
Incidence and Genomic Background of Antibiotic Resistance in Food-Borne and Clinical Isolates of Salmonella enterica Serovar Derby from Spain

 ,
,  , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Antimicrobial Susceptibility Testing and Detection of Resistance Genes
2.3. Whole Genome Sequencing and Bioinformatics Analysis
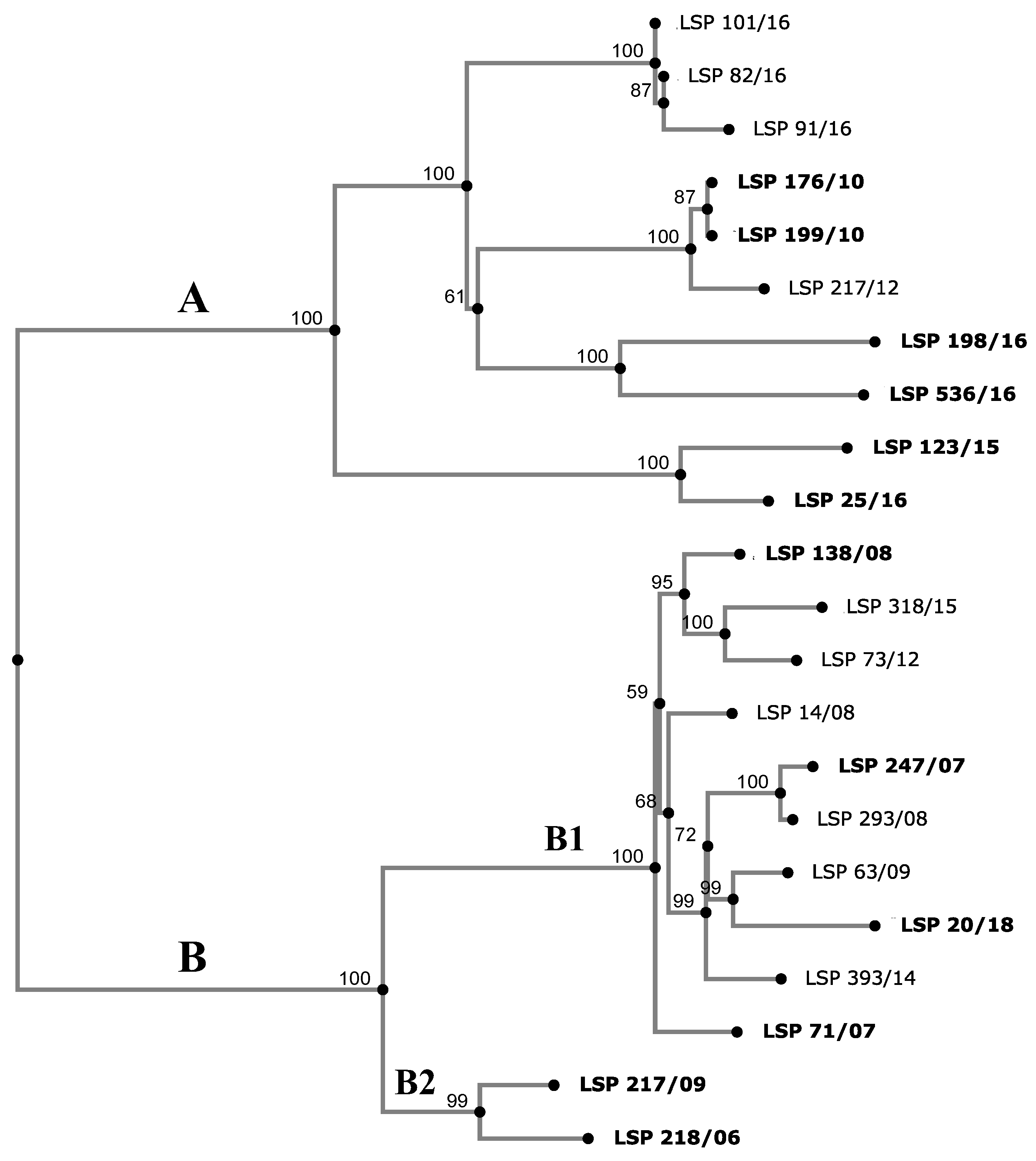
2.4. Phylogenetic Analysis
3. Results
3.1. Origin of the Isolates and General Features of the Sequenced Genomes
3.2. Resistance Phenotypes and Genetic Bases of the Observed Resistances
3.3. Genetic Environment of the Resistance Genes
3.3.1. Chromosomally Located Genes
3.3.2. Plasmid-Located Genes
3.4. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M.; International Collaboration on Enteric Disease ‘Burden of Illness’ Studies. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef]
- WHO. (World Health Organization): Salmonella (Non-Typhoidal)—Fact Sheet. 2018. Available online: http://www.who.int/mediacentre/factsheets/fs139/en/ (accessed on 5 June 2023).
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, e07666. [Google Scholar] [CrossRef]
- WHO. (World Health Organization): Global Action Plan on Antimicrobial Resistance. World Health Organization. 2015. Available online: https://www.who.int/publications/i/item/9789241509763 (accessed on 5 June 2023).
- Grimont, P.A.D.; Weill, F.X. Antigenic formulae of the Salmonella serovars. In World Health Organization Collaborating Center for Reference and Research on Salmonella, 9th ed.; Institut Pasteur: Paris, France, 2007. [Google Scholar]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, 31. [Google Scholar] [CrossRef] [PubMed]
- Threlfall, E.J. Epidemic Salmonella typhimurium DT 104—A truly international multiresistant clone. J. Antimicrob. Chemother. 2000, 46, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Boyd, D.; Peters, G.A.; Cloeckaert, A.; Boumedine, K.S.; Chaslus-Dancla, E.; Imberechts, H.; Mulvey, M.R. Complete nucleotide sequence of a 43-kilobase genomic island associated with the multidrug resistance region of Salmonella enterica serovar Typhimurium DT104 and its identification in phage type DT120 and serovar Agona. J. Bacteriol. 2001, 183, 5725–5732. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.M. Salmonella genomic islands and antibiotic resistance in Salmonella enterica. Future Microbiol. 2010, 5, 1525–1538. [Google Scholar] [CrossRef]
- Mulvey, M.R.; Boyd, D.A.; Olson, A.B.; Doublet, B.; Cloeckaert, A. The genetics of Salmonella genomic island 1. Microbes Infect. 2006, 8, 1915–1922. [Google Scholar] [CrossRef]
- Beutlich, J.; Jahn, S.; Malorny, B.; Hauser, E.; Huhn, S.; Schroeter, A.; Rodicio, M.R.; Appel, B.; Threlfall, J.; Mevius, D.; et al. Antimicrobial resistance and virulence determinants in European Salmonella genomic island 1-positive Salmonella enterica isolates from different origins. Appl. Environ. Microbiol. 2011, 77, 5655–5664. [Google Scholar] [CrossRef]
- Cummins, M.L.; Hamidian, M.; Djordjevic, S.P. Salmonella genomic island 1 is broadly disseminated within Gammaproteobacteriaceae. Microorganisms 2020, 8, 161. [Google Scholar] [CrossRef]
- Hauser, E.; Hebner, F.; Tietze, E.; Helmuth, R.; Junker, E.; Prager, R.; Schroeter, A.; Rabsch, W.; Fruth, A.; Malorny, B. Diversity of Salmonella enterica serovar Derby isolated from pig, pork and humans in Germany. Int. J. Food Microbiol. 2011, 151, 141–149. [Google Scholar] [CrossRef]
- Kerouanton, A.; Rose, V.; Weill, F.X.; Granier, S.A.; Denis, M. Genetic diversity and antimicrobial resistance profiles of Salmonella enterica serotype Derby isolated from pigs, pork, and humans in France. Foodborne Pathog. Dis. 2013, 10, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Michael, G.B.; Cardoso, M.; Rabsch, W.; Schwarz, S. Phenotypic and genotypic differentiation of porcine Salmonella enterica subsp. enterica serovar Derby isolates. Vet. Microbiol. 2006, 118, 312–318. [Google Scholar] [CrossRef]
- Valdezate, S.; Vidal, A.; Herrera-León, S.; Pozo, J.; Rubio, P.; Usera, M.A.; Carvajal, A.; Echeita, M.A. Salmonella Derby clonal spread from pork. Emerg. Infect. Dis. 2005, 11, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Trost, E.; Bender, J.; Fuchs, S.; Malorny, B.; Rabsch, W.; Prager, R.; Tietze, E.; Flieger, A. Evaluation of WGS based approaches for investigating a food-borne outbreak caused by Salmonella enterica serovar Derby in Germany. Food Microbiol. 2018, 71, 46–54. [Google Scholar] [CrossRef] [PubMed]
- González-Santamarina, B.; García-Soto, S.; Hotzel, H.; Meemken, D.; Fries, R.; Tomaso, H. Salmonella Derby: A comparative genomic analysis of strains from Germany. Front. Microbiol. 2021, 12, 591929. [Google Scholar] [CrossRef] [PubMed]
- Hayward, M.R.; Jansen, V.; Woodward, M.J. Comparative genomics of Salmonella enterica serovars Derby and Mbandaka, two prevalent serovars associated with different livestock species in the UK. BMC Genom. 2013, 14, 365. [Google Scholar] [CrossRef]
- Hayward, M.R.; Petrovska, L.; Jansen, V.A.; Woodward, M.J. Population structure and associated phenotypes of Salmonella enterica serovars Derby and Mbandaka overlap with host range. BMC Microbiol. 2016, 16, 15. [Google Scholar] [CrossRef]
- Sevellec, Y.; Felten, A.; Radomski, N.; Granier, S.A.; Le Hello, S.; Petrovska, L.; Mistou, M.Y.; Cadel-Six, S. Genetic diversity of Salmonella Derby from the poultry sector in Europe. Pathogens. 2019, 8, 46. [Google Scholar] [CrossRef]
- Sevellec, Y.; Granier, S.A.; Le Hello, S.; Weill, F.X.; Guillier, L.; Mistou, M.Y.; Cadel-Six, S. Source attribution study of sporadic Salmonella Derby cases in France. Front. Microbiol. 2020, 11, 889. [Google Scholar] [CrossRef]
- Sevellec, Y.; Vignaud, M.L.; Granier, S.A.; Lailler, R.; Feurer, C.; Le Hello, S.; Mistou, M.Y.; Cadel-Six, S. Polyphyletic nature of Salmonella enterica serotype Derby and lineage-specific host-association revealed by genome-wide analysis. Front. Microbiol. 2018, 9, 891. [Google Scholar] [CrossRef]
- Hayward, M.R.; AbuOun, M.; La Ragione, R.M.; Tchorzewska, M.A.; Cooley, W.A.; Everest, D.J.; Petrovska, L.; Jansen, V.A.; Woodward, M.J. SPI-23 of S. Derby: Role in adherence and invasion of porcine tissues. PLoS ONE 2014, 9, e107857. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Tyson, G.H.; Hsu, C.H.; Harrison, L.; Strain, E.; Tran, T.T.; Tillman, G.E.; Dessai, U.; McDermott, P.F.; Zhao, S. Long-read sequencing reveals evolution and acquisition of antimicrobial resistance and virulence genes in Salmonella enterica. Front. Microbiol. 2021, 12, 777817. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; CLSI Supplement M100: Wayne, PA, USA, 2019. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Junker, E.; Miko, A.; Helmuth, R.; Mendoza, M.C. Characterization and localization of drug resistance determinants in multidrug-resistant, integron-carrying Salmonella enterica serotype Typhimurium strains. Microb. Drug Resist. 2004, 10, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.K.; Mulvey, M.R.; Martin, I.; Peters, G.A.; Johnson, W. Genetic characterization of antimicrobial resistance in Canadian isolates of Salmonella serovar Typhimurium DT104. Antimicrob. Agents Chemother. 1999, 43, 3018–3021. [Google Scholar] [CrossRef] [PubMed]
- Perreten, V.; Boerlin, P. A new sulfonamide resistance gene (sul3) in Escherichia coli is widespread in the pig population of Switzerland. Antimicrob. Agents Chemother. 2003, 47, 1169–1172. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Liebert, C.A.; Hall, R.M.; Summers, A.O. Transposon Tn21, flagship of the floating genome. Microbiol. Mol. Biol. Rev. 1999, 63, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.A.; Yin, X.; Persaud-Lachhman, M.G.; Diarra, M.S. First detection of a fosfomycin resistance gene, fosA7, in Salmonella enterica serovar Heidelberg isolated from broiler chickens. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Wang, Z.Y.; Wu, H.; Mei, C.Y.; Shen, P.C.; Pan, Z.M.; Jiao, X. Chromosomally located fosA7 in Salmonella isolates from China. Front. Microbiol. 2021, 12, 781306. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Fang, L.X.; Jiang, Y.W.; Wu, D.S.; Jiang, Q.; Sun, R.Y.; Wang, M.G.; Sun, J.; Liu, Y.H.; Liao, X.P. Comparison of the prevalence and molecular characteristics of fosA3 and fosA7 among Salmonella isolates from food animals in China. J. Antimicrob. Chemother. 2022, 77, 1286–1295. [Google Scholar] [CrossRef] [PubMed]
- Sampei, G.; Furuya, N.; Tachibana, K.; Saitou, Y.; Suzuki, T.; Mizobuchi, K.; Komano, T. Complete genome sequence of the incompatibility group I1 plasmid R64. Plasmid. 2010, 64, 92–103. [Google Scholar] [CrossRef]
- Cohen, S.N.; Chang, A.C. Revised interpretation of the origin of the pSC101 plasmid. J. Bacteriol. 1977, 132, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, A.; Bernardi, F. Complete sequence of pSC101. Nucleic Acids Res. 1984, 12, 9415–9426. [Google Scholar] [CrossRef]
- Nguyen, S.V.; Harhay, D.M.; Bono, J.L.; Smith, T.P.; Fields, P.I.; Dinsmore, B.A.; Santovenia, M.; Kelley, C.M.; Wang, R.; Bosilevac, J.M.; et al. Complete and closed genome sequences of 10 Salmonella enterica subsp. enterica serovar Anatum isolates from human and bovine sources. Genome Announc. 2016, 4. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Ito, R.; Mustapha, M.M.; Tomich, A.D.; Callaghan, J.D.; McElheny, C.L.; Mettus, R.T.; Shanks, R.M.Q.; Sluis-Cremer, N.; Doi, Y. Widespread fosfomycin resistance in Gram-negative bacteria attributable to the chromosomal fosA gene. mBio. 2017, 8. [Google Scholar] [CrossRef]
- Sastry, S.; Doi, Y. Fosfomycin: Resurgence of an old companion. J. Infect. Chemother. 2016, 22, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Rahube, T.O.; Yost, C.K. Characterization of a mobile and multiple resistance plasmid isolated from swine manure and its detection in soil after manure application. J. Appl. Microbiol. 2012, 112, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Salipante, S.J.; Hall, B.G. Determining the limits of the evolutionary potential of an antibiotic resistance gene. Mol. Biol. Evol. 2003, 20, 653–659. [Google Scholar] [CrossRef]
- Magnet, S.; Courvalin, P.; Lambert, T. Activation of the cryptic aac(6′)-Iy aminoglycoside resistance gene of Salmonella by a chromosomal deletion generating a transcriptional fusion. J. Bacteriol. 1999, 181, 6650–6655. [Google Scholar] [CrossRef]
- WHO. Critically Important Antimicrobials for Human Medicine. License: CC BY-NC-SA 30 IGO; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Baucheron, S.; Chaslus-Dancla, E.; Cloeckaert, A.; Chiu, C.H.; Butaye, P. High-level resistance to fluoroquinolones linked to mutations in gyrA, parC, and parE in Salmonella enterica serovar Schwarzengrund isolates from humans in Taiwan. Antimicrob. Agents Chemother. 2005, 49, 862–863. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Park, J.H.; Kwak, H.S.; Woo, G.J. Characterization of the quinolone resistance mechanism in foodborne Salmonella isolates with high nalidixic acid resistance. Int. J. Food Microbiol. 2011, 146, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, X.; Fernández, J.; Hernáez, S.; Rodicio, R.; Rodicio, M.R. Plasmid-mediated quinolone resistance (PMQR) in two clinical strains of Salmonella enterica serovar Corvallis. Microorganisms. 2022, 10, 579. [Google Scholar] [CrossRef]
- Vázquez, X.; García, P.; García, V.; de Toro, M.; Ladero, V.; Heinisch, J.J.; Fernández, J.; Rodicio, R.; Rodicio, M.R. Genomic analysis and phylogenetic position of the complex IncC plasmid found in the Spanish monophasic clone of Salmonella enterica serovar Typhimurium. Sci. Rep. 2021, 11, 11482. [Google Scholar] [CrossRef]
- Weill, F.X.; Bertrand, S.; Guesnier, F.; Baucheron, S.; Cloeckaert, A.; Grimont, P.A. Ciprofloxacin-resistant Salmonella Kentucky in travelers. Emerg. Infect. Dis. 2006, 12, 1611–1612. [Google Scholar] [CrossRef]
- Bonardi, S. Salmonella in the pork production chain and its impact on human health in the European Union. Epidemiol. Infect. 2017, 145, 1513–1526. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Quan, Y.; Yang, S.; Guo, L.; Zhang, X.; Liu, S.; Chen, S.; Zhou, K.; He, L.; Li, B.; et al. Antibiotic resistance in Salmonella from retail foods of animal origin and its association with disinfectant and heavy metal resistance. Microb. Drug Resist. 2018, 24, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Newell, D.G.; Koopmans, M.; Verhoef, L.; Duizer, E.; Aidara-Kane, A.; Sprong, H.; Opsteegh, M.; Langelaar, M.; Threfall, J.; Scheutz, F.; et al. Food-borne diseases—The challenges of 20 years ago still persist while new ones continue to emerge. Int. J. Food Microbiol. 2010, 139 (Suppl. 1), S3–S15. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Shepard, S.M.; Rivet, B.; Danzeisen, J.L.; Carattoli, A. Comparative genomics and phylogeny of the IncI1 plasmids: A common plasmid type among porcine enterotoxigenic Escherichia coli. Plasmid 2011, 66, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef]
- Kazimierczak, K.A.; Scott, K.P.; Kelly, D.; Aminov, R.I. Tetracycline resistome of the organic pig gut. Appl. Environ. Microbiol. 2009, 75, 1717–1722. [Google Scholar] [CrossRef] [PubMed]
- Sandvang, D.; Aarestrup, F.M.; Jensen, L.B. Characterisation of integrons and antibiotic resistance genes in Danish multiresistant Salmonella enterica Typhimurium DT104. FEMS Microbiol. Lett. 1997, 157, 177–181. [Google Scholar] [CrossRef]
- Walker, R.A.; Lindsay, E.; Woodward, M.J.; Ward, L.R.; Threlfall, E.J. Variation in clonality and antibiotic-resistance genes among multirresistant Salmonella enterica serotype Typhimurium phage-type U302 (MR U302) from humans, animals and foods. Microb. Drug Resist. 2001, 7, 13–21. [Google Scholar] [CrossRef]
- Madsen, L.; Aarestrup, F.M.; Olsen, J.E. Characterisation of streptomycin resistance determinants in Danish isolates of Salmonella Typhimurium. Vet. Microbiol. 2000, 75, 73–82. [Google Scholar] [CrossRef]
- Arlet, G.; Phillippon, A. Construction by polymerase chain reaction and intragenic DNA probes for three main types of transferable β-lactamases (TEM, SHV, CARB). FEMS Microbiol. Lett. 1991, 82, 19–26. [Google Scholar] [CrossRef]
- Chu, C.; Chiu, C.H.; Wu, W.Y.; Chu, C.H.; Liu, T.P.; Ou, J.T. Large drug resistance virulence plasmids of clinical isolates of Salmonella enterica serovar Choleraesuis. Antimicrob. Agents Chemother. 2001, 45, 2299–2303. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell Probes. 2001, 15, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Kado, C.I.; Liu, S.T. Rapid procedure for detection and isolation of large and small plasmids. J. Bacteriol. 1981, 145, 1365–1373. [Google Scholar] [CrossRef] [PubMed]
- Threlfall, E.J.; Rowe, B.; Ferguson, J.L.; Ward, L.R. Characterization of plasmids conferring resistance to gentamicin and apramycin in strains of Salmonella typhimurium phage type 204c isolated in Britain. J Hyg. 1986, 97, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, F.; Jiménez, G.; Aguilar, A.; Baquero, F.; Rubio, V. Plasmid pVA517C from Escherichia coli V517 is required for the expression of an antibiotic microcin. J. Antibiot. 1986, 39, 1028–1030. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| R-Profile (N) a | R-Phenotype/R-Genes a,b | Origin (N) a,c | Isolates (LSP) d |
|---|---|---|---|
| R0 (11) | Susceptible/- | PB (8), HF (3) | 48/11, 496/14; 32/15, 33/15, 82/16, 83/16, 91/16, 101/16 102/16, 103/16, 536/16 |
| R1 (2) | TET/tet(B) | HF (2) | 218/06, 217/09 |
| R2 (3) | TET/tet(C) | HF (2), HU (1) | 123/15, 25/16, 198/16 |
| R3 (1) | TET NAL/tet(C) nd | PS (1) | 217/12 |
| R4 (1) | AMP NAL/blaTEM-1B nd | HF (1) | 176/10 |
| R5 (1) | AMP TET NAL/blaTEM-1B tet(A) nd | HF (1) | 199/10 |
| R6 (13) | STR SUL TET/aadA2 sul1 tet(A) | PC (2), PB (2), HF (9) | 71/07, 107/07, 70/08, 138/08, 293/08, 63/09, 20/10, 231/13, 270/13, 110/14, 393/14, 396/14, 20/18 |
| R7 (1) | AMP STR SUL TET/blaTEM-1A aadA2 sul1 tet(A) | PC (1) | 73/12 |
| R8 (1) | STR SUL TET NAL/aadA2 sul1 tet(A) nd | PS (1) | 14/08 |
| R9 (2) | STR SUL TET NAL/aadA2 sul1 tet(A) nd | WB (1), HF (1) | 247/07, 318/15 |
| Isolate a | Origin b | R-Profile: Phenotype/R-Genes c/In d/Tn e | Plasmid(s) f |
|---|---|---|---|
| LSP 82/16 | PB | R0: susceptible | - |
| LSP 91/16 | PB | R0: susceptible | - |
| LSP 101/16 | PB | R0: susceptible | IncI1-I(α)/ST134 |
| LSP 536/16 | HF | R0: susceptible | Col (pHAD28) |
| LSP 218/06 | HF | R1: TET/tet(B)/Tn10 | IncI1-I(α)/uknST |
| LSP 217/09 | HF | R1: TET/tet(B)/Tn10 | IncI1-I(α)/uknST |
| LSP 123/15 | HF | R2: TET/tet(C) | pSC101-like |
| LSP 25/16 | HU | R2: TET/tet(C) | pSC101-like |
| LSP 198/16 | HF | R2: TET/tet(C) | pSC101-like |
| LSP 217/12 | PS | R3: TET NAL/tet(C), gyrA (Asp87 to Asn) | pSC101-like, p0111, ColE10 |
| LSP 176/10 | HF | R4: AMP NAL/blaTEM-1B, gyrA (Asp87 to Asn)/ΔTn2 | IncI1-I(α)/ST25, p0111 |
| LSP 199/10 | HF | R5: AMP TET NAL/blaTEM-1B tet(A) gyrA (Asp87 to Asn) /ΔTn2/ΔTn1721 | IncI1-1(α)/ST25, p0111 |
| LSP 71/07 | HF | R6: STR SUL TET/aadA2 sul1 tet(A)/ + /ΔTn1721 | IncQ1 |
| LSP 138/08 | HF | R6: STR SUL TET/aadA2 sul1 tet(A)/ + /ΔTn1721 | - |
| LSP 293/08 | PC | R6: STR SUL TET/aadA2 sul1 tet(A)/ + /ΔTn1721 | - |
| LSP 63/09 | PC | R6: STR SUL TET/aadA2 sul1 tet(A)/ + /ΔTn1721 | IncY |
| LSP 393/14 | PB | R6: STR SUL TET/aadA2 sul1 tet(A)/ + /ΔTn1721 | ColE |
| LSP 20/18 | HF | R6: STR SUL TET/aadA2 sul1 tet(A)/ + /ΔTn1721 | - |
| LSP 73/12 | PC | R7: AMP STR SUL TET/blaTEM-1A aadA2 sul1 tet(A) / + /ΔTn1721/Tn3 | IncI1-I(α)/nST278 |
| LSP 14/08 | PS | R8: STR SUL TET NAL/aadA2 sul1 tet(A) gyrA (Asp87 to Asn) / + /ΔTn1721 | IncP, ColE |
| LSP 247/07 | HF | R9: STR SUL TET NAL/aadA2 sul1 tet(A) ni/ + /ΔTn1721 | - |
| LSP 318/15 | WB | R9: STR SUL TET NAL/aadA2 sul1 tet(A) ni/ + /ΔTn1721 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez, X.; García-Fierro, R.; Fernández, J.; Bances, M.; Herrero-Fresno, A.; Olsen, J.E.; Rodicio, R.; Ladero, V.; García, V.; Rodicio, M.R. Incidence and Genomic Background of Antibiotic Resistance in Food-Borne and Clinical Isolates of Salmonella enterica Serovar Derby from Spain. Antibiotics 2023, 12, 1204. https://doi.org/10.3390/antibiotics12071204
Vázquez X, García-Fierro R, Fernández J, Bances M, Herrero-Fresno A, Olsen JE, Rodicio R, Ladero V, García V, Rodicio MR. Incidence and Genomic Background of Antibiotic Resistance in Food-Borne and Clinical Isolates of Salmonella enterica Serovar Derby from Spain. Antibiotics. 2023; 12(7):1204. https://doi.org/10.3390/antibiotics12071204
Chicago/Turabian StyleVázquez, Xenia, Raquel García-Fierro, Javier Fernández, Margarita Bances, Ana Herrero-Fresno, John E. Olsen, Rosaura Rodicio, Víctor Ladero, Vanesa García, and M. Rosario Rodicio. 2023. "Incidence and Genomic Background of Antibiotic Resistance in Food-Borne and Clinical Isolates of Salmonella enterica Serovar Derby from Spain" Antibiotics 12, no. 7: 1204. https://doi.org/10.3390/antibiotics12071204
APA StyleVázquez, X., García-Fierro, R., Fernández, J., Bances, M., Herrero-Fresno, A., Olsen, J. E., Rodicio, R., Ladero, V., García, V., & Rodicio, M. R. (2023). Incidence and Genomic Background of Antibiotic Resistance in Food-Borne and Clinical Isolates of Salmonella enterica Serovar Derby from Spain. Antibiotics, 12(7), 1204. https://doi.org/10.3390/antibiotics12071204