S.; Heo, G.-J. Virulence and Antimicrobial Resistance Potential of Aeromonas Spp. Associated with Shellfish. Lett. Appl. Microbiol. 2021, 73, 176–186. [Google Scholar] [CrossRef]
- Ramos-Vivas, J.; Chapartegui-González, I.; Fernández-Martínez, M.; González-Rico, C.; Barrett, J.; Fortún, J.; Escudero, R.; Marco, F.; Linares, L.; Nieto, J.; et al. Adherence to Human Colon Cells by Multidrug Resistant Enterobacterales Strains Isolated From Solid Organ Transplant Recipients With a Focus on Citrobacter Freundii. Front. Cell Infect. Microbiol. 2020, 10, 447. [Google Scholar] [CrossRef] [PubMed]
- Chapartegui-González, I.; Lázaro-Díez, M.; Bravo, Z.; Navas, J.; Icardo, J.M.; Ramos-Vivas, J. Acinetobacter Baumannii Maintains Its Virulence after Long-Time Starvation. PLoS ONE 2018, 13, e0201961. [Google Scholar] [CrossRef] [PubMed]
- Pintor-Cora, A.; Álvaro-Llorente, L.; Otero, A.; Rodríguez-Calleja, J.M.; Santos, J.A. Extended-Spectrum Beta-Lactamase-Producing Enterobacteriaceae in Fresh Produce. Foods 2021, 10, 2609. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | PRL (30) * | CXM (30) | CTX (5) | CAZ (10) | FEP (30) | ATM (30) | IPM (10) | CN (10) | CIP (5) | SXT (23.75–1.25) | C (30) | AmpC/ESBL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LE20E | 24.2 | 26.6 | 27.1 | 30 | 38.3 | 39.4 | 33.9 | 26.6 | 35.5 | 19.2 | 32 | inducible |
| ES19E | 17.5 | 19.2 | 28.7 | 30 | 39 | 40.2 | 35.3 | 28.3 | 40.7 | 22.8 | 32.5 | inducible |
| ES20E | 12.3 | 22.4 | 26.3 | 30.4 | 43.4 | 39.8 | 35.2 | 20.5 | 41.5 | 25.5 | 31.2 | inducible |
| ES42E | 14.8 | 27.4 | 30.5 | 30.2 | 38.4 | 40.8 | 37.2 | 30.5 | 39.5 | 30.2 | 31.5 | inducible |
| AG26E | 17.8 | 22.8 | 30.6 | 31.4 | 39.3 | 39.2 | 32.5 | 26.1 | 35.8 | 24.5 | 38.3 | inducible |
| CI20E | 22.4 | 33 | 36.1 | 35.3 | 43.3 | 43.3 | 28.4 | 19.3 | 33.3 | 23.2 | 37.1 | inducible |
| CI21E | 25.8 | 37.6 | 40.3 | 37.3 | 45.3 | 42.7 | 28.4 | 18.2 | 45.3 | 23.7 | 35.1 | inducible |
| AG29E1 | 0 | 10.8 | 9.9 | 13.5 | 27.3 | 28.6 | 22.3 | 20.4 | 31.4 | 23.6 | 29.4 | AmpC + ESBL |
| Nº | Strain | Source | Species |
|---|---|---|---|
| 1 | LE20E | Lettuce | A. veronii |
| 2 | ES19E | Endives | A. hydrophila |
| 3 | ES20E | Endives | A. veronii |
| 4 | ES42E | Endives | A. veronii |
| 5 | AG26E | Water | A. hydrophila |
| 6 | CI20E | Celery | A. salmonicida |
| 7 | CI21E | Celery | A. salmonicida |
| 8 | AG29E1 | Water | A. hydrophila |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pintor-Cora, A.; Tapia, O.; Elexpuru-Zabaleta, M.; Ruiz de Alegría, C.; Rodríguez-Calleja, J.M.; Santos, J.A.; Ramos-Vivas, J. Cytotoxicity and Antimicrobial Resistance of Aeromonas Strains Isolated from Fresh Produce and Irrigation Water. Antibiotics 2023, 12, 511. https://doi.org/10.3390/antibiotics12030511
Pintor-Cora A, Tapia O, Elexpuru-Zabaleta M, Ruiz de Alegría C, Rodríguez-Calleja JM, Santos JA, Ramos-Vivas J. Cytotoxicity and Antimicrobial Resistance of Aeromonas Strains Isolated from Fresh Produce and Irrigation Water. Antibiotics. 2023; 12(3):511. https://doi.org/10.3390/antibiotics12030511
Chicago/Turabian StylePintor-Cora, Alberto, Olga Tapia, María Elexpuru-Zabaleta, Carlos Ruiz de Alegría, Jose M. Rodríguez-Calleja, Jesús A. Santos, and Jose Ramos-Vivas. 2023. "Cytotoxicity and Antimicrobial Resistance of Aeromonas Strains Isolated from Fresh Produce and Irrigation Water" Antibiotics 12, no. 3: 511. https://doi.org/10.3390/antibiotics12030511
APA StylePintor-Cora, A., Tapia, O., Elexpuru-Zabaleta, M., Ruiz de Alegría, C., Rodríguez-Calleja, J. M., Santos, J. A., & Ramos-Vivas, J. (2023). Cytotoxicity and Antimicrobial Resistance of Aeromonas Strains Isolated from Fresh Produce and Irrigation Water. Antibiotics, 12(3), 511. https://doi.org/10.3390/antibiotics12030511